UV-Protective Compounds in Marine Organisms from the Southern Ocean
by
 , , ,
, , ,
Laura Núñez-Pons
1 ,
,
Conxita Avila
2 ,
,
Giovanna Romano
3 ,
,
Cinzia Verde
1,4 and 
Daniela Giordano
1,4,*
1
Department of Biology and Evolution of Marine Organisms (BEOM), Stazione Zoologica Anton Dohrn (SZN), 80121 Villa Comunale, Napoli, Italy
2
Department of Evolutionary Biology, Ecology, and Environmental Sciences, and Biodiversity Research Institute (IrBIO), Faculty of Biology, University of Barcelona, Av. Diagonal 643, 08028 Barcelona, Catalonia, Spain
3
Department of Marine Biotechnology (Biotech), Stazione Zoologica Anton Dohrn (SZN), 80121 Villa Comunale, Napoli, Italy
4
Institute of Biosciences and BioResources (IBBR), CNR, Via Pietro Castellino 111, 80131 Napoli, Italy
*
Author to whom correspondence should be addressed.
Mar. Drugs 2018, 16(9), 336; https://doi.org/10.3390/md16090336
Submission received: 12 July 2018
/
Revised: 3 September 2018
/
Accepted: 12 September 2018
/
Published: 14 September 2018
(This article belongs to the Special Issue Chilling Allelochemicals: Natural Products and Bioactivities from Polar and Sub-Polar Latitudes)
Abstract
:Solar radiation represents a key abiotic factor in the evolution of life in the oceans. In general, marine, biota—particularly in euphotic and dysphotic zones—depends directly or indirectly on light, but ultraviolet radiation (UV-R) can damage vital molecular machineries. UV-R induces the formation of reactive oxygen species (ROS) and impairs intracellular structures and enzymatic reactions. It can also affect organismal physiologies and eventually alter trophic chains at the ecosystem level. In Antarctica, physical drivers, such as sunlight, sea-ice, seasonality and low temperature are particularly influencing as compared to other regions. The springtime ozone depletion over the Southern Ocean makes organisms be more vulnerable to UV-R. Nonetheless, Antarctic species seem to possess analogous UV photoprotection and repair mechanisms as those found in organisms from other latitudes. The lack of data on species-specific responses towards increased UV-B still limits the understanding about the ecological impact and the tolerance levels related to ozone depletion in this region. The photobiology of Antarctic biota is largely unknown, in spite of representing a highly promising reservoir in the discovery of novel cosmeceutical products. This review compiles the most relevant information on photoprotection and UV-repair processes described in organisms from the Southern Ocean, in the context of this unique marine polar environment.
1. Introduction
Ultraviolet radiation (UV-R) is one of the most critical abiotic factors for life on Earth. In spite of the beneficial effects, sunlight can also threaten living organisms, and excessive UV-R of certain wavelengths can promote damage in their molecular machineries. Such deleterious processes can alter marine ecosystems productivity, thus affecting species diversity, ecosystem stability, trophic interactions, and global biogeochemical cycles [1] (Figure 1).
The ozone layer in Earth’s atmosphere acts as a shield by absorbing biologically harmful solar UV-B (290–315 nm). However, each spring, large ozone holes develop over the Southern Hemisphere, increasing the amount of UV-B that reach the Antarctic marine environments [2]. The ecological consequences of springtime ozone depletion are directly correlated with the tolerance of species to UV-B via photoprotective strategies to minimize UV exposure, and repair mechanisms for correcting UV-B-induced damage (Figure 1).
Antarctic species have developed a variety of adaptive strategies to mitigate the effects of solar UV-B radiation, including avoidance mechanisms, synthesis of UV-absorbing substances, enzymatic and non-enzymatic quenching of reactive oxygen species (ROS), and the activation of DNA repair pathways. However, there is a critical lack of information about the UV-B photobiology of Southern Ocean biota before the occurrence of springtime ozone depletion, and about the ecological consequences after the first depletion events in the 1970s [3]. Current research on the UV photobiology of Antarctic marine organisms is still poor and characterized by an old literature on the theme. In this review, we describe the major characteristics of the Antarctic marine environment by outlining the principal geophysical properties influencing the Southern Ocean (e.g., currents, UV-R, photoperiod, seasonality, ozone depletion, temperature, sea ice dynamics and global climate change) in the context of marine photobiology. The most relevant available information on UV-protective strategies has been summarized, with particular emphasis on suncreen compounds, molecular quenching and photodamage-repairing mechanisms. Such processes have been further compared to those described at other latitudes, in order to identify analogies/differences in the chemical structure of the molecules involved, their function and specific concentration.
1.1. Antarctic Marine Environment
Antarctica is detached geographically from the other continents, and isolated oceanographically and thermally by currents (e.g., the Antarctic circumpolar current (ACC)) defining sub-zero temperatures. It was originally part of the supercontinent Gondwana, which originated from Pangaea and began to break up c. 135 million of years ago (mya), during the early Tertiary. Antarctica reached the current geographic position at the beginning of the Cenozoic, 65 mya [4]. The region currently comprises the major continental land, the Maritime Antarctic, the sub-Antarctic islands, and the southern cold-temperate islands. Over the Cenozoic, in the last 40 my, the Antarctic shelf experienced cyclical glaciations that led to isolation from other oceans, as well as the establishment of colder conditions, and to major episodes of extinction of marine fauna [5]. The earliest cold-climate marine faunas are thought to date back to latest Eocene—Oligocene (35 mya) [6]. Extensive and thick ice sheets began to form periodically every 1–3 my, after the middle Miocene. Climate records from ice and sediment cores indicate that over the past 800,000 years, polar regions have gone through eight glacial cycles [7]. The last glacial cycle is dated on ~120,000–110,000 years ago, and culminated approximately 15,000 years ago [8]. At that time, the ice sheet thickened to more or less its recent configuration. The progressive separation from other continental masses allowed the establishment of the ACC and, at its northern border, the Antarctic Polar Front (APF), creating the isolated and cold habitat we know today. A southward shift of the fronts of the ACC has been suggested as the key mechanism for some of the observed Southern Ocean warming [9].
Antarctica is renowned as being the driest, windiest and coldest continent, boasting the lowest recorded temperature on Earth, −89.2 °C [10]. In terms of water surface, the Southern Ocean (including the Weddell and Ross Seas) is the planet’s fourth largest ocean. Water temperatures range between +1.5 and −1.9 °C at the most northerly and southerly latitudes, respectively [11]. There is little variation in temperature during seasons or as function of depth, because of the presence of the ACC and APF [12,13]. Along the APF, the surface layers of the north-moving Antarctic waters sink beneath the less cold and less dense sub-Antarctic waters, generating almost permanent turbulence [14]. Here the Ocean plays an important role in the global carbon cycle being responsible for ~20% of carbon dioxide (CO2) drawdown [10]. The deep water south of the APF, a roughly circular oceanic system extending to 2000 m in depth, brings to the surface dissolved nutrients and CO2, and then releases CO2 to the atmosphere. In contrast, water north of the APF takes up CO2 from the atmosphere thus making the Southern Ocean both a source and a sink for atmospheric CO2.
APF acts as a barrier for migration of marine organisms between the cooler Antarctic and the lower warmer latitudes [14]. Moreover, the cooling process over the last ~5 my, characterized by cyclical freezing and warming events accompanied by advances and retreats of the continental ice sheet, introduced new niches for faunal radiation [15,16]. More than 9700 species, mostly benthic, have been described from the Southern Ocean. The number of known species has significantly increased in the last years thanks to initiatives, such as the Census of Antarctic Marine Life [17]. Marine organisms thriving in the Southern Ocean are exposed to extreme conditions of isolation, harsh climate, low stable water temperature and viscosity, deep continental shelf and disturbance from scouring by icebergs [18,19]. Several studies revealed that despite some taxonomic connectivity that remains with South America through the Scotia Arc (which acted as biogeographic bridge between Antarctica and the Magellanic region [20]), the current benthic marine invertebrate fauna is largely ancient and endemic [21]. As such, the biota has had chances to co-adapt to this unique severe environment [22].
1.2. UV Radiation, Penetration, Photoperiod
Solar UV-R, the portion of the electromagnetic spectrum between X rays and visible light, was an important factor in driving the evolution and ecology of the biosphere until the development of the ozone layer and photoprotective mechanisms. UV wavelengths range from below 200 nm to 400 nm and are divided in vacuum UV (less than 200 nm), UV-C (200–290 nm), UV-B (290–315 nm), and UV-A (315–400 nm). The energy associated with a photon is inversely proportional to its wavelength; the higher is the energy, the greater is the capacity of UV-R to cause damage [23]. UV-R at the Earth’s surface varies with the season, time of day, latitude, and altitude. The local incidence of UV is determined by the total ozone column, cloudiness, ground reflectivity (i.e., the albedo), and local aerosols. In Antarctica, aerosols are almost absent, and the role of clouds is less important than in other parts of the planet. Consequently, surface UV in Antarctica is mostly driven by ozone and albedo [24].
The effects of UV-R on biogeochemical reactions in the sea (that is, mainly dissolved organic matter (DOM), bacterio- and phytoplankton) depend on the attenuation of the water column, that in turn depends on the optical properties of seawater itself, dissolved material, concentration of phytoplankton and suspended particles [1]. Colored or chromophoric dissolved organic matter (CDOM), yellow substance, or gelbstoff, is a result of tannin-stained decaying detritus from macroalgae and plankton, due to microbial degradation. CDOM can strongly absorb short wavelength light (from blue to ultraviolet), controlling the optical characteristics of freshwater and coastal habitats. This leads to the attenuation of both, photosynthetically active radiation (PAR, 400–700 nm) and UV-R, thus reducing UV exposure of organisms in the water. Absorption of solar UV-R causes the bleaching—photodegradation—of CDOM, reducing its optical density and absorptive properties. This not only increases the transmission of radiation in the water column, but it also originates low-molecular-weight organic compounds and ROS, which can be deleterious to marine organisms [1]. Even though concentrations are highly variable, coastal water and estuaries are usually richer in CDOM than the open ocean, where UV-R can penetrate deeper. Organisms living in oceanic waters may hence be adapted to higher and stable levels of UV-R [25]. Oligotrophic tropical waters (e.g., coral reefs) are in general more transparent to UV-R than temperate waters, because the optical properties depend on the water itself and not on dissolved constituents [26] (and references herein).
Antarctic waters are characterized by a low attenuation of UV-R, particularly during episodes of ozone hole. Here, the surface incidence can increase by 35% during spring time ozone depletion events [25]. In a comparative study around the Southern Ocean, Fildes Bay (South Shetland Is.) exhibited by far the highest UV penetration, recording 11 m for UV-B (313 nm) and 27 m for UV-A (395 nm), versus 2 and 4 m respectively in a sub-Antarctic Chilean fjord [27]. Smith et al. [28] found that in the ozone hole at Bellingshausen Sea effective UVR penetration could increase by 7 m and could be detected down to 60–70 m depth. Optical properties in Antarctic waters are further regulated by snow, ice cover and particulate material from runoff during melting [27]. Although annual sea ice may be considered a physical and protective barrier to UV-R transmission, it has been demonstrated that UV-B is transmitted through the Austral spring annual ice of McMurdo Sound, when the ozone hole occurs, causing mortality and DNA damage in the embryos of the sea urchin Sterechinus neumayeri. Higher mortality and DNA damage occur at 1 m below the ice with respect to 3 m and 5 m [29].
Seasonal change in day length (photoperiod) is widely used by organisms to regulate temporal patterns of development and behavior. The Antarctic continent experiences 24 h of continuous daylight at the summer solstice in December and 24 h of dark at the winter solstice in June. Therefore, at high latitudes, sunlight is strongly seasonal, and ice-free days around the summer solstice receive orders of magnitude more light than those in winter [30]. To remain viable under extended dark conditions, cells must retain membrane, organelle and DNA integrity, as damage is constantly occurring, due to metabolic heat and oxidation. Low temperatures in some way reduce such processes, but still polar phototrophic cells rely on different strategies to survive long periods of darkness. These include production of cysts or dormant stages, switch to heterotrophy (mixitrophic) or reduction of metabolic rates and rely on energy storage products. Spore production is quite uncommon in Antarctic phytoplankton, and has been observed only in some diatom species, and a few dinoflagellates. Facultative heterotrophy, in lieu, is more widespread. During the dark season pigments and UV-absorbing products decline to minimum levels, and upon the appearance of the first rays of light, most phytoplankton taxa are able to resume photosynthesis in hours. Furthermore, one seaweed species was able to reactivate within 24 h after a six-month period of total darkness (reviewed in Reference [31]). Despite lower temperatures and stronger seasonality, the overall photosynthetic efficiencies (the proportion of captured photons channeled to photosynthesis) are similar between higher and lower latitude ecosystems thus implying that the polar species (phytoplankton) are shade adapted [32].
1.3. Sea Ice Dynamics
Antarctic ice is one of the main features of the Southern Polar continent, covering ~99.6% of its land area and surrounding seas. This ice sheet extends an average of 30 × 106 km3 and represents 70% of the Earth’s freshwater. There are several typologies of ice, which prompt impacts to marine biota living on or in the ice, and to subtidal and underlying ecosystems. Freshwater ice derives from glaciers, it can extend up to a kilometer thick, and form the ice shelves attached to the land, and freely floating icebergs when large masses of ice detach [33]. The direct effects of iceberg scouring, and anchor ice, are major factors of physical disruption for benthic communities in Antarctic seafloors [34]. Sea ice, instead, is frozen seawater a few meters thick, and similarly subdivided into fast ice (attached to land) and ice floes (non-attached) [33]. In the Southern Ocean the ice cover is highly seasonal, spreading each winter far northward (to approximately 60° S) and experiencing a retreat almost to the coastline in the summer [35]. Antarctic sea ice mediates physical disturbance to the benthos. On the one hand it prevents drifting icebergs from scouring the seabed; and on the other it forms a barrier between the water column and the atmosphere, restricting wind-induced turbulence and water-column turnover [36]. These reduced land–ocean–atmosphere interactions interfere with the supply of nutrients (particulaly iron) to the marine environments [37]. Algae and particles trapped within sea ice caps seasonally constitute the basis of the Antarctic marine food chain when the packs melt, and phytoplankton blooms and organic matter reach the benthos [38,39]. Regarding the effects on sunlight incidence, ice sheets strongly modify the radiation budget and energy balance on the ocean surface by reflecting light (albedo). The regular albedo without snow is 6–7%, but it can exceed 85% in the presence of sea ice [40]. In underwater ecosystems, sea ice significantly attenuates solar irradiance [40] while protecting from UV wavelengths [41]. For instance, the Weddell Sea pack ice in the austral spring (September) showed an almost dark under-ice light regime with light transmittances bellow 0.1% [42]. For all the mentioned, the ecology and productivity in the Southern Ocean are strongly influenced by the sea-ice cover and its periodicity [43,44]. Each year Antarctic ecosystems go through a sort of lethargy marked by long dark cold winters, and reactivate in the summer upon the return of sunlight and the melting of the ice. These cyclic events influence all marine biota, but in particular photoautrotrophs and organisms depending on these for nutrition or light protection [43,45]. Contradicting most global climate models (including recent reports of Arctic ice declines [46]), Antarctic sea ice extent increased in the last decades [47]. These episodes of sea-ice expansion increased surface albedo, reduced ventilation, and enhanced CO2 sequestration to the deep ocean [48]. Nonetheless, since 2016/17 unprecedented springtime retreats in the Antarctic ice packs [49] highlighed the possibility of a switch to future declines in sea ice extent [39]. Reductions in the fraction of ice and snow cover will definitely influence the exposure of marine ecosystems to solar UV-R, and consequently the biology of photosynthetic organisms at the base of the food web, invertebrates and large predators along the ecological web (reviewed in References [35,50]).
1.4. The Ozone Hole and the Impacts of a Changing Environment
Although there are some contrasting opinions, it is now well-accepted that in early atmosphere there was no oxygen (O2) and therefore no UV protective ozone layer [51,52]. In these conditions, cells may have been confined to dimly lit regions of the oceans. The evolution of photosynthetic organisms as cyanobacteria resulted in oxygenation of the atmosphere and the formation of the ozone layer. When O2 accumulated in the upper atmosphere, it was photochemically transformed to ozone (O3), filtering out the shortest wavelengths of UV-R and thus changing the evolution of life on Earth [26,51,52]. The stratospheric ozone layer screens harmful UV-R from the Earth’s surface thus protecting against adverse effects on cells. The shortest and most damaging wavelengths UV-C are strongly absorbed in the upper atmosphere not reaching the stratospheric ozone layer. UV-B wavelengths are absorbed by ozone, which modify both the spectral quality and the intensity of UV-B radiation, allowing only a small amount to reach the Earth’s surface. The longest wavelengths UV-A are not absorbed by ozone layer and thus are not dependent by ozone concentrations [3].
In the early 1970s, scientists recognized that human actions producing chlorofluorocarbons (CFCs) could deplete the protective layer of ozone [53], destroying virtually all ozone between heights of 14 and 22 km over Antarctica [46], especially within the south polar vortex (persistent, large-scale cyclone) where temperatures are coldest. UV-R breaks down CFCs producing significant amounts of chlorine radicals that in turn react with ozone, catalyzing its destruction [54].
Currently, the ozone layer is diminished, particularly over the Southern Hemisphere and occasionally develops over the Arctic [55]. The “ozone hole” over most of Antarctica has grown in size (up to 27 million km2 in 2006, which is nearly twice the area of the Antarctic continent) and duration (from August through early December) over the past decades. At present (November 2017), the area is between 15–18 million km2 [56].
Although the photodissociation of CFCs can occur all across the Earth, the cold dark and long Antarctic winter favours, more than in any other part, the accumulation of chlorine in the Southern Hemisphere [57,58]. This leads to the destruction of ozone molecules in the spring, when the sunlight returns to the Austral latitudes from September to October [59].
The depletion in polar regions is larger than at lower latitudes, yet it accounts for only about 13% of Earth’s surface. Ozone depletion also develops at latitudes between the equator and polar regions: total ozone averaged for 2008–2012 has been about 3.5% lower in northern midlatitudes (35° N–60° N) and about 6% lower at southern midlatitudes (35° S–60° S) [60]. In the tropics (20° N–20° S latitude), total ozone has been only weakly affected by chemical ozone depletion, because of the lower conversion of ozone depleting substances (ODSs) to reactive halogen gases [60]. In tropical regions, however, coral reefs are currently experiencing the highest irradiance of UV-R at sea level on Earth even in comparison with the Antarctic region [26].
Thanks to the Montreal Protocol (1987), the chemicals responsible for the depletion of the ozone layer are now largely regulated. The tendency is to reduce the production and use of CFCs and regulate the emission of ODSs, including greenhouse gases (GHG) [61]. The Protocol has led to a successfully reduction of concentration of GHG in the atmosphere, this way mitigating the climate-forcing across the globe [62]. Recently, Kuttippurath and Nair [63] reported the first practical results from the Montreal Protocol. In fact, Antarctic ozone depletion has started to recover in both spring and summer thanks to the reductions in global ODS emissions and continuing recovery is expected to occur. However, changes in ozone levels are not only due to the halocarbons, since significant changes had already taken place, due to other source gases (e.g., N2O, CO2, CH4). Recently, the interest in the long-term recovery of the ozone layer refocused the attention on the effects of these gases on global mean ozone levels [64].
Interaction between climate change, ozone, and UV-R may be of considerable importance on the fate of marine organisms and entire ecosystems, and must be studied in a multifactorial manner in order to understand the impacts on our future oceans [65]. In the last 30 years, the Southern Ocean has changed notoriously [10,49], with profound implications in marine ecosystems, although some effects seem to be more regionally specific [37].
Ozone depletion is one of the major drivers of climate change in the Southern Hemisphere [66]. Higher air temperatures and incoming solar radiation are increasing the surface water temperatures of lakes and oceans, reducing annual snow and ice cover and, thus, increasing exposure to UV-R. As a consequence, warmer oceans are changing the composition of many marine ecosystems and their services and functions [65]. In the tropics and also the Mediterranean, temperature raises of the seawater have devastating effects, causing bleaching, disease and stress-related outbreaks on marine organisms [67,68,69]. Biotic networks in high latitudes seem to be considerably vulnerable [70,71], given that polar species are in general stenothermal and therefore, less capable of enduring temperature shifts [72,73]. The warming of the water along the Antarctic Peninsula has been five times faster than the global average over the past 50 years [65], with an increase of ~0.5 °C/decade since 1950 [74]. Reduction of the seasonal sea-ice, increased ocean temperatures both in the Weddell and Bellingshausen seas, regional retreat of glaciers, disintegration of floating ice shelves, expansion of terrestrial flora, and permafrost degradation are effects of the fast climate changes observed in the Antarctic Peninsula [49,75]. Finally, higher atmospheric CO2 concentration induces ocean acidification, alters seawater chemistry impairing the formation of UV-absorbing exoskeletons in many marine organisms, including phytoplankton, macroalgae, and animals, such as molluscs and corals. As an example, the shells of pteropods, key species in the food web, are already dissolving in areas of the Southern Ocean surrounding Antarctica [76].
2. Effects of Light in Marine Organisms
Life on Earth relies on sunlight [77]. The infrared rays of longer wavelengths (700 nm to 1 mm) are responsible for warming contributing to the benevolent temperatures of our planet; whereas the visible spectrum (400–700 nm, visible for human eye) supports the sense of sight. Remarkably, the visible light is also essential for photosynthesis, the process whereby autotrophic solar-powered organisms that are at the basis of most food networks derive their energy from photons. However, on the other side of the spectrum below 400 nm, UV sunlight exerts mostly deleterious effects on biological systems [77,78].
2.1. Beneficial Effects of Light
The sea covers about 71% of the planet’s surface and contributes to about one third of the global productivity. In marine systems above the aphotic zone, UV-R penetrates deeply and biota at all trophic levels become potentially exposed to UV-R [79]. The principal marine primary producers comprise planktonic diatoms, dinoflagellates, coccolithophorids, silicoflagellates, and blue-green and other bacteria, while benthic phototrophs include micro- and macroalgae, higher plants, and symbiotic producers, such as zooxanthellae in corals. All these organisms live in the euphotic zone to remain photosynthetically active. Zooplankton, herbivores, and other heterotrophs, in turn, largely depend on those photoautotrophs as their primary source of food. Marine biota depends directly or indirectly on light, for a number of biological processes [80,81]. Besides photosynthesis-related effects, there are other beneficial processes powered by light. For instance, beyond the photoautotrophic nourishment zooxanthellae provide to their coral hosts, these symbiotic dinoflagellates can further potentiate calcification and lipogenesis processes in the presence of light [82]. UV-R has also been shown to be necessary for spicule formation in some gorgonians, as colonies maintained in the absence of UV-R had significantly more “irregular” spicules when compared to colonies grown in the presence of UV-R [83]. For swimming organisms, the capacity of phototaxis may allow them to control their position in the water column, while avoiding excess of radiation [84]. UV photoreceptors have been described in bacteria, cyanobacteria, and algae, as well as in protozoans, annelids, cnidarians, molluscs, crustaceans, and fish, suggesting that UV vision may be relevant in aquatic systems [79,85]. UV photoreceptors may be used for navigation, communication, enhanced foraging, and possibly for UV-R avoidance. For instance, in the Antarctic krill Euphausia superba, a complex photoreception system, composed of different opsin photopigments, enables to respond to the daily and seasonal changes in light, moving downward during the day and upward during the night within the top 200 m of the water column [86]. Both negative phototactic behaviors and UV vision, suggest that UV-R may influence behaviour, migration and abundance patterns, as well as predator-prey and intraspecific interactions in marine environment [79,85]. UV-R may also play an essential role in the ecology of several infectious diseases of aquatic organisms, particularly when there is a pronounced difference in the UV tolerance of the host and the pathogen or parasite [65]. Simultaneously, solar radiation is very effective at reducing viral infections in some organisms, including fish viruses and harmful algal blooms, as well as some trematode worms infections [65].
Light is essential for the synthesis of vitamin D (calciferol) in most organisms, which has a significant role in calcium homeostasis, inmune system, and metabolism [87]. Its precursor, 7-dehydrocholesterol reacts with UV-B light at wavelengths between 270 and 300 nm, with peak synthesis occurrying between 295 and 297 nm [88].
Seasonal cycles in many organims may also be controlled by light, since light actually initiates different kinds of cycles as it increases in spring [80]. Duration and extent of the effects are variable. Vertical migrations, for example, happen usually within a day (diurnal cycles), while horizontal migrations are seasonal or annual. Intertidal organims also use light to adjust their optimum position relative to tidal height [82]. There are different types of mechanisms based on light to synchronize individuals of a given species, or to regulate a large number of activities [77]. Photokinesis, photoperiodicity, photosensibilization are among these mechanisms. Furthermore, circadian rhythms, which are endogenous, are also often regulated by light, and may also have intermediate controls, such as hormonal regulation [77]. Somatic growth and reproduction are usually coupled with seasonal cycles [82]. Synchronization may be also very useful for reproduction and for survival of the offspring, and many sponges, corals, and echinoderms, for example, spawn in a coordinated way related to light [82].
2.2. Negative Effects of Light
The ozone layer is continuously depleting and, as a consequence, there is an increase in the incidence of UV-R reaching the Earth’s biota that can be then absorbed by selected biomolecules (e.g., DNA, proteins porphyrins, carotenoids, steroids, quinones), causing direct damage in both plants and animals, and sunburn in humans [77,78]. The highly energetic wavelengths when absorbed by DNA [89] can cause damage (e.g., cyclobutane pyrimidine dimers, CPDs), and mutations either directly by absorption or indirectly due to the production of ROS [90]. Indeed, a routine way in which UV-R can harm marine organisms is via photochemical reactions and generation of ROS. Reduced O2 intermediates, such as hydrogen peroxide (H2O2), superoxide radicals (O2−•), hydroxyl radicals (•OH) and singlet oxygen (1O2) are produced as a result of electronic excitation after UV-R absorption and reduction of molecular O2. Most of the production of ROS involves the activation of intermediate molecules in cells (e.g., aromatic amino acids), which absorb UV-R, and enter into an excited state leading to the production of extremely reactive hydroxyl radicals in an iron-catalyzed Fenton reaction. UV-A-generated ROS trigger several toxic responses in organisms, including impair of DNA, enzymes, membrane proteins and lipids (especially those containing polyunsaturated fatty acids), as well as photooxidative stress of photosystem components in photoautotrophs (see in more detail in sections below; [91,92]).
In organisms that perform oxygenic photosynthesis, an excess of UV-B light can interfere with the thylakoid photochemistry, leading to a decrease in O2, electron transport, Rubisco activity and CO2 fixation rates [93,94]. Such processes consequently lead to photoinhibition or photoinactivation, bringing repercussions to the first levels of foodwebs [81,95]. Similarly, corals are affected by strong UV light if this impairs the photosynthetic capacity of their symbiotic algae resulting in reduced carbon supply. This consequently leads to decreased growth and calcification, reduced photosynthesis and changes in respiration, DNA damage, oxidative stress and eventual mortality, as well as adverse effects on reproduction, larval development, and settlement [26].
The tolerance levels and responsive behaviours to UV stress can be different from species to species. UV-R can induce relevant changes in population compositions and trophodynamics, shifting communities towards more UV-tolerant species [79]. UV-R may affect organisms directly by producing cellular and/or tissue damages, or it may also affect organisms indirectly by constraining them to suboptimal habitats where temperature and food abundance may be low and the predation risk high [79]. Elevated UV-B radiation affects the survival of phytoplankton by decreasing their motility and inhibiting their phototactic and photophobic responses [78,96]. For zooplancton, UV-B irradiation may cause irrevesible damage and/or death, and decrease the fecundity of survivors [97,98,99]. These phenomena could eventually lead to a decrease in invertebrate and fish populations that feed on zooplankton, and be transmitted along the trophic chain, eventually affecting humans. Juvenile polychaetes showed reduced growth and development of tentacles when fed detritus derived from diatoms previously exposed to artificial UV-B radiation versus diatoms that were not pre-exposed to UV-B radiation [100]. Apoptosis is promoted due to UV-R in developing sea urchin embryos [101]. Algae and seagrasses experience physiological, biochemical, morphological, and anatomical changes towards UV-R, with deleterious effects on growth, reducing leaf size and limiting the area available for energy capture [78]. The vertical distribution of seaweeds in their ecosystem is indeed strongly determined by solar UV-R. Specially developing brown and red algae are particularly sensitive [102]. Increased solar UV-R can also reduce recruitment and impact heterotrophic species, as well as primary producers [103].
Very few studies exist on the repercussions of UV-R in Antarctic marine communites. Some examples include the study on the viability of bacterioplankton, which was shown to decrease with depth, with no significant inhibition at 9.5 m depth [104]. UV-B radiation inhibits the growth of Antarctic sea ice microalgae Chlamydomonas sp. ICE-L, especially at high intensity [105]. In some macroalgae UV-R lowered germination, showing that at unicellular life stage there was a strong species-specific susceptibility to changes in the UV-R [106]. This is important in determining the upper distribution limit of Antarctic seaweeds, which affects the community structure. Propagules of three Antarctic intertidal macroalgal species Adenocystis utricularis, Monostroma hariotii and Porphyra endiviifolium, particurarly sensitive to environmental perturbations, showed no long-lasting negative effects to UV demonstrating the possession of good repair and protective mechanisms, necessary condition for the ecological success in intertidal habitats [107]. UV-B-driven DNA damage and mortality in Antarctic sea urchin embryos has been found to vary from year to year, depending on the thickness of the sea ice and total column ozone [29].
3. UV Photoprotection in Marine Organisms: Antarctic and Non-Antarctic Strategies
Marine organisms have developed physiological and biochemical traits to cope with UV. The choice of habitat is the most effective defence, and consists of avoidance mechanisms, such as cyclic migrating behaviours from high to low UV-R levels in a diel or seasonal manner, or translocation to shaded or deeper zones along the water column. Many cyanobacterial communities in Antarctica live in dim-light environments, such as within or beneath rocks, in permanently ice-covered lakes, beneath the surface of the soil or at the base of the plants within moss banks [108]. In the Antarctic cyanobacteria Oscillatoria sp. vertical migration of the microbial mat reduces the exposition to UV [109], whereas vertical mixing of the water column provides similar effects in planktonic organisms [96].
For organisms living exposed to sunlight, mechanisms for minimizing UV damage include: (1) Screening mechanisms for reducing UV exposure by physical barriers or chemical barriers with UV-absorbing compounds; (2) quenching mechanisms by non-enzymatic (carotenoids, α-tocopherol, ascorbic acid, glutathione) and enzymatic antioxidants, such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase and other enzymes that can neutralize effects of radicals produced by UV photochemical reactions; and (3) repair mechanisms to deal with UV-induced damage that occur in DNA, proteins and lipids.
3.1. Physical Structures for Light Avoidance
Physical barriers consist of morphological or structural features that represent the first line of defence for preventing physical external injuries (e.g., from predators and enemies’ attacks), such as the calcified shells of gastropods and crustaceans or spines of echinoderms. Some structural devices also protect from harmful light forming an opaque barrier between the UV rays of the Sun and the body [3]. For instance, ostracods living in shallow benthic habitats possess shells that block 60–80% of UV-R, whereas the exoskeleton of planktonic Daphnia can block up to 35% [110]. Embryos of intertidal gastropods are also protected by egg capsules [111], even if they combine presence of photoprotective molecules [112]. Similarly, embryonic developing modes in limpets, Crepipatella spp., rely on sunscreen products prior to developing a protective shell, and adult females can transfer them to capsule walls, embryos and nurse eggs, according to their type of embryonic development [113]. The spicules, at least in the colonial didemnid ascidian Didemnum mole, are potentially related to UV-R photoprotection in shallow water colonies [114]. The tunics of some ascidians are pigmented and can absorb UV-R providing photoprotection [115].
A number of marine organisms produce copious mucous secretions that provide physical protection against sediments, desiccation and predation and might also decrease the damaging effects of UV-R. This UV-R-screening capacity may be attained by the optical properties of the mucous itself, but also because of the presence of UV-R-absorbing compounds that are excreted within mucous, e.g., very common in corals [26,116,117,118].
Cyanobacteria synthesize extracellular polysaccharides (EPS), high-molecular-mass heteropolysaccharides, which provide a protective matrix from UV stress [119]. In Antarctic cyanobacterial mats from the McMurdo ice shelf, EPS were found to be involved in matrix formation and also in the attachment to the substrate [120]. Other Antarctic marine bacteria produce EPS, which have proved to serve as ligands for trace metal nutrients and cryoprotection at low temperature and high salinity [121,122], but they may be as well involved in UV protection even if this function has not been reported yet.
Some EPS from marine bacteria are already commercially available in cosmetics under the name of Abyssine® for soothing and reducing irritation of sensitive skin against chemical, mechanical and UV-B aggression [123].
In large-celled species, including some microalgae, plant pollen and spores, sporopollenin, biopolymer of variable composition that functions as antimicrobial agent, and confers rigidness to the cell wall [124] may further protect from UV-R by increasing the optical density. It was reported that species of microalgae, that were highly tolerant to UV-R had substantial amounts of sporopollenin, whereas species containing little or no sporopollenin were highly UV-R susceptible [124]. In the pollen and spores of Antarctic plants, sporopollenin is considered as bio-indicator of solar UV-B and a valuable archive for the reconstruction of past solar UV-B [125].
Some unicellular organisms display transitory mechanisms to screen UV. Symbiotic Symbiodinium algae, for instance, can produce multiple layered cell walls when exposed to UV-R, which disappear when returned to lower light conditions [126]. Moreover, the dinoflagellate Scrippsiella sweeneyae can increase in cytoplasm volume when exposed to UV-R, possibly to lengthen the path that damaging photons have to travel to reach internal components, e.g., DNA [127]. Actually, the amount of DNA damage in Antarctic phytoplankton is inversely correlated with the cellular size; smaller cells with higher concentrations of photoproducts in their DNA are more sensitive to UV-B than larger phytoplankton species and undergo greater damage [96].
3.2. UV-Absorbing Substances (Sunscreen)
The best known photoprotective response in marine organisms is the production or accumulation of UV-absorbing compounds, including mycosporine-like amino acids (MAAs) as the most common compounds with such properties, but also others, such as scytonemin, 3-hydroxykynurenine, melanin, various secondary metabolites and fluorescent pigments [118,128,129,130,131].
3.2.1. Mycosporine-Like Amino Acids (MAAs)
Structure, Biosynthetic Pathways of MAAs and Their Regulation
MAAs are small (<400 Da) intracellular, colorless water-soluble secondary metabolites of low molecular weight, commonly found in marine environments. Their name comes for being imino carbonyl derivatives of mycosporines—a group of compounds first identified in the mycelia of fungi as compound P310, and hypothesized to act as photoprotectants during sporogenesis [132]. Novel molecular species (characterized solely by their maximal light absorbance) are constantly being discovered; to date ~30 MAAs have been resolved, of which 11 (1–11), shown in Figure 2, have been reported in the Southern Ocean (see below). Their designation consists on the name and value of absorbance (e.g., euhalothece-362; Reference [133]). Characterization of MAAs should be treated with caution, as often it has been done by indirect comparisons on co-chromatography with sub-standards and/or with published UV spectral data and HPLC retention times [45,134,135]. MAAs are enamino ketones that contain a central aromatic aminocyclohexenimine or amonicyclohexenone ring and a wide variety of substitutions. These aromatic cyclohexenimine or cyclohexenone structures are responsible for the light absorption properties and accommodation of free radicals related to enhanced solar UV-R (see below; Reference [136]). The core cyclohexenone unit is derived from the first steps of the shikimate pathway, where 4-deoxygadusol—a strong antioxidant—is a direct precursor [137]. The most common MAAs contain a glycine moiety on the C3 of the cyclohexenimine ring and a second amino acid (porphyra-334 (9), shinorine (10), mycosporine-2-glycine (3), mycosporine-glycine-glutamic acid), amino alcohol (palythinol (8), asterina-330 (1)) or an enaminone system (palythene (5), usujirene (11)) linked to the C1. Eventually, further substitutions added as as side chains can take place [138].
The shikimic pathway is a metabolic route used by bacteria, cyanobacteria, fungi, algae, some protozoan parasites and plants for the biosynthesis of folates and aromatic amino acids (phenylalanine, tyrosine, and tryptophan), as well as of higher plant photoprotectants (i.e., flavonoids; [137]). It involves several enzymes that have been recently reported also in metazoan organisms; however, these are believed to still derive mostly from associated microbiota (bacteria, dinoflagellates). Indeed, MAAs are commonly described as “microbial sunscreens” [139,140]. Interestingly, Osborn et al. (2015) [141] recently reported that fish can de novo produce on their own gadusol—an antioxidant and UV-protective compound related to MAAs synthesis, and that analogous pathways are shared in amphibians, reptiles, and birds.
In many microorganisms MAAs are found in the cytoplasm where they are produced [142], yet in some cases, as in the cyanobacterium Nostoc commune, they can be excreted and extracellularly accumulated, showing more effective protection against UV-R [143,144]. The induction of MAAs synthesis has been proposed to be triggered via two disparate signal transduction pathways: One activated by UV-R and the other related to salt stress [145]. A UV-B specific photoreceptor called pterin involved in MAA production has been described in cyanobacteria [146]. In the red alga Chondrus crispus a receptor with absorbance peaks at 320, 340 and 400 nm (UV-A) has been linked to the formation of shinorine (10) [147]. Light-induced synthesis of MAAs can be as rapid as several hours in organisms like dinoflagellates Alexandrium excavatum and Prorocentrum micans, that experience rapid light changes during vertical migration [148], but it is usually slower [149,150,151].
Biological Functions of MAAs
MAAs have the capability to absorb light between 309 and 362 nm and dissipate radiation as heat without producing ROS [134]. The presence of a suite of MAAs extends the photoprotective potential and allows the harboring organisms thrive across a broader light spectrum [45,162,163].
The photoprotective role of MAAs has been demonstrated in several assays where extracts enriched in these UV-R absorbing substances significantly reduced the production of deleterious thymine photodimers by direct molecule-to-molecule energy transfer process [164]. MAAs are multi-functional sunscreens that exhibit photostability and resistance to several abiotic stressors [45]. Besides protecting cells from mutation caused by UV-R and free radicals, they are also effective antioxidant molecules via stabilizing free radicals within their ring structure scavenging ROS [133]. Mycosporine-glycine (2) in particular, can display several antioxidative properties, and this probably explains why it is the most frequently observed (and often abundant) MAA, in tropical environments, especially in cnidarians [165,166,167,168,169,170]. Mycosporine-glycine (2) yields rapid protection against oxidative stress, even prior the intervention of antioxidant enzymes [171] and this is achieved by scavenging free radicals [172] and quenching singlet O2 [173].
It has beed proposed that MAAs can further act as osmolytes and boost cellular tolerance to desiccation, salt, and heat stress [142]. Even if mycosporine-glycine (2) and shinorine (10) can be induced by salt stress, their role as osmolytes seems to be still ambiguous [145]. There are studies demonstrating their efficacy in reducing osmotic stress [174], however others report an insignificant contribution of MAAs as compared to other osmolytes [145]. MAAs can also act as osmoprotectants under freezing conditions [143,175]. Due to their high nitrogen content, a role as nitrogen intracellular reservoirs [143,176,177]. In marine reefs, under nitrogen-limiting conditions starved corals have beed observed to preferentially accumulate MAAs in higher rates than protein and chlorophyll a (Chl a) [178]. Lastly, particularly in species establishing symbiosis with Symbiodinium zooxanthellae, MAAs together with other free amino acids have been proposed to facilitate the exchange of photosynthates between symbiont and host, and hence act as “host factors” [179].
Environmental Distribution of MAAs and Their Occurrence in Organisms
MAAs have been found from tropical (e.g., Reference [165]) to Antarctic waters (e.g., Reference [84]) and in a variety of organisms [180], spanning from cyanobacteria (e.g., Reference [181]), microalgae (e.g., Reference [182]), fungi (e.g., Reference [183]), as well as macroalgae (e.g., Reference [184]) and animals—invertebrates and vertebrates (e.g., References [185,186]). Among animals, protozoans, poriferans, cnidarians, platyhelminthes, nemerteans, polychaetes, molluscs, bryozoans, rotifers, arthropods, echinoderms, tunicates, and fish have also been reported to protect themselves from UV-R by MAAs [180]. To date, few reports have described the production of MAAs in bacteria [187,188,189]. In most organisms, there is an expected positive correlation between MAA concentration and solar UV-R: MAAs contents vary seasonally, peaking during the summer months [190], they diminish with depth [165,167,168,191,192], and under shaded conditions [157,193], as well as if UV-R irradiance is filtered out manipulatively [149,166]. Interestingly, some free-living dinoflagellates synthesize and release MAAs into the water, which was interpreted to contribute to the attenuation of UV-R during algal bloom events [194]. Organisms may further compensate the levels of dietary MAAs with behavioural traits to reduce UV damage. For example, the sessile anemone Actinia tenebrosa displayed larger seasonal fluctuations of MAAs than the mobile intertidal gastropod Diloma aethiops from New Zealand [195].
About 95% of tropical, 80% temperate and 82% of polar species studied have detectable amounts of MAAs [184,185,196]. Palythine (7), shinorine (10), mycosporine-glycine (2), porphyra-334 (9), asterina-330 (1), palythinol (8), palythene (5), and mycosporine-2-glycine (3) are by order the most common MAAs in marine biota, and are found in all latitudes. Other MAAs, instead, have more restricted distributions, but this might be biased, due to limiting representative data. Characteristically, mycosporine-glycine-valine (11) seems to be more associated to Antarctic ecosystems [185]. By phylum, cnidarians seem to possess the highest diversity, in part because there are more publications on this phylum [45]. In particular, scleractinian corals have attracted attention for their study, as they are the major bioconstructors of the impressive tropical reefs. MAAs are common in microalgal-invertebrate symbioses on coral reefs and other habitats [131]. At least 11 different MAAs have been reported in corals particularly in their mucous, being palythine (7) and mycosporine-glycine (2) the most abundant [118,169,170,197]. There are species reflecting dietary MAAs with presence exclusively in the coral host [126,161], while others require provision of these sunscreen products from their symbiotic zooxanthellae cells (Symbiodinium) [162,198], and/or prokaryotic partners (e.g., bacteria in the genus Vibrio sp.; [159]). The host organism may also play an important role in protecting symbionts from UV-R damage, for instance some ascidians accumulate MAAs in the tunic, which prevents photoinhibition of Prochloron symbionts [199]. In macroalgae (from tropics to the poles) shinorine (10) and porphyra-334 (9) are by far the most common MAAs. Red algae are the largest producers (Rhodophyceae—but with notable variability among species), followed by Phaeophyceae, and last a few green algae—Chlorophyte and Charophyte [84,180,200]. Banaszak et al. [170] however, reported mycosporine-glycine (2) as the most abundant MAAs in temperate macrophytes, especially in Chlorophyte and Phaeophyte.
The dietary uptake of MAAs can be selective [160,201]. For example, the sea urchin Strongylocentrotus droebachiensis principally accumulated shinorine (10) in the ovaries, but not palythine (7), asterina-330 (1) and usujirene (11) that were available in high concentration in their diet [201]. Moreover, subsequent biochemical or possibly bacterial (endosymbionts) conversion of acquired MAAs can sometimes increase the variability of these compounds in certain organisms [202]. This is likely the reason why some host species harbor more MAAs than their symbionts or prey [135,149,161]. On the contrary, when an organism contains fewer MAAs than the symbiont or its prey it is possible that some MAAs were not eventually incorporated or were degraded [140]. Photoprotectants can also be transmited vertically to the offspring, as a mechanism to favour larval survival [158]. For instance, in several species of soft corals MAAs contents exhibited a peak in female colonies prior to spawning in comparison to male counterparts (e.g., up to a 67% and 56% difference for Lobophytum compactum and Sinularia flexibilis, respectively) [190,203]. When embryos of some echinoderms fed on algal diets with richer MAAs contents, these obtained photoprotection against abnormalities induced by UV-B in the first stages of life [191,201,204]. Shinorine (10) indeed, was prevalent in the ovaries and eggs of sea urchins from polar to tropical biomes [170,185,191,201]. Other than to eggs and reproductive tissues, marine animals have been described to allocate MAAs to specific tissues—e.g., holothuroids in the epidermis [131,157], tridacnid clams in siphonal mantle and kidney [205], didemnid ascidians in tunic bladder cells [206], corals in mucous exhudates [207], and teleost fish in ocular tissues [160,186].
MAAs in Antarctic Marine Organisms
The deleterious effects of UV-R are presumed to be particularly acute in the Southern Ocean, and even more in the superficial layers of the water column [63]. Consequently, the planktonic fraction in this region is likely to rely much on MAAs and other photoprotective agents, and there are several studies corroborating this assumption (e.g., References [116,208,209,210,211,212,213,214]. Antarctic phytoplankton assemblages are rich in MAAs, especially when dominated by prymnesiophytes in the genus Phaeocystis [116,211], as well as by chain-forming Thalassiosira diatoms (e.g., Thalassiosira gravida; [209]). These assemblages exhibit though very different MAA compositions—e.g., Phaeocystis antarctica blooms have more complex profiles (mycosporine-glycine (2), shinorine (10), porphyra-334 (9), palythine (7), palythinol (8) and palythenic acid (6)), whereas those formed by Thalassiosira diatoms are dominated by shinorine (10) and porphyra-334 (9), and at times include mycosporine-glycine (2) [154,213,215]). Hernando et al. [154] found that UV-B induced the expression and accumulation of MAA in a Thalassiosira sp. diatom. Instead, more latitudinal cosmopolitan blooms, such as those formed by the prymnesiophyte coccolithophorid Emiliania huxleyi display restricted UV-R tolerance, linked with low concentrations of only shinorine (10) [153]. Contrasting to bloom-forming algae, MAAs seem to play a minor role as photoprotectants in sea ice algae, where they are found in low amounts. It has been suggested that the ice algal structure may provide a self-shading effect, thus confering protection on the community as a whole [216]. The Antarctic krill Euphausia superba recorded a variety of at least nine MAAs [84], likely from dietary phytolankton uptake [217]. However, it was particularly rich in rare isomeric forms of palythenic acid, suggesting Z/E isomerization during assimilation [218]. A characteristic case study among Antarctic plancto-pelagic organisms is that of the pteropod predator Clione antarctica, and its exclusive prey, the herbivorous pteropod Limacina helicina, both containing the same five MAAs, mycosporine-glycine (2), shinorine (10), porphyra-334 (9), palythenic acid (6), and palythine (7). Strickingly, the phytoplankton assemblage, on which Limacina helicina feeds on has only shinorine (10) and porphyra-334 (9) [135], suggesting that there is likely the possibility of subsequent biochemical or bacterial interconversions of shinorine (10) and porphyra-334 (9) [202], or perhaps other minor food sources.
Most of the knowledge about the use of UV protectants by subtidal benthic Antarctic marine organisms comes from the surveys of Karentz and co-workers [84,219] near Palmer Station and from McMurdo Sound areas. Regarding seaweeds, around 25 species have been studied for MAAs contents [84,219,220,221,222]. While shinorine (10) and porphyra-334 (9) are the most common in macroalgae from tropical to polar waters [84,180,184,223], Antarctic red algae (i.e., Palmaria decipiens, Iridaea cordata, Curdiea racovitzae, Kallymenia antarctica) showed more complex profiles, comprising shinorine (10) and porphyra-334 (9), palythine (7), asterina-330 (1), palythinol (8), palythene (5), usujirene (11) and the unusual M335/360 [84,180,219,220,221,222]. Such higher MAAs diversity may be consequence of interconversions among primary and secondary MAAs [202].
Karentz and co-workers [84,219] found in general low levels and abundances of MAAs in marine organisms inhabiting subtidal (>20 m depth) Antarctic seafloors from Palmer Station and McMurdo Sound. The reason for this trend was suggested to be trophic related, as the majority of those benthic species were not herbivorous. To the best of our knowledge, the species analyzed for MAAs contents so far include: 18 Porifera, 2 Cnidaria, 2 Platyhelminthes, 3 Nemertea, 15 Mollusca, 6 Annelida, 14 Arthropoda, 2 Bryozoa, 7 Echinodermata and 5 Chordata (of which three are ascidians and two are fish). Eleven MAAs (mycosporine-glycine, mycosporine-2-glycine, shinorine, porphyra-334, palythine, mycosporine-glycine-valine, asterina-330, palythene, palythinol, palythenic acid and usujirene (1–11)) were detected among these species, but four—mycosporine-glycine (2), shinorine (10), porphyra-334 (9) and palythine (7)—where present in >50% of the cases [84,219]. Similar as in some echinoderms from non-Antarctic latitudes, the ripe ovaries and testes of the sea urchin Sterechinus neumayeri and the ripe ovaries and brooded young of the sea cucumber Cucumaria ferrari were enriched in MAAs [219]. In a subsequent study monitoring temporal changes of MAAs in tissues and depth, Sterechinus neumayeri reported the highest concentrations in the ovaries, varing according to the spawning cycle and with depth [191].
Table A1 reports the list of MAAs found in Antarctic marine organisms available in the literature up to now.
3.2.2. Scytonemin
Some terrestrial cyanobacteria living in habitats exposed to full sunlight produce scytonemin (12) [224], shown in Figure 3. It is a yellow-brown lipid soluble sheath pigment that absorbs maximally in the UV-A and UV-C regions, but with some absorbance in the UV-B region [225]. Scytonemin is composed of a dimeric structure of indolic and phenolic subunits having a molecular mass of 544–546 Da, with an in vivo absorption maximum at 370 nm. Both, temperature increase and oxidative stress combined with UV-A have a synergistic effect on the synthesis of scytonemin (12) [226]. In Antarctica the primary genera rich in sheath pigments are Gloeocapsa and Calothrix, which form black or brown crusts over rocky streambeds, and Nostoc, which forms black, mucilaginous films and mats up to several centimeters thick along stream banks and in slow moving waters [108]. Scytonemin (12) has been identified in Nostoc commune and N. microscopicum isolated from fresh water pond fringe in Mc-Murdo Ice Shelf, Antarctica [224]. Cyanobacteria may exist in Antarctic sea ice, but contributing insignificantly to the marine ecosystems [227]. They are instead important components of Antarctic terrestrial and freshwater microflora, colonizing rocks, and found in lakes, ponds, meltwater holes and streams [224].
3.2.3. Erebusinone
A yellow pigment, erebusinone (13), shown in Figure 4, has been found in the Antarctic sponge Isodictya erinacea [228]. It shares the same aromatic substitution pattern of 3-hydroxykynurenine. 3-Hydroxykynurenine is a water soluble, low molecular weight, tryptophan derivative that occurs in the lens pigments of several species of marine and freshwater fish [229,230]. It absorbs in the UV-A region with a peak absorbance of 370 nm. Such photoprotective properties increase visual acuity by reducing glare, scatter and chromatic aberration while maximizing contrast, this way aiding prey detection or functioning as a stabilizing lens protein [230]. Erebusinone shares similar absorbance properties as 3-hydroxykynurenine, and biogenesis from the tryptophan catabolic pathway. Erebusinone (13) was evaluated for bioactivity against the sympatric predator amphipod Orchomene plebs, causing reduced molting and increased mortality at ecologically relevant concentrations. This seems to be the first example of molt inhibition as a mechanism of chemical defence in the marine environment [228].
Table A2 shows all known UV-absorbing compounds found in the Antarctic marine environment that are not MAAs.
3.2.4. Pigments
Melanin (from the Greek melas, “black, dark”) (14), shown in Figure 5, is produced by the oxidation and polymerization of tyrosine. Melanin is a UV-absorbing compound and belongs to a group of pigments responsible for dark, tan, and even yellowish or reddish pigmentations, due to the aerobic oxidation of phenols. It is a polymer of either or both of two monomer molecules: Indolequinone and dihydroxyindole carboxylic acid [231].
Because melanin is an aggregate of smaller component molecules, by changing the proportion and bonding pattern of the component molecules, a wide number of different types of melanins can be produced [231]. It is an effective light absorbent at all UV-R and PAR wavelengths able to dissipate over 99.9% of absorbed UV-R [232], and thus is a wide-ranging sunscreen in non-photosynthetic organisms (e.g., Reference [233]). In the skin of fish (e.g., genus Xiphophorus), for instance, melanin could lower the rate of pyrimidine dimers formation caused by exposure to various UV-R [234]. Melanin can have diverse functions in various organisms. It has been described to act as a free radical scavenger and energy transducer, majorly in microbial fungi and bacteria [235,236,237]. Some arthropod species have deposits of melanin in layers that yield an iridescent color by alternating refractive index effect—Bragg reflector [238]. In invertebrates, an immune response to invading pathogens called “melanisation” has been described, by which microbes are encapsulated within melanin. The consequent generation of free radical byproducts is thought to aid in their elimination [239]. In cephalopods instead, melanin takes part of the ink as distracting-scape defence against predators [240].
Alteromonas stellipolaris, a bacterium from Antarctic seas, produces a brown-black pigment characterized as melanin [241]. Melanin has been also identified in Lysobacter oligotrophicus, a Gram-negative bacterium isolated from an Antarctic freshwater lake [242], and in black fungi found in the Antarctic terrestrial biotopes [243,244].
Examples of melanin found in the Antarctic marine environment are reported in Table A2.
Some other UV-absorbing pigments with no specified name were detected in the Antarctic algae Palmaria decipiens and Enteromorpha bulbosa [245]. Many pigments that confer bright colourations to Antarctic sessile organisms (e.g., sponges), such as the mentioned erebusinone (13) from Isodictya erinacea, but also variolins from Kirkpatrickia variolosa, discorhabdins in Latrunculia apicalis, suberitenones from Suberites sp., and the yellow isoquinoline pigment from Dendrilla membranosa possess intriguingly striking bioactivities. It has been proposed that such colourful molecules could be the result of relict pigments originally retained for aposematism or UV screening, and then conserved because of their beneficial defensive properties, such as feeding deterrents or antifouling (revised in Reference [231]).
3.2.5. Other Secondary Metabolites
Phlorotannins
Phlorotannins are polymers of phloroglucinol (1,3,5-trihydroxybenzene), a type of tannins analogous to the shikimate-derived polyphenolics, including more than 150 compounds (e.g., (15–19), shown in Figure 6). Their molecular weight ranges from 10 to 650 kDa. They exhibit strong absorption from 280 to 320 nm [246,247] and their production is induced by exposure to UV-B, in some cases probably after external wounding or herbivory [248]. Their presence is particularly abundant in cell walls of brown algae (~10–20% dry weight; [246]) and can be found in low amounts in red algae. This may explain why MAAs are virtually absent in brown algae in comparison to red and green algae [84,170,220,249]. Phlorotannins protect algal cells against UV-R damage, but they can also be exudated in the surrounding water [246,250]. Moreover, phlorotannins may have several other roles (reviewed in Reference [251] including antioxidants as efficient ROS scavengers [252], antiherbivory defences (e.g., Reference [247]), structural function as part of brown algal cell walls and implicated in cytokinesis [249], reproduction agents in fertilization processes as spermatozoan inhibitors [253], wound healing factors [254], algicidal effect against some dinoflagellates [255], and heavy metals sequesters [246,256]. In fact, a combination of both UV-screening phlorotannins and major antioxidant enzymes may be used to respond to unfavourable light conditions [257]. As an example, it has been demonstrated that the influence of UV-R on biological processes is dependent on the exposure time: Short-term responses are mediated by down-regulation of the photochemical machinery and the increase in the synthesis of antioxidant enzymes, while long-term responses are mediated primarily by an increase in the induction of soluble phlorotannins [257].
Antarctic seaweeds—in particular endemic brown algae—reveal a tremendous bathymetric range (e.g., Desmarestiales, such as Himantothallus grandifolius, Desmarestia anceps and Desmarestia menziesii, and some fucoid species, such as Ascoseira mirabilis and Cystosphaera jacquinotii extend from 2 to 40 m), indicating a remarkable photobiological adaptation [258,259]. Furthermore, brown macroalgae are able to allocate large amounts of phlorotannins in selected parts of their thalli [247,248]. This strategical storage in algal fronds can change over short periods of time, with highest contents during low tides and higher UV stress in summer [260,261,262]. In fact, the synthesis and accumulation of phlorotannins and their antioxidant capacity were found to follow a diurnal course in several brown algae [261].
Several studies on Phaeophyceae, and also other common rhodophytes (e.g., Iridaea cordata, Trematocarpus antarcticus and Palmaria decipiens) reveal that such acclimatory flexibility to depth and seasonal patterns for light demands and UV tolerance appears to be strongly related to the high phlorotannin contents and antioxidant potential [262,263,264]. These functional traits provide benefits not only at an individual scale, but also explain both the stability and resilience capacity of the whole benthic community that depends on these organisms [264]. In relation to this, Iken et al. [265,266] found that nine abundant Antarctic brown algae contained higher levels of phlorotannins than most tropical and North Pacific species, but were comparable to levels in Australasian species. Rautenberger et al. [262] suggested that UV tolerance in macroalgae, which are sensitive to UV-R, is modulated by temperature, producing species-specific effects. But not all types of phlorotannins behave in the same way. Soluble phlorotannins exhibit a more dynamic short- and mid-term responses towards UV exposures, as observed in the kelps Lessonia nigrescens, Lessonia spicata and Macrocystis pyrifera and the fucoid Durvillaea antarctica [267,268,269,270]; whereas the insoluble content has a more stable pattern, displaying little differences upon variations of irradiance, depth or season [267,268,269]. Indeed, algae tend to anatomically allocate these two phlorotannins fractions accordingly, e.g., soluble phlorotannins tend to be more concentrated in reproductive and active structures than in vegetative tissues, where the insoluble phlorotannins are predominant [268,269].
Regarding their precise chemical structure, in contrast to hydrolysable and condensed tannins, which are easily analysed with separation methods, such as HPLC and capillary electrophoresis, phlorotannins are still commonly analysed as the total amount of the whole compound group by colorimetric methods using phloroglucinol as a standard agent [271]. In fact, current knowledge of their ecology is based almost entirely on the total contents of phlorotannins, since it usually is a very complex mixture, challenging structural elucidation of the individual compunds [272]. Examples of the diversity of phlorotannins in brown algae include fucols (with phenyl linkages) (16), phlorethols (19) and fuhalols (18), (with ether linkages), eckols (with dibenzodioxin linkages) (15), and fucophlorethols (with ether and phenyl linkages) (17). As far as we know, the reported Antarctic phlorotannins have not been identified to individual level so far.
Flavonoids
Flavonoids (from the Latin flavus = yellow) are a type of secondary metabolites found in plants and fungi. They have an absorption spectrum from 280 to 340 nm, providing UV screening, while allowing transmission of PAR for photosynthesis. Flavonoids are synthesized stimulated by UV-R through the phenylpropanoid pathway. These phenolic compounds fulfill many other roles, including resistance to predators and pathogens, pollinator enhancers, and seed dispersal agents [273]. Flavonoids are not reported in Antarctic systems, due to the lack of marine phanerogams.
Tridentatols
Tridentatols A to D are unique phenolic metabolites with an uncommon sulfur-containing functional group isolated from the hydroid Tridentata marginata, which lives commonly associated with the pelagic Sargassum community around the Caribbean. They display a strong absorption in the UV-A and UV-B regions ranging 313–342 nm, and have been hypothesized to function in photoprotection, as well as serving as deterrent agents from predators, thereby performing a dual role [274]. Similar compounds have not been reported in Antarctic organisms so far.
3.2.6. Fluorescent Proteins
Green fluorescent-like proteins, widely distributed amongst symbiotic cnidarians, fluoresce in the presence of UV-R or PAR [275]. Highly resistant to extreme pH and temperature, they were originally isolated and described from the hydromedusae Aequorea victoria [276,277]. Moreover, their SOD-like activity can quench superoxide radicals [278]. To our knowledge, green fluorescent-like proteins have not been identified yet in Antarctic marine organisms.
3.3. Quenching Mechanisms
Once UV-R reaches the inside the cell, it interacts with O2 and other organic compounds to produce harmful ROS, such as superoxide (O2−•), hydroxyl radical (•OH), hydroperoxyl radical (HO2−) or hydrogen peroxide (H2O2) provoking oxidative stress. ROS can damage important biomolecules, such as DNA, proteins and lipids [279]. Polyunsaturated fatty acids (PUFAs) are one of the primary targets of ROS by removing a proton from conjugated double bond systems, forming a peroxyl radical that then activates lipid peroxidation chain reactions [280], causing tissue damage [281] and alterations of the integrity of cell membranes [282]. This is of particular relevance for marine organisms living in Antarctica, since they have a high content of PUFAs to improve membrane fluidity at low temperature [283]. Differences in PUFAs and in their relations to other fatty acids were also described between Antarctic and Mediterranean gastropod molluscs [284]. To counteract the oxidative stress by ROS, marine organisms have evolved antioxidant systems based on both non-enzymatic and enzymatic antioxidants. Examples of Antarctic marine antioxidants are reported in Table A3.
3.3.1. Non-Enzymatic Antioxidants
Carotenoids
Carotenoids are structurally and functionally very diverse natural pigments and important components of the photosynthetic apparatus, playing a dual role by: (i) Enhancing cellular photosynthetic production, and (ii) providing photooxidative protection. They are derived from five carbon isoprene units that are polymerized enzymatically to form regular highly conjugated 40-carbon structures with up to 15 conjugated double bonds [285]. They are grouped in two main classes: carotenes, which are hydrocarbons that may go through cyclization to form β-ionone ring end groups, which additionally may be substituted by oxo, hydroxy or epoxy groups at dissimilar positions to form different xanthophylls, oxygenated derivatives of the former carotenes, which constitute the other classe of carotenoids [286].
One of the most characteristic features of carotenoids is their strong coloration, which is a consequence of light absorption, due to the presence of an extensive system of conjugated double bonds, which is crucial for the proper functioning in light absorption in photosynthetic organisms and photoprotection in all living organisms [287]. Nearly all carotenoids absorb light in the 400–500 nm range, generating their typical UV and visible spectra, with three absorption maxima [286]. The best studied carotenoids include α-, β- and γ-carotene (21, 26, 32), lutein (35), zeaxanthin (38), violaxanthin (37), diadinoxanthin (29), diatoxanthin (30), anteraxanthin (22), astaxantin (23), and flavoxanthin (31), shown in Figure 7. Their quenching properties allow dissipate excess energy from UV-B, which would otherwise generate toxic single O2, thus protecting the photosynthetic machinery from irreversible inhibition. Energy dissipation in light-harvesting antenna systems occurs via direct energy transfer from the Chl an excited state to the carotenoid S1 (lowest excited) state. The safe dissipation of excess energy as heat, also observed as a reduction in fluorescence, is a process known as non-photochemical quenching [288].
Under excess light condition, the formation of a pH gradient across the thylakoid membrane activates the xanthophyll cycle, consisting in the reversible de-epoxidation of violaxanthin (37) to zeaxanthin (38) and anteraxanthin (22) [289,290]. The amount of excitation energy dissipated by this process depends on the pool size and on the de-epoxiation state: More epoxides (violaxanthin (37)) means less energy dissipation and less photoprotection. The epoxidation state (epoxidated/(epoxidated + de-epoxidated pigments)) can, therefore, be considered an indicator of xanthophyll cycle activity under excess irradiance conditions [291].
Increases in xanthophyll/Chl a ratio is commonly observed in microalgae and macroalgae and higher plants subjected to high irradiance, including UV-R [291].
Carotenoids with photoprotective activity have been described in the Antarctic red algae Leptosomia simplex [245]. Among Antarctic microalgae, Polarella glacialis was shown to possess a very high xanthophyll/Chl a ratio, together with a high content of UV-absorbing compounds, at the highest PAR acclimation levels, appearing thus well equipped to cope with high irradiance. In addition, low intracellular concentrations of the lipid peroxidation by-product malondialdehyde were observed in this species, possibly indicating that antioxidant mechanisms are able to prevent rapid accumulation of harmful oxy-radicals that could otherwise oxidise cellular membranes [291]. It is worth to mention that UV-B vulnerability is known to be species specific, although may also be affected by a range of environmental growth conditions, including the light history of the cells. For example, under fixed light, MAA synthesis seems the most effective photoprotective mechanism activated by a microalga Eutreptiella sp. from the Southern Ocean, which under variable light conditions was instead successfully protected by the synthesis of photoprotective pigments of the xanthophyll cycle [292].
A study conducted on the Antarctic microalgae Chaetoceros dichaeta, Pyramimonas gelidicola, Phaeocystis antarctica and Polarella glacialis, confirmed species-specific sensitivity to UV-B and CPDs formation, also showing that acclimation to high PAR induced an increased sensitivity of the species Pyramimonas gelidicola to UV-B with a consequent increase of DNA damage [291].
In high irradiance acclimated cells of the marine diatoms Thalassiosira weissflogii and Thalassiosira antarctica, the diadino-diatoxanthin (29–33) pool was increased compared with cells grown under low irradiance [293]. These authors also suggested that light harvesting pigment ratio is a sensitive indicator of excessive irradiance sensitivity, and small species-specific differences in pigment composition affect photo-induced viability loss.
Carotenoids with an absorption peak at 384 nm were found in seven-year old sample of cyanobateria of the genera Nostoc, from coastal lowland adjacent to the Ross Ice Shelf [294]. Survival to UV-stress of Nostoc spp. and other cyanobacterial species common in habitat fully exposed to maximum UV-B during the ozone minimum of early spring, may be due partly to their carotenoid content. Antarctic benthic mats of cyanobacteria contain high concentrations of carotenes and xanthophylls [295,296] with the highest concentrations in the upper surface strata that confers the bright orange or pink coloration of many of these Antarctic communities [108]. Some pigmented bacteria of Antarctic soil samples owe their colors to the presence of carotenoids [297]. A similar phenomenon is described for Antarctic marine bacteria Antarcticimonas flava [298] and Muricauda antarctica [299], which are marine members of the Flavobacteriaceae isolated from Antarctic seawater, and from heterotrophic bacteria isolated in water samples from lakes and supraglacial fluvial system [300]. Other examples include the Antarctic cyanobacteria Anabaena, Nostoc and Phormidium, which contain higher carotenoid content than their corresponding tropical strains [301]. Also, the Antarctic algae Delesseria lancifolia showed a more complex xanthophyll pattern respect to species from other areas, with violaxanthin (37), antheraxanthin (22), and zeaxanthin (38) as major compounds, and without evidence of chlorophylls other than Chl a [302]. All these xanthophylls are derivatives of β-carotene (26), indicating that the alga is unable to perform the α-cyclization of lycopene in the biosynthesis pathway [302].
Aquatic animals contain significant amounts of carotenoids derived from dietary source, primary from algae and as secondary source from other animals, which accumulate the pigments from phytoplankton. More than 100 carotenoids have been isolated from sponges, cnidarians, molluscs, crustaceans, echinoderms, tunicates and fishes [303]. Antarctic krill (Euphausia superba), and other Antarctic zooplankton, accumulate in the head and shell significant amounts of carotenoids, especially astaxanthin (23), deriving from their algal food and use them as antioxidant or photoprotector [217]. Antarctic krill is considered as a new alternative, sustainable source of antioxidants, such as astaxanthin (23), vitamins A and E, and long chain n-3 PUFA [304,305].
The presence of various carotenoids was evaluated in certain species of Antarctic fish as in the Rajidae (Raja georgiana), Muraenolepidae (Muraenolepis microps), Notothenidae (Dissostichus eleginoides, Notothenia gibberifrons, Notothenia rossi-marmorata, Trematomus hansoni) and Channichthyidae (Chaenocephalus aceratus, Champsocephalus gunnari, Pseudochaenichthys georgianus). Carotenoids identified were: β-carotene (26), α-cryptoxanthin (28), canthaxanthin (27), flavoxanthin (31), isozeaxanthin (33), zeaxanthin (38), tunaxanthin (36), lutein-5, 6-epoxide (34), aurochrome (24), aurochrome-like, auroxanthin (25), astaxanthin (23), astaxanthin ester and 4-hydroxy-α-carotene (20). The total carotenoid content of these fishes ranged from 0.066 to 0.122 µg/g fresh weight [306]. Finally, an astaxanthin-protein-calcium carbonate complex was purified from the aragonite skeleton of the coral Errina antarctica [307].
α-Tocopherol
α-tocopherol (vitamin E) (39), as shown in Figure 8, is a non-enzymatic antioxidant. It refers to a group of lipid-soluble compounds that include both tocopherols and tocotrienols, produced exclusively by plants, algae, and some cyanobacteria [308]. The lipid soluble antioxidant α-tocopherol (39) is the most biologically active form of vitamin E, and it is located in the thylakoid membranes of photosynthetic organisms, where counteracts the effects of ROS by removing oxidized substrates or stopping the lipid peroxidation chains initiated by ROS [309]. Vitamin E acts as a peroxyl radical scavenger, disabling the production of damaging free radicals in tissues, by reacting with them to form a tocopheryl radical, which will then be reduced by a hydrogen donor (e.g., vitamin C) and thus return to its reduced state. Interestingly, vitamin E is not only a radical scavenger, but has a variety of other effects, which could also be an explanation for its photoprotective capacity. Initially, vitamin E absorbs UV light with a maximum at 295 nm [310], which is not often considered in the discussion of its photoprotective effect. Moreover, the inhibition of prostaglandin formation by vitamin E, resulting in the inhibition of inflammatory processes, might be associated with another photoprotective activity of this agent from the damaging effects of UV exposure in animals [311].
In the diatom Thalassiosira sp., isolated from natural phytoplankton assemblages from Potter Cove (Antarctic Peninsula) α-tocopherol showed a marked decrease during the exponential growth phase after exposure to solar UV-R, while the initial content of β-carotene did not show significant differences over time [312]. In addition, MAA production increased, suggesting that for this species photoprotection against UV-induced damage is characterized by short-term consumption of α-tocopherol and longer-term synthesis of MAAs. The UV-B damage/repair ratio during long-term exposure involves the combined action of several endogenous factors within the cell, with MAAs synthesis being the most effective factor related to photoprotection.
A higher content of α-tocopherol and β-carotene was also found in the Antarctic bivalve Laternula elliptica with respect to the temperate mud clam Mya arenaria. The higher vitamins content mirrored lipid radical content that was higher in Laternula elliptica. This study showed that lipid peroxidation extent in Antarctic bivalves can also depend on iron availability [313].
Biochemical adaptation of cellular membranes to function at low-temperature implies a corresponding need for enhanced lipid-phase antioxidant protection, and this is demonstrated by the increased need for dietary vitamin E by cold-water teleosts [314]. Antarctic fish, that have a high content of PUFAs in the plasma membrane to improve membrane fluidity at low temperature [283], are at an elevated risk of UV induced oxidative stress. To avoid lipid peroxidation, fishes and also invertebrates use vitamin E obtained from the diet as the most efficient lipid soluble antioxidant [315]. Vitamin E concentrations were five to six times higher in the Antarctic fish species, Pagothenia borchgrevinki and Trematomus bernacchii than in the plasma from two New Zealand temperate water fish species, blue cod (Parapercis colias) and banded wrasse (Notolabrus fucicola) [316].
α-tocopherol was found to co-exist with a new derivative of vitamin E (α-tocomonoenol (40), shown in Figure 9) in Antarctic notothenioid fish Chaenocephalus aceratus, Champsocephalus gunnari and Gobionotothen gibberifrons and extracts of Antarctic krill Euphausia superba, as well as phytoplankton collected from the Antarctic Peninsula, ranging from 2.8 to 22.3% of the total vitamin E composition [317].
α-tocomonoenol bears an unusual methylene unsaturation at the isoprenoid-chain terminus, and it was firstly isolated from salmon eggs [318], being specific of marine organisms. Due to its specific occurrence, this antioxidant compound was named “marine-derived tocopherol” (MDT). MDT has a broader distribution in marine organisms with comparatively higher concentrations occurring in fish inhabiting cold-water environments, suggesting a specific metabolic function in low temperature adaptation [315]. MDT has greater reactivity with peroxyl radicals than α-tocopherol at low rates of radical flux and performed better than α-tocopherol also in preventing peroxidation of high-density, cholesterol-incorporated liposomes.
Ascorbic Acid
Ascorbic acid (41), shown in Figure 10, is a water-soluble six-carbon compound related to glucose. Its biologically active form, vitamin C, functions as a potent reducing and antioxidant agent and as co-enzyme in several metabolic pathways [319]. It is synthesized by all species except for higher-order primates, guinea pigs, and some bat, fish, and bird species that consequently rely on a supply of vitamin C from their diet [320]. Apart from its antioxidant role, vitamin C also functions as cofactor for the enzymatic reduction of violaxanthin (37) to antheraxanthin (22) and zeaxanthin (38) involved in thermal dissipation of excess energy [321], and in the regeneration of vitamin E [316].
In Antarctic species, ascorbate content may significantly decrease after UV-B irradiation, as for the Antarctic Chlorella sp., cells in which a concomintant decrease of the α-tocopherol, β-carotene and total thiols contents was recorded, leading to the onset of oxidative stress state in its cells [322].
Vitamin C was found to be higher in the plasma of the benthic Antarctic fish Trematomus bernacchii than those found in the plasma of the cryopelagic Pagothenia borchgrevinki and of two New Zealand temperate water fish species, Parapercis colias and Notolabrus fucicola [316].
Glutathione (GSH)
GSH (42), shown in Figure 11, is a tripeptide thiol involved in the maintenance of the cellular redox homeostasis [323], protecting thiol groups in various enzymes and with a relevant role in cellular protection by conjugation with toxic compounds or by quenching ROS. It is also involved in α-tocopherol (39) and ascorbate regeneration through the glutathione-ascorbate cycle [324] and it is in turn regenerated via the pentose phosphate pathway GSH [325]. Together with vitamin C is one of the main hydrophilic antioxidants, found in cytosolic, mitochondrial, chloroplastic, and nuclear aqueous compartments [326]. In non-cold-acclimated terrestrial plants, GSH protects from damage at low temperatures, but in cold-acclimated plants it is required in recovery from high temperature-induced damage suggesting a role for repair and redox homeostasis [327].
3.3.2. Enzymatic Antioxidants
The enzymatic antioxidants comprise SOD, CAT, glutathione peroxidase, glutathione-S-transferase and the enzymes involved in the ascorbate-glutathione cycle to detoxify ROS, such as ascorbate peroxidase, monodehydroascorbate reductase, dehydroascorbate reductase and glutathione reductase [328]. SOD scavenges superoxide radicals and converts them to H2O2, which is then converted to water and O2 via a combined CAT-peroxide system [329], (Figure 12). The ascorbate-glutathione cycle is the most important antioxidant cycle, especially in plants. The process starts with the production of the first ROS, the superoxide radical anion (O2−•), which is dismutated to H2O2 by SOD. The H2O2 is then reduced to H2O by ascorbate peroxidase (APX). The ascorbate oxidized by APX is reduced by the reduced form of GSH, which is yet again reduced by glutathione reductase [330], (Figure 12).
The SOD, peroxidase and CAT activities in the Antarctic sea ice microalgae Chlamydomonas sp. ICE-L were seen to be enhanced under UV-B radiation stress [105]. SOD seems to be an important ROS scavenging mechanism also in the Antarctic marine diatom Chaetoceros brevis, because the activity of SOD was rapidly up-regulated (4 h) in response to irradiance transitions [331]. In Antarctic clam Laternula elliptica, antioxidant capacities (CAT and GSH) are higher than those found in the temperate mud clam Mya arenaria whereas the level of lipofuscin, marker of oxidative damage, is lower in the polar than in the temperate clam [332]. To date, the majority of studies on antioxidant enzymes have been done on Antarctic fish [333,334,335,336,337], where additionally, the antioxidant enzyme system plays an important role in the adaptation to low temperature. As O2 solubility is inversely related to temperature, the Southern Ocean is O2 rich, with concentrations approximately 1.6-fold higher than in seawater at 20 °C [11]. From the biological standpoint, the advantage of a higher availability of O2 for oxidative metabolism is counterbalanced by the formation of high levels of ROS that are added to those produced by UV-R. Therefore, Antarctic marine organisms have an effective and intricate network of defence mechanisms that protects them against oxidative stress [338] and references herein. Transcript levels of proteins involved in antioxidant metabolism and multiple gene copies of ROS scavengers, as well as proteins that mediate iron processing—as iron promotes the production of free radicals—are higher in Antarctic fish than in warm bodied counterparts. Among the proteins involved in the response to oxidative stress, hemoproteins play a relevant role in Antarctic marine organisms [339].
3.4. Mechanisms of Repair from Photodamage
When neither physical nor chemical barriers are able to avoid or reduce UV-induced damage, a variety of repair mechanisms and resynthesis of sensitive targets are carried out to restore the cellular components after UV damage.
DNA is the primary lethal target and its damage is wavelength dependent: UV-A causes indirect damage to DNA by radicals (e.g., singlet O2, hydroxyl ions) in intra- or extracellular fluids [340], damaging also proteins and lipids. UV-B and UV-C cause both indirect and direct damage, because of the strong absorption of DNA at wavelengths below 315 nm. In Antarctic environments, the cool temperatures promote a decrease in the effectivity of repairing processes of damaged molecules, and adverse UV impact may be further exacerbated with episodes of ozone depletion [27]. Molecules involved in the mechanisms of repair—of DNA, protein, and others—developed in Antarctic marine organisms against UV-B damage are listed in Table A4.
3.4.1. DNA Repair
The main target of solar UV-B radiation is DNA, whose peak of absorbance is at 260 nm maintaining a good absorption into all UV-B range [3]. When DNA absorbs UV light, structural changes occur in the molecule that, if uncorrected, can interfere with DNA synthesis and RNA transcription, with consequences for the translation of the genetic code. Besides loss of bases and single- or double-strand breakage, the most common UV-B-induced damage is represented by CPDs, pyrimidine 6-4 pyrimidone photoproducts (6-4PPs) and Dewar isomers [341]. CPDs are the most frequent lesions causing a block in DNA replication, impairing the normal function of replication of DNA, the transcription of genes and the synthesis of proteins [118,342,343]. The Antarctic ozone reduction may strongly increase the risk for CPD accumulation in the bulk of bacterio and phytoplankton, with consequences on the reduction of community growth and the loss of biomass from the water column [344]. In the Antarctic Ryder Bay, Buma et al. [344] demonstrated that, during mid-summer (January, beginning of February 1998), high levels of CPD were detected in the plankton with the highest found in the smallest size fraction, consisting of heterotrophic bacteria. Surface levels of CPD exceed 100 CPDs per million nucleotides in the bacterioplankton fraction. At the end of February and beginning of March, DNA damage was homogeneously distributed over the first 10 m, with levels between 20 and 30 CPDs per million nucleotides for the smallest size fraction. In the Antarctic nototheniod fish Notothenia coriiceps (family Nototheniidae; rockcods) and Chaenocephalus aceratus (family Channichthyidae; icefishes) and in the krill Euphausia superba high levels of CPD have been measured during periods of increased UV-B flux [103]. The capacity for DNA repair in Antarctic organisms has been shown to be highest in those species whose early life history stages occupy the water column during periods of ozone depletion (austral spring), and lowest in species whose eggs and larvae are abundant during winter [103]. In some Antarctic macroalgae—Desmarestia menziesii, Ascoseira mirabilis, Desmarestia anceps, Iridaea cordata, Trematocarpus antarcticus, Palmaria decipiens and Himantothallus grandifolius—, sensitive to UV-R at 2 °C, enhanced UV tolerance at 7 °C may be due to a more efficient damage repair of the photosynthetic apparatus rather than to an enhanced UV screening or radical scavenging [262].
Mechanisms of DNA repair represent an evolutionary ancient defence among organisms [345,346,347], indicating a very early requirement in the evolutionary history of life. In cyanobacteria, the unusually large number of DNA copies may give an additional protection against long-term UV-B damage, particularly in cold polar environments where the rates of DNA repair may be significantly decreased by the low temperature [108]. Common repair mechanisms include photoreactivation and excision repair (dark repair), which are the major pathways used to remove UV-induced DNA lesions, and others, such as recombinational repair and dimer bypass [3].
Photoreactivation
Photoreactivation can remove DNA lesions, such as CPDs or 6-4PPs, thanks to the activity of photolyase, flavin-dependent enzyme, that recognizes and binds to the DNA lesion and, using UV-A and blue light energy, splits the dimers and reverses the lesion [118]. Photolyases are ancient enzymes widely distributed in eukaryotes, eubacteria and archaea [348] but missing in placental mammals [349,350]. They contain a chromophore, such as 5,10-methenyltetrahydrofolate [351], or 8-hydroxy-5-deaza-riboflavin [352], able to absorb blue light and transfer the energy [353] to a non-covalently bound flavin adenine dinucleotide (FAD), which subsequently transfers an electron to the dimer, restoring the normal configuration of DNA [354]. Photolyases have been identified in the Antarctic marine bacteria Pseudomonas, Janthinobacterium, Flavobacterium, Hymenobacter and Sphingomonas [3,355], in different species of Antarctic diatoms [96], in the microalga Chlamydomonas sp. ICE-Lis [356], in the bipolar terrestrial moss Sanionia uncinata (Hedw.) Loeske [357], in the notothenioid fish, Notothenia coriiceps and Chaenocephalus aceratus, and in the krill Euphausia superba [103]. In the Antarctic sea urchin Sterechinus neumayeri, photoreactivation is the primary mechanism of removing CPDs, being able to repair all CPDs in less than 24 h [358]. In this organism, photolyase is constitutively expressed in all tissues and also induced in embryos in response to in situ exposure to UV-R, especially in shallower water depths or sea ice-free regions [359].
Dark Repairs
Another mechanism of DNA repair, common in mammalian cells and also present in prokaryotes and eukaryotes, is the nucleotide excision repair (NER). This is a light-independent mechanism, which involves a series of DNA replication enzymes that recognize DNA lesions and replace damaged DNA with new undamaged nucleotides. Endonuclease cuts the DNA strand at the lesion, DNA polymerase resynthesizes the correct sequence using the complementary undamaged strand of DNA, exonuclease cuts the damaged fragment, and ligase closes the new DNA strand. This kind of repair mechanism is present in bacterial isolates from Antarctic waters and different species of Antarctic diatoms [3]. Xeroderma pigmentosum protein C and RAD23 are crucial proteins of NER in metazoans, and are involved in damage recognition and the recruitment of repair factors [360,361]. Recently, the transcriptome of a sea anemone revealed full-length sunlight stress response genes codifying a chromoprotein and a photoprotein as gene families in cnidarians, while CPD photolyase and UV excision repair RAD23 were present as single copy genes [362].
In bacteria, but not in eukaryotic cells [363], NER together with postreplication recombinational repair and error-prone repair, belong to the dark repair mechanisms light-independent pathways that are triggered by the SOS response, well described in Escherichia coli [364]. Induction of the SOS response involves more than forty independent SOS genes, most of which encode proteins involved in protection, repair, replication, mutagenesis and metabolism of DNA [364]. However, there are two crucial proteins involved, LexA repressor and RecA, that can regulate transcription of cell cycle and DNA repair genes, and correct DNA damage [343]. When DNA damage is little, the genes of the SOS response are transcribed at basal level, due to the binding of the LexA repression protein to SOS promoters [365]. When large amount of DNA damage accumulates within cells inhibiting replication, the RecA protein activates the autocatalysis of the LexA protein, the SOS promoters are activated, and transcription of SOS genes occurs [366]. In Antarctic microbial populations, the timing of induction of genes involved in the SOS response is an important factor in the level of UV-tolerance SOS response [104]. The expression of recA gene and quantification of RecA protein have been used as an indicator of repair in Antarctic marine bacterioplankton communities. Levels of RecA protein vary during the day. It accumulates after sunset when it is still inactive, and it is consumed during the night as it operates [367].
3.4.2. Other Mechanisms of Repair and Defence
In addition to repairing DNA, organisms can replace damaged proteins by de novo synthesis. Photosystem II (PSII) repair cycle and the de novo synthesis of the D1 protein are an important defence mechanism in UV-B-tolerance of Antarctic phytoplankton communities [368], as well as of the Antarctic cyanobacterium Synechocystis salina and the Antarctic green alga Chlorella vulgaris from moss and soil samples, respectively [369]. PSII is a protein-pigment complex involved in the electron transfer during photosynthesis that releases O2 catalyzing the transfer of electrons from water to plastoquinone, and D1 protein, located at its core has a rapid, light-dependent turnover [370]. Under all light intensities, the PSII repair mechanism is continuously active [371], but under UV-R stress, the D1 protein must be rapidly degraded and synthesized de novo via the PSII repair cycle to prevent the accumulation of damaged PSII and maintain the pool of active PSII [372,373]. Apostolova et al. [369] monitored the changes in the photochemistry of PSII of the Antarctic cyanobacterium Synechocystis salina and the Antarctic green alga Chlorella vulgaris and their mesophilic counterparts and demonstrated that the PSII activity of cyanobacteria was more vulnerable to UV-B radiation than that of green algae, whereas the mesophilic strain of Synechocystis salina was more susceptible to UV-B radiation than the Antarctic isolates.
Another mechanism used by the cells to respond to stressful conditions is to synthesize heat shock proteins (Hsps). The expression of Hsps in response to UV is assumed to be part of the endogenous UV response [374]. They are known to function primarily as molecular chaperones to assist protein folding/refolding of non-functional proteins [375], but are now recognized as sensors of cell stress in many systems [376] and as promoters of damaged proteins degradation, contrasting the toxic effects due to exposure to heat, UV-R, and other environmental stressors [377,378]. Hsps increase the resistance of cells to stress by influencing the DNA repair mechanisms or the induction of apoptosis [379]. Small Hsps have protein-protective activity and the capacity to stabilize lipid membranes [380]. UV-B radiation is able to increase the expression level of cytosolic Hsp70 gene in the Antarctic ice algae Chlamydomonas sp. ICE-L [381]. The Antarctic psychrophilic marine ciliate Euplotes focardii, a model organism well adapted to cold, is unable to respond to heat stress with the activation of Hsp70 genes, but under UV stress it is able to induce their expression [382].
When the cellular damage is too high to be repaired, apoptosis can occur to activate a programmed cell death, protecting the whole organism and sacrificing an individual cell [383,384]. Apoptosis in response to UV damage has been reported for several marine organisms [385], including various phytoplankton species [386], green algae (e.g., Dunaliella tertiolecta), and dinoflagellates (e.g., Peridinium gatunense) [387]. UV-induced oxidative stress is also known to play a role in apoptosis in sea urchin embryos activating genes, such as p53 and p21 [101].
4. Conclusions and Future Perspectives
In response to UV-R several photoprotective, quenching and repair mechanisms have evolved and persisted across many taxonomic groups as discussed in the above sections (and previous reviews, e.g., References [118,343]). Actually, the need of evasion from UV-R is one of the reasons for suggesting an origin of life in submarine vents, benthic crevices, and in deep environments [51]. The environment in future scenarios is pronosticated to continue a trend of global warming and ocean acidification, which will result in faster degradation of DOM, in particular CDOM. This will potentially enhance the penetration of UV-R, and in particular UV-B into the water column, emphasizing the need of marine ecosystems to retain photoprotective traits [388].
Antarctic species use the same mechanisms for protection and repair from UV as species thriving at lower latitudes [3]. However, the lack of data on species-specific responses to increased UV-B represents a limitation to the quantitative evaluation of the tolerance mechanisms with respect to other ecosystems. In this sense, the ecological impact of environmental harsh polar conditions and the ozone depletion in this region may favour towards the selection of more efficient biomolecular pathways for the production of photoprotectants. All in all, the photobiology of marine organisms from the Southern Ocean is still poorly explored and characterized by old specialized literature, even if the communities here are a huge reservoir of biodiversity. To date, there has been very little research on biomes from Antarctica for the discovery of new photoprotection products, due to the limited accessibility to those ecosystems. Anti-UV molecules as those found, up to date, in Antarctic organisms are found in other latitudes, and have been commercialized. The discovery of new sources of natural sunscreens is highly timely for cosmeceuticals, as many sunscreen bioactives used by humans, such as the widely used octocrylene, imply risks for marine ecosystems. Octocrylene, an ester formed by the reaction of 3,3-diphenylcyanoacrylate with 2-ethylhexanol, has been already found in the liver of dolphins off the coast of Brazil [389], suggesting a potential risk also for humans.
MAAs are the best understood and the most common photoprotective molecules found in marine organisms. Detection of MAAs in fossils supports their protection function against the harmful effects of UV-R in the early geological eras [390]. Thanks to their multiple roles, MAAs are attractive molecules, showing a promising application in the pharmaceutical and cosmetic industries as natural sunscreens, activators of cell proliferation, anti-cancer agents, anti-photoaging molecules, and stimulators of skin renewal [391]. Indeed, several candidates have been tested for sunscreen suitability for potential use in human skin-care and cosmetic products [159,392,393,394,395,396,397,398]. Among these, porphyra-334 from the red algae Porphyra umbilicalis associated to shinorine has been commercialized as Helioguard® 365 [394,397,398] with protective properties against UV-A-induced loss of cell viability and DNA damage. Helionori® is another product containing as active ingredients the MAAs sunscreens, palythine, porphyria-334 and shinorine, extracted from Porphyra umbilicalis, which protects against UV-A, preserving membrane lipids of keratinocytes and fibroblasts, as well as protecting DNA [399].
The potential application of MAAs with photoprotective and antioxidant activities has been listed in a large number of patents [391]. Besides the application as suncare and cosmetic products, MAAs have been used in the manufacture of several non-biological materials, such as photostabilizing additives in plastics, paints and varnishes [134].
Besides MAAs, also carotenoids have applications in the healthcare and nutraceuticals industry [400]. The global carotenoid market value was $1.5 billion in 2014 and it is expected to increase to nearly $1.8 billion in 2019, with a compound annual growth rate of 3.9% [401]. Microalgae are a valuable source of carotenoids [402] and of some of the most innovative skincare products today. Some examples include Dermochlorella® from CODIF Recherche et Nature (Brittany, France), an extract from green microalgae Chlorella vulgaris containing oligopeptides that increases firmness and skin tone [403]; Alguronic acid from Algenist (San Francisco, CA, USA), mix of polysaccharides produced by microalgae with anti-ageing properties; Alguard from FRUTAROM, a natural sulphated polysaccharide from red microalgae Porphyridium sp. protecting against skin ageing and photodamage [404]. Analogous sunscreen molecules and photoprotective mechanisms have never been tested from Antarctic species, even if polar sources could potentially yield richer concentrations, or exhibit diverse reaction kinetics [266].
There are only few examples of Antarctic photoprotective molecules that have been already used in biotechnological applications, such as carotenoids isolated from UV-resistant Antarctic bacteria used to develop green solar cells called photosensitizers in Dye Sensitized Solar Cells [297]. Another example is represented by the SeaCode® launched by LIPOTEC, an EPS with a market impact characterized by a mixture of extracellular glycoproteins and other glucidic exopolymers produced by biotechnological fermentation of a Pseudoalteromonas sp. isolated in Antarctic waters [405]. This mixture improves the synthesis of collagen I, contributing to the amelioration of skin structural properties [123]. Besides SeaCode®, LIPOTEC has developed another anti-aging product named Antarcticine® obtained from extracts of marine Pseudoalteromonas antarctica [406].
Currently, there is a need for extensive work to understand the photobiology of Antarctic communities and to investigate on the potential of photoprotective compounds and related molecular and enzymatic machineries from those marine habitats to be used in biotechnological discovery pipelines and pharmaceutical applications. Further development of marine biotechnology for human photoprotection and research must be specially focused on the analysis, biosynthesis, and mode of action of several unknown photoprotective compounds against several abnormalities induced by UV-R. All these studies should be always carried out respecting the environment, and protecting biodiversity and chemical diversity for future generations.
Author Contributions
All authors gave an important contribution in the consideration of the available bibliographic information for the review and in the preparation of the final version of the manuscript. L.N-P. and D.G. developed the original idea and organized all the materials.
Funding
This study grant support from the Italian National Programme for Antarctic Research (PNRA) (2016/AZ1.06-Project PNRA16_00043). Grant support was also provided by the Spanish Government through the BLUEBIO project (CTM2016-78901/ANT) to C.A. This study was carried out in the framework of the SCAR Programmes “Antarctic Thresholds–Ecosystem Resilience and Adaptation” (AnT-ERA) and “State of the Antarctic Ecosystem” (AntEco).
Acknowledgments
D.G. wishes to thank Valentina Brasiello and Francesca Segreti for support and assistance.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ACC | Antarctic Circumpolar Current |
| ACCE | Antarctic Climate Change and the Environment |
| APF | Antarctic Polar Front |
| APX | Ascorbate peroxidase |
| CAT | Catalase |
| CDOM | Coloured dissolved organic matter |
| CFCs | Chlorofluorocarbons |
| Chl | Chlorophyll |
| CPDs | Cyclobutane pyrimidine dimers |
| DOM | Dissolved organic matter |
| EPS | Extracellular polysaccharides |
| GHG | Greenhouse gases |
| GSH | Glutathione |
| GSH-Px | Glutathione peroxidase |
| HPLC | High pressure liquid chromatography |
| MAAs | Mycosporine-like amino acids |
| MDT | Marine-derived tocopherol |
| my | Million of years |
| mya | Million of years ago |
| NASA | National Aeronautics and Space Administration |
| NER | Nucleotide excision repair |
| NOAA | National Oceanic and Atmospheric Administration |
| ODSs | Ozone depleting substances |
| PAR | Photosynthetically active radiation |
| 6-4PPs | Pyrimidine 6-4 pyrimidone photoproducts |
| PUFA | Polyunsaturated fatty acids |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| UNEP | United Nations Environment Programme |
| UV, UV-R | Ultraviolet, UV radiation |
| WMO | World Meteorological Organization |
Appendix A

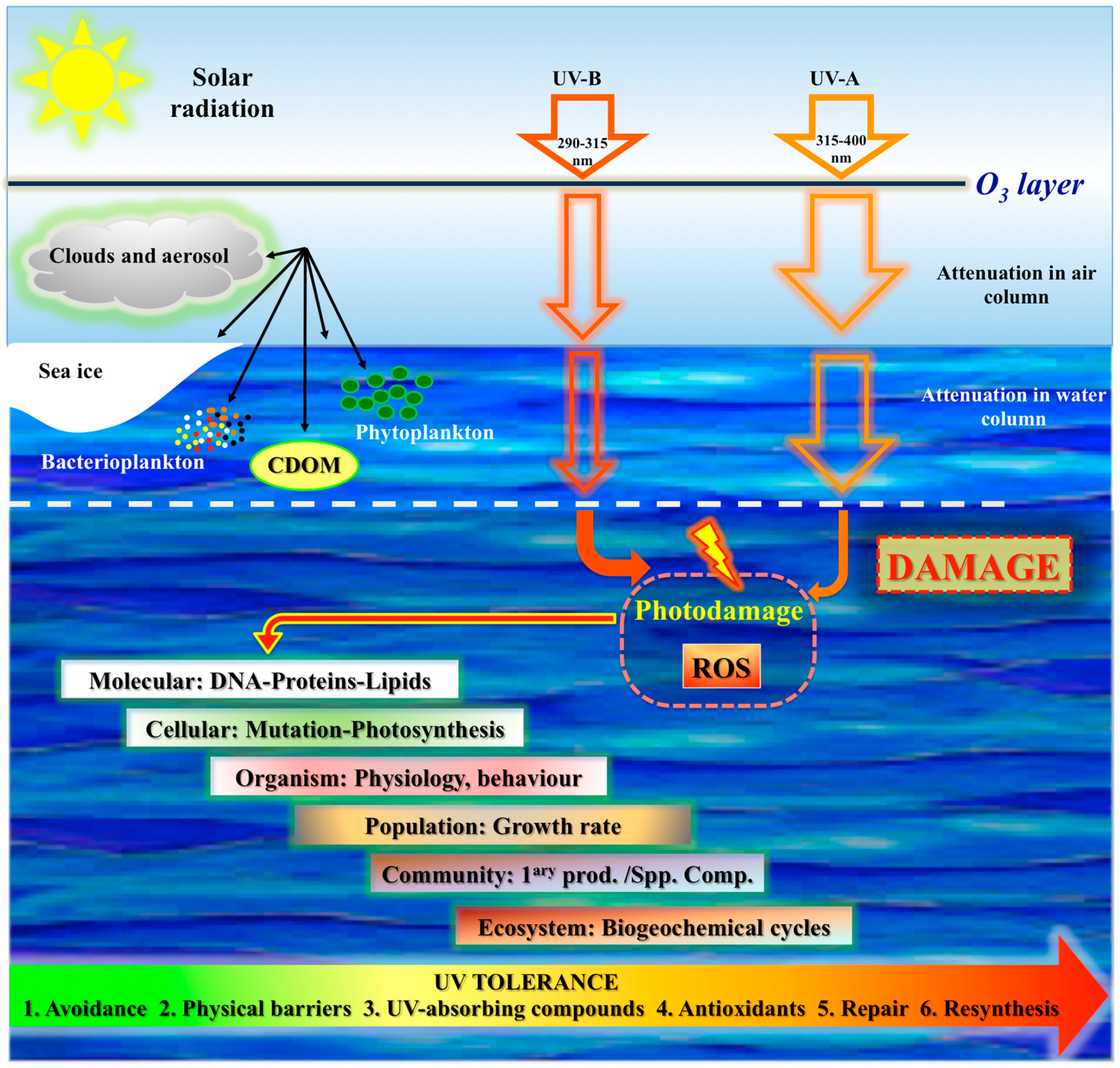{kind=link}
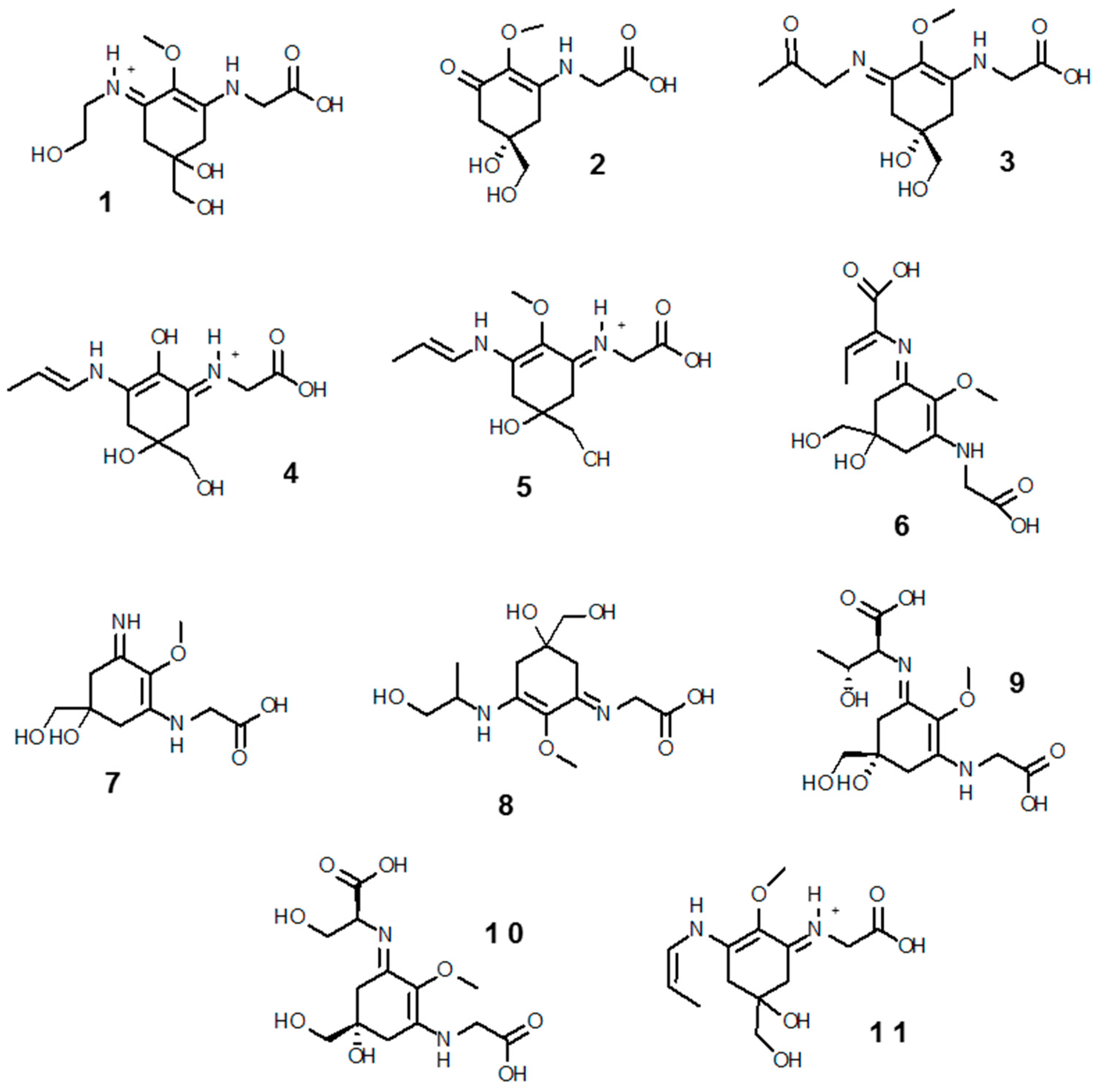{kind=link}
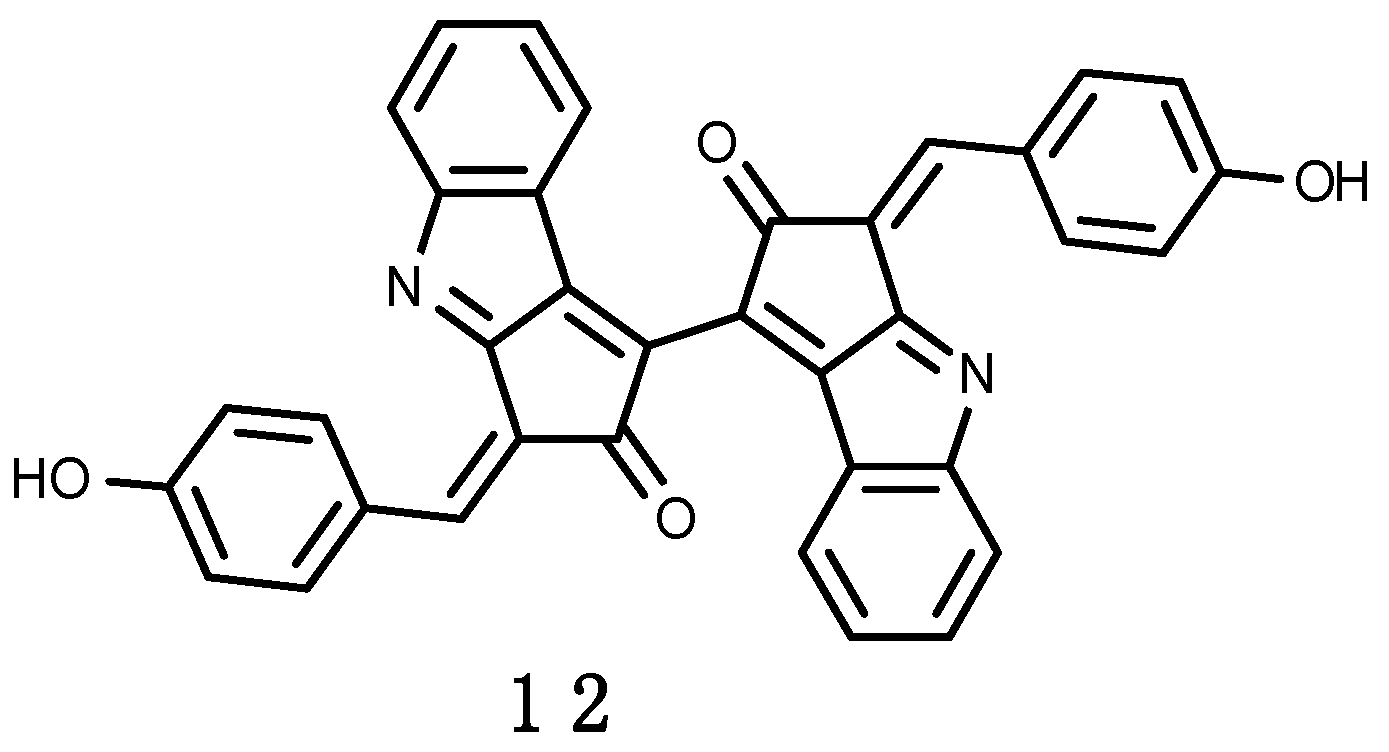{kind=link}
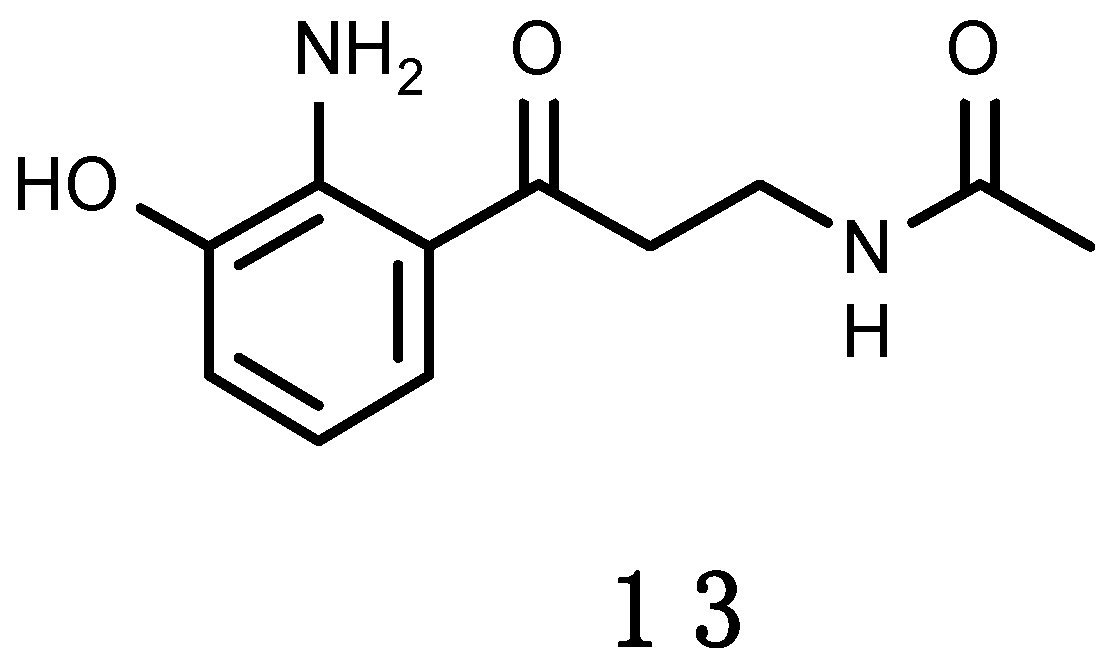{kind=link}
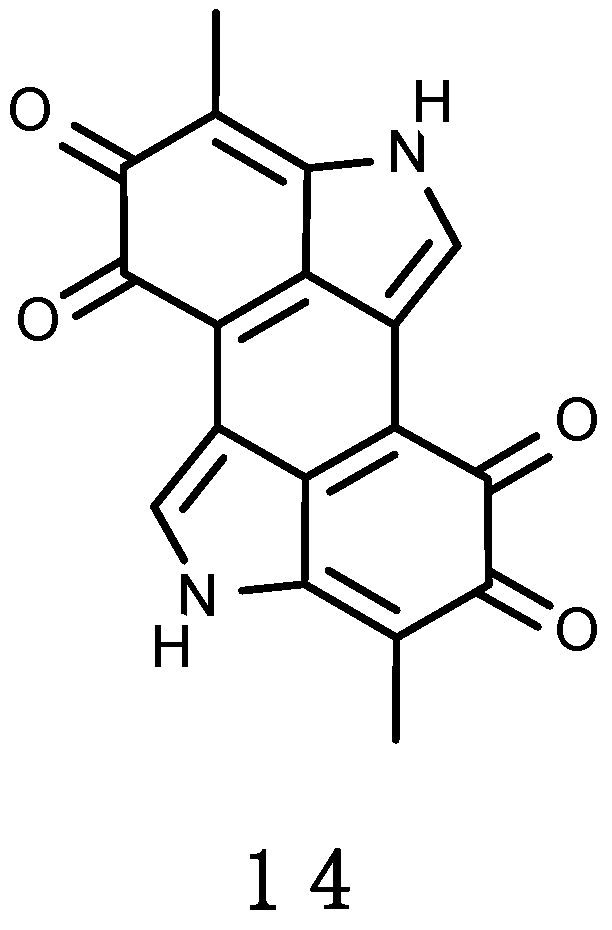{kind=link}
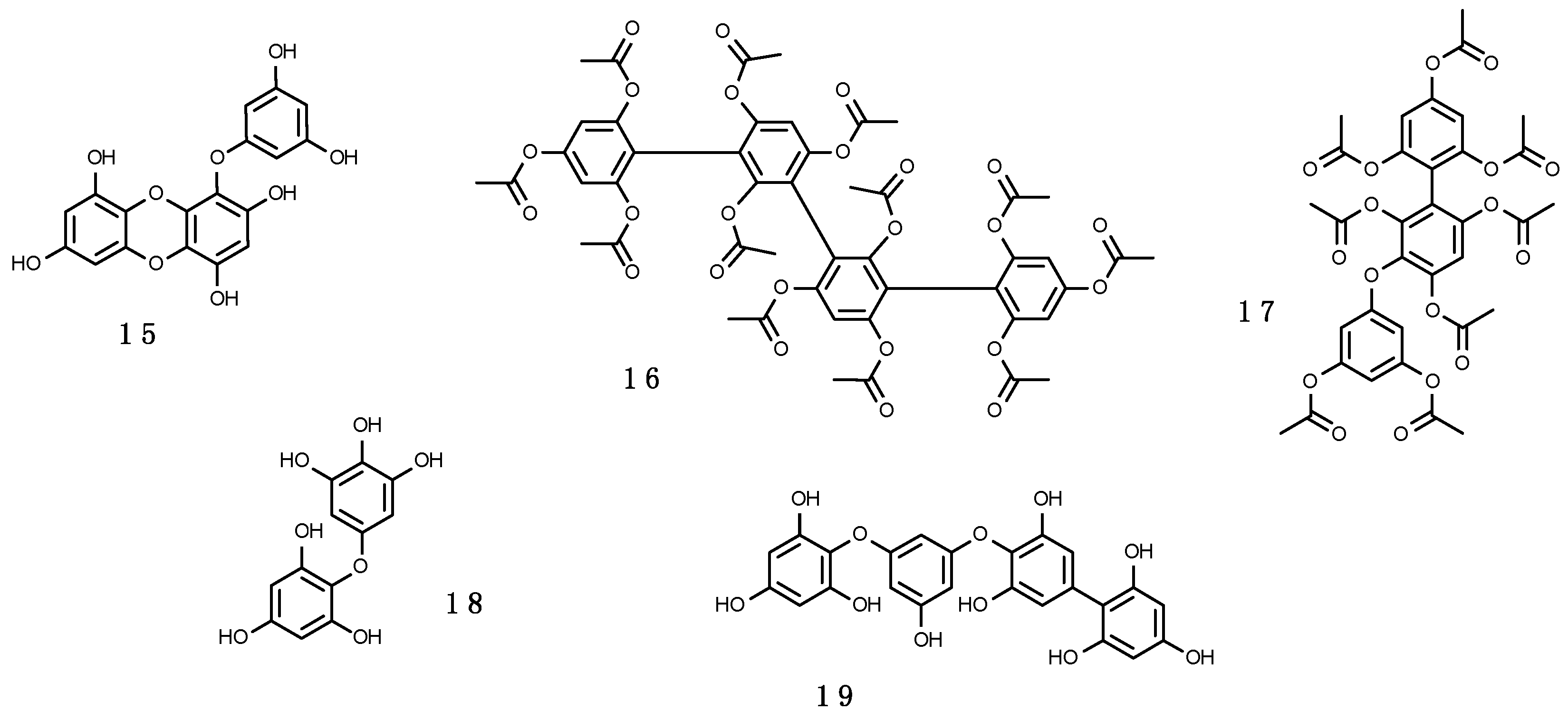{kind=link}
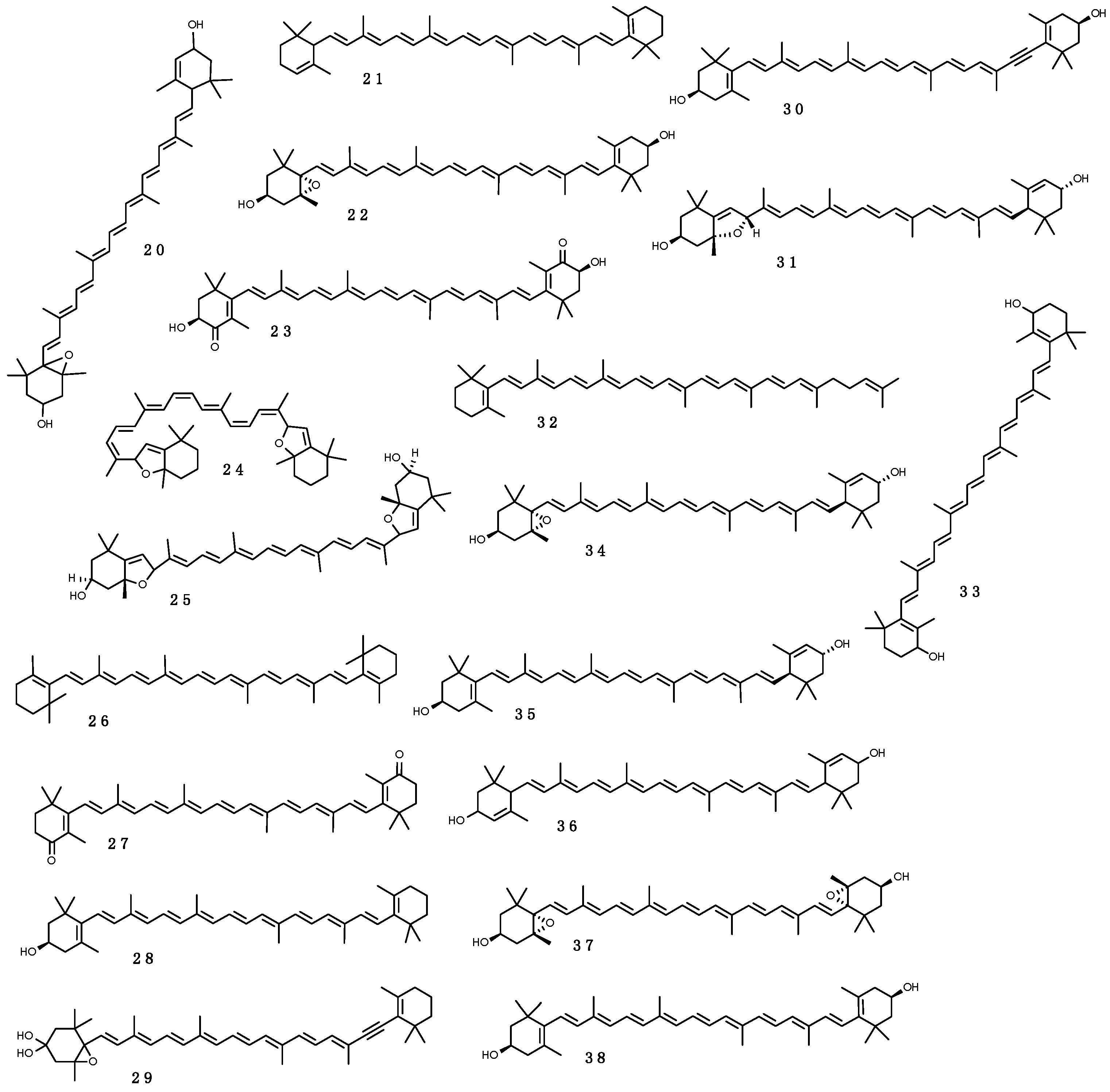{kind=link}
{kind=link}
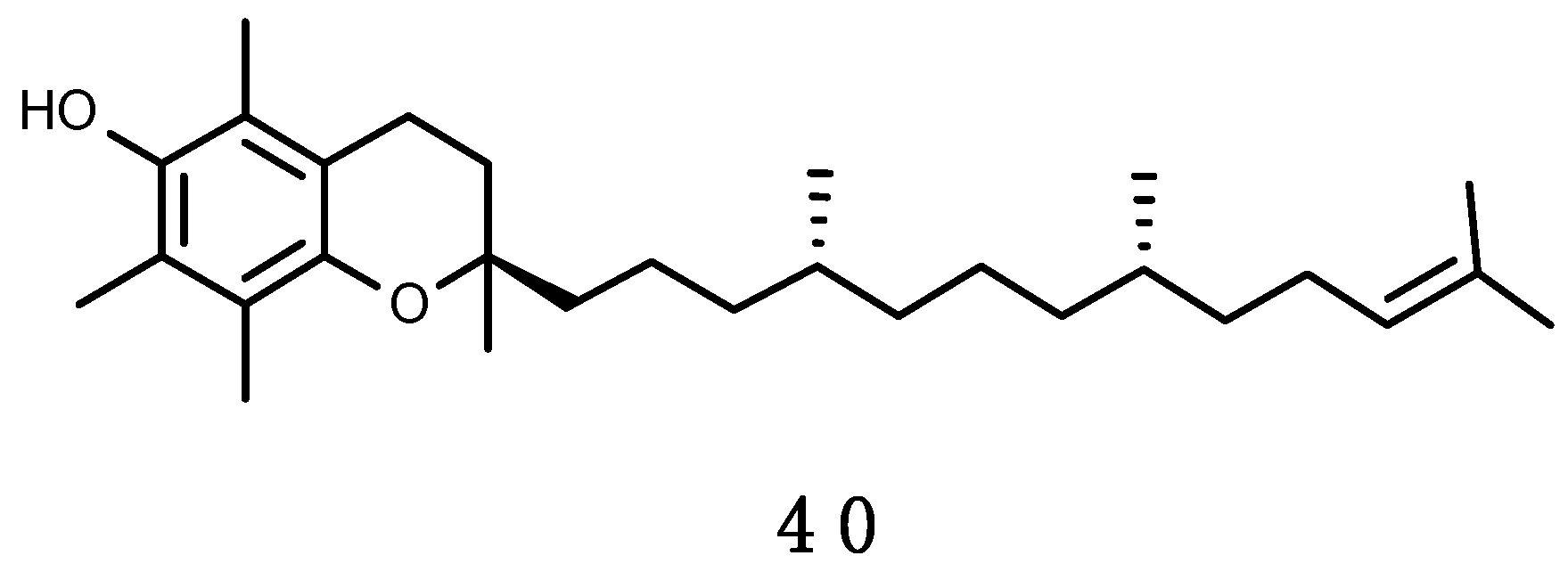{kind=link}
{kind=link}
{kind=link}
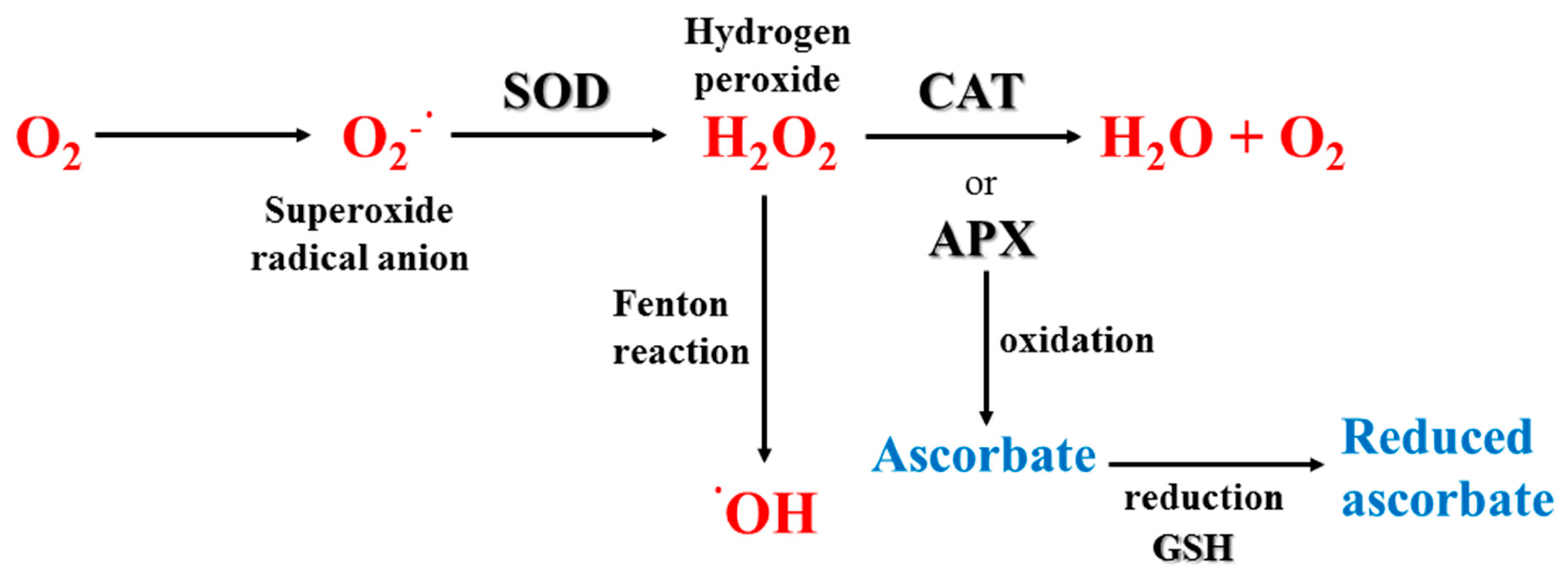{kind=link}
Table A1.
MAAs, UV-absorbing molecules from Antarctic marine organisms.
| Taxonomy | Compound | Reference |
|---|---|---|
| Phylum: Heterokontophyta | ||
| Class: Bacillariophyceae | ||
| Chaetoceros sp. 1 | Porphyra-334, Shinorine | [152] |
| Chaetoceros sp. 2 | Porphyra-334, Shinorine | [152] |
| Coretron cryophilum | Porphyra-334, Shinorine | [213] |
| Coscinodiscus centralis | Porphyra-334, Shinorine | [152] |
| Fragilariopsis cylindrus | Porphyra-334, Shinorine | [152,213] |
| Fragilariopsis linearis | Porphyra-334, Shinorine | [152] |
| Porosira glacialis | Porphyra-334, Shinorine | [152] |
| Porosira pseudodenticulata | Porphyra-334, Shinorine | [152] |
| Proboscia inermis | Porphyra-334, Shinorine | [152] |
| Pseudonitzschia sp. | Porphyra-334, Shinorine | [213] |
| Stellarima microtrias | Porphyra-334, Shinorine | [152] |
| Thalassiosira antarctica | Porphyra-334, Shinorine | [152] |
| Thalassiosira tumida | Porphyra-334, Shinorine | [152] |
| Thalassiosira sp. | Porphyra-334, Shinorine | [213] |
| Diatom mat (mixture of Achnantes sp., Licmophora sp., Navicula sp.) | Porphyra-334, Shinorine, Palythine | [84] |
| Phylum: Chlorophyta | ||
| Class: Chlorophyceae | ||
| Enteromorpha bulbosa | Porphyra-334 | [84] |
| Monostroma hariotii | Shinorine | [220] |
| Green algal mat (mixture of Ulothrix cf. australis, Urospora cf. penicilliformis) | Porphyra-334, Shinorine, Palythine, Asterina-330 | [84] |
| Phylum: Ochrophyta | ||
| Class: Phaeophyceae | ||
| Desmarestia menziesii | Porphyra-334 | [84] |
| Himantothallus grandifolius | Porphyra-334 | [220] |
| Phylum: Rhodophyta | ||
| Class: Rhodophyceae | ||
| Bangia atropurpurea | Porphyra-334 | [220] |
| Curdiea racovitzae | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Palythene, Palythinol | [84,220] |
| Georgiella confluens | Porphyra-334, Shinorine, Palythine | [220] |
| Gigartina skottsbergii | Porphyra-334, Shinorine, Palythine, Asterina-330 | [220] |
| Gymnogongrus antarctica | Shinorine, Palythine, Asterina-330 | [220,221] |
| Gymnogongrus turquetii | Porphyra-334, Shinorine, Mycosporine-glycine | [221,222] |
| Iridaea cordata | Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Palythene, Palythinol | [84,219,220] |
| Kallymenia antarctica | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Palythinol | [221,222] |
| Lithothamnion cf. antarcticum | Porphyra-334, Shinorine | [84] |
| Myriogramme manginii | Porphyra-334, Shinorine, Palythine, Asterina-330 | [220] |
| Neuroglossum ligulatum | Porphyra-334, Shinorine, Palythine | [220,221,222] |
| Notophycus fimbriatus | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine | [220] |
| Pachymenia orbicularis | Porphyra-334, Palythine, Mycosporine-glycine | [220] |
| Palmaria decipiens | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Palythene, Palythinol, Usujirene | [222] |
| Phyllophora antarctica | Shinorine, Palythine | [219] |
| Phyllophora appendiculata | Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Palythene | [84] |
| Plocamium cartilagineum | Porphyra-334, Shinorine, Palythine, Asterina-330, Palythinol | [220,221] |
| Porphyra endiviifolium | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Palythinol | [222] |
| Porphyra plocamiestris | Porphyra-334, Shinorine | [221] |
| Rhodymenia subantarctica | Porphyra-334 | [220] |
| Sarcothalia papillosa | Shinorine, Palythine | [220] |
| Phylum: Porifera | ||
| Class: Demospongia | ||
| Latrunculia (Latrunculia) apicalis | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine-valine | [219] |
| Cinachyra antarctica | Porphyra-334, Shinorine, Palythine | [219] |
| Tetilla leptoderma | Porphyra-334, Shinorine, Palythine | [219] |
| Haliclona (Gellius) benedeni | Shinorine, Palythine, Mycosporine-glycine-valine | [219] |
| Haliclona sp. | Palythine, Mycosporine-glycine | [219] |
| Homaxinella balfourensis | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine-valine | [219] |
| Inflatella belli | Palythine | [219] |
| Isodictya erinacea | Palythine, Palythinol | [219] |
| Kirkpatrickia variolosa | Shinorine, Palythine, Mycosporine-glycine-valine | [219] |
| Mycale (Oxymycale) acerata | Palythine | [219] |
| Sphaerotylus antarcticus | Porphyra-334 | [219] |
| Dendrilla membranosa | Porphyra-334, Shinorine, Palythine | [219] |
| Polymastia invaginata | Porphyra-334, Shinorine, Palythine | [219] |
| Unidentified Sponge #1 | Porphyra-334, Shinorine, Palythine, Palythene, Palythinol, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Unidentified Sponge #3 | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine | [84] |
| Unidentified Sponge #5 | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Unidentified Sponge #6 | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Class: Hexactinellida | ||
| Rossella nuda | Palythine | [219] |
| Rossella racovitzae | Palythine, Mycosporine-glycine | [219] |
| Phylum: Cnidaria | ||
| Class: Anthozoa | ||
| Isotealia antarctica | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Mycosporine-glycine-valine | [219] |
| Unidentified Cnidarian #1 | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine-valine | [219] |
| Phylum: Platyhelminthes | ||
| Class: Rhabditophora | ||
| Obrimoposthia wandeli | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine-valine | [84] |
| Plagiostomum n. sp. | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine-valine | [84] |
| Phylum: Nemertea | ||
| Class: Anopla | ||
| Parborlasia corrugatus | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine-valine | [84] |
| Parborlasia fueguina | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine-valine | [84] |
| Amphiporus michaelseni | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine-valine | [84] |
| Phylum: Mollusca | ||
| Class: Polyplacophora | ||
| Tonicina zschaui | Porphyra-334, Shinorine, Palythine | [84] |
| Class: Gastropoda | ||
| Margarella antarctica | Porphyra-334, Shinorine, Palythine, Asterina-330 | [84] |
| Nacella concinna | Porphyra-334, Shinorine | [407] |
| Marseniopsis mollis | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine | [219] |
| Paludestrina antarctica | Porphyra-334, Shinorine, Palythine, Asterina-330, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Trophon cf. geversianus | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Palythene, Palythinol | [84] |
| Limacina helicina | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Palythenic acid | [135] |
| Limacina helicina ssp. antarctica | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Clione antarctica | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Palythenic acid | [135] |
| Telarma antarctica | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Notaeolidia gigas | Mycosporine-glycine | [219] |
| Tritoniella belli | Shinorine | [219] |
| Mycosporine-glycine | [219] | |
| Class: Bivalvia | ||
| Limatula hodgsoni | Shinorine, Palythine | [219] |
| Limatula cf. ovalis | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Cyamium cf. commune | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Phylum: Anellida | ||
| Class: Polychaeta | ||
| Aglaophamus ornatus | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine | [84] |
| Neanthes kerguelensis | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Tomopteris carpenteri | Porphyra-334, Palythine, Mycosporine-glycine | [84] |
| Terebella ehlersi | Porphyra-334, Shinorine, Palythine, Palythene, Asterina-330, Palythinol, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Unidentified Polychaete #2 | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Class: Clitellata | ||
| Trachelobdella autralis | Porphyra-334, Shinorine, Palythine, Asterina-330, Palythene, Palythinol, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Phylum: Artropoda | ||
| Class: Crustacea | ||
| Calanus propinquus | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine | [84] |
| Euphasia superba | Porphyra-334, Shinorine, Palythine, Asterina-330, Palythene, Palythinol, Mycosporine-glycine, Mycosporine-glycine:valine, Palythenic acid | [84] |
| Cymodocella tubicauda | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Notasellus sarsii | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Glyptonotus antarcticus | Palythine | [219] |
| Bovallia gigantea | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Halirages sp. | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Jassa sp. | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Orchomene sp. | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Paraceradocus sp. | Porphyra-334, Shinorine, Palythine, Asterina-330, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Pariphimedia integricauda | Porphyra-334, Shinorine, Palythine, Asterina-330, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine, Palythinol | [84] |
| Pontogeneia sp. | Porphyra-334, Shinorine, Palythine, Asterina-330, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine, Palythinol | [84] |
| Class: Pycnogonida | ||
| Achelia spicata | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Unidentified Pycnogonid | Shinorine, Palythine | [219] |
| Phylum: Bryozoa | ||
| Class: Gymnolaemata | ||
| Beania livingstonei | Porphyra-334, Shinorine | [84] |
| Inversiula nutrix | Porphyra-334, Shinorine, Palythine, Palythene, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Phylum: Echinodermata | ||
| Class: Crinoidea | ||
| Promachocrinus kerguelensis | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine | [219] |
| Class: Asteroidea | ||
| Granaster nutrix | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Odontaster validus | Palythine | [219] |
| Mycosporine-glycine | [219] | |
| Class: Echinoidea | ||
| Sterechinus neumayeri | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine | [191,219] |
| Class: Holothuroidea | ||
| Cucumaria ferrari | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine | [219] |
| Ekmocucumis steineni | Porphyra-334, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine | [84] |
| Amphioplus affinis | Palythine | [84] |
| Phylum: Chordata | ||
| Class: Ascidiacea | ||
| Cnemidocarpa verrucosa | Porphyra-334, Shinorine, Palythine | [219] |
| Molgula enodis | Porphyra-334, Shinorine, Palythine, Asterina-330, Mycosporine-glycine, Mycosporine-glycine:valine, Palythene | [84] |
| Unidentified ascidia | Shinorine, Mycosporine-glycine | [219] |
| Class: Actinopterygii | ||
| Unidentified ice-fish larvae | Porphyra-334, Shinorine, Palythine, Mycosporine-glycine, Mycosporine-glycine:valine, Palythene | [84] |
| Trematomus bernacchii | Shinorine, palythine | [219] |
Table A2.
UV-absorbing molecules, that are not MAAs, from Antarctic marine organisms.
| Taxonomy | Compound | Reference |
|---|---|---|
| Phylum: Porifera | ||
| Class: Demospongia | ||
| Isodictya erinacea | Erebusinone | [228] |
| Phylum: Proteobacteria | ||
| Class: Gammaproteobacteria | ||
| Alteromonas stellipolaris sp. nov. | Melanin | [241] |
| Phylum: Rhodophyta | ||
| Class: Florideophyceae | ||
| Palmaria decipiens | UV-adsorbing pigment | [245] |
| Phylum: Chlorophyta | ||
| Class: Ulvophyceae | ||
| Enteromorpha bulbosa | UV-adsorbing pigment | [245] |
| Phylum: Ochrophyta | ||
| Class: Phaeophyceae | ||
| Adenocystis utricularis | Phlorotannins | [266] |
| Ascoseira mirabilis | Phlorotannins | [266] |
| Chordaria linearis | Phlorotannins | [266] |
| Cystosphaera jacquinotii | Phlorotannins | [266] |
| Desmarestia anceps | Phlorotannins | [266,408,409] |
| Desmarestia antarctica | Phlorotannins | [266] |
| Desmarestia menziesii | Phlorotannins | [266,408,409] |
| Himantothallus grandifolius | Phlorotannins | [266] |
| Phaeurus antarcticus | Phlorotannins | [266] |
Table A3.
Antioxidant compounds from Antarctic marine organisms.
| Taxonomy | Compound | Reference |
|---|---|---|
| Phylum: Bacteroidetes | ||
| Class: Flavobacteria | ||
| Antarcticimonas flava | Carotenoids | [298] |
| Muricauda antarctica | Carotenoids | [299] |
| Phylum: Proteobacteria | ||
| Class: Gammaproteobacteria | ||
| Pseudoalteromonas haloplanktis TAC125 | Enzymatic antioxidant defence | [338] |
| Phylum: Cyanobacteria | ||
| Class: Hormogoneae | ||
| Nostoc sp. | Carotenoids | [294] |
| Nostoc sp. | Carotenoids | [295] |
| Nostoc commune | Carotenoids | [296] |
| Nostoc sp. | Carotenoids | [301] |
| Class: Cyanophyceae | ||
| Anabaena sp. | Carotenoids | [301] |
| Phormidium sp. | Carotenoids | [301] |
| Oscillatoria sp. | Carotenoids | [295] |
| Phylum: Bacillariophyta | ||
| Class: Mediophyceae | ||
| Thalassiosira antarctica | Diadinoxantin, diatoxanthin | [293] |
| Thalassiosira weissflogii | Diadinoxantin, diatoxanthin | [293] |
| Thalassiosira sp. | α-tocopherol | [312] |
| Chaetoceros brevis | SOD | [331] |
| Phylum: Euglenozoa | ||
| Class: Euglenophyceae | ||
| Eutreptiella sp. | Xanthophylls | [292] |
| Phylum: Miozoa | ||
| Class: Dinophyceae | ||
| Polarella glacialis | Xanthophylls | [291] |
| Phylum: Chlorophyta | ||
| Class: Ulvophyceae | ||
| Enteromorpha bulbosa | UV-absorbing pigments | [245] |
| Class: Trebouxiophyceae | ||
| Chlorella sp. | Ascorbic acid | [322] |
| Class: Chlorophyceae | ||
| Chlamydomonas sp. ICE-L | SOD, CAT, peroxidase | [105] |
| Phylum: Rhodophyta | ||
| Class: Florideophyceae | ||
| Palmaria decipiens | UV-absorbing pigments | [245] |
| Leptosomia simplex | Carotenoids | [410] |
| Delesseria lancifolia | Violaxanthin, antheraxanthin, zeaxanthin | [302] |
| Phylum: Cnidaria | ||
| Class: Hydrozoa | ||
| Errina antarctica | Astaxanthin | [307] |
| Phylum: Mollusca | ||
| Class: Bivalvia | ||
| Laternula elliptica | α-tocopherol | [313] |
| Enzymatic antioxidant defence | [332] | |
| Phylum: Arthropoda | ||
| Class: Malacostraca | ||
| Euphausia superba | Astaxantin | [217] |
| Astaxanthin | [305] | |
| Vitamin A | [305] | |
| Vitamin E | [305] | |
| α-tocomonoenol | [317] | |
| Phylum: Chordata | ||
| Class: Chondrichthyes | ||
| Raja georgiana | Carotenoids | [306] |
| Class: Actinopterygii | ||
| Muraenolepis microps | Carotenoids | [306] |
| Dissostichus eleginoides | Carotenoids | [306] |
| Notothenia gibberifrons | Carotenoids | [306] |
| Notothenia rossi-marmorata | Carotenoids | [306] |
| Trematomus hansoni | Carotenoids | [306] |
| Chaenocephalus aceratus | Carotenoids | [306] |
| Champsocephalus gunnari | Carotenoids | [306] |
| Pseudochaenichthys georgianus | Carotenoids | [306] |
| Pagothenia borchgrevinki | Vitamin E | [316] |
| Trematomus bernacchii | Vitamin E | [316] |
| Champsocephalus gunnari | α-tocomonoenol | [317] |
| Gobionotothen gibberifrons | α-tocomonoenol | [317] |
| Chaenocephalus aceratus | α-tocomonoenol | [317] |
| Antarctic notothenioid fishes | Enzymatic antioxidant defence | [333,334,335,336,337] |
Table A4.
Molecules involved in the mechanisms of repair in Antarctic marine organisms.
| Taxonomy | Compound | Activity | Reference |
|---|---|---|---|
| Phylum: Proteobacteria, Bacteroidetes | |||
| Class: Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, Flavobacteriia | |||
| Pseudomonas, Janthinobacterium, Flavobacterium, Hymenobacter, Sphingomonas | Photolyase | DNA repair | [3,355] |
| Phylum: Ciliophora | |||
| Class: Spirotrichea | |||
| Euplotes focardii | Heat shock protein, Hsp70 | Chaperone in the UV stress | [382] |
| Phylum: Chlorophyta | |||
| Class: Chlorophyceae | |||
| Chlamydomonas sp. ICE-L | Photolyase | DNA repair | [356] |
| Heat shock protein, Hsp70 | Chaperone in the UV stress | [381] | |
| Phylum: Ochrophyta | |||
| Class: Bacillariophyceae | |||
| Chaetoceros neglectus, Corethron cryophilum, Coscinodiscus ocutus-iridis, Odontella weissflogii, Porosira pseudodenticulata, Thalassiosira subtilis, Eucampia antarctica, Chaetoceros convolutus, Chaetocerois socialis, Thalassiosira australis, Licmophora decora, Nitzchia kerguelensis | Photolyase/NER | DNA repair | [96] |
| Phylum: Echinodermata | |||
| Class: Echinoidea | |||
| Sterechinus neumayeri | Photolyase | DNA repair | [359] |
| Phylum: Arthropoda | |||
| Class: Crustacea | |||
| Euphausia superba | Photolyase | DNA repair | [103] |
| Phylum: Chordata | |||
| Class: Actinopterygii | |||
| Chaenocephalus aceratus, Notothenia coriiceps | Photolyase | DNA repair | [103] |
| Organismal assemblages | |||
| Antarctic marine bacterioplankton communities | RecA protein | DNA repair | [367] |
| Antarctic phytoplankton communities | Photosystem II (PSII) repair cycle | de novo synthesis | [368] |
References
- Häder, D.P.; Helbling, E.W.; Williamson, C.E.; Worrest, R.C. Effects of UV radiation on aquatic ecosystems and interactions with climate change. Photochem. Photobiol. Sci. 2011, 10, 242–260. [Google Scholar] [CrossRef] [PubMed]
- Karentz, D.; Bosch, I. Influence of Ozone-Related Increases in Ultraviolet Radiation on Antarctic Marine Organisms. Am. Zool. 2001, 41, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Karentz, D. Ultraviolet tolerance mechanisms in Antarctic marine organisms. Ultrav. Radiat. Antarct. Meas. Boil. Eff. 1994, 62, 93–110. [Google Scholar]
- Eastman, J.T. Antarctic notothenioid fishes as subjects for research in evolutionary biology. Antarct. Sci. 2000, 12, 276–287. [Google Scholar] [CrossRef]
- Rogers, A.D. Evolution and biodiversity of Antarctic organisms: A molecular perspective. Philos. Trans. R. Soc. B 2007, 362, 2191–2214. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T. The nature of the diversity of Antarctic fishes. Pol. Biol. 2005, 28, 94–107. [Google Scholar] [CrossRef]
- Barbante, C.; Fischer, H.; Masson-Delmotte, V.; Waelbroeck, C.; Wolff, E.W. Climate of the last million years: New insights from EPICA and other records. Quat. Sci. Rev. 2010, 29, 1–7. [Google Scholar] [CrossRef]
- Anderson, J.B.; Shipp, S.S.; Lowe, A.L.; Smith Wellner, J.; Mosola, A.B. The Antarctic ice sheet during the Last Glacial Maximum and its subsequent retreat history: A review. Quat. Sci. Rev. 2002, 21, 49–70. [Google Scholar] [CrossRef]
- Sura, P.; Gille, S.T. Stochastic dynamics of sea surface height variability. J. Phys. Oceanogr. 2010, 40, 1582–1596. [Google Scholar] [CrossRef]
- Convey, P.; Bindschadler, R.; di Prisco, G.; Fahrbach, E.; Gutt, J.; Hodgson, D.A.; Mayewski, P.A.; Summerhayes, C.P.; Turner, J.; the ACCE Consortium. Antarctic climate change and the environment. Antarct. Sci. 2009, 21, 541–563. [Google Scholar] [CrossRef] [Green Version]
- Littlepage, J.L. Oceanographic Observations in McMurdo Sound, Antarctica. In Biology of the Antarctic Seas; Llano, G.A., Ed.; American Geophysical Union: Washington, DC, USA, 1965; pp. 1–37. [Google Scholar]
- DeWitt, H.H. The character of the midwater fish fauna of the Ross Sea, Antarctica. In Antarctic Ecology; Holdgate, M.W., Ed.; Academic Press: London, UK, 1970; pp. 305–314. [Google Scholar]
- Hunt, B.M.; Hoefling, K.; Cheng, C.H.C. Annual warming episodes in seawater temperatures in McMurdo Sound in relationship to endogenous ice in notothenioid fish. Antarct. Sci. 2003, 15, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Gordon, A.L. Oceanography of Antarctic waters. Antarct. Res. Ser. 1971, 15, 169–203. [Google Scholar]
- Eastman, J.T. Antarctic Fish Biology: Evolution in a Unique Environment; Academic Press: San Diego, CA, USA, 1993; p. 322. [Google Scholar]
- Thatje, S.; Hillenbrand, C.D.; Mackensen, A.; Larter, R. Life hung by a thread: Endurance of Antarctic fauna in glacial periods. Ecology 2008, 89, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Census of Marine Life (CAML). Available online: http://www.coml.org/census-antarctic-marine-life-caml/ (accessed on 15 May 2018).
- Beers, J.M.; Jayasundara, N. Antarctic notothenioid fish: What are the future consequences of losses and gains acquired during long-term evolution at cold and stable temperatures? J. Exp. Biol. 2015, 218, 1834–1845. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.K.A.; Clarke, A. Antarctic marine biology. Curr. Biol. 2011, 21, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Arntz, W.; Thatje, S.; Gerdes, D.; Gili, J.M.; Gutt, J.; Jacob, U.; Montiel, A.; Orejas, C.; Teixido, N. The Antarctic-Magellan connection: Macrobenthos ecology on the shelf and upper slope, a progress report. Sci. Mar. 2005, 69, 237–269. [Google Scholar] [CrossRef]
- Aronson, R.B.; Thatje, S.; Clarke, A.; Peck, L.S.; Blake, D.B.; Wilga, C.D.; Seibel, B.A. Climate change and invasibility of the antarctic benthos. Ann. Rev. Ecol. Evol. Syst. 2007, 38. [Google Scholar] [CrossRef]
- Peck, L.S.; Convey, P.; Barnes, D.K.A. Environmental constraints on life histories in Antarctic ecosystems: Tempos, timings and predictability. Biol. Rev. 2006, 81, 75–109. [Google Scholar] [CrossRef] [PubMed]
- C.I.E. Standardization of the Terms UV-A1, UV-A2 and UV-B. Commission Internationale de l’Eclairage (International Commission on Illumination) Collection in Photobiology and Photochemistry 134-1999: TC 6-26. 1999. Available online: http://www.cie.co.at/publications/cie-collection-photobiology-photochemistry-1999/ (accessed on 23 August 2018).
- Cordero, R.R.; Damiani, A.; Ferrer, J.; Jorquera, J.; Tobar, M.; Labbe, F.; Carrasco, J.; Laroze, D. UV irradiance and albedo at Union Glacier Camp (Antarctica): A case study. PLoS ONE 2014, 9, e90705. [Google Scholar] [CrossRef] [PubMed]
- Tedetti, M.; Sempéré, R. Penetration of Ultraviolet Radiation in the marine environment. A Review. Photochem. Photobiol. 2006, 82, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Banaszak, A.T.; Lesser, M.P. Effects of solar ultraviolet radiation on coral reef organisms. Photochem. Photobiol. Sci. 2009, 8, 1276–1294. [Google Scholar] [CrossRef] [PubMed]
- Huovinen, P.; Ramírez, J.; Gómez, I.; Ban, S. Underwater Optics in Sub-Antarctic and Antarctic Coastal Ecosystems. PLoS ONE 2016, 11, e0154887. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.C.; Prézelin, B.B.; Baker, K.S.; Bidigare, R.R.; Boucher, N.P.; Coley, T.L.; Karentz, D.; Macintyre, S.; Matlick, H.A.; Menzies, D.; et al. Ozone depletion: Ultraviolet radiation and phytoplankton biology in Antarctic waters. Science 1992, 255, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P.; Lamare, M.D.; Barker, M.F. Transmission of ultraviolet radiation through the Antarctic annual sea ice and its biological effects on sea urchin embryos. Limnol. Oceanogr. 2004, 49, 1957–1963. [Google Scholar] [CrossRef] [Green Version]
- Clark, G.F.; Stark, J.S.; Johnston, E.L.; Runcie, J.W.; Goldsworthy, P.M.; Raymond, B.; Riddle, M.J. Light-driven tipping points in polar ecosystems. Glob. Chang. Biol. 2013, 19, 3749–3761. [Google Scholar] [CrossRef] [PubMed]
- McMinn, A.; Martin, A. Dark Survival in a Warming World. Proc. R. Soc. B Biol. Sci. 2013, 280, 1755. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.K.A.; Tarling, G.A. Polar oceans in a changing climate. Curr. Biol. 2017, 27, R431–R510. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.L. Oceanography-the brawniest retroflection. Nature 2003, 421, 904–905. [Google Scholar] [CrossRef] [PubMed]
- Smale, D.A. Ice disturbance intensity structures benthic communities in nearshore Antarctic waters. Mar. Ecol. Prog. Ser. 2007, 349, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Constable, A.J.; Melbourne-Thomas, J.; Corney, S.P.; Arrigo, K.R.; Barbraud, C.; Barnes, D.K.A.; Bindoff, N.L.; Boyd, P.W.; Brandt, A.; Costa, D.P.; et al. Climate change and Southern Ocean ecosystems I: How changes in physical habitats directly affect marine biota. Glob. Chang. Biol. 2014, 20, 3004–3025. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.F.; Stark, J.S.; Palmer, A.S.; Riddle, M.J.; Johnston, E.L. The Roles of Sea-Ice, Light and Sedimentation in Structuring Shallow Antarctic Benthic Communities. PLoS ONE 2017, 12, e0168391. [Google Scholar] [CrossRef] [PubMed]
- Smetacek, V.; Nicol, S. Polar ocean ecosystems in a changing world. Nature 2005, 437, 362–368. [Google Scholar] [CrossRef] [PubMed]
- McMahon, K.W.; Ambrose, W.G.J.; Johnson, B.J.; Sun, M.; Lopez, G.R.; Clough, L.M.; Carrol, M.L. Benthic community response to ice algae and phytoplankton in Ny Alesund, Svalbard. Mar. Ecol. Prog. Ser. 2006, 310, 1–14. [Google Scholar] [CrossRef]
- Gutt, J.; Isla, E.; Bertler, A.N.; Bodeker, G.E.; Bracegirdle, T.J.; Cavanagh, R.D.; Comiso, J.C.; Convey, P.; Cummings, V.; De Conto, R.; et al. Cross-disciplinarity in the advance of Antarctic ecosystem research. Mar. Gen. 2018. [Google Scholar] [CrossRef] [PubMed]
- Massom, R.A.; Eicken, H.; Hass, C.; Jeffries, M.O.; Drinkwater, M.R.; Sturm, M.; Worby, A.P.; Wu, X.; Lytle, V.I.; Ushio, S.; et al. Snow on Antarctic Sea ice. Rev. Geophys. 2001, 39, 413–445. [Google Scholar] [CrossRef]
- Buckley, R.; Trodahl, H. Scattering and absorption of visible light by sea ice. Nature 1987, 326, 867–869. [Google Scholar] [CrossRef]
- Arndt, S.; Meiners, K.M.; Ricker, R.; Krumpen, T.; Katlein, C.; Nicolaus, M. Influence of snow depth and surface flooding on light transmission through Antarctic pack ice. J. Geophys. Res. Oceans 2017, 122, 2108–2119. [Google Scholar] [CrossRef] [Green Version]
- Brierley, A.S.; Thomas, D.N. Ecology of southern ice pack. Adv. Mar. Biol. 2002, 43, 171–276. [Google Scholar] [PubMed]
- Smith, W., Jr.; Comiso, J.C. The influence of sea ice on primary production in the Southern Ocean: A satellite perspective. J. Geophys. Res. 2008, 113, C05S93. [Google Scholar] [CrossRef]
- Carreto, J.I.; Carignan, M.O. Mycosporine-like amino acids: Relevant secondary metabolites. Chemical and ecological aspects. Mar. Drugs 2011, 9, 387–446. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Bindschadler, R.; Convey, P.; di Prisco, G.; Fahrbach, E.; Gutt, J.; Hodgson, D.; Mayewski, P.; Summerhayes, C. Antarctic Climate Change and the Environment; SCAR & Scott Polar Research Institute: Cambridge, UK, 2009; pp. 1–526. [Google Scholar]
- Holland, M.M.; Landrum, L.; Raphael, M.; Stammerjohn, S. Springtime winds drive Ross Sea ice variability and change in the following autumn. Nat. Commun. 2017, 8, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crampton, J.S.; Cody, R.D.; Levy, R.; Harwood, D.; McKay, R.; Naish, T.R. Southern Ocean phytoplankton turnover in response to stepwise Antarctic cooling over the past 15 million years. Proc. Natl. Acad. Sci. USA 2016, 113, 6868–6873. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Lu, H.; White, I.; King, J.C.; Phillips, T.; Hosking, J.S.; Bracegirdle, T.J.; Marshall, G.J.; Mulvaney, R.; Deb, P. Absence of 21st century warming on Antarctic Peninsula consistent with natural variability. Nature 2016, 535, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, A.; Murphy, E.J.; Meredith, M.P.; King, J.C.; Peck, L.S.; Barnes, D.K.A.; Smith, R.C. Climate change and the marine ecosystem of the western Antarctic Peninsula. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 149–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleaves, H.J.; Miller, S.L. Oceanic protection of prebiotic organic compounds from UV radiation. Proc. Natl. Acad. Sci. USA 1998, 95, 7260–7263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, P.D. Out of Thin Air: Dinosaurs, Birds, and Earth’s Ancient Atmosphere; Joseph Henry Press: Washington, DC, USA, 2006; pp. 1–296. [Google Scholar]
- Scientific Assessment of Ozone Depletion: 2010. WMO Global Ozone Research and Monitoring Project—Report No. 52. World Meteorological Organization: Geneva, Switzerland. Available online: https://www.wmo.int/pages/prog/arep/gaw/ozone_2010/documents/Ozone-Assessment-2010-complete.pdf (accessed on 5 June 2018).
- Molina, M.J.; Rowland, F.S. Stratospheric sink for chlorofluoromethanes: Chlorine atom-catalysed destruction of ozone. Nature 1974, 249, 810–812. [Google Scholar] [CrossRef]
- Manney, G.L.; Santee, M.L.; Rex, M.; Livesey, N.J.; Pitts, M.C.; Veefkind, P.; Nash, E.R.; Wohltmann, I.; Lehmann, R.; Froidevaux, L.; et al. Unprecedented Arctic ozone loss in 2011. Nature 2011, 478, 467–475. [Google Scholar] [CrossRef] [PubMed]
- National Oceanic and Atmospheric Administration NOAA, USA. Available online: http://www.ozonelayer.noaa.gov/data/antarctic.htm (accessed on 18 May 2018).
- Farman, J.; Gardiner, B.; Shanklin, J. Large losses of total ozone in Antarctica reveal seasonal CIO/NO Interaction. Nature 1985, 315, 207–210. [Google Scholar] [CrossRef]
- Robinson, S.A.; Erickson III, D.J. Not just about sunburn—The ozone hole’s profound effect on climate has significant implications for Southern Hemisphere ecosystems. Glob. Chang. Biol. 2015, 21, 515–527. [Google Scholar] [CrossRef] [PubMed]
- National Aeronautics and Space Administration (NASA). Available online: https://www.nasa.gov/content/goddard/2014-antarctic-ozone-hole-holds-steady/ (accessed on 23 April 2018).[Green Version]
- United Nations Environment Programme (UNEP). Available online: http://ozone.unep.org/ (accessed on 23 April 2018). update 2014.
- Velders, G.J.M.; Andersen, S.O.; Daniel, J.S.; Fahey, D.W.; McFarland, M. The importance of the Montreal Protocol in protecting climate. Proc. Natl. Acad. Sci. USA 2007, 104, 4814–4819. [Google Scholar] [CrossRef] [PubMed]
- Estrada, F.; Perron, P.; Martínez-López, B. Statistically derived contributions of diverse human influences to twentieth-century temperature changes. Nat. Geosci. 2013, 6, 1–6. [Google Scholar] [CrossRef]
- Kuttippurath, J.; Nair, P.J. The signs of Antarctic ozone hole recovery. Sci. Rep. 2017, 7, 585. [Google Scholar] [CrossRef] [PubMed]
- Portmann, R.W.; Daniel, J.S.; Ravishankara, A.R. Stratospheric ozone depletion due to nitrous oxide: Influences of other gases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.P.; Williamson, C.E.; Wängberg, S.A.; Rautio, M.; Rose, K.C.; Gao, K.; Helbling, E.W.; Sinhah, R.P.; Worresti, R. Effects of UV radiation on aquatic ecosystems and interactions with other environmental factors. Photochem. Photobiol. Sci. 2015, 14, 108–126. [Google Scholar] [Green Version]
- United Nations Environment Programme (UNEP). Environmental effects of ozone depletion and its interactions with climate change: Progress report 2016. Environmental Effects Assessment Panel. Photochem. Photobiol. Sci. 2017, 16, 107–145. [Google Scholar] [CrossRef] [PubMed]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate warming and disease risks for terrestrial and marine biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef] [PubMed]
- Dubinsky, Z.; Stambler, N. Coral Reefs: An. Ecosystem in Transition; Springer: Dordrecht, The Netherlands, 2011; p. 552. [Google Scholar]
- Marbà, N.; Jordà, G.; Agustí, S.; Girard, C.; Duarte, C.M. Footprints of climate change on Mediterranean Sea biota. Front. Mar. Sci. 2015, 2, 56. [Google Scholar] [CrossRef]
- Meredith, M.; King, J. Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef] [Green Version]
- Peck, L.S. Prospects for surviving climate change in Antarctic aquatic species. Front. Zool. 2005, 2, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, L.S.; Conway, L.Z. The myth of metabolic cold adaptation: Oxygen consumption in stenothermal Antarctic bivalves. In The Evolutionary Biology of the Bivalvia; Harper, E.M., Taylor, J.D., Crame, J.A., Eds.; Special Publication; Geological Society: London, UK, 2000; pp. 441–445. [Google Scholar]
- Peck, L.S.; Morley, S.A.; Richard, J.; Clark, M.S. Acclimation and thermal tolerance in Antarctic marine ectotherms. J. Exp. Biol. 2014, 217, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Barrand, N.E.; Bracegirdle, T.J.; Convey, P.; Hodgson, D.A.; Jarvis, M.; Jenkins, A.; Marshall, G.; Meredith, M.P.; Roscoe, H.; et al. Antarctic climate change and the environment: An update. Polar Rec. 2014, 50, 237–259. [Google Scholar] [CrossRef]
- Oliva, M.; Navarro, F.; Hrbáček, F.; Hernández, A.; Nývlt, D.; Pereira, P.; Ruiz-Fernández, J.; Trigo, R. Recent regional climate cooling on the Antarctic Peninsula and associated impacts on the cryosphere. Sci. Total Environ. 2017, 580, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Bednaršek, N.; Tarling, G.A.; Bakker, D.C.E.; Fielding, Z.S.; Jones, E.M.; Venables, H.J.; Ward, P.; Kuzirian, A.; Lézé, B.; Feely, R.A.; et al. Extensive dissolution of live pteropods in the Southern Ocean. Nat. Geosci. 2012, 5, 881–885. [Google Scholar] [CrossRef] [Green Version]
- Margalef, R. Ecología; Ediciones Omega: Barcelona, Spain, 1998; pp. 1–968. [Google Scholar]
- Diffey, B.L. Solar ultraviolet radiation effects on biological systems. Phys. Med. B 1991, 36, 299–328. [Google Scholar] [CrossRef]
- Leech, D.M.; Johnsen, S. Behavioral responses: UV avoidance and vision. In UV Effects in Aquatic Organisms and Ecosystems; Helbling, H., Zagarese, H., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2003; pp. 455–481. [Google Scholar]
- Valiela, I. Marine Ecological Processes; Springer Advanced Texts in Life Sciences; Springer: New York, NY, USA, 1984; pp. 1–702. [Google Scholar]
- Franklin, L.A.; Osmond, C.B.; Larkum, A.W.D. Photoinhibition, UV-B and algal photosynthesis. In Photosynthesis in algae; Larkum, A.W., Douglas, S.E., Raven, J.A., Eds.; Springer: Kluwer, The Netherlands, 2003; pp. 351–384. [Google Scholar]
- Levinton, J.S. Marine Biology: Function, Biodiversity, Ecology; Oxford University Press: Oxford, UK, 2009; pp. 1–588. [Google Scholar]
- Kingsley, R.J.; Corcoran, M.L.; Krider, K.L.; Kriechbaum, K.L. Thyroxine and vitamin D in the gorgonian Leptogorgia virgulata. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 129, 897–907. [Google Scholar] [CrossRef]
- Karentz, D.; McEue, F.S.; Land, M.C.; Dunlap, W.C. Survey of mycosporine like amino acid compounds in Antarctic marine organisms: Potential protection from ultraviolet exposure. Mar. Biol. 1991, 108, 157–166. [Google Scholar] [CrossRef]
- Leech, D.M.; Johnsen, S. Light, Biological Receptors. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Elsevier: Oxford, UK, 2009; pp. 671–681. [Google Scholar]
- Biscontin, A.; Frigato, E.; Sales, G.; Mazzotta, G.M.; Teschke, M.; De Pittà, C.; Jarman, S.; Meyer, B.; Costa, R.; Bertolucci, C. The opsin repertoire of the Antarctic krill Euphausia superba. Mar. Gen. 2016, 29, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, R.R.; Lemonovich, T.L.; Salata, R.A. An update on the association of vitamin D deficiency with common infectious diseases. Can. J. Physiol. Pharmacol. 2015, 93, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Hume, E.M.; Lucas, N.S.; Smith, H.H. On the Absorption of Vitamin D from the Skin. Biochem. J. 1927, 21, 362–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, W.F.; Neale, P.J. Mechanisms of UV damage to aquatic organisms. In The Effects of UV Radiation in the Marine Environment; de Mora, S., Demers, S., Vernet, M., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 149–176. [Google Scholar]
- Sinha, R.P.; Häder, D.P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, R.M. UV-A (320–380 nm) as an oxidative stress. In Oxidative Stress: Oxidants and Antioxidants; Sies, H., Ed.; Academic Press: San Diego, CA, USA, 1991; pp. 57–83. [Google Scholar]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Bischof, K.; Krábs, G.; Wiencke, C.; Hanelt, D. Solar ultraviolet radiation affects the activity of ribulose-1,5-bisphosphate carboxylase-oxygenase and the composition of photosynthetic and xanthophyll cycle pigments in the intertidal green alga Ulva lactuca L. Planta 2002, 215, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Gómez, I.; Orostegui, M.; Huovinen, P. Morpho-functional patterns of photosynthesis in the South Pacific kelp Lessonia nigrescens: Effects of UV radiation on 14C fixation and primary photochemical reactions. J. Phycol. 2007, 43, 55–64. [Google Scholar] [CrossRef]
- Dring, M.J.; Wagner, A.; Boeskov, J.; Lüning, K. Sensitivity of intertidal and subtidal red algae to UV-A and UV-B radiation, as monitored by chlorophyll fluorescence measurements: Influence of collection depth, and season, and length of irradiance. Eur. J. Phycol. 1996, 31, 293–302. [Google Scholar] [CrossRef]
- Karentz, D.; Cleaver, J.E.; Mitchell, D.L. Cell survival characteristics and molecular responses of Antarctic phytoplankton to Ultraviolet-B radiation. J. Phycol. 1991, 27, 326–341. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Survival and sex ratios of the intertidal copepod, Tigriopus californicus, following ultraviolet-B (290–320 nm) radiation exposure. Mar. Biol. 1995, 123, 799–804. [Google Scholar] [CrossRef]
- Alonso Rodríguez, C.; Browman, H.I.; Runge, J.A.; St-Pierre, J.F. Impact of solar ultraviolet radiation on hatching of a marine copepod, Calanus finmarchicus. Mar. Ecol. Progr. Ser. 2000, 193, 85–93. [Google Scholar] [CrossRef]
- Hader, D.P.; Kumar, H.D.; Smith, R.C.; Worrest, R.C. Aquatic ecosystems: Effects of solar ultraviolet radiation and interactions with other climatic change factors. Photochem. Photobiol. Sci. 2003, 2, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Nahon, S.; Pruski, A.M.; Duchene, J.C.; Mejanelle, L.; Vetion, G.; Desmalades, M.; Charles, F. Can UV radiation affect benthic deposit-feeders through biochemical alteration of food resources? An experimental study with juveniles of the benthic polychaete Eupolymnia nebulosa. Mar. Environ. Res. 2011, 71, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P.; Kruse, V.A.; Barry, T.M. Exposure to ultraviolet radiation causes apoptosis in developing sea urchin embryos. J. Exp. Biol. 2003, 206, 4097–4103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessoa, M.F. Harmful effects of UV radiation in Algae and aquatic macrophytes—A review. Emir. J. Food Agric. 2012, 24, 510–526. [Google Scholar] [CrossRef]
- Malloy, K.D.; Holman, M.A.; Mitchell, D.; Detrich, H.W., III. Solar UV-B-induced DNA damage and photoenzymatic DNA repair in antarctic zooplankton. Proc. Natl. Acad. Sci. USA 1997, 94, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Helbling, E.W.; Marguet, E.R.; Villafañe, V.E.; Holm-Hansen, O. Bacterioplankton viability in Antarctic waters as affected by solar ultraviolet radiation. Mar. Ecol. Prog. Ser. 1995, 126, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Hou, Y.; Miao, J.; Li, G. Effect of UV-B radiation on the growth and antioxidant enzymes of Antarctic sea ice microalgae Chlamydomonas sp. ICE-L. Acta Physiol. Plant. 2009, 31, 1097–1102. [Google Scholar] [CrossRef]
- Zacher, K. The susceptibility of spores and propagules of Antarctic seaweeds to UV and photosynthetically active radiation—Field versus laboratory experiments. J. Exp. Mar. Biol. Ecol. 2014, 458, 57–63. [Google Scholar] [CrossRef]
- Zacher, K.; Roleda, M.Y.; Hanelt, D.; Wiencke, C. UV effects on photosynthesis and DNA in propagules of three Antarctic seaweeds (Adenocystis utricularis, Monostroma hariotii and Porphyra endiviifolium). Planta 2007, 225, 1505–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, W.F.; Quesada, A. Ultraviolet Radiation Effects on Cyanobacteria: Implications for Antarctic Microbial Ecosystems. In Ultraviolet Radiation in Antarctica: Measurements and Biological Effects; Weiler, C.S., Penhale, P.A., Eds.; Washington Geophysical Union: Washington, DC, USA, 1994; pp. 111–124. [Google Scholar]
- Nadeau, T.L.; Howard-Williams, C.; Castenholz, R.W. Effects of solar UV and visible irradiance on photosynthesis and vertical migration of Oscillatoria sp. (Cyanobacteria) in an Antarctic microbial mat. Aquat. Microb. Ecol. 1999, 20, 231–239. [Google Scholar] [CrossRef]
- Van Den Broecke, L.; Martens, K.; Pieri, V.; Schön, I. Ostracod valves as efficient UV protection. J. Limnol. 2012, 71. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, T.A. Adaptations to Physical Stresses in the Intertidal Zone: The Egg Capsules of Neogastropod Molluscs. Am. Zool. 1999, 39, 230–243. [Google Scholar] [CrossRef]
- Wraith, J.; Przeslawski, R.; Davis, A.R. UV-Induced Mortality in Encapsulated Intertidal Embryos: Are Mycosporine-Like Amino Acids an Effective Sunscreen? J. Chem. Ecol. 2006, 32, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Molina, F.J.; Cubillos, V.M.; Montory, J.A.; Andrade-Villagrán, P.A. Are embryonic developing modes determinant in the acquisition and levels of photoprotective compounds in slipper limpets of the Crepipatella genus? J. Photochem. Photobiol. B Biol. 2016, 162, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Hirose, E.; Hirabayashi, S.; Hori, K.; Kasai, F.; Watanabe, M.M. UV protection in the photosymbiotic ascidian Didemnum molle inhabiting different depths. Zool. Sci. 2006, 23, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Hirose, E.; Ohtsuka, K.; Ishikura, M.; Maruyama, T. Ultraviolet absorption in ascidian tunic and ascidian-Prochloron simbiosis. J. Mar. Biol. Assoc. UK 2004, 84, 789–794. [Google Scholar] [CrossRef]
- Marchant, H.J.; Davidson, A.T.; Kelly, G.J. UV-B protecting compounds in the marine alga Phaeocystis pouchetii from Antarctica. Mar. Biol. 1991, 109, 391–395. [Google Scholar] [CrossRef]
- Drollet, J.H.; Glaziou, P.; Martin, P.M.V. A study of mucus from the solitary coral Fungia fungites (Scleractinia: Fungiidae) in relation to photobiological UV adaptation. Mar. Biol. 1993, 115, 263–266. [Google Scholar] [CrossRef]
- Banaszak, A.T. Photoprotective physiological and biochemical responses of aquatic organisms. In UV Effects in Aquatic Organisms and Ecosystems; Helbling, E.W., Zagarese, H., Eds.; Royal Society of Chemistry: Cambridge, UK, 2003; pp. 329–356. [Google Scholar]
- Mandal, S.; Rath, J. Extremophilic cyanobacteria for novel drug development. In Springer Brief in Pharmaceutical Science & Drug Development; Springer: Basel, Switzerland, 2014; pp. 1–89. ISBN 978-3-319-12009-6. [Google Scholar]
- de los Ríos, A.; Ascaso, C.; Wierzchos, J.; Fernández-Valiente, E.; Quesada, A. Microstructural Characterization of Cyanobacterial Mats from the McMurdo Ice Shelf, Antarctica. Appl. Environ. Microbiol. 2004, 70, 569–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancuso Nichols, C.A.; Garon, S.; Bowman, J.P.; Raguénès, G.; Guézennec, J. Production of exopolysaccharides by Antarctic marine bacterial isolates. J. Appl. Microbiol. 2004, 96, 1057–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Extracellular polymeric substances with metal adsorption capacity produced by Pseudoalteromonas sp. MER144 from Antarctic seawater. Environ. Sci. Pollut. Res. Int. 2018, 25, 4667–4677. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Komenda, J.; Kopecky, J.; Nedbal, L. Strategies of ultraviolet-B protection in microscopic algae. Physiol. Plant. 1997, 100, 378–388. [Google Scholar] [CrossRef]
- Rozema, J.; Noordjik, A.J.; Broekman, R.A.; van Beem, A.; Meijkamp, B.M.; de Bakker, N.V.J.; van de Staaij, J.W.M.; Stroetenga, M.; Bohncke, S.J.P.; Konert, M.; et al. (Poly)phenolic compounds in pollen and spores of Antarctic plants as indicators of UV-B: A new proxy for the reconstruction of past solar UV-B? Plant. Ecol. 2001, 154, 11–26. [Google Scholar]
- Banaszak, A.T.; Trench, R.K. Effects of ultraviolet (UV) radiation on marine microalgal-invertebrate symbioses. I.Response, of the algal symbionts in culture and in hospite. J. Exp. Mar. Biol. Ecol. 1995, 194, 213–232. [Google Scholar] [CrossRef]
- Taira, H.; Goes, J.I.; Gomes, H.R.; Yabe, K.; Taguchi, S. Photoinduction of mycosporine-like amino acids and cell volume increases by ultraviolet radiation in the marine dinoflagellate Scrippsiella sweeneyae. Plankton Biol. Ecol. 2004, 51, 82–94. [Google Scholar]
- Dunlap, W.C. Sunscreens, oxidative stress and antioxidant functions in marine organisms of the Great Barrier Reef. Redox Rep. 1999, 4, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Klisch, M.; Gröniger, A.; Häder, D.P. Ultraviolet absorbing/screening substances in cyanobacteria, phytoplankton and macroalgae. J. Photochem. Photobiol. B 1998, 47, 83–94. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C.; Buettner, G.R. Ultraviolet (UV) protection in marine organisms II. Biosynthesis, accumulation, and sunscreening function of mycosporine-like amino acids. In Free Radicals in Chemistry, Biology and Medicine; OICA International: London, UK, 2000; pp. 215–228. [Google Scholar]
- Shick, J.M.; Dunlap, W.C. Mycosporine-like amino acids and related gadusols: Biosynthesis, accumulation, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 2002, 64, 223–262. [Google Scholar] [CrossRef] [PubMed]
- Trione, E.J.; Leach, C.M.; Mutch, J.T. Sporogenic substances isolated from fungi. Nature 1966, 212, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Sakamoto, T.; Matsugo, S. Mycosporine-Like Amino Acids and Their Derivatives as Natural Antioxidants. Antioxidants 2015, 4, 603–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandaranayake, W.M. Mycosporines: Are they nature’s sunscreens? Nat. Prod. Rep. 1998, 15, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, K.; Karentz, D.; Hedges, J.I. Mycosporine-like amino acids (MAAs) in phytoplankton, a herbivorous pteropod (Limacina helicina), and its pteropodpredator (Clione antarctica) in McMurdo Bay, Antarctica. Mar. Biol. 2001, 139, 1013–1019. [Google Scholar]
- Teai, T.; Drollet, J.H.; Bianchini, J.P.; Cambon, A.; Martin, M.V.P. Widespread occurrence of mycosporine-like amino acid compounds in scleractinians from French Polynesia. Coral Reefs 1997, 16, 169–176. [Google Scholar] [CrossRef]
- Bentley, R. The shikimate pathway—A metabolic tree with many branches. Crit. Rev. Biochem. Mol. Biol. 1990, 25, 307–384. [Google Scholar] [CrossRef] [PubMed]
- Portwich, A.; Garcia-Pichel, F. Biosynthetic pathway of mycosporines (mycosporine-like amino acids) in the cyanobacterium Chlorogloeopsis sp. strain PCC 6912. Phycologia 2003, 42, 384–392. [Google Scholar] [CrossRef]
- Starcevic, A.; Dunlap, W.C.; Shick, J.M.; Hranueli, D.; Cullum, J.; Long, P.F. Enzymes of the shikimic acid pathway encoded in the genome of a basal metazoan, Nematostella vectensis, have microbial origins. Proc. Natl. Acad. Sci. USA 2008, 105, 2533–2537. [Google Scholar] [CrossRef] [PubMed]
- Banaszak, A.T.; Barba Santos, M.G.; LaJeunesse, T.C.; Lesser, M.P. The distribution of mycosporine-like amino acids (MAAs) and the phylogenetic identity of symbiotic dinoflagellates in cnidarian hosts from the Mexican Caribbean. J. Exp. Mar. Biol. Ecol. 2006, 337, 131–146. [Google Scholar] [CrossRef]
- Osborn, A.R.; Almabruk, K.H.; Holzwarth, G.; Asamizu, S.; LaDu, J.; Kean, K.M.; Karplus, P.A.; Tanguay, R.L.; Bakalinsky, A.T.; Mahmud, T. De novo synthesis of a sunscreen compound in vertebrates. eLife 2015, 4, e05919. [Google Scholar] [CrossRef] [PubMed]
- García-Pichel, F.; Wingard, C.E.; Castenholz, R.W. Evidence regarding the UV sunscreen role of a mycosporine-like compound in the Cyanobacterium Gloeocapsa sp. Appl. Environ. Microbiol. 1993, 59, 170–176. [Google Scholar] [PubMed]
- Oren, A.; Gunde-Cimerman, N. Mycosporines and mycosporine-like amino acids: UV protectants or multipurpose secondary metabolites? FEMS Microbiol. Lett. 2007, 269, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehling-Schulz, M.; Bilger, W.; Scherer, S. UV-B-induced synthesis of photoprotective pigments and extracellular polysaccharides in the terrestrial cyanobacterium. Nostoc Commun. J. Bacteriol. 1997, 179, 1940–1945. [Google Scholar] [CrossRef]
- Portwich, A.; Garcia-Pichel, F. Ultraviolet and osmotic stresses induce and regulate the synthesis of mycosporines in the cyanobacterium Chlorogloeopsis PCC 6912. Arch. Microbiol. 1999, 172, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Portwich, A.; Garcia-Pichel, F. A novel prokaryotic UV-B photoreceptor in the cyanobacterium Chlorogloeopsis PCC 6912. Photochem. Photobiol. 2000, 71, 493–498. [Google Scholar] [CrossRef]
- Kräbs, G.; Watanabe, M.; Wiencke, C. A Monochromatic Action Spectrum for the Photoinduction of the UV-Absorbing Mycosporine-like Amino Acid Shinorine in the Red Alga Chondrus crispus. Photochem. Photobiol. 2004, 79, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Carreto, J.I.; De Marco, S.G.; Lutz, V.A. UV-absorbing pigments in the dinoflagellates Alexandrium excavatum and Prorocentrum micans. Effects of light intensity. In Red Tides: Biology, Environmental Science, and Toxicology; Okaichi, T., Anderson, D.M., Nemoto, T., Eds.; Elsevier Science: New York, NY, USA, 1989; pp. 333–336. [Google Scholar]
- Banaszak, A.T.; Trench, R.K. Effects of ultraviolet (UV) radiation on marine microalgal-invertebrate symbioses. II. The, synthesis of mycosporine-like amino acids in response to exposure to UV in Anthopleura elegantissima and Cassiopeia xamachana. J. Exp. Mar. Biol. Ecol. 1995, 194, 233–250. [Google Scholar] [CrossRef]
- Gröniger, A.; Hallier, C.; Häder, D.P. Influence of UV radiation and visible light on Porphyra umbilicalis: Photoinhibition and MAA concentration. J. Appl. Phycol. 1999, 11, 437–445. [Google Scholar] [CrossRef]
- Zudaire, L.; Roy, S. Photoprotection and long-term acclimation to UV radiation in the marine diatom Thalassiosira weissflogii. J. Photochem. Photobiol. B Biol. 2001, 62, 26–34. [Google Scholar] [CrossRef]
- Riegger, L.; Robinson, D. Photoinduction of UV-absorbing compounds in Antarctic diatoms and Phaeocystis antarctica. Mar. Ecol. Prog. Ser. 1997, 160, 13–25. [Google Scholar] [CrossRef]
- Hannach, G.; Sigleo, A.C. Photoinduction of UV-absorbing compounds in six species of marine phytoplankton. Mar. Ecol. Prog. Ser. 1998, 174, 207–222. [Google Scholar] [CrossRef] [Green Version]
- Hernando, M.; Carreto, J.I.; Carignan, M.O.; Ferreyra, G.A.; Gross, C. Effects of solar radiation on growth and mycosporine-like amino acids content in Thalassiosira sp., an Antarctic diatom. Polar Biol. 2002, 25, 12–20. [Google Scholar]
- Shick, J.M.; Romaine-Lioud, S.; Ferrier-Pages, C.; Gattuso, J.P. Ultraviolet-B radiation stimulates shikimate pathway-dependent accumulation of mycosporine-like amino acids in the coral Stylophora pistillata despite decreases in its population of symbiotic dinoflagellates. Limnol. Oceanogr. 1999, 44, 1667–1682. [Google Scholar] [CrossRef] [Green Version]
- Lesser, M.P. Depth-dependent photoacclimatization to solar ultraviolet radiation in the Caribbean coral Montastrea faveolata. Mar. Ecol. Prog. Ser. 2000, 192, 137–151. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C.; Chalker, B.E.; Banaszak, A.T.; Rosenzweig, T.K. Survey of ultraviolet radiation-absorbing mycosporinelike amino acids in organs of coral reef holothuroids. Mar. Ecol. Prog. Ser. 1992, 90, 139–148. [Google Scholar] [CrossRef]
- Carroll, A.K.; Shick, J.M. Dietary accumulation of UV-absorbing mycosporine-like amino acids (MAAs) by the green sea urchin (Strongylocentrotus droebachiensis). Mar. Biol. 1996, 124, 561–569. [Google Scholar] [CrossRef]
- Dunlap, W.C.; Shick, J.M. Ultraviolet radiation-absorbing mycosporine-like amino acids in coral reef organisms: A biochemical and environmental perspective. J. Phycol. 1998, 34, 418–430. [Google Scholar] [CrossRef]
- Mason, D.S.; Schafer, F.; Shick, J.M.; Dunlap, W.C. Ultraviolet radiation-absorbing mycosporine-like amino acids (MAAs) are acquired from their diet by medaka fish (Oryzias latipes) but not by SKH-1 hairless mice. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 120, 587–598. [Google Scholar] [CrossRef]
- Shick, J.M. The continuity and intensity of ultraviolet irradiation affect the kinetics of biosynthesis, accumulation, and conversión of mycosporine-like amino acids (MAAs) in the coral Stylophora pistillata. Limnol. Oceanogr. 2004, 49, 442–458. [Google Scholar] [CrossRef]
- Yakovleva, I.; Hidaka, M. Diel fluctuations of mycosporine-likeamino acids in shallow-water scleractinian corals. Mar. Biol. 2004, 145, 863–873. [Google Scholar] [CrossRef]
- Rezanka, T.; Temina, M.; Tolstikov, A.G.; Dembitsky, V.M. Natural Microbial UV Radiation Filters—Mycosporine-like Amino Acids. Folia Microbiol. 2004, 49, 339–352. [Google Scholar] [CrossRef]
- Misonou, T.; Saitoh, J.; Oshiba, S.; Tokitomo, Y.; Maegawa, M.; Inoue, Y.; Hori, H.; Sakurai, T. UV-absorbing substance in the red alga Porphyra yezoensis (Bangiales, Rhodophyta) block thymine photodimer production. Mar. Biotechnol. 2003, 5, 194–200. [Google Scholar]
- Dunlap, W.C.; Chalker, B.E. Identification and quantitation of near-UV absorbing compounds (S-320) in a hermatypic scleractinian. Coral Reefs 1986, 5, 155–159. [Google Scholar] [CrossRef]
- Shick, J.M.; Lesser, M.P.; Stochaj, W.R. Ultraviolet radiation and photooxidative stress in zooxanthellate Anthozoa: The sea anemone Phyllodiscus semoni and the octocoral Clavularia sp. Symbiosis 1991, 10, 145–173. [Google Scholar]
- Shick, J.M.; Lesser, M.P.; Dunlap, W.C.; Stochaj, W.R.; Chalker, B.E.; Wu Won, J. Depth-dependent responses to solar ultraviolet radiation and oxidative stress in the zooxanthellate coral Acropora microphthalma. Mar. Biol. 1995, 122, 41–51. [Google Scholar] [CrossRef]
- Gleason, D.F. Differential effects of ultraviolet radiation on Green and brown morphs of the Caribbean coral Porites astreoides. Limnol. Oceanogr. 1993, 38, 1452–1463. [Google Scholar] [CrossRef]
- Teai, T.; Raharivelomanana, P.; Bianchini, J.P.; Faura, R.; Martín, P.M.V.; Cambon, A. Structure de deux nouvelles iminomycosporines isolées de Pocillopora eydouxy. Tetrahedron Lett. 1997, 38, 5799–5800. [Google Scholar] [CrossRef]
- Banaszak, A.T.; Lesser, M.P.; Kuffner, I.B.; Ondrusek, M. Relationship between ultraviolet (UV) radiation and mycosporinelike amino acids (MAAs) in marine organisms. Bull. Mar. Sci. 1998, 63, 617–628. [Google Scholar]
- Yakovleva, I.; Bhagooli, R.; Takemura, A.; Hidaka, M. Differential susceptibility to oxidative stress of two scleractinian corals: Antioxidant functioning of mycosporine-glycine. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 139, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, W.C.; Yamamoto, Y. Small-molecule antioxidants in marine organisms: Antioxidant activity of mycosporine-glycine. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 112, 105–114. [Google Scholar] [CrossRef]
- Suh, H.J.; Lee, H.W.; Jung, J. Mycosporine glycine protects biological systems against photodynamic damage by quenching singlet oxygen with a high efficiency. Photochem. Photobiol. 2003, 78, 109–113. [Google Scholar] [CrossRef]
- Oren, A. Mycosporine-like amino acids as osmotic solutes in a community of halophilic cyanobacteria. Geomicrobiol. J. 1997, 14, 231–240. [Google Scholar] [CrossRef]
- Neale, P.J.; Banaszak, A.T.; Jarriel, C.R. Ultraviolet sunscreens in Gymnodinium sanguineum (Dynophyceae): Mycosporine-like amino acids protect against inhibition of photosynthesis. J. Phycol. 1998, 938, 928–938. [Google Scholar] [CrossRef]
- Peinado, N.K.; Abdala Díaz, R.T.; Figueroa, F.L.; Helbling, E.W. Ammonium and UV radiation stimulate the accumulation of mycosporine-like amino acids in Porphyra columbina (Rhodophyta) from Patagonia, Argentina. J. Phycol. 2004, 40, 248–259. [Google Scholar] [CrossRef]
- Korbee, N.; Figueroa, F.L.; Aguilera, J. Accumulation of mycosporine-like amino acids (MAAs): Biosynthesis, photocontrol and ecophysiological functions. Rev. Chil. Hist. Nat. 2006, 79, 119–132. [Google Scholar]
- Shick, J.M.; Ferrier-Pagés, C.; Grover, R.; Allemand, D. Effects of starvation, ammonium concentration, and photosynthesis on the UV dependent accumulation of mycosporine-like amino acids (MAAs) in the coral Stylophora pistillata. Mar. Ecol. Prog. Ser. 2005, 295, 135–156. [Google Scholar] [CrossRef]
- Cook, C.B.; Davy, S.K. Are free amino acids responsible for the ‘host factor’ effects on symbiotic zooxanthellae in extracts of host tissue? Hydrobiologia 2001, 461, 71–78. [Google Scholar] [CrossRef]
- Sinha, R.P.; Singh, S.P.; Häder, D.P. Database on mycosporines and mycosporine-like amino acids (MAAs) in fungi, cyanobacteria, macroalgae, phytoplankton and animals. J. Photochem. Photobiol. B Biol. 2007, 66, 47–53. [Google Scholar] [CrossRef]
- Shibata, K. Pigments and a UV-absorbing substance in corals and a blue–green alga living on the Great Barrier Reef. Plant. Cell. Physiol. 1969, 10, 325–335. [Google Scholar]
- Llewellyn, C.A.; Airs, R.L. Distribution and abundance of MAAs in 33 species of microalgae across 13 classes. Mar. Drugs 2010, 8, 1273–1291. [Google Scholar] [CrossRef] [PubMed]
- Bernillon, J.; Bouillant, M.L.; Pittet, J.L.; Favre-Bonvin, J.; Arpin, N. Mycosporine glutamine and related mycosporines in the fungus Pyronema omphalodes. Phytochemistry 1984, 23, 1083–1087. [Google Scholar] [CrossRef]
- Karsten, U.; Sawall, T.; Hanelt, D.; Bischof, K.; Figueroa, F.L.; Flores-Moya, A.; Wiencke, C. An inventory of UV-absorbing mycosporine-like amino acids in macroalgae from polar to warm-temperate regions. Bot. Mar. 1998, 41, 443–453. [Google Scholar] [CrossRef]
- Karentz, D. Chemical defences of marine organisms against solar radiation exposure: UV-absorbing mycosporine-like amino acids and scytonemin. In Marine Chemical Ecology; McClintock, J.B., Baker, B.J., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 481–520. [Google Scholar]
- Dunlap, W.C.; Williams, D.M.; Chalker, B.E.; Banaszak, A.T. Biochemical photoadaptations in vision: UV-absorbing pigments in fish eye tissues. Comp. Biochem. Physiol. 1989, 93, 601–607. [Google Scholar] [CrossRef]
- Arai, T.; Nishijima, M.; Adachi, K.; Sano, H. Isolation and structure of a UV absorbing substance from the marine bacterium Micrococcus sp. AK-334. MBI Rep. 1992, 88–94. [Google Scholar]
- Miyamoto, K.T.; Komatsu, M.; Ikeda, H. Discovery of gene cluster for mycosporine-like amino acid biosynthesis from Actinomycetales microorganisms and production of a novel mycosporine-like amino acid by heterologous expression. J. Appl. Environ. Microbiol. 2014, 80, 5028–5036. [Google Scholar] [CrossRef] [PubMed]
- Corinaldesi, C.; Barone, G.; Marcellini, F.; Dell’Anno, A.; Danovaro, R. Marine microbial-derived molecules and their potential use in cosmeceutical and cosmetic products. Mar. Drugs 2017, 15, 118. [Google Scholar] [CrossRef] [PubMed]
- Michalek-Wagner, K. Seasonal and sex-specific variations in levels of photo-protecting mycosporine-like amino acids (MAAs) in soft corals. Mar. Biol. 2001, 139, 651–660. [Google Scholar]
- Karentz, D.; Dunlap, W.C.; Bosch, I. Temporal and spatial occurrence of UV-absorbing mycosporine-like amino acids in tissues of the Antarctic sea urchin Sterechinus neumayeri during springtime ozone-depletion. Mar. Biol. 1997, 129, 343–353. [Google Scholar] [CrossRef]
- Corredor, J.E.; Bruckner, A.W.; Muszynski, F.Z.; Armstrong, R.A.; Garcia, R.; Morell, J.M. UV-absorbing compounds in three species of Caribbean zooxanthellate corals: Depth distribution and spectral response. Bull. Mar. Sci. 2000, 67, 821–830. [Google Scholar]
- Muszynski, F.Z.; Bruckner, A.; Armstrong, R.A.; Morell, J.M.; Corredor, J.E. Within-colony variations of UV absorption in a reef building coral. Bull. Mar. Sci. 1998, 63, 589–594. [Google Scholar]
- Vernet, M.; Whitehead, K. Release of ultraviolet-absorbing compounds by the red-tide dinoflagellate Lingulodinium polyedra. Mar. Biol. 1996, 127, 35–44. [Google Scholar] [CrossRef]
- Cubillos, V.M.; Burritt, D.J.; Lamare, M.D.; Peake, B.M. The relationship between UV-irradiance, photoprotective compounds and DNA damage in two intertidal invertebrates with contrasting mobility characteristics. J. Photochem. Photobiol. B Biol. 2015, 149, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Avila, C.; Taboada, S.; Núñez-Pons, L. Antarctic marine chemical ecology: What is next? Mar. Ecol. 2008, 29, 1–71. [Google Scholar] [CrossRef]
- Teai, T.; Drollet, J.H.; Bianchini, J.P.; Cambon, A.; Martin, P.M.V. Occurrence of ultraviolet radiation-absorbing mycosporinelike amino acids in coralmucus and whole corals of French Polynesia. Mar. Freshw. Res. 1998, 49, 127–132. [Google Scholar] [CrossRef]
- Yakovleva, I.M.; Baird, A.H. Ontogenetic change in the abundance of mycosporine-like amino acids in non-zooxanthellate coral larvae. Coral Reefs 2005, 24, 443–452. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Ishikura, M.; Maruyama, T.; Miyachi, S. UV absorbing substances in the tunic of a colonial ascidian protect its symbiont, Prochloron sp. from damage by UV-B radiation. Mar. Biol. 1997, 128, 455–461. [Google Scholar] [CrossRef]
- Karsten, U.; Sawall, T.; Wiencke, C. A survey of the distribution of UV-absorbing substances in tropical macroalgae. Phycol. Res. 1998, 46, 271–279. [Google Scholar] [CrossRef]
- Adams, N.L.; Shick, J.M. Mycosporine-like amino acids prevent UVB-induced abnormalities during early development of the green sea urchin Strongylocentrotus droebachiensis. Mar. Biol. 2001, 138, 267–280. [Google Scholar] [CrossRef]
- Franklin, L.A.; Yakovleva, I.; Karsten, U.; Lüning, K. Synthesis of mycosporine-like amino acids in Chondrus crispus (Florideophyceae) and the consequences for sensitivity to ultraviolet B radiation. J. Phycol. 1999, 35, 682–693. [Google Scholar] [CrossRef]
- Michalek-Wagner, K.; Willis, B.L. Impacts of bleaching on the soft coral Lobophytum compactum. II. Biochemical, changes in adults and their eggs. Coral Reefs 2001, 19, 240–246. [Google Scholar] [CrossRef]
- Adams, N.L.; Shick, J.M. Mycosporine-like amino acids provide protection against ultraviolet radiation in eggs of the sea urchin Strongylocentrotus droebachiensis. Photochem. Photobiol. 1996, 64, 149–158. [Google Scholar] [CrossRef]
- Ishikura, M.; Kato, C.; Maruyama, T. UV-absorbing substances in zooxanthellate and azooxanthellate clams. Mar. Biol. 1997, 128, 649–655. [Google Scholar] [CrossRef]
- Maruyama, T.; Hirose, E.; Ishikura, M. Ultraviolet-light-absorbing tunic cells in didemnid ascidians hosting a symbiotic photo-oxygenic prokaryote. Prochloron. Biol. Bull. 2003, 204, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Drollet, J.H.; Teai, T.; Faucon, M.; Martin, P.M.V. Field study of compensatory changes in UV-absorbing compounds in the mucus of the solitary coral Fungia repanda (Scleractinia: Fungiidae) in relation to solar UV radiation, sea-water temperature, and other coincident physico-chemical parameters. Mar. Freshw. Res. 1997, 48, 329–333. [Google Scholar] [CrossRef]
- Bidigare, R. Potential effects of UV-B Radiation on marine organisms of the Southern Ocean: Distributions of phytoplankton and krill during spring. Photochem. Photobiol. 1989, 50, 469–477. [Google Scholar] [CrossRef]
- Ferreyra, G.A.; Schloss, I.; Demers, S.; Neale, P.J. Phytoplankton responses to natural ultraviolet irradiance during early spring in the Weddell-Scotia Confluence: An experimental approach. Antarct. J. U. S. 1994, 29, 268–270. [Google Scholar]
- Davidson, A.T.; Marchanht, H.J. The impact of ultraviolet radiation on Phaeocystis and selected species of Antarctic marine diatoms. Antarc. Res. Ser. 1994, 62, 187–205. [Google Scholar]
- Vernet, M.; Brody, E.A.; Holm-Hansen, O.; Mitchell, B.G. The response of Antarctic phytoplankton to ultraviolet radiation: Absorption, photosynthesis, and taxonomic composition. Antarct. Res. Ser. 1994, 62, 143–158. [Google Scholar]
- Helbling, E.W.; Villafan, V.; Holm-Hansen, O. Effects of ultraviolet radiation on Antarctic marine phytoplankton photosynthesis with particular attention to the influences of mixing. Antarc. Res. Ser. 1994, 62, 207–227. [Google Scholar]
- Helbling, E.W.; Chalker, B.E.; Dunlap, W.C.; Holm-Hansen, O.; Villafañe, V.E. Photoacclimation of antarctic marine diatoms to solar ultraviolet radiation. J. Exp. Mar. Biol. Ecol. 1996, 204, 85–101. [Google Scholar] [CrossRef]
- Bidigare, R.R.; Iriarte, J.L.; Kangs, S.H.; Ondrusemk, M.E.; Karentdz, D.; Fryxell, G.A. Phytoplankton: Quantitative and qualitative assessments. Antarct. Res. Ser. 1996, 70, 173–198. [Google Scholar]
- Frame, E.R. Mycosporine-Like Amino Acids (MAAs) in Bloom Forming Phytoplankton: The Influence of Nitrogen, Ultraviolet Radiation and Species Composition. Ph.D. Thesis, University of California, San Diego, CA, USA, 2004. [Google Scholar]
- Ryan, K.G.; McMinn, A.; Mitchell, K.A.; Trenerry, L. Mycosporine-like amino acids inAntarctic Sea ice algae, and their response to UV-B radiation. Z. Fuer Naturforschung C J. Biosci. 2002, 57, 471–477. [Google Scholar] [CrossRef]
- Newman, S.J.; Dunlap, W.C.; Nicol, S.; Ritz, D. Antarctic krill (Euphausia superba) acquire a UV-absorbing mycosporine-like amino acid from dietary algae. J. Exp. Mar. Biol. Ecol. 2000, 255, 93–110. [Google Scholar] [CrossRef]
- Nakamura, H.; Kobayashi, J.; Hirata, Y. Separation of mycosporine-like amino acids in marine organisms using reversed-phase high-performance liquid chromatography. J. Chromatogr. 1982, 250, 113–118. [Google Scholar] [CrossRef]
- McClintock, J.; Karentz, D. Mycosporine-like amino acids in 38 species of subtidal marine organisms from McMurdo Sound, Antarctica. Antarc. Sci. 1997, 4, 392–398. [Google Scholar] [CrossRef]
- Hoyer, K.; Karsten, U.; Sawall, T.; Wiencke, C. Photoprotective substances in Antarctic macroalgae and their variation with respect to depth distribution, different tissues and developmental stages. Mar. Ecol. Prog. Ser. 2001, 211, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Hoyer, K.; Karsten, U.; Wiencke, C. Induction of sunscreen compounds in Antarctic macroalgae by different radiation conditions. Mar. Biol. 2002, 141, 619–627. [Google Scholar]
- Hoyer, K.; Karsten, U.; Wiencke, C. Inventory of UV-absorving mycosporine-like amino acids in polar macroalgae and factors controlling their content. In Antarctic Biology in a Global Context; Huiskes, A.H.L., Gieskes, W.W.C., Rozema, J., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 56–62. [Google Scholar]
- Helbling, E.W.; Barbieri, E.S.; Sinha, R.J.; Villafañe, V.E.; Häder, D.P. Dynamics of potentially protective compounds in Rhodophyta species from Patagonia (Argentina) exposed to solar radiation. J. Photochem. Photobiol. B 2004, 75, 63–71. [Google Scholar] [CrossRef] [PubMed]
- García-Pichel, F.; Castenholz, R.W. Characterization and biological implications of scytonemin, and cyanobacterial sheath pigment. J. Phycol. 1991, 27, 395–409. [Google Scholar] [CrossRef]
- Sinha, R.P.; Häder, D.P. UV-protectants in cyanobacteria. Plant Sci. 2008, 174, 278–289. [Google Scholar] [CrossRef]
- Dillon, J.G.; Tatsumi, C.M.; Tandingan, P.G.; Castenholz, R.W. Effect of environmental factors on the synthesis of scytonemin, a UV-screening pigment, in a cyanobacterium (Chroococcidiopsis sp.). Arch. Microbiol. 2002, 177, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.Y.; Cowie, R.O.; Simpson, A.M.; O’Toole, R.; Ryan, K.G. The origin of cyanobacteria in Antarctic sea ice: Marine or freshwater? Environ. Microbiol. Rep. 2012, 4, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Moon, B.; Park, Y.C.; McClintock, J.B.; Bakera, B.J. Structure and bioactivity of erebusinone, a pigment from the antarctic sponge Isodictya erinacea. Tetrahedron 2000, 56, 9057–9062. [Google Scholar] [CrossRef]
- Truscott, R.J.W.; Carver, J.A.; Thorpe, A.; Douglas, R.H. Identification of 3-Hydroxykynurenine as the lens pigment in the Gourami Trichogaster trichopterus. Exp. Eye Res. 1992, 54, 1015–1017. [Google Scholar] [CrossRef]
- Thorpe, A.; Douglas, R.H.; Truscott, R.J.W. Spectral transmission and shortwave absorbing pigments in the fish lens—I. Phylogenetic distribution and identity. Vis. Res. 1993, 33, 289–300. [Google Scholar] [CrossRef]
- Bandaranayake, W.M. The nature and role of pigments of marine invertebrates. Nat. Prod. Rep. 2006, 23, 223–255. [Google Scholar] [CrossRef] [PubMed]
- Meredith, P.; Riesz, J. Radiative relaxation quantum yields for synthetic eumelanin. Photochem. Photobiol. 2004, 79, 211–216. [Google Scholar] [CrossRef]
- Tartarotti, B.; Laurion, I.; Sommaruga, R. Large variability in the concentration of mycosporine-like amino acids among zooplankton from lakes located across an altitude gradient. Limnol. Oceanogr. 2001, 46, 1546–1552. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, F.E.; Setlow, R.B. Ultraviolet radiation-induced DNAdamage and its photorepair in the skin of the platyfish Xiphophorus. Cancer Res. 1993, 53, 2249–2255. [Google Scholar] [PubMed]
- Hamilton, A.J.; Gomez, B.L. Melanins in fungal pathogens. J. Med. Microbiol. 2002, 51, 189–191. [Google Scholar] [PubMed] [Green Version]
- Castelvecchi, D. Dark Power: Pigment seems to put radiation to good use. Sci. News 2007, 171, 325. [Google Scholar] [CrossRef]
- Dadachova, E.; Bryan, R.A.; Huang, X.; Moadel, T.; Andrew, D.; Schweitzer, A.D.; Aisen, P.; Nosanchuk, J.; Casadevall, A. Ionizing radiation changes the electronic properties of melanin and enhances the growth of melanized fungi. PLoS ONE 2007, 2, e457. [Google Scholar] [CrossRef] [PubMed]
- Neville, A.C. Biology of the Arthropod Cuticle; Springer: Berlin, Germany, 1975; pp. 1–450. [Google Scholar]
- Cerenius, L.; Söderhäll, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, R.T.; Messenger, J.B. Cephalopod Behaviour; Cambridge University Press: Cambridge, UK, 1999; pp. 1–365. [Google Scholar]
- Van Trappen, S.; Tan, T.L.; Yang, J.; Mergaert, J.; Swings, J. Alteromonas stellipolaris sp. nov., a novel, budding, prosthecate bacterium from Antarctic seas, and emended description of the genus Alteromonas. Int. J. Syst. Evol. Microbiol. 2004, 54, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Fukuda, W.; Sanada, T.; Imanaka, T. Characterization of water-soluble dark-brown pigment from Antarctic bacterium, Lysobacter oligotrophicus. J. Biosci. Bioeng. 2015, 120, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Tashirev, A.B.; Romanovskaia, V.A.; Rokitko, P.V.; Matveeva, N.A.; Shilin, S.O.; Tashireva, A.A. Synthesis of melanin pigments by Antarctic black yeast. Mikrobiol. Z. 2012, 74, 2–8. [Google Scholar] [PubMed]
- Tashirev, A.B.; Romanovskaia, V.A.; Shilin, S.O.; Chernaia, N.A. Screening of yeast-producers of melanin in the Antarctic terrestrial biotopes. Mikrobiol. Z. 2010, 72, 3–8. [Google Scholar] [PubMed]
- Post, A.; Larkum, A.W. UV-absorbing pigments, photosynthesis and UV exposure in Antarctica: Comparison of terrestrial and marine algae. Aquat. Bot. 1993, 45, 231–243. [Google Scholar] [CrossRef]
- Ragan, A.M.; Glombitza, K.W. Phlorotannins, brown algal polyphenols. Prog. Phycol. Res. 1986, 4, 129–241. [Google Scholar]
- Targett, N.M.; Arnold, T.M. Predicting the effects of brown algal phlorotannins on marine herbivores in tropical and temperate oceans. J. Phycol. 1998, 834, 195–205. [Google Scholar] [CrossRef]
- Pavia, H.; Cervin, G.; Lindgren, A.; Aberg, P. Effects of UV-B radiation and simulated herbivory on phlorotannins in the brown alga Ascophyllum nodosum. Mar. Ecol. Prog. Ser. 1997, 157, 139–146. [Google Scholar] [CrossRef]
- Shibata, T.; Kawaguchi, S.; Hama, Y.; Inagaki, M.; Yamaguchi, K.; Nakamura, T. Local and chemical distribution of phlorotannins in brown algae. J. Appl. Phycol. 2004, 16, 291. [Google Scholar] [CrossRef]
- Swanson, A.K.; Druehl, L.D. Induction, exudation and the UV protective role of kelp phlorotannins. Aquat. Bot. 2002, 73, 241. [Google Scholar] [CrossRef]
- Amsler, C.D.; Fairhead, V.A. Defensive and sensory chemical ecology of brown algae. Adv. Bot. Res. 2006, 43, 1–91. [Google Scholar]
- Rice–Evans, R.E.; Miller, C.A.; Paganga, N.J.G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Schoenwaelder, M.E.A.; Clayton, M.N. Secretion of phenolic substances into the zygote wall and cell plate in embryos of Hormosira and Acrocarpis (Fucales, Phaeophyceae). J. Phycol. 1998, 34, 969–980. [Google Scholar] [CrossRef]
- Lüder, U.H.; Clayton, M.N. Induction of phlorotannins in the brown macroalga Ecklonia radiata (Laminariales, Phaeophyta) in response to simulated herbivory—The first microscopic study. Planta 2004, 218, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, K.; Shibata, T.; Fujimoto, K.; Honj, T.; Nakamura, T. Algicidal effect of phlorotannins from the brown alga Ecklonia kurome on red tide microalgae. Aquaculture 2003, 218, 601–611. [Google Scholar] [CrossRef]
- Ragan, M.A.; Smidsrød, O.; Bjørn, L. Chelation of divalent metal ions by brown algal polyphenols. Mar. Chem. 1979, 7, 265. [Google Scholar] [CrossRef]
- Cruces, E.; Rautenberger, R.; Rojas-Lillo, Y.; Cubillos, V.M.; Arancibia-Miranda, N.; Ramírez-Kushel, E.; Gómez, I. Physiological acclimation of Lessonia spicata to diurnal changing PAR and UV radiation: Differential regulation among down- regulation of photochemistry, ROS scavenging activity and phlorotannins as major photoprotective mechanisms. Photosynth. Res. 2017, 131, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Amsler, C.D.; Rowley, R.J.; Laur, D.R.; Quetin, L.B.; Ross, R.M. Vertical distribution of Antarctic peninsular macroalgae: Cover, biomass and species composition. Phycologia 1995, 34, 424–430. [Google Scholar] [CrossRef]
- Klöser, H.; Quartino, M.L.; Wiencke, C. Distribution of macroalgae and macroalgal communities in gradients of physical conditions in Potter Cove, King George Island, Antarctica. Hydrobiologia 1996, 333, 1–17. [Google Scholar] [CrossRef]
- Connan, S.; Goulard, F.; Stiger, V.; Deslandes, E.; Gall, E. Interspecific and temporal variation in phlorotannin levels in an assemblage of brown algae. Bot. Mar. 2004, 47, 410–416. [Google Scholar] [CrossRef]
- Connan, S.; Deslandes, E.; Gall, E. Influence of day-night and tidal cycles on phenol content and antioxidant capacity in three temperate intertidal brown seaweeds. J. Exp. Mar. Biol. Ecol. 2007, 349, 359–369. [Google Scholar] [CrossRef]
- Rautenberger, R.; Huovinen, P.; Gómez, I. Effects of increased seawater temperature on UV tolerance of Antarctic marine macroalgae. Mar. Biol. 2015, 162, 1087–1097. [Google Scholar] [CrossRef]
- Gómez, I.; Huovinen, P. Lack of physiological depth patterns in conspecifics of endemic antarctic brown algae: A trade-off between UV stress tolerance and shade adaptation? PLoS ONE 2015, 10, e0134440. [Google Scholar] [CrossRef] [PubMed]
- Flores-Molina, M.R.; Rautenberger, R.; Muñoz, P.; Huovinen, P.; Gómez, I. Stress tolerance of the endemic antarctic brown alga Desmarestia anceps to UV radiation and temperature is mediated by high concentrations of phlorotannins. Photochem. Photobiol. 2016, 92, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Iken, K.; Amsler, C.D.; Amsler, M.O.; McClintock, J.B.; Baker, B.J. Field studies on deterrent properties of phlorotannins in Antarctic brown algae. Bot. Mar. 2009, 52, 547–557. [Google Scholar] [CrossRef]
- Iken, K.; Amsler, C.D.; Hubbard, J.M.; McClintock, J.B.; Baker, B.J. Allocation patterns of phlorotannins in Antarctic brown algae. Phycologia 2007, 46, 386–395. [Google Scholar] [CrossRef]
- Cruces, E.; Huovinen, P.; Gómez, I. Phlorotannin and antioxidant responses upon short-term exposure to UV radiation and elevated temperature in three south Pacific kelps. Photochem. Photobiol. 2012, 88, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Huovinen, P.; Gómez, I. UV sensitivity of vegetative and reproductive tissues of two Antarctic Brown Algae is related to differential allocation of phenolic substances. Photochem. Photobiol. 2015, 91, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Gómez, I.; Español, S.; Véliz, K.; Huovinen, P. Spatial distribution of phlorotannins and its relationship with photosynthetic UV tolerance and allocation of storage carbohydrates in blades of the kelp Lessonia spicata. Mar. Biol. 2016, 163, 1–14. [Google Scholar] [CrossRef]
- Tala, F.; Penna-Díaz, M.A.; Luna-Jorquera, G.; Rothäusler, E.; Thiel, M. Daily and seasonal changes of photobiological responses in floating bull kelp Durvillaea antarctica (Chamisso) Hariot (Fucales: Phaeophyceae). Phycologia 2017, 56, 271–283. [Google Scholar] [CrossRef]
- Stern, J.L.; Hagerman, A.E.; Steinberg, P.D.; Winter, F.C.; Estes, J.A. A new assay for quantifying brown algal phlorotannins and comparisons to previous methods. J. Chem. Ecol. 1996, 22, 1273–1293. [Google Scholar] [CrossRef] [PubMed]
- Heffernan, N.; Brunton, N.P.; FitzGerald, R.J.; Smyth, T.J. Profiling of the molecular weight and structural isomer abundance of macroalgae-derived phlorotannins. Mar. Drugs 2015, 13, 509–528. [Google Scholar] [CrossRef] [PubMed]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 28, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Stachowicz, J.J.; Lindquist, N. Chemical defence among hydroids on pelagic Sargassum: Predator deterrence and absorption of solar UV radiation by secondary metabolites. Mar. Ecol. Prog. Ser. 1997, 155, 115–126. [Google Scholar] [CrossRef]
- Matz, M.V.; Fradkov, A.F.; Labas, Y.A.; Savitsky, A.P.; Zaraisky, A.G.; Markelov, M.L.; Lukyanov, S.A. Fluorescent proteins from nonbiolumninescent Anthozoa species. Nat. Biotechnol. 1999, 17, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, O.; Johnson, F.H.; Saiga, Y. Extraction, purification and properties of aequorin, a bioluminescent protein from the luminous hydromedusan. Aequorea J. Cell. Comp. Physiol. 1962, 59, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Tsein, R.Y. The green fluorescent protein. Annu. Rev. Biochem. 1998, 67, 509–544. [Google Scholar] [CrossRef] [PubMed]
- Bou-Abdallah, F.; Chasteen, N.D.; Lesser, M.P. Quenching of superoxide radicals by green fluorescent protein. Biochim. Biophys. Acta 2006, 1760, 1690–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 3rd ed.; Oxford Science Publications: Oxford, UK, 1999; pp. 1–936. [Google Scholar]
- McCord, J.M. Superoxide dismutase, lipid peroxidation, and bell-shaped dose response curves. Intern. Hormesis Soc. 2008, 6, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Staniek, K.; Nohl, H. H2O2 detection from intact mitochondria as a measure for one-electron reduction of dioxygen requires a non-invasive assay system. Biochim. Biophys. Acta 1999, 1413, 70–80. [Google Scholar] [CrossRef]
- Barata, C.; Lekumberri, I.; Vila-Escale, M.; Prat, N.; Porte, C. Trace metal concentration, antioxidant enzyme activities and susceptibility to oxidative stress in the trichoptera larvae Hydropsyche exocellata from the Llobregat river basin (NE Spain). Aquat. Toxicol. 2005, 74, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Storelli, C.; Acierno, R.; Maffia, M. Membrane lipid and protein adaptations in Antarctic fish. In Cold Ocean Physiology; Pörtner, H., Playle, R., Eds.; Cambridge University Press: Cambridge, UK, 1998; pp. 166–189. [Google Scholar]
- Avila, C.; Fontana, A.; Esposito, M.; Ciavatta, M.L.; Cimino, G. Fatty acids of Antarctic gastropods: Distribution and comparison with Mediterranean species. Iberus 2004, 22, 33–44. [Google Scholar]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, D.M.; Valentão, P.; Andrade, P.B. Marine natural pigments: Chemistry, distribution and analysis. Dyes Pigments 2014, 111, 124–134. [Google Scholar] [CrossRef]
- Britton, G. Structure and properties of carotenoids in relation to function. FASEB J. 1995, 9, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Magdaong, N.C.M.; Blankenship, R.E. Photoprotective, excited-state quenching mechanisms in diverse photosynthetic organisms. J. Biol. Chem. 2018, 293, 5018–5025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmore, A.M. Mechanistic aspects of xanthopyll cycle-dependent photoprotection in higher plant chloroplasts and leaves. Physiol. Plant. 1997, 99, 197–209. [Google Scholar] [CrossRef]
- Verhoeven, A.S.; Adams, W.W.; Demmig-Adams, B.; Croce, R.; Bassi, R. Xanthophyll cycle pigment localization and dynamics during exposure to low temperatures and light stress in Vinca major. Plant Physiol. 1999, 120, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Buma, A.G.J.; Wright, S.W.; van den Enden, R.; van de Poll, W.H.; Davidson, A.T. PAR acclimation and UVBR-induced DNA damage in Antarctic marine microalgae. Mar. Ecol. Prog. Ser. 2006, 315, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Hernando, M.; Schloss, I.; Roy, S.; Ferreyra, G. Photoacclimation to Long-Term Ultraviolet Radiation Exposure of Natural Sub-Antarctic Phytoplankton Communities: Fixed Surface Incubations versus Mixed Mesocosms. Photochem. Photobiol. 2006, 82, 923–935. [Google Scholar] [CrossRef] [PubMed]
- van de Poll, W.H.; Alderkamp, A.C.; Janknegt, P.J.; Roggeveld, J.; Buma, A.G.J. Photoacclimation modulates excessive photosynthetically active and ultraviolet radiation effects in a temperate and an Antarctic marine diatom. Limnol. Oceanogr. 2006, 51, 1239–1248. [Google Scholar] [CrossRef] [Green Version]
- Wynn-Williams, D.D.; Edwards, H.G.M. Environmental UV Radiation: Biological Strategies for Protection and Avoidance. In Astrobiology; Horneck, G., Baumstark-Khan, C., Eds.; Springer: Berlin, Heidelberg, Germany, 2002; pp. 245–260. [Google Scholar]
- Palmisano, A.C.; Wharton, R.A.; Cronin, S.E.; des Marais, D.J. Lipophilic pigments from the benthos of a perennially ice-covered Antarctic lake. Hydrobiologia 1989, 178, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Potts, M.; Olie, J.J.; Nickels, J.S.; Parsons, J.; White, D.C. Variation in phospholipid ester-linked fatty acids and carotenoids of dessicated Nostoc commune (Cyanobacteria) from different geographic locations. Appl. Environ. Microbiol. 1987, 53, 4–9. [Google Scholar] [PubMed]
- Órdenes-Aenishanslins, N.; Anziani-Ostuni, G.; Vargas-Reyes, M.; Alarcón, J.; Tello, A.; Pérez-Donoso, J.M. Pigments from UV-resistant Antarctic bacteria as photosensitizers in Dye Sensitized Solar Cells. J. Photochem. Photobiol. B Biol. 2016, 162, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Oh, H.M.; Chung, S.; Cho, J.C. Antarcticimonas flava gen. nov., sp. nov., isolated from Antarctic coastal seawater. J. Microbiol. 2009, 47, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Yu, P.S.; Zhou, Y.D.; Xu, L.; Wang, C.S.; Wu, M.; Oren, A.; Xu, X.W. Muricauda antarctica sp. nov., a marine member of the Flavobacteriaceae isolated from Antarctic seawater. Int. J. Syst. Evol. Microbiol. 2013, 63, 3451–3456. [Google Scholar] [CrossRef] [PubMed]
- Dieser, M.; Greenwood, M.; Foreman, C.M. Carotenoid Pigmentation in Antarctic Heterotrophic Bacteria as a Strategy to Withstand Environmental Stresses. Arct. Antarct. Alp. Res. 2010, 42, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.P.; Kashyap, A.K. An assessment of biopotential of three cyanobacterial isolates from Antarctic for carotenoid production. Indian J. Biochem. Biophys. 2003, 40, 362–366. [Google Scholar] [PubMed]
- Marquardt, J.; Hanelt, D. Carotenoid composition of Delesseria lancifolia and other marine red algae from polar and temperate habitats. Eur. J. Phycol. 2004, 39, 285–292. [Google Scholar] [CrossRef]
- Matsuno, T. Aquatic animal carotenoids. Fish. Aquat. Sci. 2001, 67, 771–783. [Google Scholar] [CrossRef]
- Rangarao, A.; Moi, P.S.; Sarada, R.; Ravishankar, G.A. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications—A Review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [Green Version]
- Ulven, S.M. Metabolic Effects of Krill Oil. In Fish and Fish Oil in Health and Disease Prevention; Raatz, S.K., Bibus, D.M., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 333–339. [Google Scholar]
- Czeczuga, B.; Klyszejko, B. Carotenoids in fish XIV. The carotenoid content in the flesh of certain species from the Antarctic. Hydrobiologia 1978, 60, 173–175. [Google Scholar] [CrossRef]
- Czeczuga, B. Carotenoid-calcium carbonate complex from the hydrocoral Errina antarctica. Biochem. Syst. Ecol. 1985, 13, 455–457. [Google Scholar] [CrossRef]
- Mène-Saffrané, L. Vitamin E Biosynthesis and Its Regulation in Plants. Antioxidants 2017, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Fryer, M.J. Evidence for the photoprotective effects of vitamin E. Photochem. Photobiol. 1993, 58, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.; Witt, E.; Goldman, R.; Scita, G.; Packer, L. Ultraviolet light-induced generation of vitamin E radicals and their recycling. A possible photosensitizing effect of vitamin E in skin. Free Radic. Res. Commun. 1992, 16, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Podhaisky, H.P.; Wohlrab, W. Is the photoprotective effect of vitamin E based on its antioxidative capacity? J. Dermatol. Sci. 2002, 28, 84–86. [Google Scholar] [CrossRef]
- Hernando, M.; Malanga, G.; Puntarulo, S.; Ferreyra, G. Non-enzymatic antioxidant photoprotection against potential UVBR-induced damage in an Antarctic diatom (Thalassiosira sp.). Lat. Am. J. Aquat. Res. 2011, 39, 397–408. [Google Scholar] [CrossRef]
- Estevez, M.S.; Abele, D.; Puntarulo, S. Lipid radical generation in polar (Laternula elliptica) and temperate (Mya arenaria) bivalves. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 132, 729–737. [Google Scholar] [CrossRef]
- Woodward, B. Dietary vitamin requirements of cultured young fish, with emphasis on quantitative estimates for salmonids. Aquaculture 1994, 124, 133–168. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Fujisawa, A.; Hara, A.; Dunlap, W.C. An unusual vitamin E constituent provides antioxidant protection in marine organisms adapted to coldwater environments. Proc. Natl. Acad. Sci. USA 2001, 98, 13144–13148. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Cuddihy, S.; Hill, J.V.; Davison, W. A comparison of plasma vitamin C and E levels in two Antarctic and two temperate water fish species. Comp. Biochem. Physiol. B 2000, 125, 371–378. [Google Scholar] [CrossRef]
- Dunlap, W.C.; Fujisawa, A.; Yamamoto, Y.; Moylan, T.J.; Sidell, B.D. Notothenioid fish, krill and phytoplankton from Antarctica contain a vitamin E constituent (α-tocomonoenol) functionally associated with cold-water adaptation. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 133, 299–305. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Maita, N.; Fujisawa, A.; Takashima, J.; Ishii, Y.; Dunlap, W.C. A new vitamin E (α-tocomonoenol) from eggs of the Pacific salmon Oncorhynchus keta. J. Nat. Prod. 1999, 62, 1685–1687. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, J.Y.; Wang, S.Q. The role of antioxidants in photoprotection: A critical review. J. Am. Acad. Dermatol. 2012, 67, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Michels, A.J.; Frei, B. Vitamin C. Adv. Nutr. 2014, 5, 16–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahns, P.; Latowski, D.; Strzalka, K. Mechanism and regulation of the violaxanthin cycle: The role of antenna proteins and membrane lipids. Biochem. Biophys. Acta Bioenerg. 2009, 1787, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estevez, M.S.; Malanga, G.; Puntarulo, S. UV-B effects on Antarctic Chlorella sp cells. J. Photochem. Photobiol. B Biol. 2001, 62, 19–25. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 191–212. [Google Scholar] [CrossRef]
- Niyogi, K.K. Photoprotection revisited: Genetics and molecular approaches. Annu. Rev. Plant Physiol. 1999, 50, 333–359. [Google Scholar] [CrossRef] [PubMed]
- Winkler, B.S.; Orselli, S.M.; Rex, T.S. The redox couple between glutathione and ascorbic acid: A chemical and physiological perspective. Free Radic. Biol. Med. 1994, 17, 333–349. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Laureau, C.; Bligny, R.; Streb, P. The significance of glutathione for photoprotection at contrasting temperatures in the alpine plant species Soldanella alpine and Ranunculus glacialis. Physiol. Plant. 2011, 143, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Häder, D.P.; Sinha, R.P. Cyanobacteria and ultraviolet radiation (UVR) stress: Mitigation strategies. Ageing Res. Rev. 2010, 9, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Tel-Or, E.; Huflejt, M.E.; Packer, L. Hydroperoxide metabolism in cyanobacteria. Arch. Biochem. Biophys. 1986, 246, 396–402. [Google Scholar] [CrossRef]
- Alscher, R.G.; Donahue, J.H.; Cramer, C.L. Reactive oxygen species and antioxidants: Relationships in green cells. Physiol. Plant. 1997, 100, 224–233. [Google Scholar] [CrossRef]
- Janknegt, P.J.; van de Poll, W.H.; Visser, R.J. Oxidative stress responses in the marine Antarctic diatom Chaetoceros brevis (bacillariophyceae) during photoacclimation. J. Phycol. 2008, 44, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Philipp, E.; Brey, T.; Pörtner, H.O.; Abele, D. Chronological and physiological ageing in a polar and a temperate mud clam. Mech. Ageing Dev. 2005, 126, 598–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassini, A.; Favero, M.; Albergoni, V. Comparative studies of antioxidant enzymes in red-blooded and white-blooded Antarctic teleost fish Pagothenia bernacchii and Chionodraco hamatus. Comp. Biochem. Physiol. 1993, 106, 333–336. [Google Scholar] [CrossRef]
- Ansaldo, M.; Luquet, C.M.; Evelson, P.A.; Polo, J.M.; Llesuy, S. Antioxidant levels from different Antarctic fish caught around South Georgia Island and Shag Rocks. Polar Biol. 2000, 23, 160–165. [Google Scholar] [CrossRef]
- Abele, D.; Puntarulo, S. Formation of reactive species and induction of antioxidant defence systems in polar and temperate marine invertebrates and fish. Comp. Biochem. Physiol. Part A 2004, 138, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.; Zaleski, T.; Rodrigues, E.; Carvalho, C.; Cadena, S.M.; Gozzi, G.; Krebsbach, P.; Rios, F.S.; Donatti, L. Effect of temperature acclimation on the liver antioxidant defence system of the Antarctic nototheniids Notothenia coriiceps and Notothenia rossii. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2014, 172–173, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.D.; Borges, V.D.; Rosa, C.E.; Colares, E.P.; Robaldo, R.B.; Martinez, P.E.; Bianchini, A. Effects of increasing temperature on antioxidant defense system and oxidative stress parameters in the Antarctic fish Notothenia coriiceps and Notothenia rossii. J. Therm. Biol. 2017, 68, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Giordano, D.; Coppola, D.; Russo, R.; Tinajero-Trejo, M.; di Prisco, G.; Lauro, F.; Ascenzi, P.; Verde, C. The globins of cold-adapted Pseudoalteromonas haloplanktis TAC125: From the structure to the physiological functions. Adv. Microb. Physiol. 2013, 63, 329–389. [Google Scholar] [PubMed]
- Giordano, D.; Russo, R.; Coppola, D.; Altomonte, G.; di Prisco, G.; Bruno, S.; Verde, C. “Cool” adaptations to cold environments: Globins in Notothenioidei. Hydrobiologia 2015, 761, 293–312. [Google Scholar] [CrossRef]
- Cortat, B.; Garcia, C.C.M.; Quinet, A.; Schuch, A.P.; de Lima-Bessa, K.M.; Menck, C.F.M. The relative roles of DNA damage induced by UVA irradiation in human cells. Photochem. Photobiol. Sci. 2013, 12, 1483–1495. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.R.P.; Singh, S.P.; Häder, D.P. Photoprotective compounds from marine organisms. J. Ind. Microbiol. Biotechnol. 2010, 37, 537–558. [Google Scholar] [CrossRef] [PubMed]
- Richa; Sinha, R.P.; Häder, D.P. Physiological aspects of UV-excitation of DNA. In Photoinduced Phenomena in Nucleic Acids; Barbatti, B., Borin, A.C., Ullrich, A.C., Eds.; Springer: Berlin, Germany, 2014; pp. 1–46. [Google Scholar]
- Karentz, D. Beyond Xeroderma pigmentosum: DNA Damage and repair in an ecological context. A tribute to James, E. Cleaver. Photochem. Photobiol. 2015, 91, 460–474. [Google Scholar] [CrossRef] [PubMed]
- Buma, A.G.J.; Karin de Boer, M.; Boelen, P. Depth distributions of DNA damage in antarctic marine phyto and bacterioplankton exposed to summertime UV radiation. J. Phycol. 2001, 37, 200–208. [Google Scholar] [CrossRef]
- Eisen, J.A.; Hanawalt, P.C. A phylogenomic study of DNA repair genes, proteins, and processes. Mutat. Res. 1999, 435, 171–213. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, P.J. Catalytic promiscuity and the divergent evolution of DNA repair enzymes. Chem. Rev. 2006, 106, 720–752. [Google Scholar] [CrossRef] [PubMed]
- Rooney, J.P.; Patil, A.; Joseph, F.; Endres, L.; Begley, U.; Zappala, M.R.; Cunningham, R.P.; Begley, T.J. Cross-species functionome analysis identifies proteins associated with DNA repair, translation and aerobic respiration as conserved modulators of UV-toxicity. Genomics 2011, 97, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Essen, L.O.; Klar, T. Light-driven DNA repair by photolyases. Cell. Mol. Life Sci. 2006, 63, 1266–1277. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Todo, T.; Ayaki, H.; Ishizaki, K.; Morita, T.; Mitra, S.; Ikenaga, M. Cloning of a marsupial DNA photolyase gene and the lack of related nucleotide sequences in placental mammals. Nucleic Acids Res. 1994, 22, 4119–4124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleaver, J.E. Photoreactivation: A radiation repair mechanism absent from mammalian cells. Biochem. Biophys. Res. Commun. 1966, 24, 569–576. [Google Scholar] [CrossRef]
- Johnson, J.L.; Hamm-Alvarez, S.; Payne, G.; Sancar, G.B.; Rajagopalan, K.V.; Sancar, A. Identification of the second chromophore of Escherichia coli and yeast DNA photolyases as 5,10-methenyltetrahydrofolate. Proc. Natl. Acad. Sci. USA 1988, 85, 2046–2050. [Google Scholar] [CrossRef] [PubMed]
- Eker, A.P.M.; Kooiman, P.; Hessels, J.K.C.; Yasui, A. DNA photoreactivating enzyme from the cyanobacterium Anacystis nidulans. J. Biol. Chem. 1990, 265, 8009–8015. [Google Scholar] [PubMed]
- Wang, J.; Du, X.; Pan, W.; Wang, X.; Wu, W. Photoactivation of the cryptochrome/photolyase superfamily. J. Photochem. Photobiol. C Photochem. Rev. 2015, 22, 84–102. [Google Scholar] [CrossRef]
- Sancar, A.; Sancar, G.B. DNA repair enzymes. Annu. Rev. Biochem. 1988, 57, 29–67. [Google Scholar] [CrossRef] [PubMed]
- Marizcurrena, J.J.; Morel, M.A.; Braña, V.; Morales, D.; Martinez-López, W.; Castro-Sowinski, S. Searching for novel photolyases in UVC-resistant Antarctic bacteria. Extremophiles 2017, 21, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ma, L.; Mou, S.; Wang, Y.; Zheng, Z.; Liu, F.; Qi, X.; An, M.; Chen, H.; Miao, J. Cyclobutane pyrimidine dimers photolyase from extremophilic microalga: Remarkable UV-B resistance and efficient DNA damage repair. Mutat. Res. 2015, 773, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Lud, D.; Moerdijk, T.C.W.; van de Poll, W.H.; Buma, A.G.J.; Huiskes, A.H.L. DNA damage and photosynthesis in Antarctic and Arctic Sanionia uncinata (Hedw.) Loeske under ambient and enhanced levels of UV-B radiation. Plant Cell Environ. 2002, 25, 1579–1589. [Google Scholar] [CrossRef]
- Lamare, M.D.; Barker, M.F.; Lesser, M.P.; Marshall, C. DNA photorepair in echinoid embryos: Effects of temperature on repair rate in Antarctic and non-Antarctic species. J. Exp. Biol. 2006, 209, 5017–5028. [Google Scholar] [CrossRef] [PubMed]
- Isely, N.; Lamare, M.; Marshall, C.; Barker, M. Expression of the DNA repair enzyme, photolyase, in developmental tissues and larvae, and in response to ambient UV-R in the Antarctic sea urchin Sterechinus neumayeri. Photochem. Photobiol. 2009, 85, 1168–1176. [Google Scholar] [CrossRef] [PubMed]
- Dantuma, N.P.; Heinen, C.; Hoogstraten, D. The ubiquitin receptor Rad23: At the crossroads of nucleotide excision repair and proteasomal degradation. DNA Repair 2009, 8, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Bergink, S.; Toussaint, W.; Luijsterburg, M.S.; Dinant, C.; Alekseev, S.; Hoeijmakers, J.H.J.; Dantuma, N.P.; Houtsmuller, A.B.; Vermeulen, W. Recognition of DNA damage by XPC coincides with disruption of the XPC–RAD23 complex. J. Cell Biol. 2012, 196, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, J.N. An Investigation of Sunlight Stress Response Genes in the Intertidal Sea Anemone, Actinia tenebrosa. Master’s Thesis, Queensland University of Technology, Queensland, New Zealand, 2016. [Google Scholar]
- Janion, C. Inducible SOS Response System of DNA Repair and Mutagenesis in Escherichia coli. Int. J. Biol. Sci. 2008, 4, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Courcelle, J.; Khodursky, A.; Peter, B.; Brown, P.O.; Hanawalt, P.C. Comparative gene expression profiles following UV exposure in wild-type and SOS deficient Escherichia coli. Genetics 2001, 58, 41–64. [Google Scholar]
- Walker, G.C. Mutagenesis and induced responses to deoxyribonucleic acid damage in Escherichia coli. Microbiol. Rev. 1984, 48, 60–93. [Google Scholar] [PubMed]
- Miller, R.V. recA: The gene and its protein product. In Encyclopedia of Microbiology, 2nd ed.; Luria, S., Ed.; Academic Press: San Diego, CA, USA, 2000; Volume 4, pp. 43–54. [Google Scholar]
- Booth, M.G.; Hutchinson, L.; Brumsted, M.; Aas, P.; Coffin, R.B.; Downer, R.C.; Kelley, C.A.; Lyons, M.M.; Pakulski, J.D.; Sandvik, S.L.H.; et al. Quantification of recA gene expression as an indicator of repair potential in marine bacterioplankton communities of Antarctica. Aquat. Microb. Ecol. 2001, 24, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, J.N.; Roy, S.; Ferreyra, G.; Campbell, D.A.; Curtosi, A. Ultraviolet-B effects on photosystem II efficiency of natural phytoplankton communities from Antarctica. Polar Biol. 2005, 28, 607–618. [Google Scholar] [CrossRef]
- Apostolova, E.L.; Pouneva, I.; Rashkov, G.; Dankov, K.; Grigorova, I.; Misra, A.N. Effect of UV-B radiation on photosystem II functions in Antarctic and mesophilic strains of a green alga Chlorella vulgaris and a cyanobacterium Synechocystis salina. Ind. J. Plant Physiol. 2014, 19, 111–118. [Google Scholar] [CrossRef]
- Mattoo, A.K.; Hoffman-Falk, H.; Marder, J.B.; Edelman, M. Regulation of protein metabolism: Coupling of photosynthetic electron transport to in vivo degradation of the rapidly metabolized 32-kilodalton protein of chloroplast membranes. Proc. Natl. Acad. Sci. USA 1984, 81, 1380–1384. [Google Scholar] [CrossRef] [PubMed]
- Prásil, O.; Adir, N.; Ohad, I. Dynamics of photosystem II: Mechanism of photoinhibition and recovery processes. In The photosystems: Structure, Function and Molecular Biology. Topics in Photosynthesis; Barber, J., Ed.; Elsevier: Amsterdam, The Netherlands, 1992; Volume 11, pp. 295–348. [Google Scholar]
- Aro, E.M.; Virgin, I.; Andersson, B. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1993, 1143, 113–134. [Google Scholar] [CrossRef]
- Sass, L.; Spetea, C.; Maté, Z.; Nagy, F.; Vass, I. Repair of UV-B induced damage of photosystem II via de novo synthesis of D1 and D2 reaction centre subunits in Synechocystis sp. PCC 6803. Photosynth. Res. 1997, 54, 55–62. [Google Scholar] [CrossRef]
- Verschooten, L.; Claerhout, S.; Laethemii, A.V.; Agostinis, P.; Garmyn, M. New strategies of photoprotection. Photochem. Photobiol. 2006, 82, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Kiang, J.G.; Tsokos, G.C. Heat shock protein 70 kDa: Molecular biology, biochemistry, and physiology. Pharmacol. Ther. 1998, 80, 183–201. [Google Scholar] [CrossRef]
- Bierkens, J.G. Applications and pitfalls of stress-proteins in biomonitoring. Toxicology 2000, 153, 61–72. [Google Scholar] [CrossRef]
- La Terza, A.; Barchetta, S.; Buonanno, F.; Ballarini, P.; Miceli, C. The protozoan ciliate Tetrahymena thermophila as biosensor of sublethal levels of toxicants in the soil. Fresenius Environ. Bull. 2008, 17, 1144–1150. [Google Scholar]
- Sarkar, S.; Singh, D.M.; Yadav, R.; Arunkumar, K.P.; Pittman, G.W. Heat shock proteins: Molecules with assorted functions. Front. Biol. 2011, 6, 312–327. [Google Scholar]
- Torok, Z.; Goloubinoff, P.; Horvath, I.; Tsvetkova, N.M.; Glatz, A.; Balogh, G.; Varvasovszki, V.; Los, D.A.; Vierling, E.; Crowe, J.H.; et al. Synechocystis HSP17 is an amphitropic protein that stabilizes heat-stressed membranes and binds denatured proteins for subsequent chaperone-mediated refolding. Proc. Natl. Acad. Sci. USA 2001, 98, 3098–3103. [Google Scholar] [CrossRef] [PubMed]
- Horváth, I.; Glatz, A.; Varvasovszki, V.; Török, Z.; Páli, T.; Balogh, G.; Kovács, E.; Nádasdi, L.; Benkö, S.; Joó, F.; et al. Membrane physical state controls the signaling mechanism of the heat shock response in Synechocystis PCC 6803: Identification of hsp17 as a “fluidity gene”. Proc. Natl. Acad. Sci. USA 1998, 95, 3513–3518. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, P.; Cong, B.; Liu, C.; Lin, X.; Shen, J.; Huang, X. Molecular cloning and expression analysis of a cytosolic Hsp70 gene from Antarctic ice algae Chlamydomonas sp. ICE-L. Extremophiles 2010, 14, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Fulgentini, L.; Passini, V.; Colombetti, G.; Miceli, C.; La Terza, A.; Marangoni, R. UV Radiation and Visible Light Induce hsp70 Gene Expression in the Antarctic Psychrophilic Ciliate Euplotes focardii. Microb. Ecol. 2015, 70, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Dahms, H.U.; Lee, J.S. UV radiation in marine ectotherms: Molecular effects and responses. Aquat. Toxicol. 2010, 97, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P.; Moh, S.H.; Lee, T.K.; Kottuparambil, S.; Kim, Y.; Rhee, J.S.; Choi, E.M.; Brown, M.T.; Häder, D.P.; et al. Ultraviolet radiation and cyanobacteria. J. Photochem. Photobiol. B 2014, 141, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, R.; Poma, V.; Costa, C.; Matranga, V. UV-B radiation prevents skeleton growth and stimulates the expression of stress markers in sea urchin embryos. Biochem. Biophys. Res. Commun. 2005, 328, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Franklin, D.J.; Brussaard, C.P.D.; Berges, J.A. What is the role and nature of programmed cell death in phytoplankton ecology? Eur. J. Phycol. 2006, 41, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Vardi, A.; Berman-Frank, I.; Rozenberg, T.; Hadas, O.; Kaplan, A.; Levine, A. Programmed cell death of the dinoflagellate Peridinium gatunense is mediated by CO2 limitation and oxide stress. Curr. Biol. 1999, 9, 1061–1064. [Google Scholar] [CrossRef]
- Häder, D.P.; Kumar, H.D.; Smith, R.C.; Worrest, R.C. Effects on aquatic ecosystems. J. Photochem. Photobiol. B 1998, 46, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Gago-Ferrero, P.; Alonso, M.B.; Bertozzi, C.P.; Mango, J.; Barbosa, L.; Cremer, M.; Secchi, E.R.; Domit, C.; Azevedo, A.; Lailson-Brito, J.; et al. First determination of UV filters in marine mammals. Octocrylene levels in Franciscana dolphins. Environ. Sci. Technol. 2013, 47, 5619–5625. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, P.R.; Vinebrooke, R.D.; Donald, D.B.; Smol, J.P.; Schindler, D.W. Past ultraviolet radiation environments in lakes derived from fossil pigments. Nature 1997, 388, 457–459. [Google Scholar] [CrossRef]
- Chrapusta, E.; Kaminski, A.; Duchnik, K.; Bober, B.; Adamski, M.; Bialczyk, J. Mycosporine-Like Amino Acids: Potential Health and Beauty Ingredients. Mar. Drugs 2017, 15, 326. [Google Scholar] [CrossRef] [PubMed]
- Bird, G.; Fitzmaurice, N.; Dunlap, W.C.; Chalker, B.E.; Bandaranayake, W.M. Sunscreen Compositions and Compounds for Use Therein. Australian Patent 595,075, 26 September 1986. [Google Scholar]
- Chalmers, P.J.; Fitzmaurice, N.; Rigg, D.J.; Thang, S.H.; Bird, G. UV Absorbing Compounds and Compositions. Australian Patent 653,495, 7 September 1990. [Google Scholar]
- Schmid, D.; Schürch, C.; Zülli, F.; Nissen, H.P.; Prieur, H. Mycosporine-like amino acids: Natural UV-screening compounds from red algae to protect the skin against photoaging. SÖFW-J. 2003, 129, 1–5. [Google Scholar]
- Torres, A.; Enk, C.D.; Hochberg, M.; Srebnik, M. Porphyra-334, a potential natural source for UVA protective sunscreens. Photochem. Photobiol. Sci. 2006, 5, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Oyamada, C.; Kaneniwa, M.; Ebitani, K.; Murata, M.; Ishihara, K. Mycosporine-like amino acids extracted from scallop (Patinopecten yessoensis) ovaries: UV protection and growth stimulation activities on human cells. Mar. Biotechnol. 2008, 10, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Schmid, D.; Schürch, C.; Zülli, F. Mycosporine-like amino acids from red algae protect against premature skin-aging. Eur. Cosmet. 2006, 9, 1–4. [Google Scholar]
- Cardozo, K.H.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Andre, G.; Pellegrini, M.; Pellegrini, L. Algal Extracts Containing Amino Acid Analogs of Mycosporin Are Useful as Dermatological Protecting Agents against Ultraviolet Radiation. Patent No. FR2803201, 6 July 2001. [Google Scholar]
- Sathasivam, R.; Ki, J.S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- The Global Market for Carotenoids. Available online: https://www.bccresearch.com/market-research/food-and-beverage/carotenoids-global-market-report-fod025e.html (accessed on 15 April 2018).
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from marine organisms: Biological functions and industrial applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- CODIF. Available online: http://www.codif-tn.com/en?s=dermochlorella (accessed on 18 April 2018).
- Frutarom. Available online: http://www.frutarom.com/FrutaromNew/Templates/showpage.asp?DBID=1&LNGID=1&TMID=84&FID=534 (accessed on 18 April 2018).
- Lipotec. Available online: https://www.lipotec.com/en/products/seacode-trade-marine-ingredient/ (accessed on 18 April 2018).
- Lipotec. Available online: https://www.lipotec.com/en/products/antarcticine-reg-marine-ingredient/ (accessed on 18 April 2018).
- Karentz, D.; Bosch, I.; Dunlap, W.C. Distribution of UV-absorbing compounds in the Antarctic limpet, Nacella concinna. Antarct. J. U. S. 1992, 27, 121–122. [Google Scholar]
- Fairhead, V.A.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Within-thallus variation in chemical and physical defences in two species of ecologically dominant brown macroalgae from the Antarctic Peninsula. J. Exp. Mar. Biol. Ecol. 2005, 322, 1–12. [Google Scholar] [CrossRef]
- Fairhead, V.A.; Amsler, C.D.; McCLintock, J.B.; Baker, B.J. Lack of defence or phlorotannin induction by UV radiation or mesograzers in Desmarestia anceps and D. menziesii (phaeophyceae). J. Phycol. 2006, 42, 1174–1183. [Google Scholar] [CrossRef]
- Döhler, G. Effect of ultraviolet radiation on pigmentation and nitrogen metabolism of antarctic phytoplankton and ice algae. J. Plant Physiol. 1998, 153, 603–609. [Google Scholar] [CrossRef]
Figure 1.
Negative effects of ultraviolet radiation (UV-R) on marine environment. Main factors involved in the attenuation of light through the air and water column and mechanisms of UV tolerance in biological systems by mitigating strategies and repair processes. UV-R may affect organisms through molecular and/or cellular damages, genetic mutations, or by causing disturbances at population and community levels, interfering with physiological functions (e.g., growth, reproduction and behaviour), and species interactions with effects on the ecosystem and biogeochemical cycles. ROS, reactive oxygen species; CDOM, coloured dissolved organic matter.
Figure 1.
Negative effects of ultraviolet radiation (UV-R) on marine environment. Main factors involved in the attenuation of light through the air and water column and mechanisms of UV tolerance in biological systems by mitigating strategies and repair processes. UV-R may affect organisms through molecular and/or cellular damages, genetic mutations, or by causing disturbances at population and community levels, interfering with physiological functions (e.g., growth, reproduction and behaviour), and species interactions with effects on the ecosystem and biogeochemical cycles. ROS, reactive oxygen species; CDOM, coloured dissolved organic matter.

Figure 2.
Mycosporine-like amino acids (MAAs) found in Antarctic marine organisms: (1) Asterina-330; (2) mycosporine-glycine; (3) mycosporine-2-glycine; (4) mycosporine-glycine-valine; (5) palythene; (6) palythenic acid; (7) palythine; (8) palythinol; (9) porphyra-334; (10) shinorine; (11) usujirene.
Figure 2.
Mycosporine-like amino acids (MAAs) found in Antarctic marine organisms: (1) Asterina-330; (2) mycosporine-glycine; (3) mycosporine-2-glycine; (4) mycosporine-glycine-valine; (5) palythene; (6) palythenic acid; (7) palythine; (8) palythinol; (9) porphyra-334; (10) shinorine; (11) usujirene.

Figure 3.
(12) Scytonemin.

Figure 4.
(13) Erebusinone.

Figure 5.
(14) Melanin.

Figure 6.
Examples of marine phlorotannins: (15) Eckol; (16) fucol; (17) fucophlorethol; (18) fuhalol; (19) phlorethol.
Figure 6.
Examples of marine phlorotannins: (15) Eckol; (16) fucol; (17) fucophlorethol; (18) fuhalol; (19) phlorethol.

Figure 7.
Carotenoids: (20) 4-hydroxy-α-carotene; (21) α-carotene; (22) antheraxanthin; (23) astaxanthin; (24) aurochrome; (25) auroxanthin; (26) β-carotene; (27) canthaxanthin; (28) cryptoxanthin; (29) diadinoxanthin; (30) diatoxanthin; (31) flavoxanthin; (32) γ-carotene; (33) isozeaxanthin; (34) lutein-5,6-epoxide; (35) lutein; (36) tunaxanthin; (37) violaxanthin; (38) zeaxanthin.
Figure 7.
Carotenoids: (20) 4-hydroxy-α-carotene; (21) α-carotene; (22) antheraxanthin; (23) astaxanthin; (24) aurochrome; (25) auroxanthin; (26) β-carotene; (27) canthaxanthin; (28) cryptoxanthin; (29) diadinoxanthin; (30) diatoxanthin; (31) flavoxanthin; (32) γ-carotene; (33) isozeaxanthin; (34) lutein-5,6-epoxide; (35) lutein; (36) tunaxanthin; (37) violaxanthin; (38) zeaxanthin.

Figure 8.
(39) α-tocopherol.

Figure 9.
(40) α-tocomonoenol.

Figure 10.
(41) Ascorbic acid.

Figure 11.
(42) Glutathione.

Figure 12.
Antioxidant enzymes involved in the detoxification of reactive oxygen species (ROS). SOD, superoxide dismutase; CAT, catalase; APX, ascorbate peroxidase; GSH, glutathione.
Figure 12.
Antioxidant enzymes involved in the detoxification of reactive oxygen species (ROS). SOD, superoxide dismutase; CAT, catalase; APX, ascorbate peroxidase; GSH, glutathione.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Núñez-Pons, L.; Avila, C.; Romano, G.; Verde, C.; Giordano, D. UV-Protective Compounds in Marine Organisms from the Southern Ocean. Mar. Drugs 2018, 16, 336. https://doi.org/10.3390/md16090336
AMA Style
Núñez-Pons L, Avila C, Romano G, Verde C, Giordano D. UV-Protective Compounds in Marine Organisms from the Southern Ocean. Marine Drugs. 2018; 16(9):336. https://doi.org/10.3390/md16090336
Chicago/Turabian StyleNúñez-Pons, Laura, Conxita Avila, Giovanna Romano, Cinzia Verde, and Daniela Giordano. 2018. "UV-Protective Compounds in Marine Organisms from the Southern Ocean" Marine Drugs 16, no. 9: 336. https://doi.org/10.3390/md16090336
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.