Characterization of Rhamnolipids Produced by an Arctic Marine Bacterium from the Pseudomonas fluorescence Group

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Identification of the Isolate M10B774
2.2. Bioactive Extracts
2.3. Identification of Bioactive Compounds
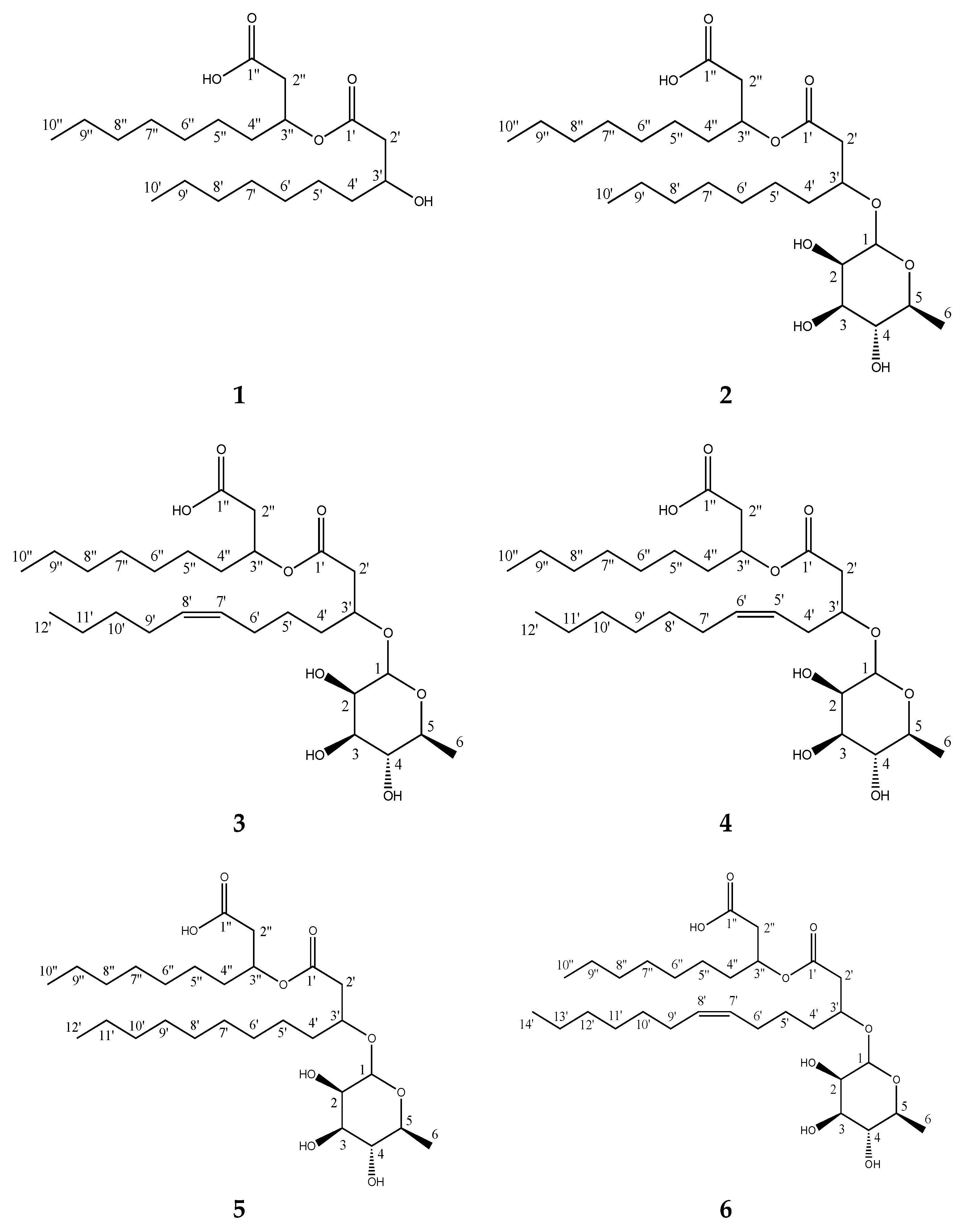
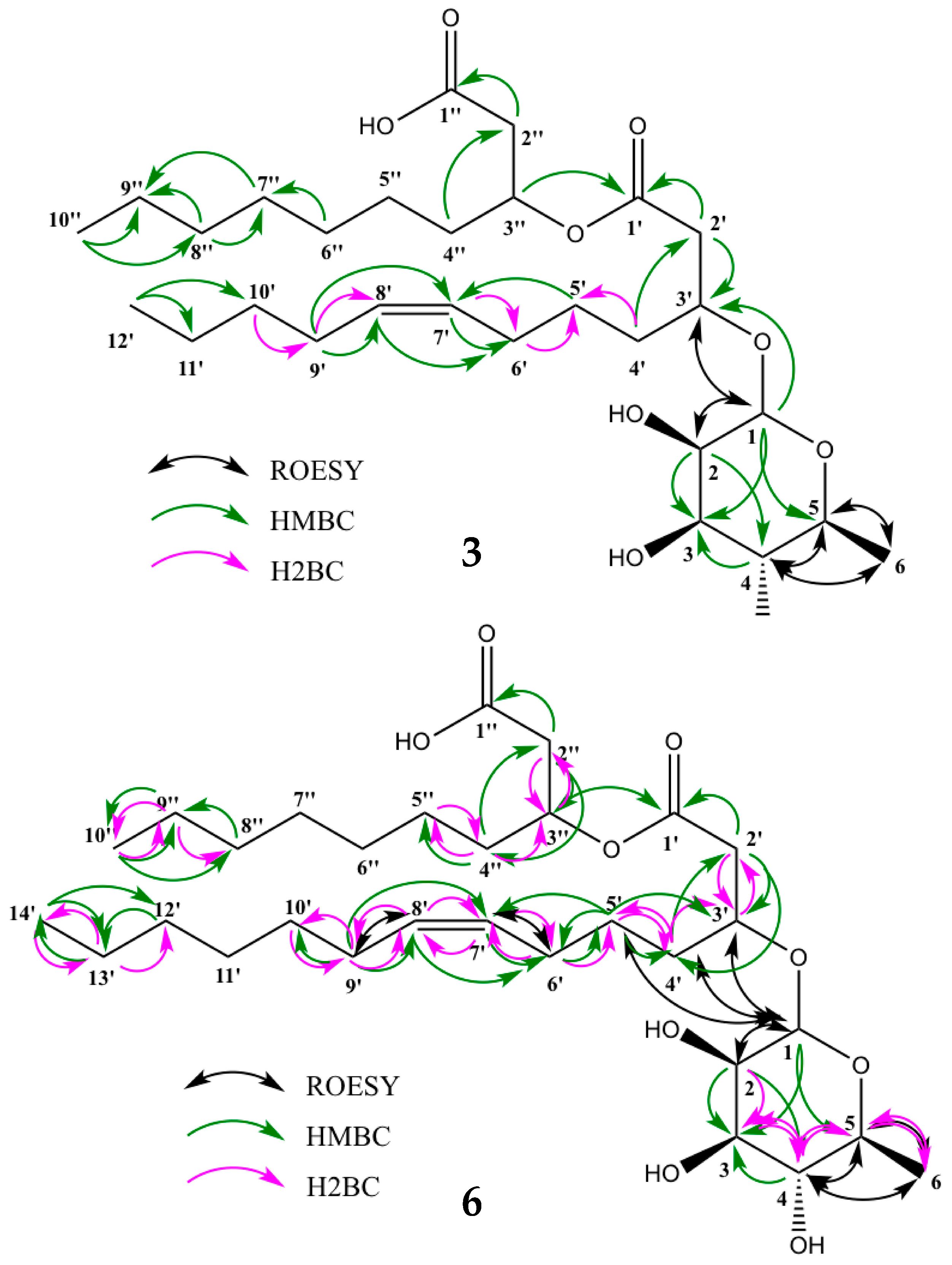
2.4. Characterization of the Isolated Compounds
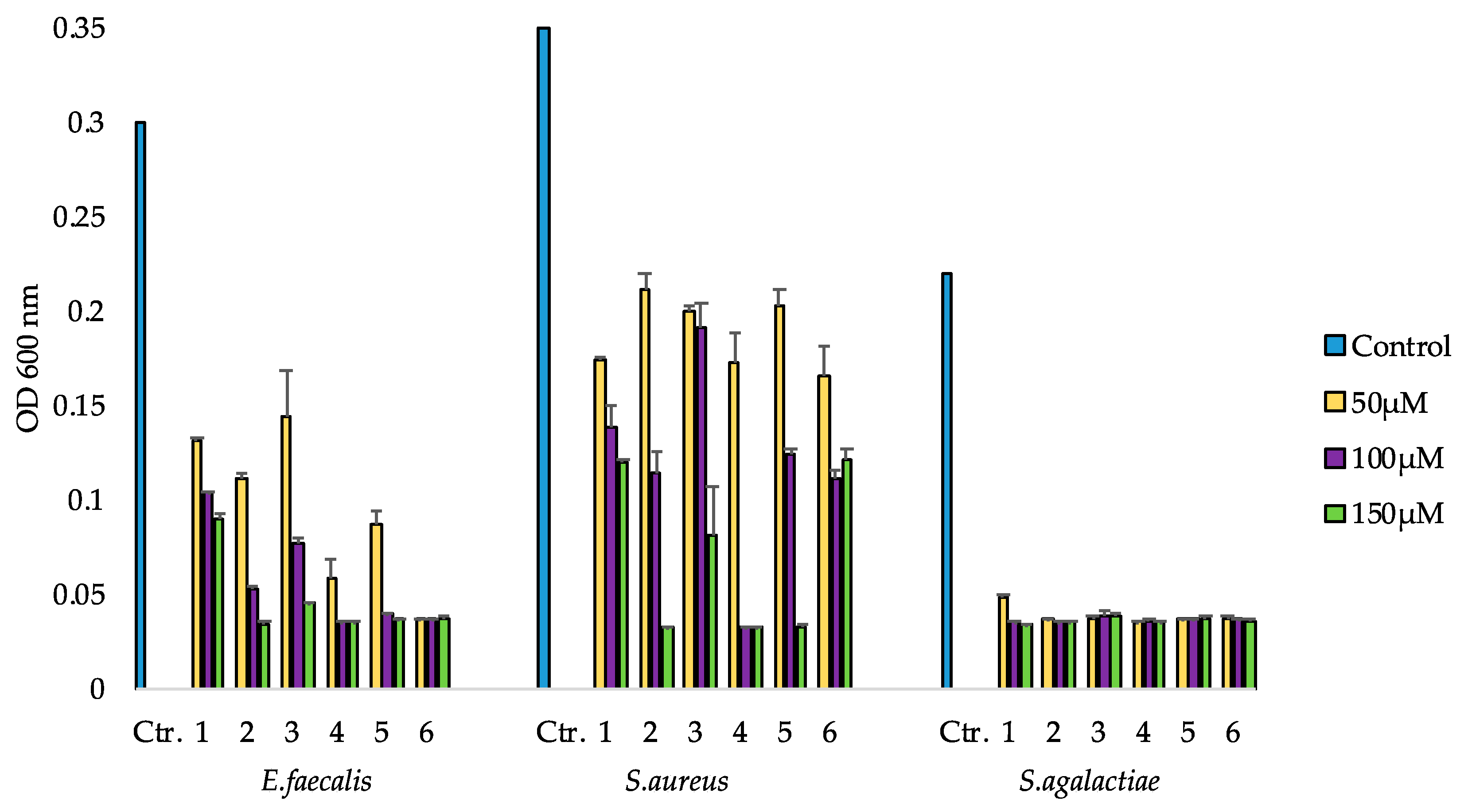
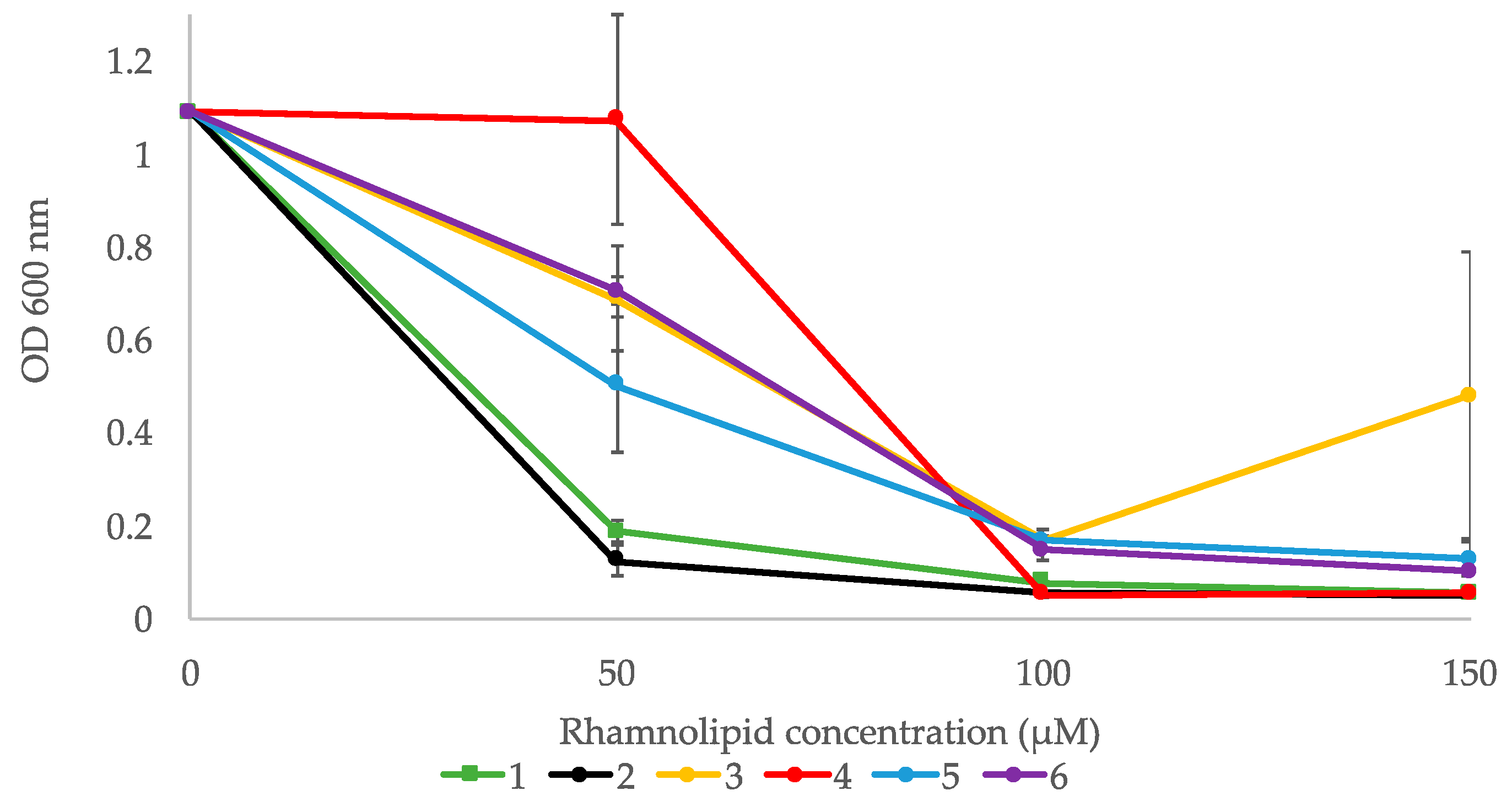
2.5. Bioactivity of Compounds 1–6
2.5.1. Antibacterial Activity
2.5.2. Inhibition of Biofilm Formation
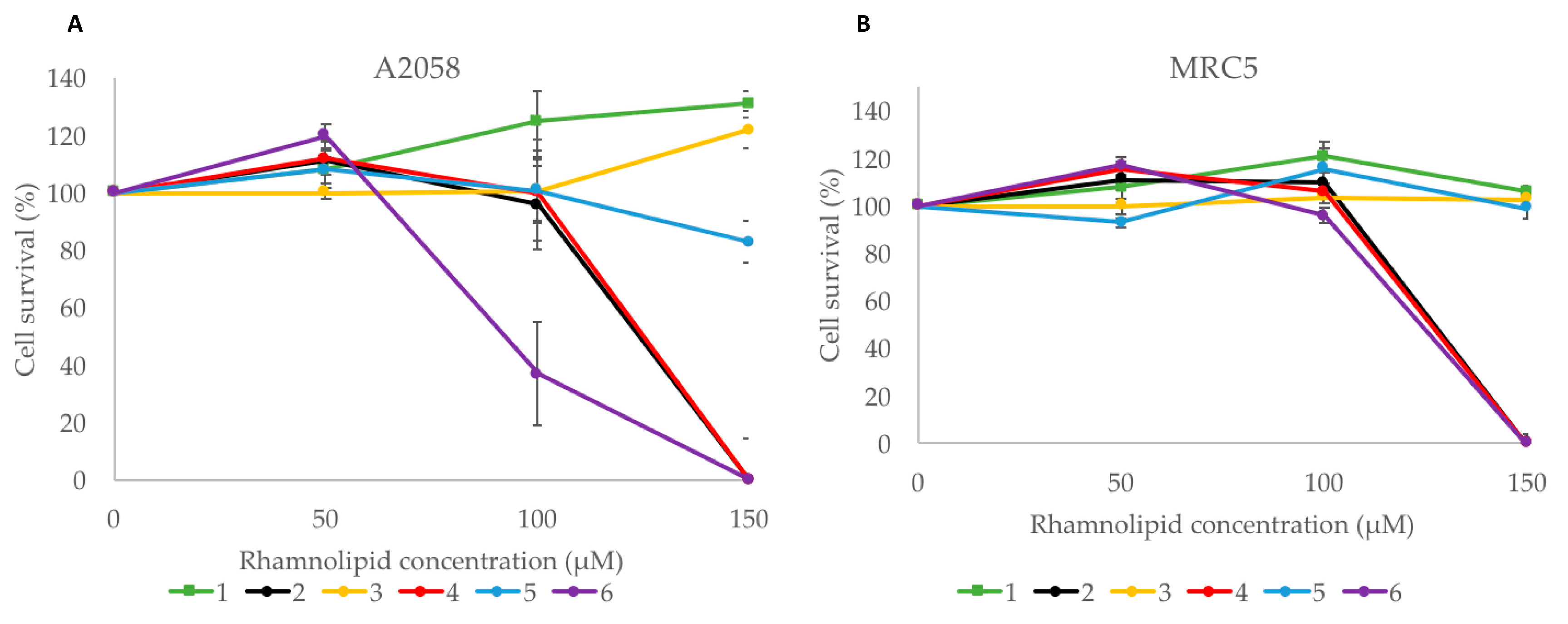
2.5.3. Cytotoxic Activity
3. Discussion
4. Materials and Methods
4.1. Microorganism
4.2. Fermentation and Extraction of Secondary Metabolites
4.3. Fractionation
4.4. Bioactivity
4.4.1. Growth Inhibition Assay
4.4.2. Biofilm Inhibition Assay
4.4.3. Cytotoxicity Assay
4.5. Dereplication, Isolation and Structure Elucidation
4.5.1. LC-MS/MS and Molecular Networking
4.5.2. HR-MS/MS
4.5.3. Isolation of Compounds 1–6
4.5.4. NMR
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rappe, M.S.; Giovannoni, S.J. The uncultured microbial majority. Annu. Rev. Microbiol. 2003, 57, 369–394. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; De Felicio, R.; Fenner, A. Molecular networking as a dereplication strategy. J. Nat. Prod. 2013, 76, 1686–1699. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F.; Månsson, M.; Rank, C.; Frisvad, J.C.; Larsen, T.O. Dereplication of microbial natural products by LC-DAD-TOFMS. J. Nat. Prod. 2011, 74, 2338–2348. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J.; Nuzillard, J.-M.; Renault, J.-H. Dereplication strategies in natural product research: How many tools and methodologies behind the same concept? Phytochem. Rev. 2017, 16, 55–95. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with GNPS. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; van der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers, J.M. Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA 2012, 109, E1743–E1752. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.D.; Wu, C.-H.; Moree, W.J.; Lamsa, A.; Medema, M.H.; Zhao, X.; Gavilan, R.G.; Aparicio, M.; Atencio, L.; Jackson, C. MS/MS networking guided analysis of molecule and gene cluster families. Proc. Natl. Acad. Sci. USA 2013, 110, E2611–E2620. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2014, 42, D32–D37. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.R.; Hayashi, J.A. Structure of a rhamnolipid from Pseudomonas aeruginosa. Arch. Biochem. Biophys. 1965, 111, 415–421. [Google Scholar] [CrossRef]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Rhamnolipids: Diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, P.; Maida, I.; Palma Esposito, F.; Tortorella, E.; Subko, K.; Ezeofor, C.C.; Zhang, Y.; Tabudravu, J.; Jaspars, M.; Fani, R. Antimicrobial activity of monoramnholipids produced by bacterial strains isolated from the Ross Sea (Antarctica). Mar. Drugs 2016, 14, 83. [Google Scholar] [CrossRef] [PubMed]
- Lanni, E.J.; Masyuko, R.N.; Driscoll, C.M.; Aerts, J.T.; Shrout, J.D.; Bohn, P.W.; Sweedler, J.V. MALDI-guided SIMS: Multiscale Imaging of Metabolites in Bacterial Biofilms. Anal. Chem. 2014, 86, 9139–9145. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of marine natural products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Chester, R. Trace elements in the oceans. In Marine Geochemistry; Springer: Dordrecht, The Netherlands, 1990; pp. 346–421. [Google Scholar]
- Bunch, A.W.; Harris, R.E. The manipulation of micro-organisms for the production of secondary metabolites. Biotechnol. Genet. Eng. Rev. 1986, 4, 117–144. [Google Scholar] [CrossRef] [PubMed]
- Abouseoud, M.; Yataghene, A.; Amrane, A.; Maachi, R. Biosurfactant production by free and alginate entrapped cells of Pseudomonas fluorescens. J. Ind. Microbiol. Biotechnol. 2008, 35, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Reiser, J. Autoinducer-mediated regulation of rhamnolipid biosurfactant synthesis in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1995, 92, 6424–6428. [Google Scholar] [CrossRef] [PubMed]
- Itoh, S.; Honda, H.; Tomita, F.; SUZUKI, T. Rhamnolipids produced by Pseudomonas aeruginosa grown on n-paraffin (mixture of C12, C13 and C14 fractions). J. Antibiot. 1971, 24, 855–859. [Google Scholar] [CrossRef]
- Maqsood, M.I.; Jamal, A. Factors affecting the rhamnolipid biosurfactant production. Pak. J. Biotechnol. 2011, 8, 1–5. [Google Scholar]
- Déziel, E.; Lépine, F.; Dennie, D.; Boismenu, D.; Mamer, O.A.; Villemur, R. Liquid chromatography/mass spectrometry analysis of mixtures of rhamnolipids produced by Pseudomonas aeruginosa strain 57RP grown on mannitol or naphthalene. Biochim. Biophys. Acta 1999, 1440, 244–252. [Google Scholar] [CrossRef]
- Behrens, B.; Engelen, J.; Tiso, T.; Blank, L.M.; Hayen, H. Characterization of rhamnolipids by liquid chromatography/mass spectrometry after solid-phase extraction. Anal. Bioanal. Chem. 2016, 408, 2505–2514. [Google Scholar] [CrossRef] [PubMed]
- Bergström, S.; Theorell, H.; Davide, H. On a metabolic product of Ps. pyocyania. Pyolipic acid active against M. tuberculosis. Arkiv Kemi Mineral. Geol. 1946, 23A, 1–12. [Google Scholar]
- Funston, S.J.; Tsaousi, K.; Rudden, M.; Smyth, T.J.; Stevenson, P.S.; Marchant, R.; Banat, I.M. Characterising rhamnolipid production in Burkholderia thailandensis E264, a non-pathogenic producer. Appl. Microbiol. Biotechnol. 2016, 100, 7945–7956. [Google Scholar] [CrossRef] [PubMed]
- Toribio, J.; Escalante, A.E.; Soberón-Chávez, G. Rhamnolipids: Production in bacteria other than Pseudomonas aeruginosa. Eur. J. Lipid Sci. Technol. 2010, 112, 1082–1087. [Google Scholar] [CrossRef]
- Marchant, R.; Banat, I.M. Microbial biosurfactants: Challenges and opportunities for future exploitation. Trends Biotechnol. 2012, 30, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, M.; Costa, S.G.V.A.O. Biosurfactants in food industry. Trends Food Sci. Technol. 2007, 18, 252–259. [Google Scholar] [CrossRef]
- Varvaresou, A.; Iakovou, K. Biosurfactants in cosmetics and biopharmaceuticals. Lett. Appl. Microbiol. 2015, 61, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Vasileva-Tonkova, E.; Sotirova, A.; Galabova, D. The effect of rhamnolipid biosurfactant produced by Pseudomonas fluorescens on model bacterial strains and isolates from industrial wastewater. Curr. Microbiol. 2011, 62, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Abouseoud, M.; Maachi, R.; Amrane, A.; Boudergua, S.; Nabi, A. Evaluation of different carbon and nitrogen sources in production of biosurfactant by Pseudomonas fluorescens. Desalination 2008, 223, 143–151. [Google Scholar] [CrossRef]
- Wilson, N.G.; Bradley, G. The effect of immobilization on rhamnolipid production by Pseudomonas fluorescens. J. Appl. Bacteriol. 1996, 81, 525–530. [Google Scholar]
- El-Amine Bendaha, M.; Mebrek, S.; Mostefa, N.; Tifrit, A.; Belaouni, H.A.; Bouziane, A. Isolation and Comparison of Rhamnolipids Production in Pseudomonas aeruginosa P.B:2 and Pseudomonas fluorescens P.V:10. Sci. Rep. 2012, 544, 1–7. [Google Scholar]
- Irorere, V.U.; Tripathi, L.; Marchant, R.; McClean, S.; Banat, I.M. Microbial rhamnolipid production: A critical re-evaluation of published data and suggested future publication criteria. Appl. Microbiol. Biotechnol. 2017, 101, 3941–3951. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, G.; Hoti, S.; Rao, H.S.P.; Vijjapu, S. Di-rhamnolipid is a mosquito pupicidal metabolite from Pseudomonas fluorescens (VCRC B426). Acta Trop. 2015, 148, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Pantazaki, A.A.; Papaneophytou, C.P.; Lambropoulou, D.A. Simultaneous polyhydroxyalkanoates and rhamnolipids production by Thermus thermophilus HB8. AMB Express 2011, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Gunther, N.W.; Nunez, A.; Fett, W.; Solaiman, D.K. Production of rhamnolipids by Pseudomonas chlororaphis, a nonpathogenic bacterium. Appl. Environ. Microbiol. 2005, 71, 2288–2293. [Google Scholar] [CrossRef] [PubMed]
- Irfan-Maqsood, M.; Seddiq-Shams, M. Rhamnolipids: Well-characterized glycolipids with potential broad applicability as biosurfactants. Ind. Biotechnol. 2014, 10, 285–291. [Google Scholar] [CrossRef]
- Remichkova, M.; Galabova, D.; Roeva, I.; Karpenko, E.; Shulga, A.; Galabov, A.S. Anti-herpesvirus activities of Pseudomonas sp. S-17 rhamnolipid and its complex with alginate. Z. Naturforsch. C 2008, 63, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Benincasa, M.; Abalos, A.; Oliveira, I.; Manresa, A. Chemical structure, surface properties and biological activities of the biosurfactant produced by Pseudomonas aeruginosa LBI from soapstock. Antonie Van Leeuwenhoek 2004, 85, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Thanomsub, B.; Pumeechockchai, W.; Limtrakul, A.; Arunrattiyakorn, P.; Petchleelaha, W.; Nitoda, T.; Kanzaki, H. Chemical structures and biological activities of rhamnolipids produced by Pseudomonas aeruginosa B189 isolated from milk factory waste. Bioresour. Technol. 2006, 97, 2457–2461. [Google Scholar] [CrossRef] [PubMed]
- Irie, Y.; O’toole, G.A.; Yuk, M.H. Pseudomonas aeruginosa rhamnolipids disperse Bordetella bronchiseptica biofilms. FEMS Microbiol. Lett. 2005, 250, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Borah, S.N.; Lahkar, J.; Handique, P.J.; Deka, S. Antifungal properties of rhamnolipid produced by Pseudomonas aeruginosa DS9 against Colletotrichum falcatum. J. Basic Microbiol. 2015, 55, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Denyer, S.P. Mechanisms of action of antibacterial biocides. Int. Biodeterior. Biodegrad. 1995, 36, 227–245. [Google Scholar] [CrossRef]
- Jung, D.; Rozek, A.; Okon, M.; Hancock, R. Structural transitions as determinants of the action of the calcium-dependent antibiotic daptomycin. Chem. Biol. 2004, 11, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Mensa, B.; Howell, G.; Scott, R.; DeGrado, W. Comparative Mechanistic Studies of Brilacidin, Daptomycin, and the Antimicrobial Peptide LL16. Antimicrob. Agents Chemother. 2014, 58, 5136–5145. [Google Scholar] [CrossRef] [PubMed]
- Schreier, S.; Malheiros, S.V.P.; de Paula, E. Surface active drugs: Self-association and interaction with membranes and surfactants. Physicochemical and biological aspects. Biochim. Biophys. Acta 2000, 1508, 210–234. [Google Scholar] [CrossRef]
- Helenius, A.; Simons, K. Solubilization of membranes by detergents. Biochim. Biophys. Acta 1975, 415, 29–79. [Google Scholar] [CrossRef]
- Sotirova, A.V.; Spasova, D.I.; Galabova, D.N.; Karpenko, E.; Shulga, A. Rhamnolipid–biosurfactant permeabilizing effects on Gram-positive and Gram-negative bacterial strains. Curr. Microbiol. 2008, 56, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Al-Tahhan, R.A.; Sandrin, T.R.; Bodour, A.A.; Maier, R.M. Rhamnolipid-induced removal of lipopolysaccharide from Pseudomonas aeruginosa: Effect on cell surface properties and interaction with hydrophobic substrates. Appl. Environ. Microbiol. 2000, 66, 3262–3268. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Shen, C.; Long, X.; Zhang, G.; Meng, Q. Rhamnolipids elicit the same cytotoxic sensitivity between cancer cell and normal cell by reducing surface tension of culture medium. Appl. Microbiol. Biotechnol. 2014, 98, 10187–10196. [Google Scholar] [CrossRef] [PubMed]
- Ferri, J.K.; Stebe, K.J. Which surfactants reduce surface tension faster? A scaling argument for diffusion-controlled adsorption. Adv. Colloid Interface Sci. 2000, 85, 61–97. [Google Scholar] [CrossRef]
- Desai, J.D.; Banat, I.M. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 1997, 61, 47–64. [Google Scholar] [PubMed]
- Ren, Q.; Ruth, K.; Thöny-Meyer, L.; Zinn, M. Enatiomerically pure hydroxycarboxylic acids: Current approaches and future perspectives. Appl. Microbiol. Biotechnol. 2010, 87, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Ibarguren, M.; López, D.J.; Escribá, P.V. The effect of natural and synthetic fatty acids on membrane structure, microdomain organization, cellular functions and human health. Biochim. Biophys. Acta 2014, 1838, 1518–1528. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Inhibition Assay | Viability Assay | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Media | E. coli N | S.aur P | P.aer N | E.F P | S.aga P | A2058 | MCF7 | HT29 | MRC5 * |
| VR_1 | − | − | − | − | + | − | − | − | − |
| VR_2 | − | − | − | + | + | − | − | − | − |
| M19 | − | + | − | + | + | + | + | + | + |
| SGC | − | − | − | − | − | − | − | − | − |
| Position | δC, Type | δH (J in Hz) | COSY | HMBC | H2BC | ROESY |
|---|---|---|---|---|---|---|
| 1 | 98.7, CH | 4.62, s | 3′,3,5 | 3′,2,4′ | ||
| 2 | 70.3, CH | 3.52 o | 3,4 | |||
| 3 | 69.3, CH | 3.41 o | 4 | 4 | ||
| 4 | 71.9, CH | 3.12, t (9.1) | 3,5 | 10,5 | 5 | 5 |
| 5 | 69.0, CH | 3.43 o | 4,6 | 4 | 6 | 3′,4,6 |
| 6 | 17.8, CH3 | 1.08, d (6.1) | 5 | 4,5 | 5 | 4,5 |
| 1′ | 170.3, C | |||||
| 2′ | 40.1, CH2 | 3′ | 1′,3′ | 3′ | 3′ | |
| 3′ | 72.9, CH | 3.91, d (5.6) | 2′,4′ | 1′ | 2′ | 2′,1,4′ |
| 4′ | 32.1, CH2 | 1.45, dt (10.0, 6.4) | 5′,3′ | 2′,3′ | 5′ | o |
| 5′ | 24.3, CH2 | 1.30 o | 6′,4′ | 7′ | o | |
| 6′ | 26.3, CH2 | 1.98 o | 7′,5′ | 8′,7′ | 7′,5′ | o |
| 7′ | 129.9, CH | 5.33 o | 6′ | 9′,6′ | 8′,6′ | o |
| 8′ | 129.3, CH | 5.32 o | 9′ | 9′,6′ | 9′,7′ | o |
| 9′ | 26.6, CH2 | 2.00 o | 10′,8′ | 11′,10′,8′,7′ | 10′,8′ | o |
| 10′ | 31.3, CH2 | 1.27 o | 9′ | 9′ | o | |
| 11′ | 21.7, CH2 | 1.27 o | 12 | 12′,10′ | 12′ | o |
| 12′ | 13.8, CH3 | 0.86, t (6.9) | 11′ | 11′,10′ | 11′ | o |
| 1″ | 170.6 *, C | |||||
| 2″ | 40.4, CH2 | 2.38 o | 1″ | |||
| 3″ | 71.0, CH | 5.11, s b | 2″,4″ | 1′ | ||
| 4″ | 33.7, CH2 | 1.52, s b | 3″ | 2″ | o | |
| 5″ | 24.7, CH2 | 1.20 o | o | |||
| 6″ | 28.6 **, CH2 | 1.23 o | 7″ | o | ||
| 7″ | 28.8 **, CH2 | 1.23 o | 9″ | o | ||
| 8″ | 31.2, CH2 | 1.22 o | 9″,7″ | o | ||
| 9″ | 22.1, CH2 | 1.25 o | 10″ | 10″,8″ | 10″ | o |
| 10″ | 14.0, CH3 | 0.85, t (7.0) | 9″ | 9″,8″ | 9″ | o |
| Position | δC. type | δH (J in Hz) | COSY | HMBC | H2BC | R/NOESY | |
|---|---|---|---|---|---|---|---|
| 1 | 99.8, CH | 4.78, d (1.5) | 2 | 3′ | 2,3′,4′,5′ | ||
| 2 | 72.8, CH | 3.74, dd (3.3, 1.7) | 1,3 | 1, ** | |||
| 3 | 72.0, CH | 3.67 o | 2,4 | 2,4 | 2,4 | ** | |
| 4 | 74.2, CH | 3.31 o | 3,5 | 5,6 | 3,5 | ** | |
| 5 | 70.1, CH | 3.67 o | 4,6 | 6 | 4,6 | ** | |
| 6 | 17.9, CH3 | 1.25, d (6.1) | 5 | 4 | 5 | 4,5 | |
| 1′ | 172.8, C | 2′ | |||||
| 2′ | 41.2, CH2 | 2.56, dd (15.1, 7.6)2.47 o | 3′ | 4′ w | 3′ | 3′,4′ | |
| 3′ | 74.7, CH | 4.11, dq (7.5, 5.6) | 2′,4′ | 2′,5′ | 2′,4′ | 1,2′,4′,5′ | |
| 4′ | 33.5, CH2 | 1.56 o | 3′,5′ | 2′,5′ | 5′ | 1,2′,3′,5′,6′ | |
| 5′ | 25.9, CH2 | 1.30, qd (7.4, 1.5) | 4′,6′ | 6′ | 4′,6′ | 1,3′,4′,6′ | |
| 6′ | 28.1, CH2 | 2.05 o | 5′,7′ | 5′,(7′,8′) | 7′ | 4′,5′,7′w,7′ | |
| 7′ | 130.3, CH | 5.34 o | 6′ | 5′,(6′,9′) | 6′,8′ | 6′ | |
| 8′ | 131.5, CH | 5.37 o | 9′ | (6′,9′),10′ w | 7′,9′ | 9′ | |
| 9′ | 28.2, CH2 | 2.07 o | 8′,10′ | (7′,8′) | 8′,10′ | 8′,10′,11′ | |
| 10′ | 30.8, CH2 | 1.31 o | 9′ | 9′ | 9′ | 9′ | |
| 11′ | 30.1, CH2 | 1.32 o | 9′ | ||||
| 12′ | 32.99, CH2 * | 1.30 o | 14′ | 13′ | |||
| 13′ | 23.7, CH2 * | 1.31 o | 14′ | 12′,14′ | 14′ | 14′ | |
| 14′ | 14.45, CH3 * | 0.91, t (7.0) * | 13′ | 13′ | 13′ | 13′ | |
| 1″ | 177.1, C | 2″ | |||||
| 2″ | 42.3, CH2 | 2.47 o | 3″ | 4″ w | 3″ | 3″,4″ | |
| 3″ | 73.6, CH | 5.31, m | 2″,4″ | 2″ | 2″,4″ | 2″ | |
| 4″ | 35.4, CH2 | 1.61, q (6.6) | 3″,5″ | 2″ | 5″ | 2″,5″ | |
| 5″ | 26.3, CH2 | 1.33 o | 4″ | 4″ | 4″ | 4″ | |
| 6″ | 30.6, CH2 | 1.31 o | |||||
| 7″ | 30.4, CH2 | 1.31 o | |||||
| 8″ | 32.95, CH2 * | 1.28 o | 10″ | 9″ | |||
| 9″ | 23.7, CH2 * | 1.31 o | 10″ | 8″,10″ | 10″ | 10″ | |
| 10″ | 14.46, CH3 * | 0.90, t (7.0) * | 9″ | 9″ | 9″ | 9″ |
| Medium | d-Mannitol (63560) | Peptone (82303) | d-Glucose (D9434) | Casein Hydrolase (22090) | Malt Extract (70167) | Yeast Extract (Y1625) | FeSO4 ⋅ 7H2O (1.03965) | KBr (22186) |
|---|---|---|---|---|---|---|---|---|
| M19 | 20 | 20 | - | - | - | - | - | - |
| VR_1 | - | 11.11 | - | - | 6.67 | 6.67 | - | - |
| VR_2 | - | 11.11 | - | - | 6.67 | 6.67 | 0.044 | 0.044 |
| SGC | - | - | 4 | 3 | - | - | - | - |
| Compound | Column | Gradient (%) ACN | Time (min) |
|---|---|---|---|
| 1 | XSelect | 55–57 | 7.00 |
| 2 | Atlantis | 70–78 | 10.00 |
| 3 | Atlantis | 68–72 | 10.00 |
| 4 | Atlantis | 70–80 | 12.30 |
| 5 | X-Terra | 70–78 | 10.00 |
| 6 | Atlantis | 80–96 | 12.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kristoffersen, V.; Rämä, T.; Isaksson, J.; Andersen, J.H.; Gerwick, W.H.; Hansen, E. Characterization of Rhamnolipids Produced by an Arctic Marine Bacterium from the Pseudomonas fluorescence Group. Mar. Drugs 2018, 16, 163. https://doi.org/10.3390/md16050163
Kristoffersen V, Rämä T, Isaksson J, Andersen JH, Gerwick WH, Hansen E. Characterization of Rhamnolipids Produced by an Arctic Marine Bacterium from the Pseudomonas fluorescence Group. Marine Drugs. 2018; 16(5):163. https://doi.org/10.3390/md16050163
Chicago/Turabian StyleKristoffersen, Venke, Teppo Rämä, Johan Isaksson, Jeanette Hammer Andersen, William H. Gerwick, and Espen Hansen. 2018. "Characterization of Rhamnolipids Produced by an Arctic Marine Bacterium from the Pseudomonas fluorescence Group" Marine Drugs 16, no. 5: 163. https://doi.org/10.3390/md16050163