HSQC-TOCSY Fingerprinting-Directed Discovery of Antiplasmodial Polyketides from the Marine Ascidian-Derived Streptomyces sp. (USC-16018)

 , , and
, , and
Abstract
:1. Introduction
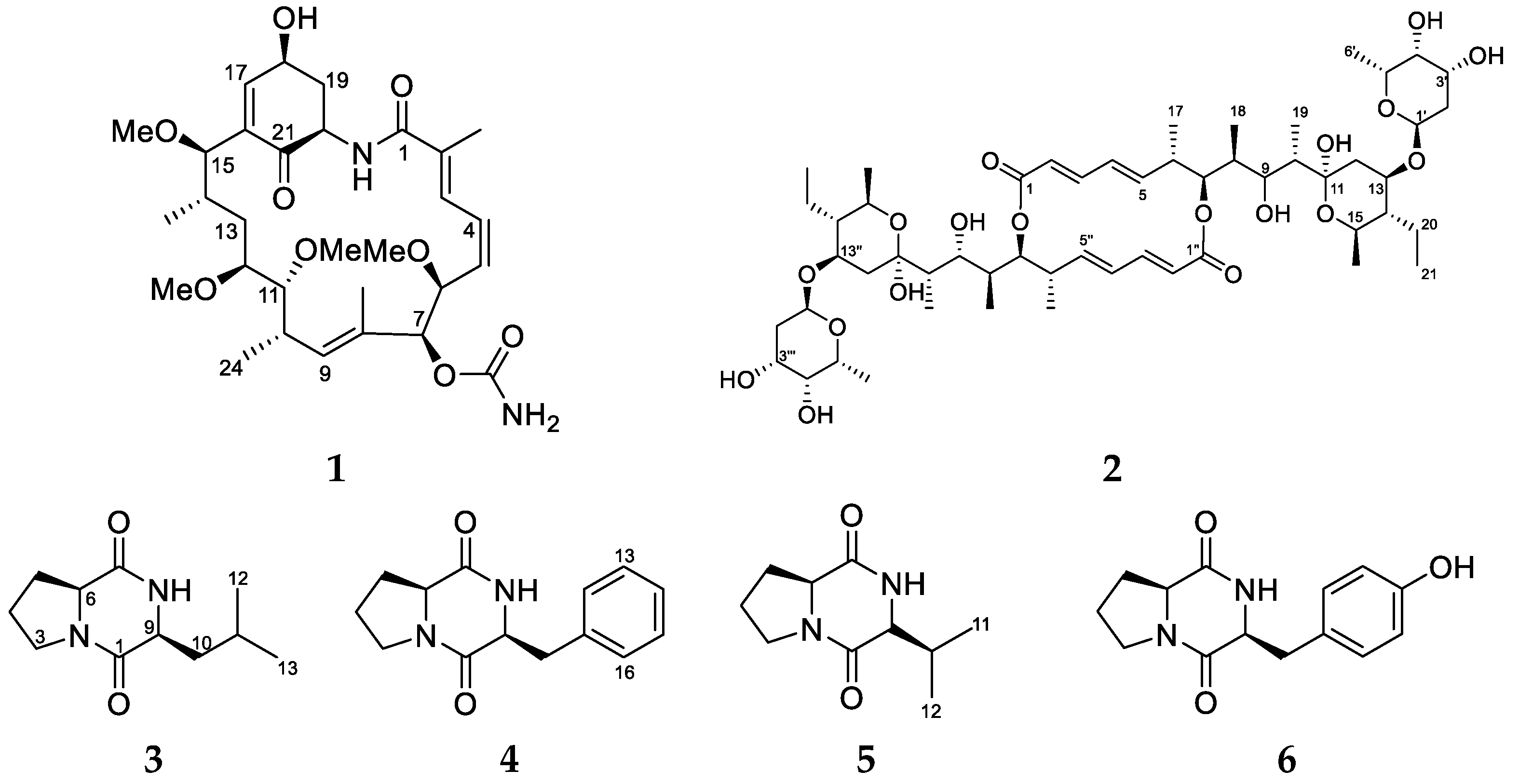
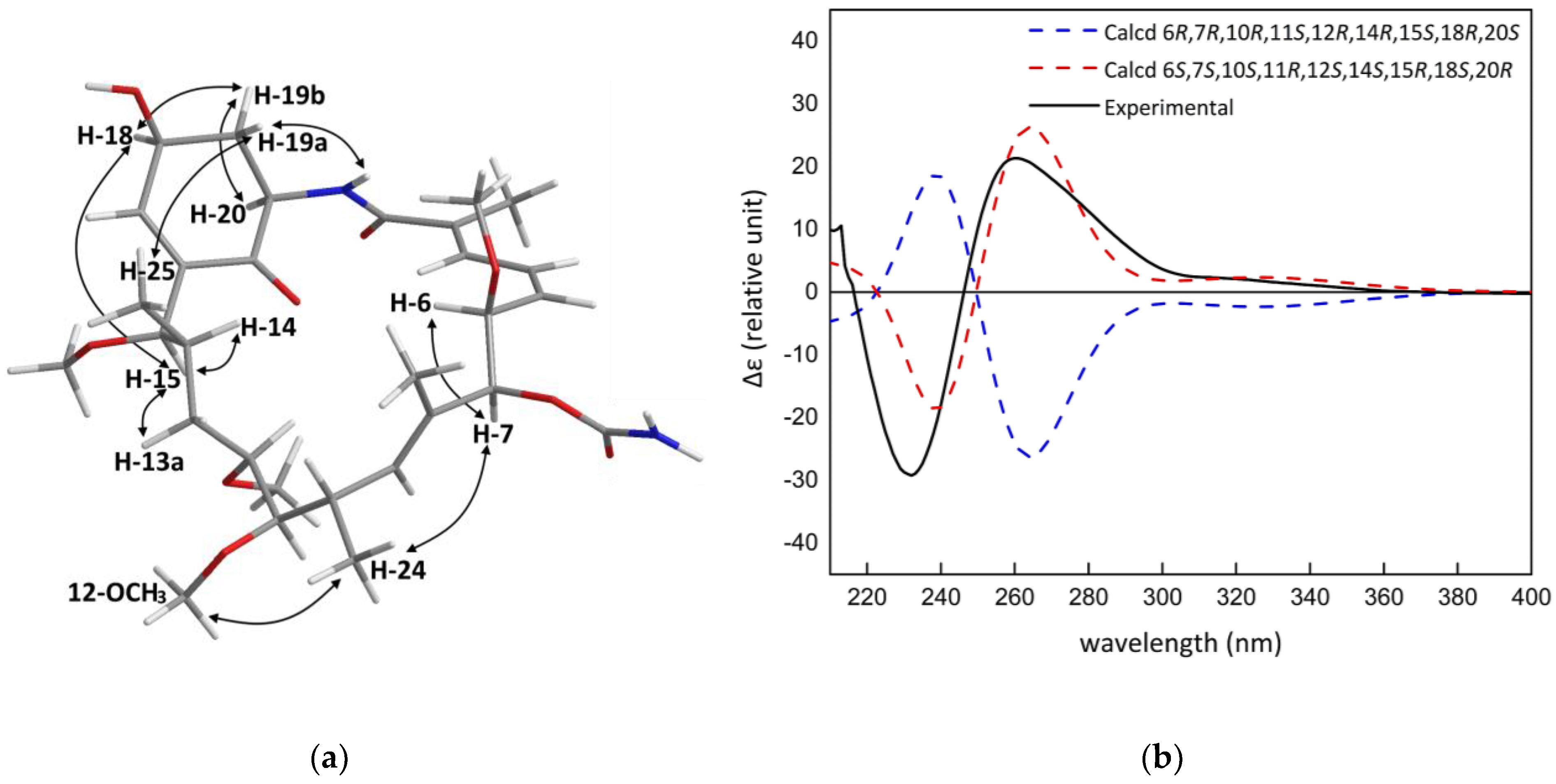
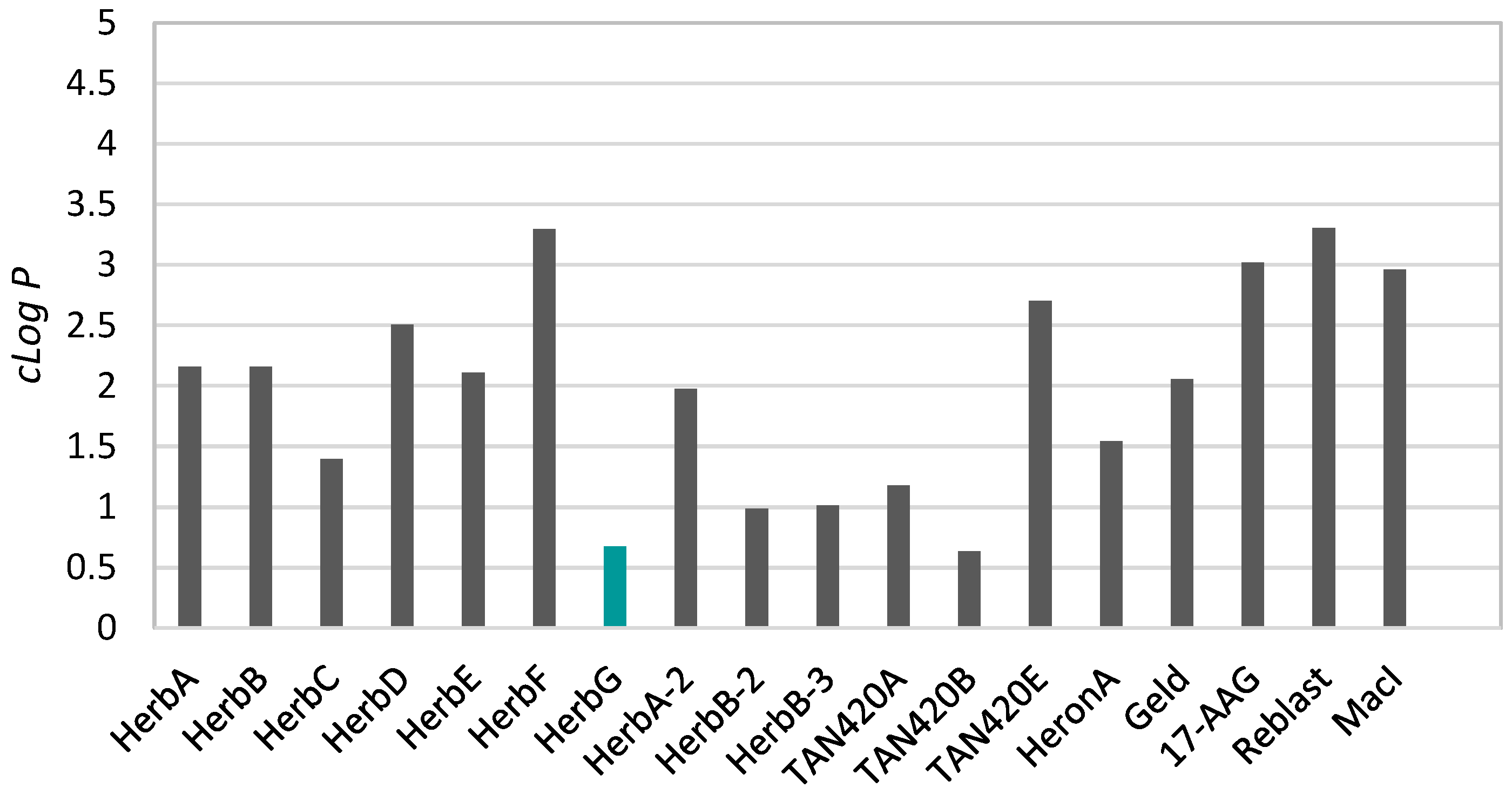
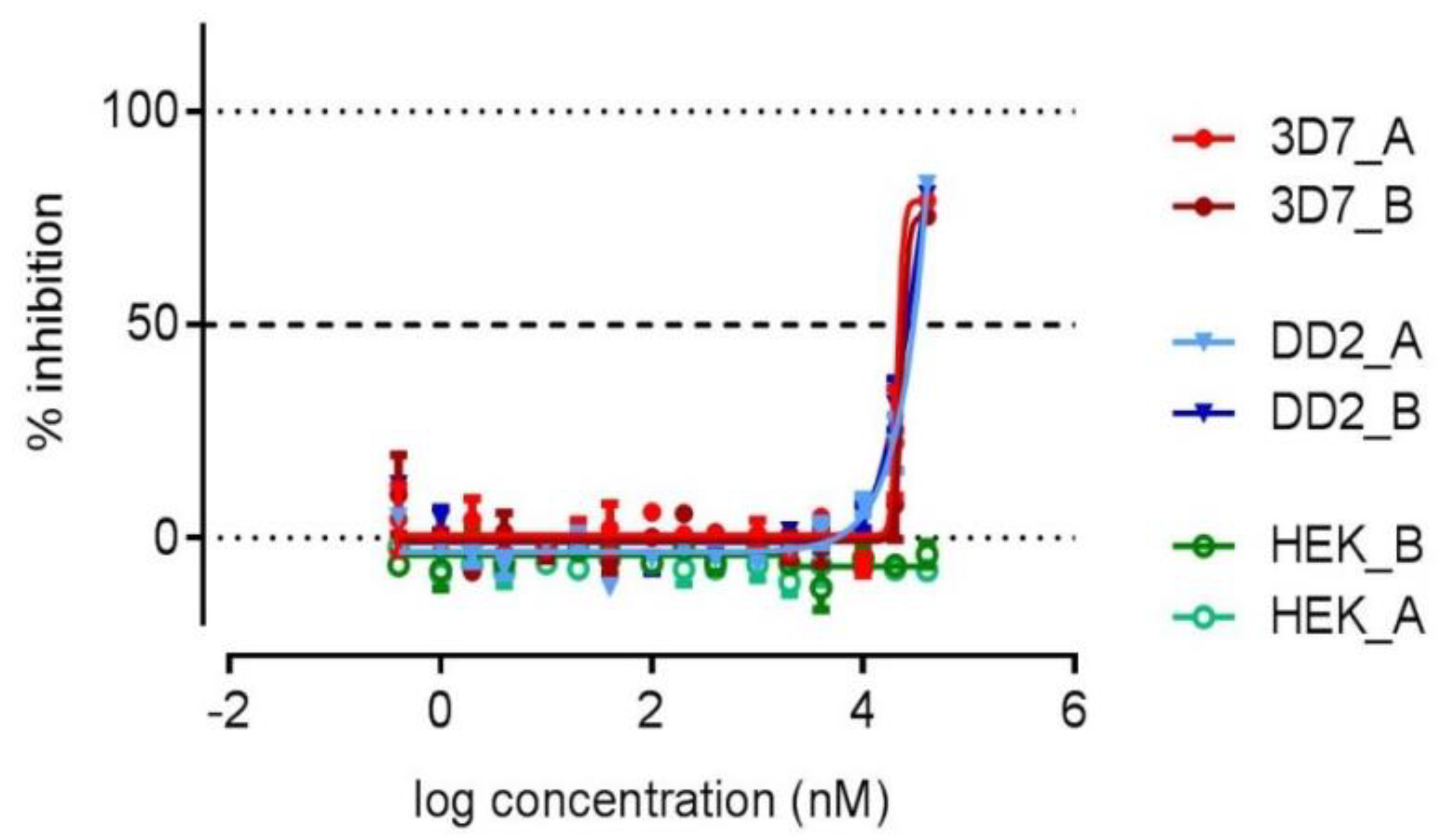
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Section
3.2. Biological Material
3.3. Fermentation, Extraction, and Isolation
3.4. Molecular Modelling Calculations
3.5. Biological Activity Testing
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2017. Available online: http://www.who.int/malaria/publications/world-malaria-report-2017/report/en/ (accessed on 1 May 2018).
- Tuteja, R. Malaria—An overview. FEBS J. 2007, 274, 4670–4679. [Google Scholar] [CrossRef] [PubMed]
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [PubMed]
- Michalakis, Y.; Renaud, F. Malaria: Evolution in vector control. Nature 2009, 462, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Menard, D.; Dondorp, A. Antimalarial drug resistance: A threat to malaria elimination. Cold Spring Harbor Perspect. Med. 2017, 7, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Goodman, C.D.; Useglio, M.; Peirú, S.; Labadie, G.R.; McFadden, G.I.; Rodríguez, E.; Gramajo, H. Chemobiosynthesis of new antimalarial macrolides. Antimicrob. Agents Chemother. 2013, 57, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, A.B.S.; Sun, Q.; Nkrumah, L.J.; Dunne, M.W.; Sacchettini, J.C.; Fidock, D.A. In Vitro Efficacy, Resistance Selection, and Structural Modeling Studies Implicate the Malarial Parasite Apicoplast as the Target of Azithromycin. J. Biol. Chem. 2007, 282, 2494–2504. [Google Scholar] [CrossRef] [PubMed]
- Mout, R.; Xu, Z.-D.; Wolf, A.K.; Davisson, V.J.; Jarori, G.K. Anti-malarial activity of geldanamycin derivatives in mice infected with Plasmodium yoelii. Malar. J. 2012, 11, 54. [Google Scholar] [CrossRef] [PubMed]
- Buedenbender, L.; Carroll, A.R.; Ekins, M.; Kurtböke, D.İ. Taxonomic and metabolite diversity of actinomycetes associated with three Australian ascidians. Diversity 2017, 9, 53. [Google Scholar] [CrossRef]
- Buedenbender, L.; Habener, L.J.; Grkovic, T.; Kurtböke, D.İ.; Duffy, S.; Avery, V.M.; Carroll, A.R. HSQC–TOCSY fingerprinting for prioritization of polyketide- and peptide-producing microbial isolates. J. Nat. Prod. 2018, 81, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Iwai, Y.; Takahashi, Y.; Sadakane, N.; Nakagawa, A.; Oiwa, H.; Hasegawa, Y.; Ikai, T. Herbimycin, a new antibiotic produced by a strain of Streptomyces. J. Antibiot. 1979, 32, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, K.A.; Wang, X.; Elshahawi, S.I.; Ponomareva, L.V.; Sunkara, M.; Copley, G.C.; Hower, J.C.; Morris, A.J.; Kharel, M.K.; Thorson, J.S. Herbimycins D–F, ansamycin analogues from Streptomyces sp. Rm-7-15. J. Nat. Prod. 2013, 76, 1619–1626. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cui, J.; Lu, X.; Zheng, Z.; Liu, X.; Ni, S.; Wang, Y.; Wu, L. Methanethiol as a catabolite of methionine provides methylthio-group for chemical formation of 19-s-methylgeldanamycin and 17,19-dimethylthioherbimycin A. J. Antibiot. 2013, 66, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Iwai, Y.; Nakagawa, A.; Sadakane, N.; Omura, S.; Oiwa, H.; Matsumoto, S.; Takahashi, M.; Kiai, T.; Ochiai, Y. Herbimycin B, a new benzoquinonoid ansamycin with anti-TMV and herbicidal activities. J. Antibiot. 1980, 33, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Jiang, B.; Wu, L.; Nan, Y.; Cui, J.; Yu, L.; Wei, Y.; Li, J.; Shan, G. Two herbimycin analogs, 4,5-dihydro-(4s)-4-hydroxyherbimycin B and (15s)-15-hydroxyherbimycin B, from Streptomyces sp. CPCC 200291. J. Antibiot. 2015, 68, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Raju, R.; Piggott, A.M.; Khalil, Z.; Bernhardt, P.V.; Capon, R.J. Heronamycin A: A new benzothiazine ansamycin from an Australian marine-derived Streptomyces sp. Tetrahedron Lett. 2012, 53, 1063–1065. [Google Scholar] [CrossRef]
- Dimopoulos, M.-A.; Mitsiades, C.S.; Anderson, K.C.; Richardson, P.G. Tanespimycin as antitumor therapy. Clin. Lymphoma Myeloma Leuk. 2011, 11, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Musiyenko, A.; Barik, S. The heat shock protein 90 of Plasmodium falciparum and antimalarial activity of its inhibitor, geldanamycin. Malar. J. 2003, 2, 30. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Levin, J.I. Macrocycles in Drug Discovery; Royal Society of Chemistry: Cambridge, UK, 2015; Volume 40. [Google Scholar]
- Furtado, N.A.; Pupo, M.T.; Carvalho, I.; Campo, V.L.; Duarte, M.C.T.; Bastos, J.K. Diketopiperazines produced by an Aspergillus fumigatus Brazilian strain. J. Braz. Chem. Soc. 2005, 16, 1448–1453. [Google Scholar] [CrossRef]
- Puopolo, G.; Cimmino, A.; Palmieri, M.; Giovannini, O.; Evidente, A.; Pertot, I. Lysobacter capsici az78 produces cyclo (l-pro-l-tyr), a 2,5-diketopiperazine with toxic activity against sporangia of Phytophthora infestans and Plasmopara viticola. J. Appl. Microbiol. 2014, 117, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Hammann, P.; Kretzschmar, G. Secondary metabolites by chemical screening. 12. 13C NMR studies of elaiophylin derivatives. Magn. Reson. Chem. 1991, 29, 667–670. [Google Scholar] [CrossRef]
- Yin, M.; Jiang, M.; Ren, Z.; Dong, Y.; Lu, T. The complete genome sequence of Streptomyces autolyticus CGMCC 0516, the producer of geldanamycin, autolytimycin, reblastatin and elaiophylin. J. Biotechnol. 2017, 252, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Rascher, A.; Hu, Z.; Buchanan, G.O.; Reid, R.; Hutchinson, C.R. Insights into the biosynthesis of the benzoquinone ansamycins geldanamycin and herbimycin, obtained by gene sequencing and disruption. Appl. Environ. Microbiol. 2005, 71, 4862–4871. [Google Scholar] [CrossRef] [PubMed]
- Supong, K.; Sripreechasak, P.; Tanasupawat, S.; Danwisetkanjana, K.; Rachtawee, P.; Pittayakhajonwut, P. Investigation on antimicrobial agents of the terrestrial Streptomyces sp. BCC71188. Appl. Microbiol. Biotechnol. 2017, 101, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, P.H.; Jansma, M.J.; Hoye, T.R. A guide to small-molecule structure assignment through computation of (1H and 13C) NMR chemical shifts. Nat. Protoc. 2014, 9, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16; Gaussian Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Tomasi, J.; Mennucci, B.; Cammi, R. Quantum mechanical continuum solvation models. Chem. Rev. 2005, 105, 2999–3094. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, T.; Schaumloeffel, A.; Hemberger, Y.; Bringmann, G. Specdis: Quantifying the comparison of calculated and experimental electronic circular dichroism spectra. Chirality 2013, 25, 243–249. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Tenderholt, A.L.; Langner, K.M. Cclib: A library for package-independent computational chemistry algorithms. J. Comput. Chem. 2008, 29, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.; Avery, V.M. Development and optimization of a novel 384-well anti-malarial imaging assay validated for high-throughput screening. Am. J. Trop. Med. Hyg. 2012, 86, 84–92. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δC | δH (Multiplicity, J in Hz) | COSY | HMBC | ROESY |
|---|---|---|---|---|---|
| NH | 8.02 (d, 9.4) | 20 | 1, 20 | 3, 19a | |
| 1 | 171.5 | ||||
| 2 | 133.6 | ||||
| 3 | 127.5 | 7.13 (dd,12.2, 1.2) | 4 | 1, 3, 5, 22 | |
| 4 | 126.4 | 6.48 (ddd, 12.2, 10.9, 1.4) | 3, 5 | 2, 6 | |
| 5 | 135.4 | 5.65 (dd, 10.9, 7.9) | 4, 6 | 4 | |
| 6 | 78.1 | 4.56 (dd, 7.9, 1.4) | 5, 7 | 5 | 7 |
| 6-OCH3 | 56.5 | 3.20 (s) | |||
| 7 | 78 | 5.16 (s) | 6 | 8 | 6, 24 |
| 7-OCONH2 | 156.5 | 6.66, 6.30 | |||
| 8 | 133 | ||||
| 9 | 129.4 | 5.20 (d, 7.7) | 10 | 23 | |
| 10 | 33.5 | 2.45 (m) | 9, 11, 24 | 8, 11 | |
| 11 | 83.8 | 3.15 (m) | 10, 12 | 12-OCH3, 13 | |
| 11-OCH3 | 59.8 | 3.35 (s) | 11 | 12, 24 | |
| 12 | 82.3 | 3.33 (m) | 11, 13 | 11-OCH3, 13a | |
| 12-OCH3 | 56.5 | 3.19 (s) | 12 | ||
| 13a | 33.8 | 1.41 (dddd, 14.2, 9.7, 3.4, 3.4) | 12, 14, 15 | ||
| 13b | 1.55 (m) | ||||
| 14 | 36.6 | 1.51 (m) | 13, 15, 25 | 13a, 15 | |
| 15 | 77.6 | 4.15 (d, 1.4) | 14 | 13, 15-OCH3, 16, 17, 25 | 13a, 14 |
| 15-OCH3 | 57.2 | 3.13 (s) | 15 | ||
| 16 | 134.0 | ||||
| 17 | 149.9 | 6.65 (ddd, 3.3, 1.9, 1.4) | 15, 16, 19, 21 | ||
| 18 | 65.1 | 4.68 (dddd, 10.6, 5.1, 4.7, 2.5) | 17, 19 | 19b | |
| 18-OH | 5.56 (d, 5.1) | 17, 18, 19 | 19a | ||
| 19a | 39.2 | 1.96 (dddd, 14.1, 11.8, 10.6, 2.5) | 18 | 18, 20 | NH, 18-OH |
| 19b | 2.26 (dddd, 11.8, 4.9, 4.7 1.9) | 18 | 17, 18, 20, 21 | 18, 20 | |
| 20 | 53.2 | 4.55 (ddd, 14.1, 9.4, 4.9) | NH,19 | 19, 21 | 19b |
| 21 | 197.1 | ||||
| 22 | 12.9 | 1.84 (d, 1.2) | 1, 2, 3 | ||
| 23 | 14.2 | 1.62 (s) | 7, 8, 9 | ||
| 24 | 17.2 | 0.94 (d, 6.7) | 10 | 9, 10, 11 | 11-OCH3 |
| 25 | 13.9 | 0.69 (d, 6.7) | 14 | 13, 14, 15 |
| Compound | % Inhibition at 40 µM (IC50 in nM) | ||
|---|---|---|---|
| P. falciparum 3D7 | P. falciparum Dd2 | HEK-293 Cells | |
| Herbimycin G (1) | 77.2 | 81.7 | no effect |
| Elaiophylin (2) | 96.6 (777.9) | 86.1 (598.5) | 101.9 (1445) |
| Cyclo-l-Pro-l-Leu (3) | 45.9 | 39.0 | no effect |
| Cyclo-l-Pro-l-Phe (4) | no effect | no effect | no effect |
| Cyclo-l-Pro-l-Val (5) | no effect | no effect | no effect |
| Cyclo-l-Pro-l-Tyr (6) | no effect | no effect | no effect |
| Artesunate | 99.6 (0.9) | 98.4 (1.3) | 50.6 |
| Chloroquine | 98.8 (10.0) | 96.5 (87.9) | 40.9 |
| Dihydroartemisinin | 99.9 (0.4) | 98.0 (0.6) | 37.9 |
| Puromycin | 99.0 (148.9) | 99.1 (114.4) | 102.6 (1409.5) |
| Pyrimethamine | 98.7 (4.7) | 23.2 | 68.4 |
| Pyronaridine | 99.9 (7.4) | 97.8 (8.3) | 98.7 (2825.5) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buedenbender, L.; Robertson, L.P.; Lucantoni, L.; Avery, V.M.; Kurtböke, D.İ.; Carroll, A.R. HSQC-TOCSY Fingerprinting-Directed Discovery of Antiplasmodial Polyketides from the Marine Ascidian-Derived Streptomyces sp. (USC-16018). Mar. Drugs 2018, 16, 189. https://doi.org/10.3390/md16060189
Buedenbender L, Robertson LP, Lucantoni L, Avery VM, Kurtböke Dİ, Carroll AR. HSQC-TOCSY Fingerprinting-Directed Discovery of Antiplasmodial Polyketides from the Marine Ascidian-Derived Streptomyces sp. (USC-16018). Marine Drugs. 2018; 16(6):189. https://doi.org/10.3390/md16060189
Chicago/Turabian StyleBuedenbender, Larissa, Luke P. Robertson, Leonardo Lucantoni, Vicky M. Avery, D. İpek Kurtböke, and Anthony R. Carroll. 2018. "HSQC-TOCSY Fingerprinting-Directed Discovery of Antiplasmodial Polyketides from the Marine Ascidian-Derived Streptomyces sp. (USC-16018)" Marine Drugs 16, no. 6: 189. https://doi.org/10.3390/md16060189