Processing Optimization and Characterization of Angiotensin-Ι-Converting Enzyme Inhibitory Peptides from Lizardfish (Synodus macrops) Scale Gelatin

Abstract
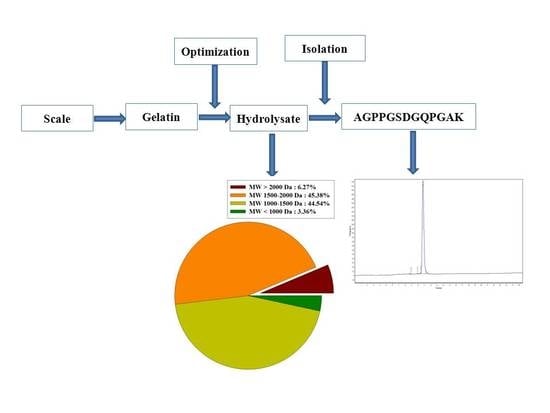
:
1. Introduction
2. Results and Discussion
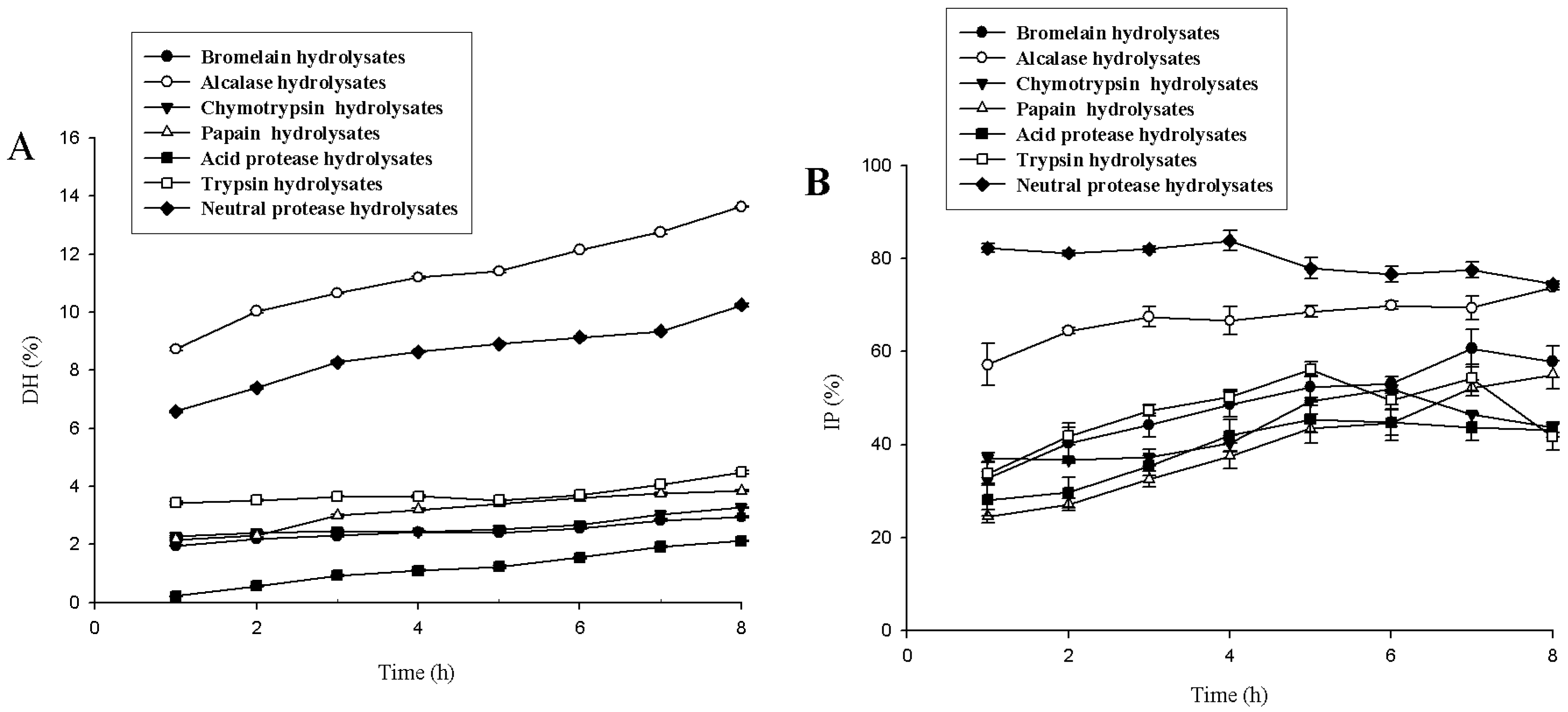
2.1. Screening of Protease
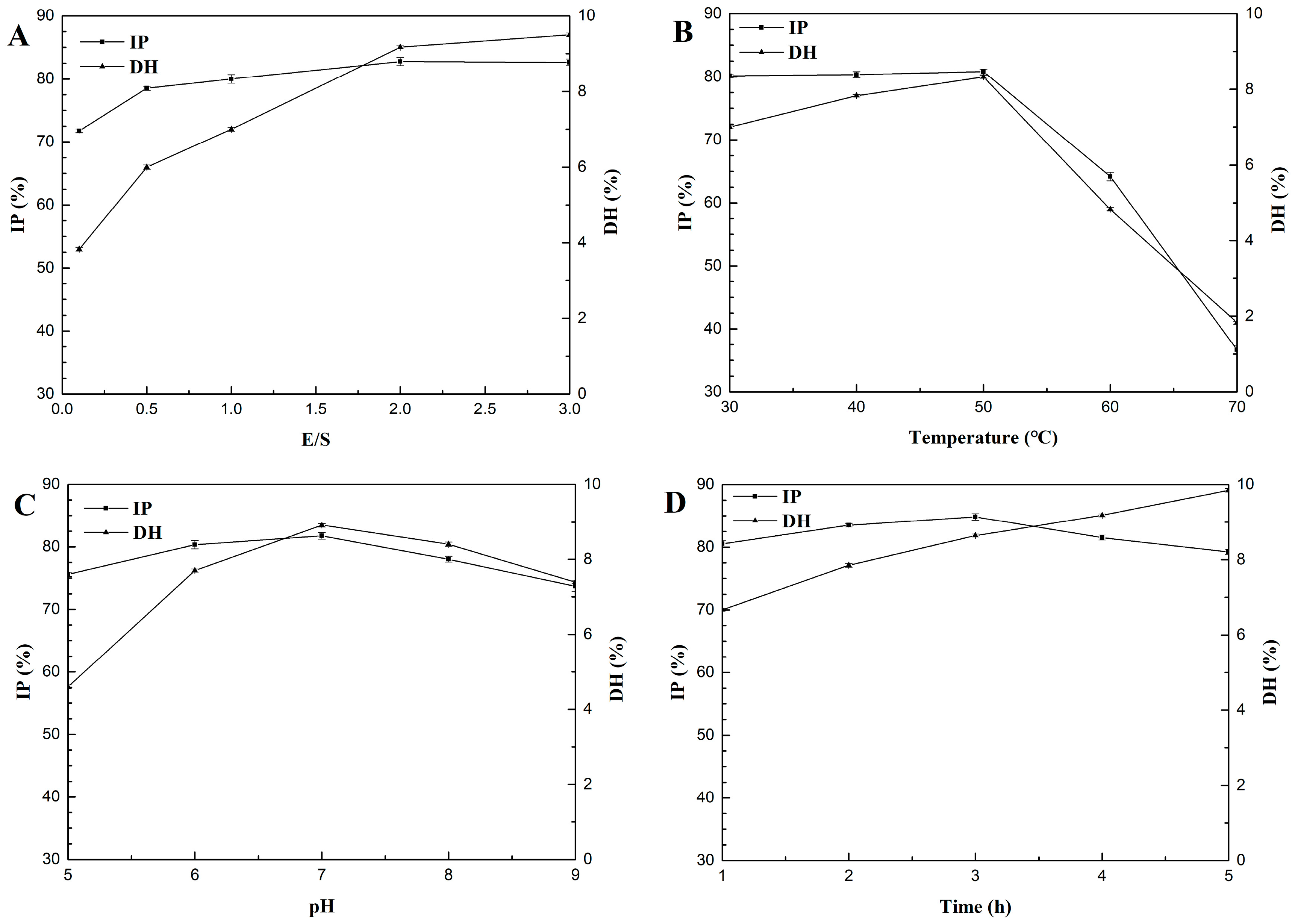
2.2. Effects of E/S Ratio, Hydrolysis Temperature, pH, and Time on DH and ACE Inhibitory Activity
2.3. Central Composite Design and Response Surface Method
X2 − 0.215 X1 X3 + 0.0275 X1 X4 − 0.4975 X22 − 0.2575 X2 X3 − 0.045 X2 X4 − 2.21875 X32
− 0.4375 X3 X4 − 0.64375 X42 R2 = 0.9830
0.175 X1 X3 − 0.3775 X1 X4 − 0.629583 X22 − 1.49 X2 X3 − 0.4175 X2 X4 − 2.424583 X32 −
0.175 X3 X4 + 0.412917 X42 R2 = 0.9326
2.4. Determination of the Optimal Process Variables for the Preparation of Gelatin Peptides
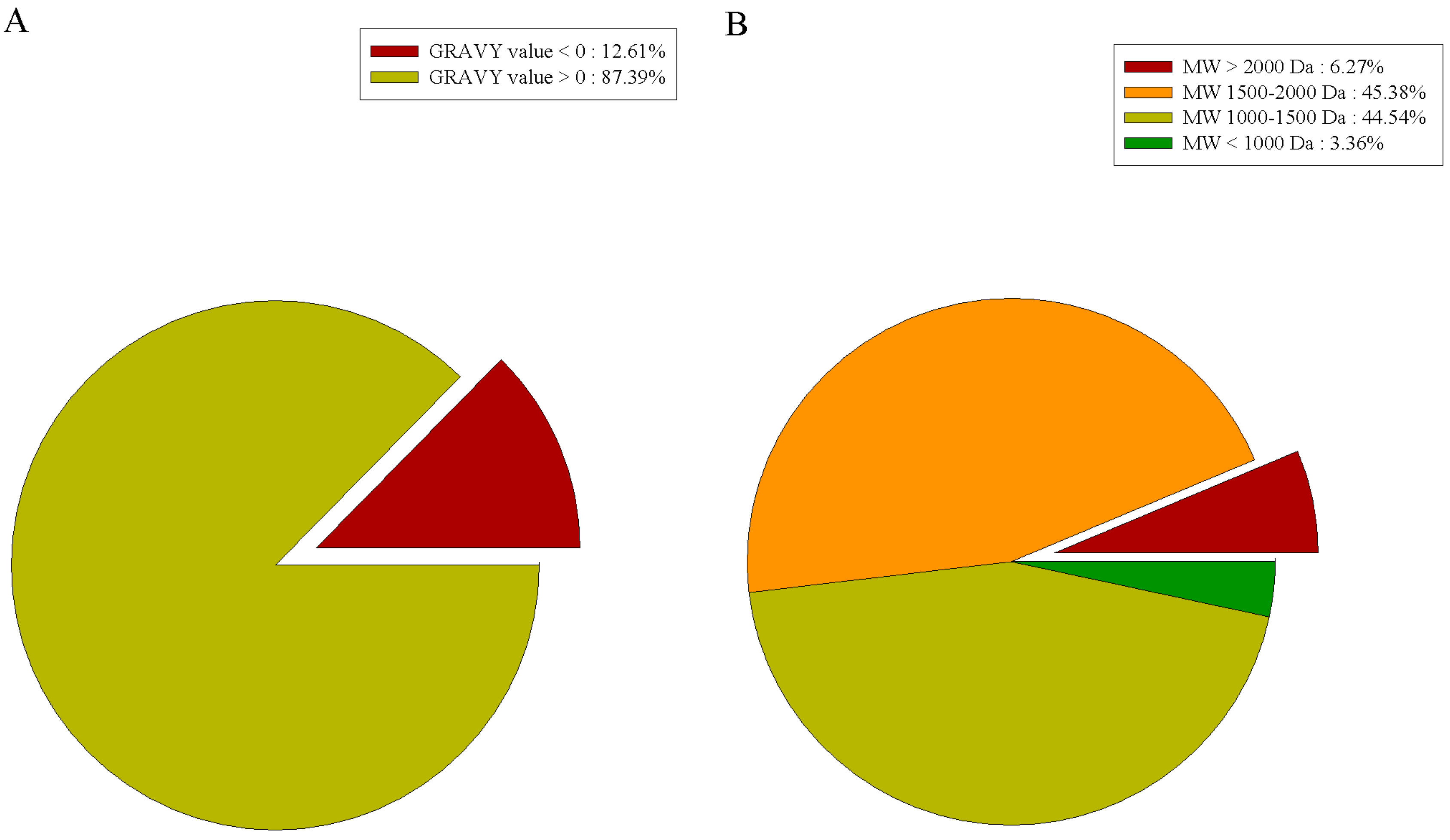
2.5. Characterization of Gelatin Peptides
2.6. Stability of ACE Inhibitory Activity
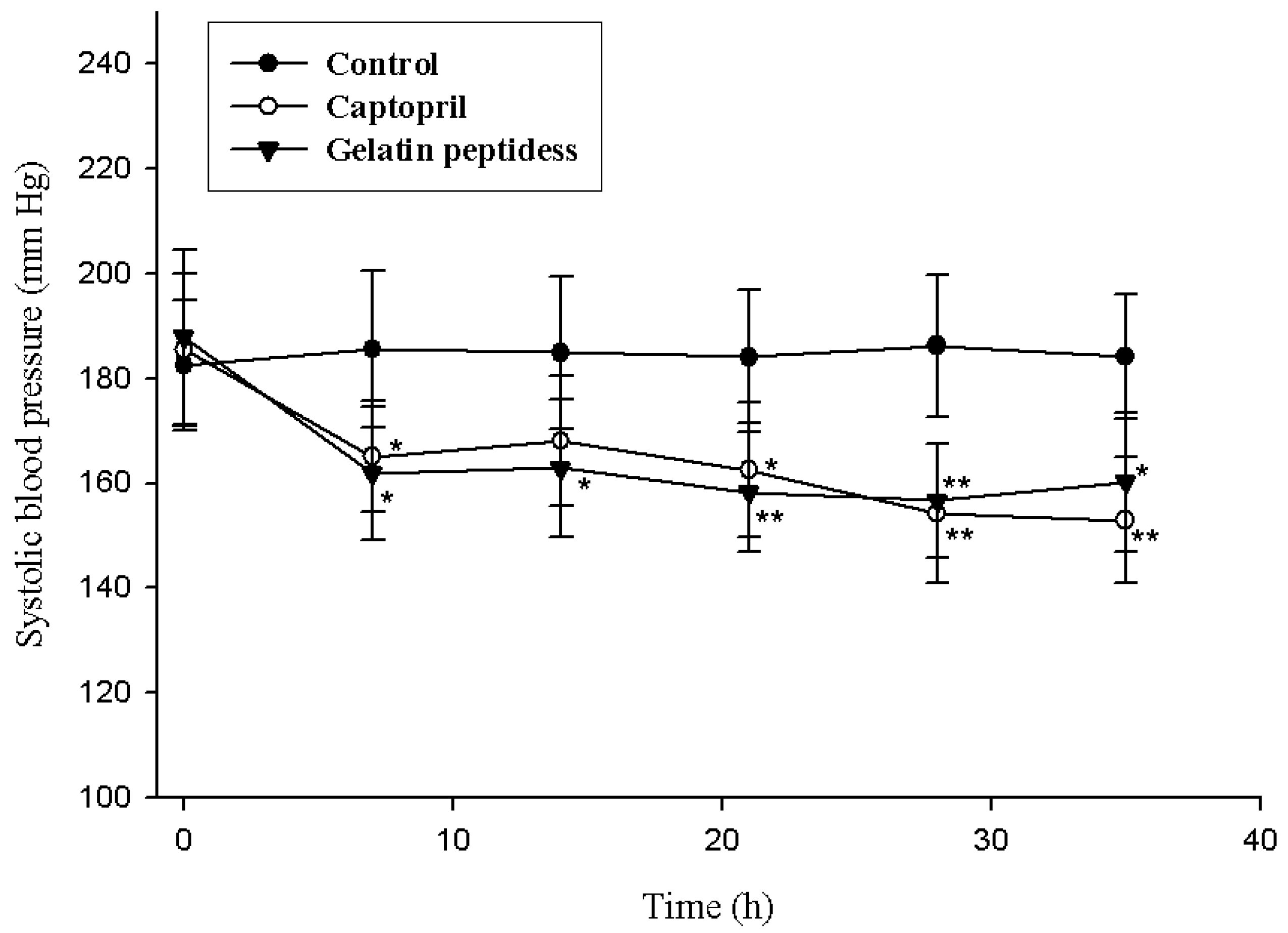
2.7. Antihypertensive Action of Gelatin Peptides in Spontaneously Hypertensive Rats (SHRs)
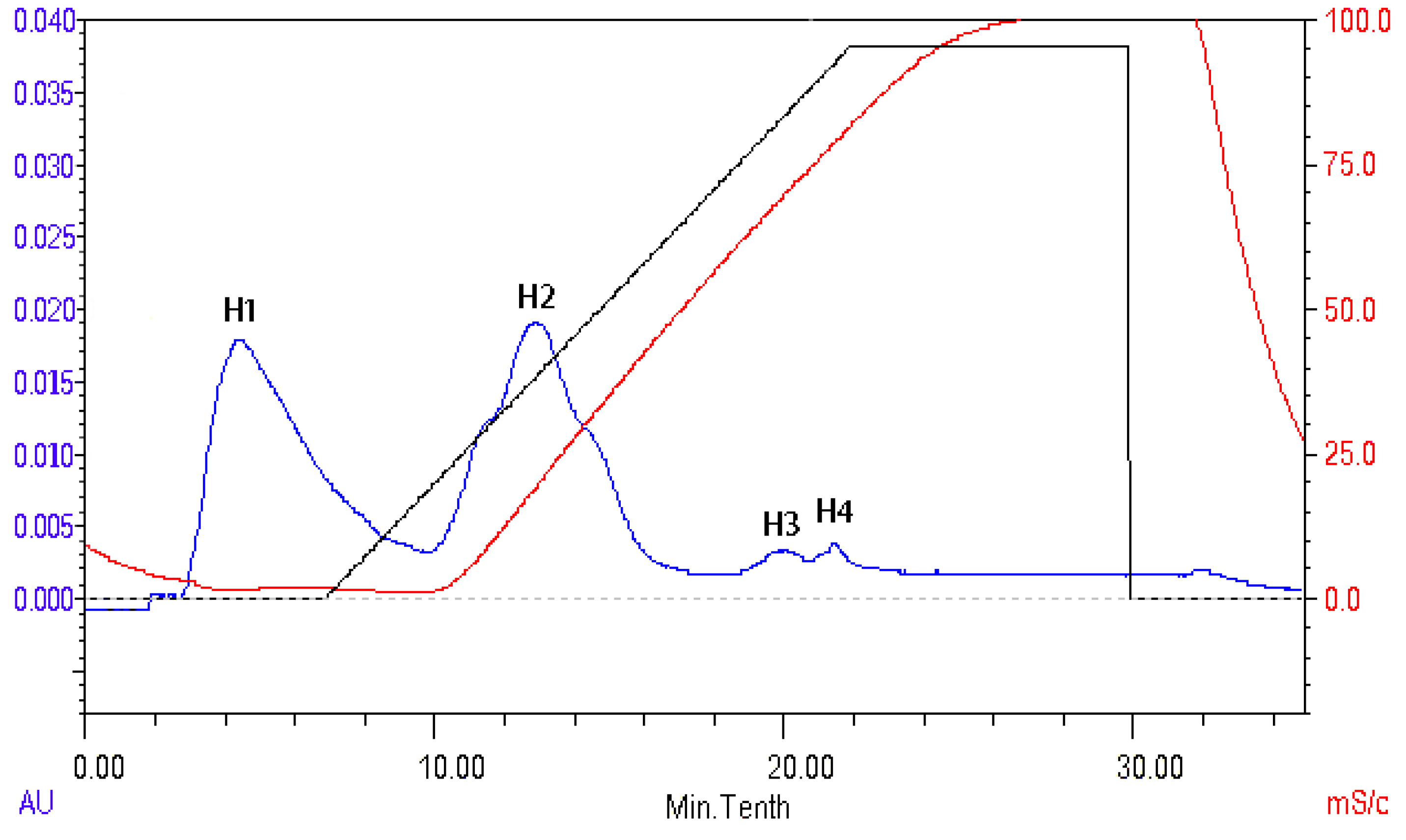
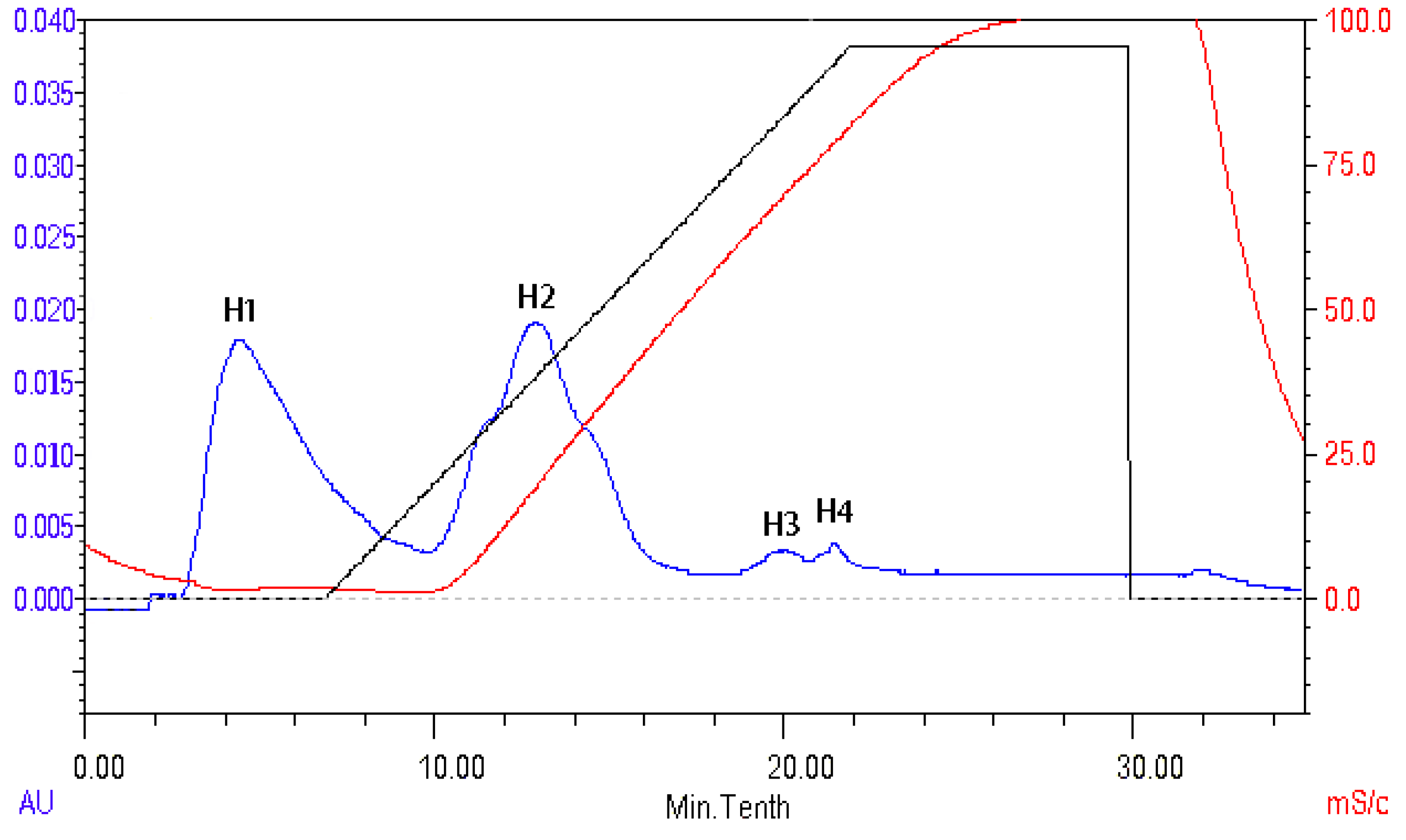
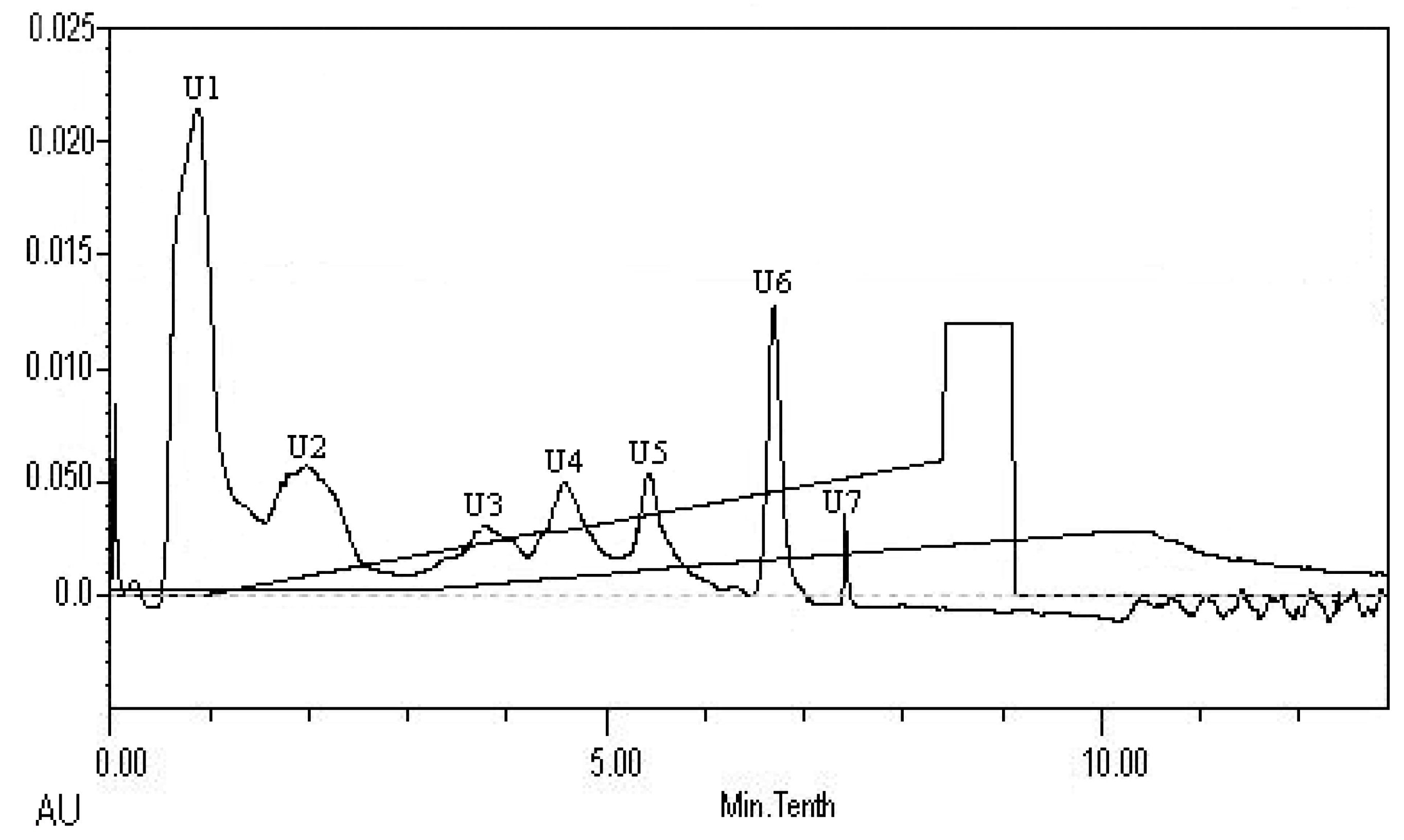
2.8. Isolation and Identification of ACE Inhibitory Peptide
3. Materials and Methods
3.1. Materials
3.2. Preparation of Scale Gelatin and Gelatin Hydrolysates
3.3. Yield of Gelatin Hydrolysates
3.4. DH of Gelatin Hydrolysates
3.5. Measurement of ACE Inhibition
3.6. Optimization Experimental Design
3.7. Peptide Characterization by Nano-LC-MS/MS
3.8. Stability of ACE Inhibitory Activity
3.8.1. Thermal Stability
3.8.2. pH Stability
3.8.3. Gastrointestinal Stability
3.9. Antihypertensive Action of Gelatin Peptides in SHRs
3.10. Isolation and Identification of ACE Inhibitory Peptide
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, S.Y.; Hur, S.J. Antihypertensive peptides from animal products, marine organisms, and plants. Food Chem. 2017, 228, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Lassoued, I.; Mora, L.; Barkia, A.; Aristoy, M.-C.; Nasri, M.; Toldrá, F. Bioactive peptides identified in thornback ray skin’s gelatin hydrolysates by proteases from Bacillus subtilis and Bacillus amyloliquefaciens. J. Proteom. 2015, 128, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Roytrakul, S.; Sutheerawattananonda, M. Production and purification of glucosamine and angiotensin—І converting enzyme (ACE) inhibitory peptides from mushroom hydrolysates. J. Funct. Foods 2017, 36, 72–83. [Google Scholar] [CrossRef]
- Luna-Vital, D.A.; Mojica, L.; De Mejía, E.G.; Mendoza, S.; Loarca-Piña, G. Biological potential of protein hydrolysates and peptides from common bean (Phaseolus Vulgaris L.): A review. Food Res. Int. 2015, 76, 39–50. [Google Scholar] [CrossRef]
- García, M.C.; Puchalska, P.; Esteve, C.; Marina, M.L. Vegetable food: A cheap source of proteins and peptides with antihypertensive, antioxidant, and other less occurrence bioactivities. Talanta 2013, 106, 328–349. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Mohan, A. Mechanisms of food protein-derived antihypertensive peptides other than ACE inhibition. J. Funct. Foods. 2014, 8C, 45–52. [Google Scholar] [CrossRef]
- Jemil, I.; Abdelhedi, O.; Nasri, R.; Mora, L.; Jridi, M.; Aristoy, M.-C.; Toldrá, F.; Nasri, M. Novel bioactive peptides from enzymatic hydrolysate of Sardinelle (Sardinella aurita) muscle proteins hydrolysed by Bacillus subtilis A26 proteases. Food Res. Int. 2017, 100, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.S.; Jia, A.; Zhang, Y.G.; Zhu, H.M.; Zhang, C.; Sun, Z.L.; Liu, C.H. Purification and characterization of angiotensin І converting enzyme inhibitory peptides from jellyfish Rhopilema esculentun. Food Res. Int. 2013, 50, 339–343. [Google Scholar] [CrossRef]
- Fujita, H.; Yamagami, T.; Ohshima, K. Effects of an ACE-inhibitory agent, katsuobushi oligopeptide, in the spontaneously hypertensive rat and in borderline and mildly hypertensive subjects. Nutr. Res. 2001, 21, 1149–1158. [Google Scholar] [CrossRef]
- Chen, J.D.; Li, L.; Yi, R.Z.; Xu, N.H.; Gao, R.; Hong, B.H. Extraction and characterization of acid-soluble collagen from scales and skin of tilapia (Oreochromis niloticus). LWT-Food Sci. Technol. 2016, 66, 453–459. [Google Scholar] [CrossRef]
- Venkatesan, J.; Anil, S.; Kim, S.K.; Shim, M.S. Marine fish proteins and peptides for cosmeceuticals: A review. Mar. Drugs 2017, 15, 143. [Google Scholar] [CrossRef] [PubMed]
- Veeruraj, A.; Arumugam, M.; Ajithkumar, T.; Balasubramanian, T. Isolation and characterization of collagen from the outer skin of squid (Doryteuthis Singhalensis). Food Hydrocolloid. 2015, 43, 708–716. [Google Scholar] [CrossRef]
- Matmaroh, K.; Benjakul, S.; Prodpran, T.; Encarnaction, A.B.; Kishimura, H. Characteristics of acid soluble collagen and pepsin soluble collagen from scale of spotted golden goatfish (Parupeneus heptacanthus). Food Chem. 2011, 129, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.D.; Li, L.; Yi, R.Z.; Gao, R.; He, J.L. Release kinetics of Tilapia scale І collagen peptides during tryptic hydrolysis. Food Hydrocolloid. 2018, 77, 931–936. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Yu, W.L.; Wu, J.P. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Turan, D.; Capanoglu, E.; Altay, F. Investigating the effect of roasting on functional properties of defatted hazelnut flour by response surface methodology (RSM). LWT-Food Sci. Technol. 2015, 63, 758–765. [Google Scholar] [CrossRef]
- Auwal, S.M.; Zarei, M.; Abdul-Hamid, A.; Saari, N. Optimization of bromelain-aided production of angiotensin І-coverting enzyme inhibitory hydrolysates from stone fish using response surface methodology. Mardrugs 2017, 15, 104. [Google Scholar]
- Fang, X.B.; Xie, N.N.; Chen, X.E.; Yu, H.; Chen, J. Optimization of antioxidant hydrolysate production from flying squid muscle protein using response surface methodology. Food Bioprod. Process. 2012, 90, 676–682. [Google Scholar] [CrossRef]
- Guo, Y.X.; Pan, D.D.; Tanokura, M. Optimisation of hydrolysis conditions for the production of the angiotensin-І coverting enzyme (ACE) inhibitory peptides from whey protein using response surface methodology. Food Chem. 2009, 114, 328–333. [Google Scholar] [CrossRef]
- Wu, S.G.; Sun, J.H.; Tong, Z.F.; Lan, X.D.; Zhao, Z.X.; Liao, D.K. Optimization of hydrolysis conditions for the production of angiotensin І-converting enzyme inhibitory peptides and isolation of a novel peptide from lizard fish (Saurida elongata) muscle protein hydrolysate. Mar. Drugs 2012, 10, 1066–1080. [Google Scholar] [CrossRef] [PubMed]
- Li, G.H.; Le, G.W.; Shi, Y.H.; Shrestha, S. Angiotensin I-converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- Liu, R.; Huang, Q.; Duan, J.A.; Zhu, Z.H.; Liu, P.; Bian, Y.; Tao, J.H.; Qian, D.W. Peptidome characterization of the antipyretic fraction of Bubali Cornu aqueous extract by nano liquid chromatography with orbitrap mass spectrometry detection. J. Sep. Sci. 2017, 40, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.J.; Zhu, Z.H.; Zhu, Y.; Qian, D.W.; Liu, R.; Peng, Y.R.; Ding, Y.H.; Oiyang, Z.; Duan, J.A. Characterization of collagen peptides in Elaphuri Davidiani Cornu aqueous extract with proliferative activity on osteoblasts using nano-liquid chromatography in tandem with orbitrap mass spectrometry. Molecules 2017, 22, 166. [Google Scholar] [CrossRef] [PubMed]
- Salampessy, J.; Reddy, N.; Kailasapathy, K.; Phillips, M. Functional and potential therapeutic ACE-inhibitory peptides derived from bromelain hydrolysis of trevally proteins. J. Funct. Foods 2015, 14, 716–725. [Google Scholar] [CrossRef]
- Liu, C.L.; Fang, L.; Min, W.H.; Liu, J.S.; Li, H.M. Exploration of the molecular interactions between angiotensin—І-converting enzyme (ACE) and the inhibitory peptides derived from hazelnut (Corylus heterophylla Fisch.). Food Chem. 2018, 245, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Pihlanto-Leppälä, A.; Koskinen, P.; Piilola, K.; Tupasela, T.; Korhonen, H. Angiotensin—I converting enzyme inhibitory properties of whey protein digests: Concentration and characterization of active peptides. J. Dairy Res. 2000, 67, 53–64. [Google Scholar]
- Ren, F.; Zhang, S.; Guo, H.; Jiang, L. Systemic screening of milk protein-derived ACE inhibitors through a chemically synthesised tripeptide library. Food Chem. 2011, 128, 761–768. [Google Scholar] [CrossRef]
- Nawaz, K.A.A.; David, S.M.; Murugesh, E.; Thandeeswaran, M.; Kiran, K.G.; Mahendran, R.; Palaniswamy, M.; Angayarkanni, J. Identification and in silico characterization of a novel peptide inhibitor of angiotensin converting enzyme from pigeon pea (Cajanus cajan). Phytomedicine 2017, 36, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Abdelmalek, B.E.; Gómez-Estaca, J.; Sila, A.; Martinez-Alvarez, O.; Gómez-Guillén, M.C.; Chaabouni-Ellouz, S.; Ayadi, M.A.; Bougatef, A. Characteristics and functional properties of gelatin extracted from squid (Loligo vulgaris) skin. LWT-Food Sci. Technol. 2016, 65, 924–931. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Zhang, H.; Wang, L.; Guo, X.N.; Qi, X.G.; Qian, H.F. Influence of the degree of hydrolysis (DH) on antioxidant properties and radical-scavenging activities of peanut peptides prepared from fermented peanut meal. Eur. Food Res. Technol. 2011, 232, 941–950. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay and properties of the angiotensin І-coverting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Wang, J.P.; Hu, J.N.; Cui, J.Z.; Bai, X.F.; Du, Y.G.; Miyaguchi, Y.; Lin, B.C. Purification and identification of a ACE inhibitory peptide from oyster proteins hydrolysate and the antihypertensive effect of hydrolysate in spontaneously hypertensive rats. Food Chem. 2008, 111, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, L.; Camp, J.V.; Morel, N.; Rougé, P.; Herregods, G.; Smaggle, G. Ala-Val-Phe and Val-Phe: ACE inhibitory peptides derived from insect protein with antihypertensive activity in spontaneously hypertensive rats. Peptides 2010, 31, 482–488. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Variable | Coded Levels | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| X1 | pH | 6 | 7 | 8 |
| X2 (%) | E/S | 1 | 2 | 3 |
| X3 (°C) | Temperature | 40 | 50 | 60 |
| X4 (h) | Time | 2 | 3 | 4 |
| Standard Order | Code Level of Variable | Response Value | ||||
|---|---|---|---|---|---|---|
| X1 | X2 (%) | X3 (°C) | X4 (h) | DH (%) | IP (%) | |
| 1 | −1 | −1 | 0 | 0 | 6.1 ± 0.1 a | 81.9 ± 0.8 a |
| 2 | −1 | 1 | 0 | 0 | 8.0 ± 0.1 b | 84.7 ± 0.3 b |
| 3 | 1 | −1 | 0 | 0 | 6.6 ± 0.1 c | 76.5 ± 0.4 c |
| 4 | 1 | 1 | 0 | 0 | 8.2 ± 0.1 d | 82.0 ± 1.0 a |
| 5 | 0 | 0 | −1 | −1 | 6.6 ± 0.1 c | 84.5 ± 0.9 b |
| 6 | 0 | 0 | −1 | 1 | 8.6 ± 0.1 e | 85.3 ± 0.8 b,d |
| 7 | 0 | 0 | 1 | −1 | 4.2 ± 0.2 f | 78.9 ± 0.9 e |
| 8 | 0 | 0 | 1 | 1 | 4.5 ± 0.0 g | 79.0 ± 0.0 e |
| 9 | −1 | 0 | 0 | −1 | 6.4 ± 0.1 h | 82.7 ± 0.4 a,f |
| 10 | −1 | 0 | 0 | 1 | 7.3 ± 0.1 i | 85.5 ± 0.8 b,g |
| 11 | 1 | 0 | 0 | −1 | 6.6 ± 0.1 c | 81.1 ± 1.2 a |
| 12 | 1 | 0 | 0 | 1 | 7.6 ± 0.0 j | 82.4 ± 1.0 a |
| 13 | 0 | −1 | −1 | 0 | 6.4 ± 0.1 h | 83.8 ± 1.2 b,f,h |
| 14 | 0 | −1 | 1 | 0 | 3.4 ± 0.0 k | 74.3 ± 1.0 i |
| 15 | 0 | 1 | −1 | 0 | 9.1 ± 0.1 l | 85.7 ± 0.9 b,g |
| 16 | 0 | 1 | 1 | 0 | 5.1 ± 0.1 m | 82.1 ± 1.3 a |
| 17 | −1 | 0 | −1 | 0 | 6.8 ± 0.1 n | 84.9 ± 0.5 b |
| 18 | −1 | 0 | 1 | 0 | 3.9 ± 0.1 o | 77.1 ± 0.4 c |
| 19 | 1 | 0 | −1 | 0 | 8.2 ± 0.1 d | 84.8 ± 0.1 b |
| 20 | 1 | 0 | 1 | 0 | 4.4 ± 0.1 g | 76.3 ± 0.7 c |
| 21 | 0 | −1 | 0 | −1 | 6.7 ± 0.0 c,n | 82.8 ± 0.7 a,h |
| 22 | 0 | −1 | 0 | 1 | 7.4 ± 0.1 i | 84.3 ± 0.5 b |
| 23 | 0 | 1 | 0 | −1 | 8.7 ± 0.1 e,p | 86.5 ± 0.4 d,g |
| 24 | 0 | 1 | 0 | 1 | 9.2 ± 0.1 l | 86.3 ± 0.2 d,g |
| 25 | 0 | 0 | 0 | 0 | 8.8 ± 0.1 p,q | 84.1 ± 0.4 b |
| 26 | 0 | 0 | 0 | 0 | 8.9 ± 0.1 q | 84.3 ± 0.4 b |
| 27 | 0 | 0 | 0 | 0 | 9.0 ± 0.1 l | 85.2 ± 0.3 b |
| Source | Mean Square | F-Value | p-Value | |||
|---|---|---|---|---|---|---|
| DH | IP | DH | IP | DH | IP | |
| Model | 5.555053 | 19.6431 | 49.53511 | 11.86095 | <0.0001 *** | <0.0001 *** |
| X1 | 0.096671 | 0.371496 | 7.187278 | 9.0352 | 0.0200 * | 0.0109 * |
| X2 | 0.096671 | 0.371496 | 103.2931 | 28.40672 | <0.0001 *** | 0.0002 *** |
| X3 | 0.096671 | 0.371496 | 300.8151 | 85.74476 | <0.0001 *** | <0.0001 *** |
| X4 | 0.096671 | 0.371496 | 22.47859 | 2.07395 | 0.0005 *** | 0.1754 |
| X1X2 | 0.167439 | 0.643451 | 0.174776 | 1.108634 | 0.6833 | 0.3131 |
| X1X3 | 0.167439 | 0.643451 | 1.648777 | 0.073968 | 0.2234 | 0.7903 |
| X1X4 | 0.167439 | 0.643451 | 0.026974 | 0.344194 | 0.8723 | 0.5683 |
| X2X3 | 0.167439 | 0.643451 | 2.365045 | 5.362187 | 0.1500 | 0.0391 * |
| X2X4 | 0.167439 | 0.643451 | 0.072229 | 0.421 | 0.7927 | 0.5287 |
| X3X4 | 0.167439 | 0.643451 | 6.827175 | 0.073968 | 0.0227 * | 0.7903 |
| X12 | 0.145007 | 0.557245 | 58.33257 | 13.20017 | 0.0001 *** | 0.0034 ** |
| X22 | 0.145007 | 0.557245 | 11.7709 | 1.276482 | 0.0050 ** | 0.2806 |
| X32 | 0.145007 | 0.557245 | 234.121 | 18.93139 | <0.0001 *** | 0.0009 *** |
| X42 | 0.145007 | 0.557245 | 19.70871 | 0.549077 | 0.0008 *** | 0.4729 |
| Residual | 0.112144 | 1.656115 | ||||
| Lack-of-fit | 0.131366 | 1.914652 | 8.193295 | 5.268234 | 0.1136 | 0.1700 |
| Pure error | 0.016033 | 0.363433 | ||||
| Cor total | 5.667197 | 21.299215 | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Liu, Y.; Wang, G.; Sun, S.; Liu, R.; Hong, B.; Gao, R.; Bai, K. Processing Optimization and Characterization of Angiotensin-Ι-Converting Enzyme Inhibitory Peptides from Lizardfish (Synodus macrops) Scale Gelatin. Mar. Drugs 2018, 16, 228. https://doi.org/10.3390/md16070228
Chen J, Liu Y, Wang G, Sun S, Liu R, Hong B, Gao R, Bai K. Processing Optimization and Characterization of Angiotensin-Ι-Converting Enzyme Inhibitory Peptides from Lizardfish (Synodus macrops) Scale Gelatin. Marine Drugs. 2018; 16(7):228. https://doi.org/10.3390/md16070228
Chicago/Turabian StyleChen, Junde, Ying Liu, Guangyu Wang, Shanshan Sun, Rui Liu, Bihong Hong, Ran Gao, and Kaikai Bai. 2018. "Processing Optimization and Characterization of Angiotensin-Ι-Converting Enzyme Inhibitory Peptides from Lizardfish (Synodus macrops) Scale Gelatin" Marine Drugs 16, no. 7: 228. https://doi.org/10.3390/md16070228