Marine-Derived Metabolites of S-Adenosylmethionine as Templates for New Anti-Infectives

Abstract
:1. Introduction
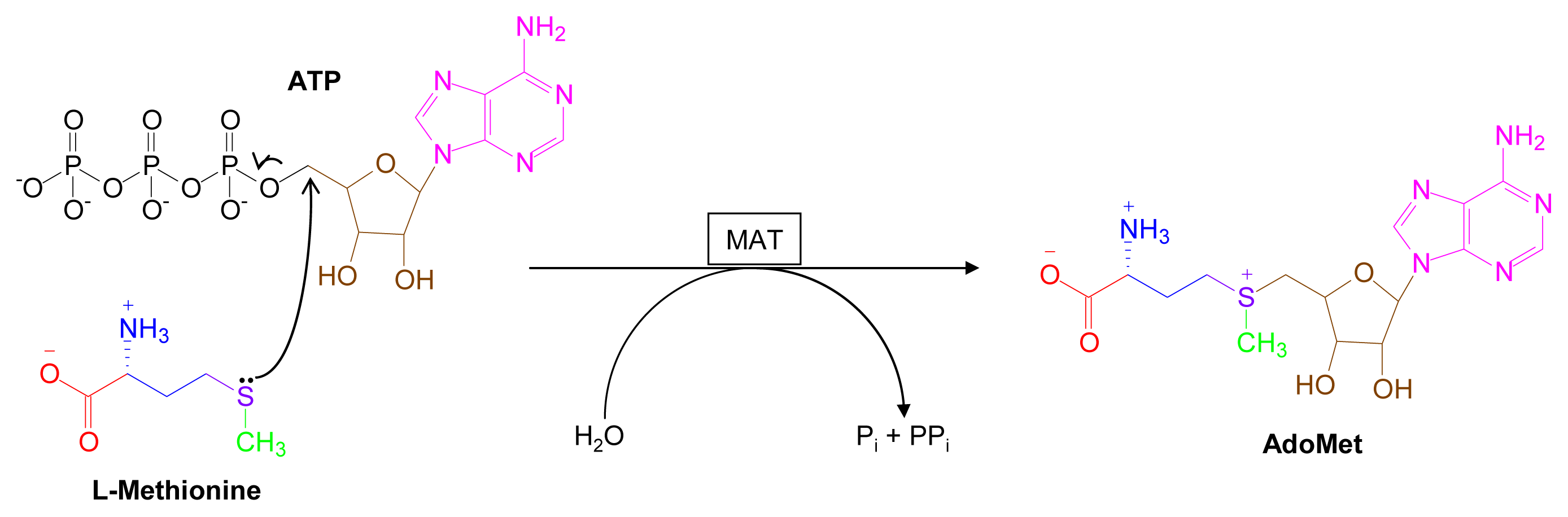
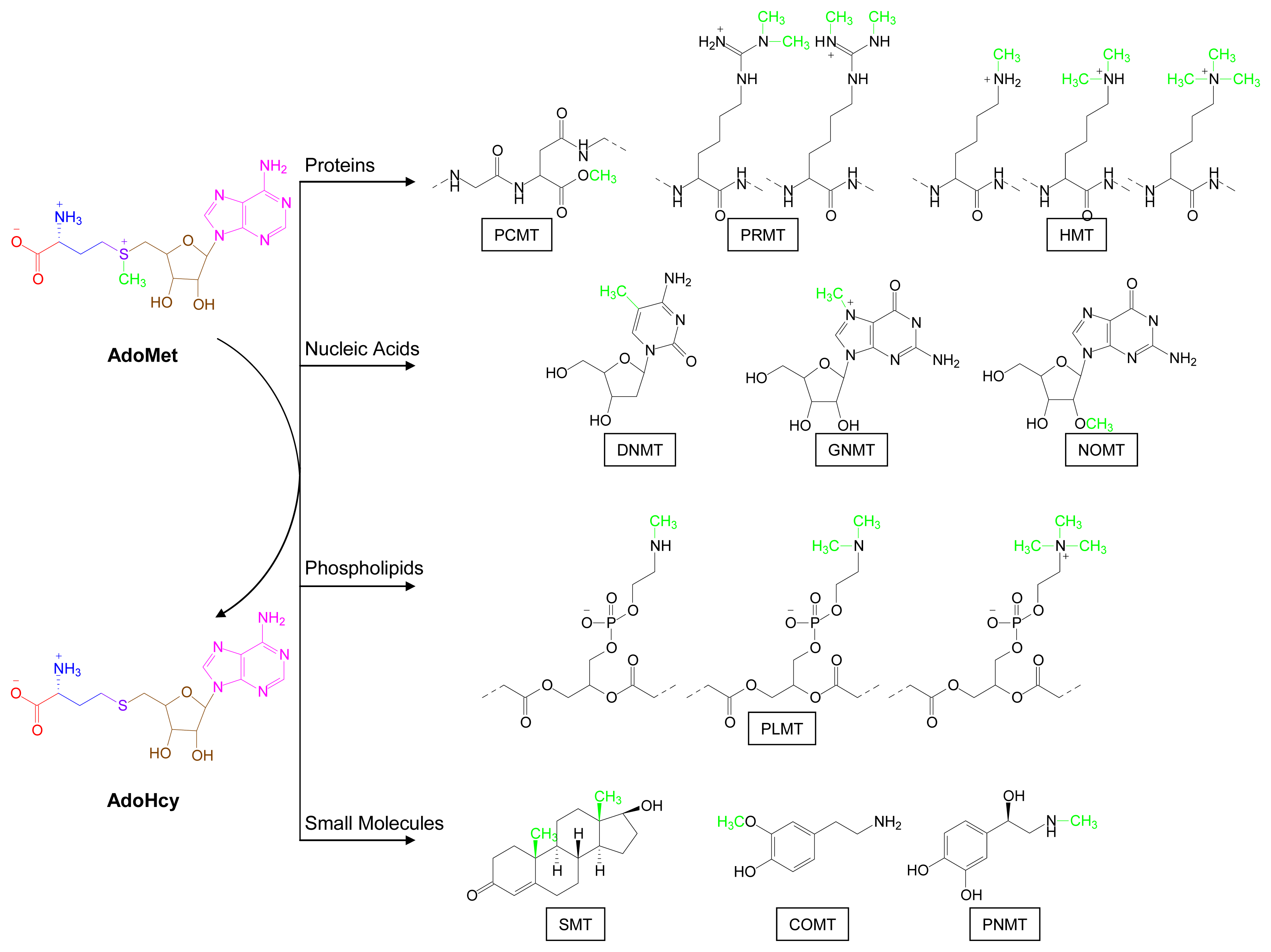
2. Overview of Major AdoMet-Utilizing Pathways
3. AdoMet Pathways and Marine-Derived AdoMet Metabolites
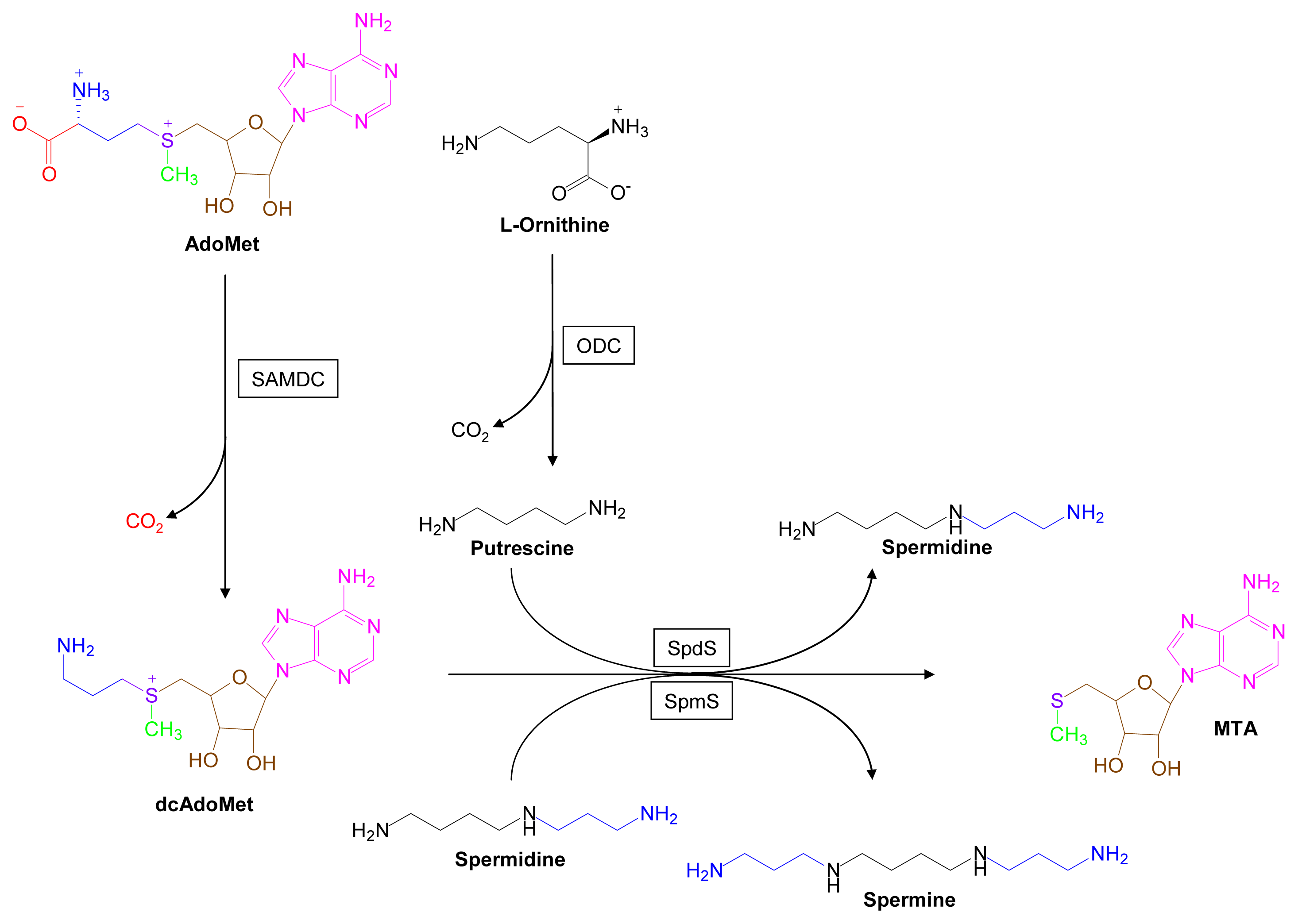
3.1. Polyamine Pathways [37–44]
3.2. Methylation Pathways
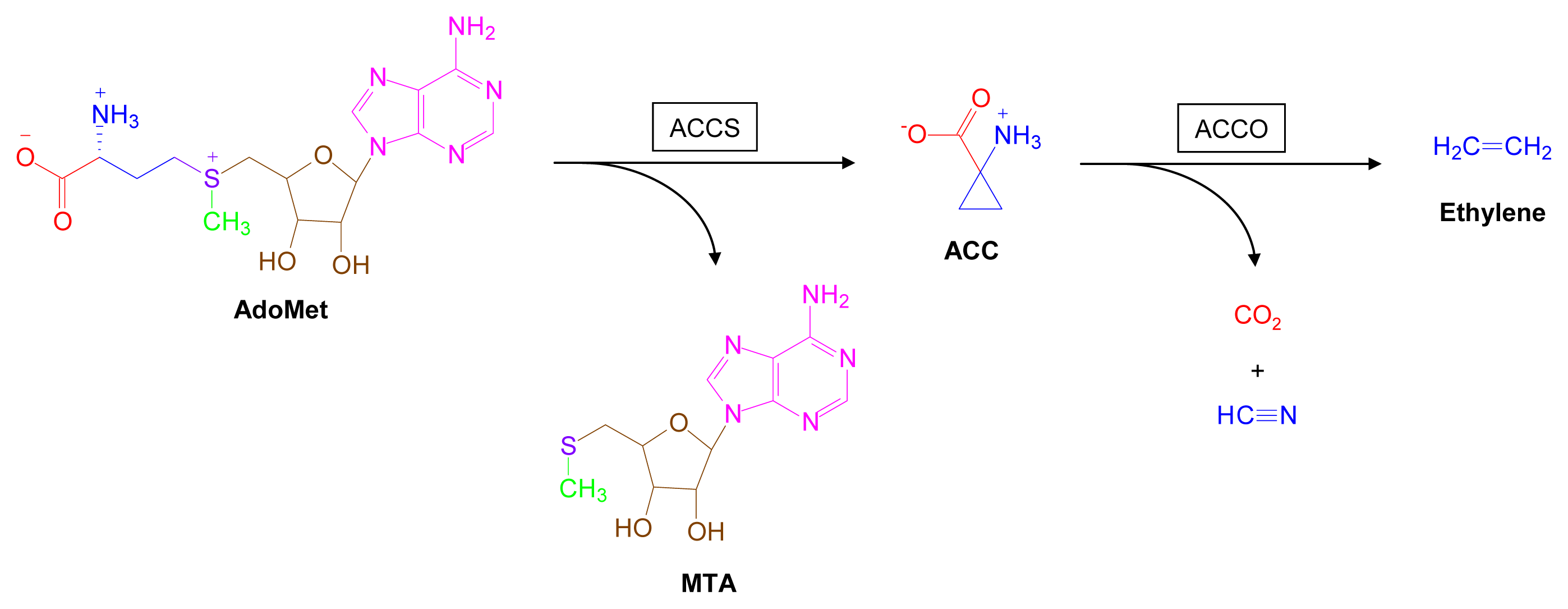
3.3. AdoMet-Dependent Ethylene Biosynthesis [113–117]
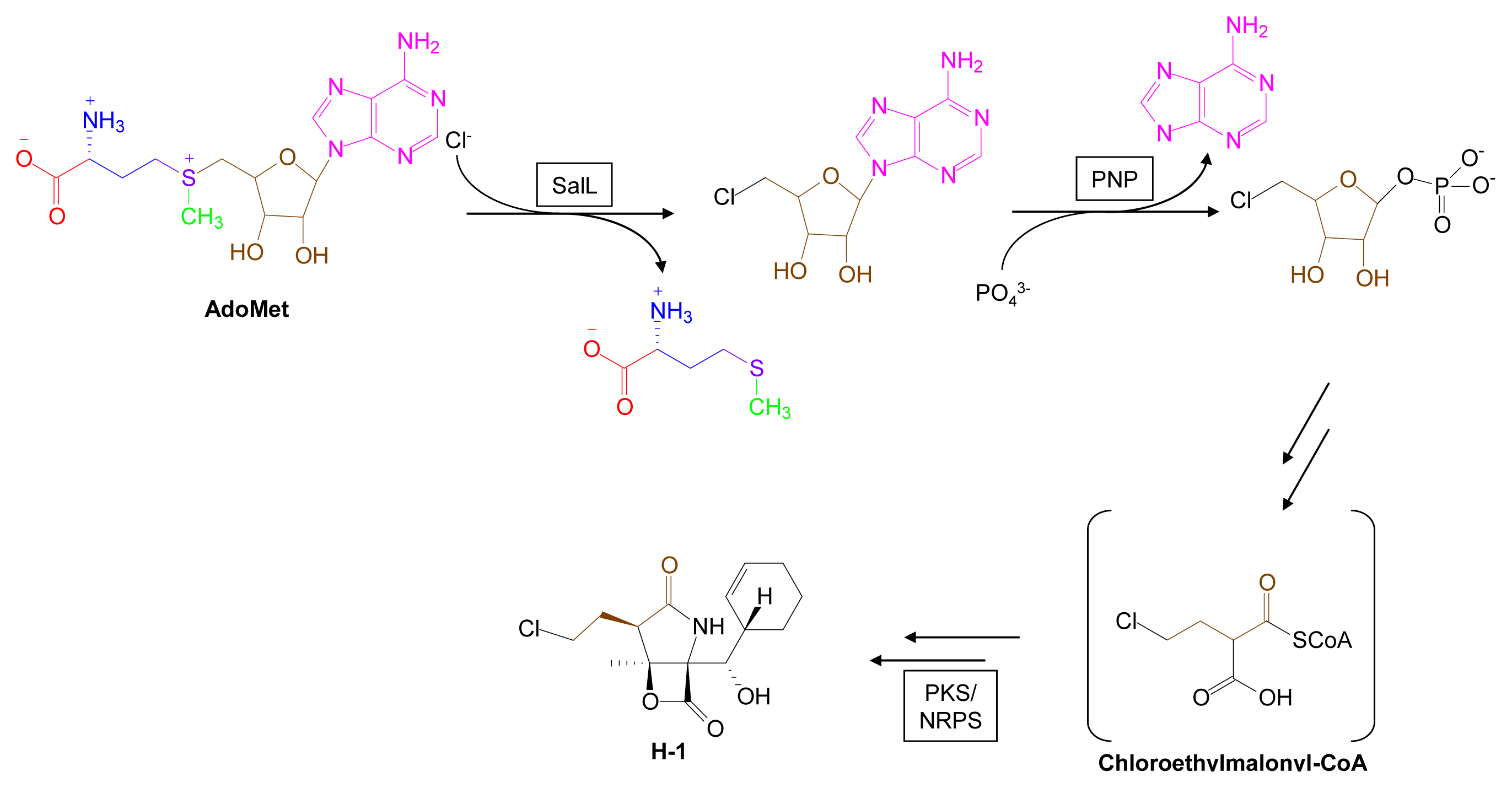
3.4. Biohalogenation Pathways [123–125]
3.5. Radical SAM Pathways [10–12,127–130]
3.6. Quorum Sensing Pathways
3.7. N-Acylhomoserine Lactones as Templates for Anti-Infective Tetramic and Tetronic Acids
3.8. Marine-Derived Quorum Sensing Antagonists
3.9. Unusual Marine Metabolites of AdoMet
4. Conclusions
References and Notes
- Eustaquio, AS; Pojer, F; Noe, JP; Moore, BS. Discovery and Characterization of a Marine Bacterial SAM-Dependent Chlorinase. Nat Chem Biol 2008, 4, 69–74. [Google Scholar]
- Kotb, M; Geller, AM. Methionine Adenosyltransferase-Structure and Function. Pharmacol Ther 1993, 59, 125–143. [Google Scholar]
- Iwig, DF; Booker, SJ. Insight into the Polar Reactivity of the Onium Chalcogen Analogues of S-Adenosyl-l-Methionine. Biochemistry 2004, 43, 13496–13509. [Google Scholar]
- Fontecave, M; Atta, M; Mulliez, E. S-Adenosylmethionine: Nothing Goes to Waste. Trends Biochem Sci 2004, 29, 243–249. [Google Scholar]
- Kagan, RM; Clarke, S. Widespread Occurrence of 3 Sequence Motifs in Diverse S-Adenosylmethionine-Dependent Methyltransferases Suggests a Common Structure for These Enzymes. Arch Biochem Biophys 1994, 310, 417–427. [Google Scholar]
- Deng, H; O'Hagan, D. The Fluorinase, the Chlorinase and the Duf-62 Enzymes. Curr Opin Chem Biol 2008, 12, 582–592. [Google Scholar]
- Hanzelka, BL; Greenberg, EP. Quorum Sensing in Vibrio Fischeri: Evidence That S-Adenosylmethionine Is the Amino Acid Substrate for Autoinducer Synthesis. J Bacteriol 1996, 178, 5291–5294. [Google Scholar]
- More, MI; Finger, LD; Stryker, JL; Fuqua, C; Eberhard, A; Winans, SC. Enzymatic Synthesis of a Quorum-Sensing Autoinducer through Use of Defined Substrates. Science 1996, 272, 1655–1658. [Google Scholar]
- Chen, X; Schauder, S; Potier, N; Van Dorsselaer, A; Pelczer, I; Bassler, BL; Hughson, FM. Structural Identification of a Bacterial Quorum-Sensing Signal Containing Boron. Nature 2002, 415, 545–549. [Google Scholar]
- Sofia, HJ; Chen, G; Hetzler, BG; Reyes-Spindola, JF; Miller, NE. Radical SAM, a Novel Protein Superfamily Linking Unresolved Steps in Familiar Biosynthetic Pathways with Radical Mechanisms: Functional Characterization Using New Analysis and Information Visualization Methods. Nucleic Acids Res 2001, 29, 1097–1106. [Google Scholar]
- Frey, PA; Hegeman, AD; Ruzicka, FJ. The Radical SAM Superfamily. Crit Rev Biochem Mol Biol 2008, 43, 63–88. [Google Scholar]
- Booker, SJ. Anaerobic Functionalization of Unactivated C-H Bonds. Curr Opin Chem Biol 2009, 13, 58–73. [Google Scholar]
- O'Hagan, D; Schaffrath, C; Cobb, SL; Hamilton, JTG; Murphy, CD. Biosynthesis of an Organofluorine Molecule-a Fluorinase Enzyme Has Been Discovered that Catalyses Carbon-Fluorine Bond Formation. Nature 2002, 416, 279–279. [Google Scholar]
- Dong, CJ; Huang, FL; Deng, H; Schaffrath, C; Spencer, JB; O'Hagan, D; Naismith, JH. Crystal Structure and Mechanism of a Bacterial Fluorinating Enzyme. Nature 2004, 427, 561–565. [Google Scholar]
- Eisenberg, MA; Stoner, GL. Biosynthesis of 7,8-Diaminopelargonic Acid, a Biotin Intermediate, from 7-Keto-8-Aminoperlargonic Acid and S-Adenosyl-l-Methionine. J Bacteriol 1971, 108, 1135–1140. [Google Scholar]
- Nishimura, S; Taya, Y; Kuchino, Y; Ohashi, Z. Enzymatic-Synthesis of 3-(3-Amino-3-Carboxypropyl)Uridine in Escherichia-Coli Phenylalanine Transfer-RNA-Transfer of 3-Amino-3-Carboxypropyl Group from S-Adenosylmethionine. Biochem Biophys Res Commun 1974, 57, 702–708. [Google Scholar]
- Iwata-Reuyl, D. An Embarrassment of Riches: The Enzymology of RNA Modification. Curr Opin Chem Biol 2008, 12, 126–133. [Google Scholar]
- van Lanen, SG; Kinzie, SD; Matthieu, S; Link, T; Culp, J; Iwata-Reuyl, D. tRNA Modification by S-Adenosylmethionine: tRNA Ribosyltransferase-Isomerase-Assay Development and Characterization of the Recombinant Enzyme. J Biol Chem 2003, 278, 10491–10499. [Google Scholar]
- Iwata-Reuyl, D. Biosynthesis of the 7-Deazaguanosine Hypermodified Nucleosides of Transfer RNA. Bioorg Chem 2003, 31, 24–43. [Google Scholar]
- Basturea, GN; Rudd, KE; Deutscher, MP. Identification and Characterization of RsmE, the Founding Member of a New RNA Base Methyltransferase Family. RNA-Publ RNA Soc 2006, 12, 426–434. [Google Scholar]
- Poole, A; Penny, D; Sjoberg, BM. Methyl-RNA: An Evolutionary Bridge between RNA and DNA? Chem Biol 2000, 7, R207–R216. [Google Scholar]
- Kerr, SJ; Borek, E. Transfer-RNA Methyltransferases. Adv Enzymol Relat Areas Mol Biol 1972, 36, 1–27. [Google Scholar]
- Colonna, A; Kerr, SJ. The Nucleus as the Site of Transfer-RNA Methylation. J Cell Physiol 1980, 103, 29–33. [Google Scholar]
- Tollervey, D. Small Nucleolar RNAs Guide Ribosomal RNA Methylation. Science 1996, 273, 1056–1057. [Google Scholar]
- Maravic, G; Flogel, M. RNA Methylation and Antibiotic Resistance: An Overview. Period Biol 2004, 106, 135–140. [Google Scholar]
- Sitachitta, N; Marquez, BL; Williamson, RT; Rossi, J; Roberts, MA; Gerwick, WH; Nguyen, VA; Willis, CL. Biosynthetic Pathway and Origin of the Chlorinated Methyl Group in Barbamide and Dechlorobarbamide, Metabolites from the Marine Cyanobacterium Lyngbya Majuscula. Tetrahedron 2000, 56, 9103–9113. [Google Scholar]
- Flatt, PM; O'Connell, SJ; McPhail, KL; Zeller, G; Willis, CL; Sherman, DH; Gerwick, WH. Characterization of the Initial Enzymatic Steps of Barbamide Biosynthesis. J Nat Prod 2006, 69, 938–944. [Google Scholar]
- Orjala, J; Gerwick, WH. Barbamide, a Chlorinated Metabolite with Molluscicidal Activity from the Caribbean Cyanobacterium Lyngbya Majuscula. J Nat Prod 1996, 59, 427–430. [Google Scholar]
- Chang, ZX; Sitachitta, N; Rossi, JV; Roberts, MA; Flatt, PM; Jia, JY; Sherman, DH; Gerwick, WH. Biosynthetic Pathway and Gene Cluster Analysis of Curacin A, an Antitubulin Natural Product from the Tropical Marine Cyanobacterium Lyngbya Majuscula. J Nat Prod 2004, 67, 1356–1367. [Google Scholar]
- Kokke, W; Shoolery, JN; Fenical, W; Djerassi, C. Biosynthetic-Studies of Marine Lipids.4. Mechanism of Side-Chain Alkylation in (E)-24-Propylidenecholesterol by a Chrysophyte Alga. J Org Chem 1984, 49, 3742–3752. [Google Scholar]
- Giner, JL; Djerassi, C. Biosynthetic-Studies of Marine Lipids. 33. Biosynthesis of Dinosterol, Peridinosterol, and Gorgosterol - Unusual Patterns of Bioalkylation in Dinoflagellate Sterols. J Org Chem 1991, 56, 2357–2363. [Google Scholar]
- Kerr, RG; Kerr, SL; Djerassi, C. Biosynthetic-Studies of Marine Lipids. 26. Elucidation of the Biosynthesis of Mutasterol, a Sponge Sterol with a Quaternary Carbon in Its Side-Chain. J Org Chem 1991, 56, 63–66. [Google Scholar]
- Lang, G; Wiese, J; Schmaljohann, R; Imhoff, JF. New Pentaenes from the Sponge-Derived Marine Fungus Penicillium Rugulosum: Structure Determination and Biosynthetic Studies. Tetrahedron 2007, 63, 11844–11849. [Google Scholar]
- Kobayashi, H; Meguro, S; Yoshimoto, T; Namikoshi, M. Absolute Structure, Biosynthesis, and Anti-Microtubule Activity of Phomopsidin, Isolated from a Marine-Derived Fungus Phomopsis Sp. Tetrahedron 2003, 59, 455–459. [Google Scholar]
- Bringmann, G; Lang, G; Gulder, TAM; Tsuruta, H; Muhlbacher, J; Maksimenka, K; Steffens, S; Schaumann, K; Stohr, R; Wiese, J; Imhoff, JF; Perovic-Ottstadt, S; Boreiko, O; Muller, WEG. The First Sorbicillinoid Alkaloids, the Antileukemic Sorbicillactones A and B, from a Sponge-Derived Penicillium Chrysogenum Strain. Tetrahedron 2005, 61, 7252–7265. [Google Scholar]
- Cacciapuoti, G; Porcelli, M; Moretti, MA; Sorrentino, F; Concilio, L; Zappia, V; Liu, ZJ; Tempel, W; Schubot, F; Rose, JP; Wang, BC; Brereton, PS; Jenney, FE; Adams, MWW. The First Agmatine/Cadaverine Aminopropyl Transferase: Biochemical and Structural Characterization of an Enzyme Involved in Polyamine Biosynthesis in the Hyperthermophilic Archaeon Pyrococcus Furiosus. J Bacteriol 2007, 189, 6057–6067. [Google Scholar]
- Kusano, T; Berberich, T; Tateda, C; Takahashi, Y. Polyamines: Essential Factors for Growth and Survival. Planta 2008, 228, 367–381. [Google Scholar]
- Shah, P; Swlatlo, E. A Multifaceted Role for Polyamines in Bacterial Pathogens. Mol Microbiol 2008, 68, 4–16. [Google Scholar]
- Kuehn, GD; Phillips, GC. Role of Polyamines in Apoptosis and Other Recent Advances in Plant Polyamines. Crit Rev Plant Sci 2005, 24, 123–130. [Google Scholar]
- Williams, K. Modulation and Block of Ion Channels: A New Biology of Polyamines. Cell Signal 1997, 9, 1–13. [Google Scholar]
- Igarashi, K; Kashiwagi, K. Polyamines: Mysterious Modulators of Cellular Functions. Biochem Biophys Res Commun 2000, 271, 559–564. [Google Scholar]
- Rhee, HJ; Kim, EJ; Lee, JK. Physiological Polyamines: Simple Primordial Stress Molecules. J Cell Mol Med 2007, 11, 685–703. [Google Scholar]
- Baron, K; Stasolla, C. The Role of Polyamines During in Vivo and in Vitro Development. In Vitro Cell Dev Biol-Plant 2008, 44, 384–395. [Google Scholar]
- Groppa, MD; Benavides, MP. Polyamines and Abiotic Stress: Recent Advances. Amino Acids 2008, 34, 35–45. [Google Scholar]
- Williams, K. Interactions of Polyamines with Ion Channels. Biochem J 1997, 325, 289–297. [Google Scholar]
- Terui, Y; Ohnuma, M; Hiraga, K; Kawashima, E; Oshima, T. Stabilization of Nucleic Acids by Unusual Polyamines Produced by an Extreme Thermophile, Thermus Thermophilus. Biochem J 2005, 388, 427–433. [Google Scholar]
- Matsunaga, S; Sakai, R; Jimbo, M; Kamiya, H. Long-Chain Polyamines (LCPAs) from Marine Sponge: Possible Implication in Spicule Formation. ChemBioChem 2007, 8, 1729–1735. [Google Scholar]
- Kroger, N; Deutzmann, R; Bergsdorf, C; Sumper, M. Species-Specific Polyamines from Diatoms Control Silica Morphology. Proc Natl Acad Sci USA 2000, 97, 14133–14138. [Google Scholar]
- Sumper, M; Lehmann, G. Silica Pattern Formation in Diatoms: Species-Specific Polyamine Biosynthesis. ChemBioChem 2006, 7, 1419–1427. [Google Scholar]
- Sumper, M; Brunner, E. Silica Biomineralisation in Diatoms: The Model Organism Thalassiosira Pseudonana. ChemBioChem 2008, 9, 1187–1194. [Google Scholar]
- Foo, CWP; Huang, J; Kaplan, DL. Lessons from Seashells: Silica Mineralization via Protein Templating. Trends Biotechnol 2004, 22, 577–585. [Google Scholar]
- Ohnuma, M; Terui, Y; Tamakoshi, M; Mitome, H; Niitsu, M; Samejima, K; Kawashima, E; Oshima, T. N-1-Aminopropylagmatine, a New Polyamine Produced as a Key Intermediate in Polyamine Biosynthesis of an Extreme Thermophile, Thermus Thermophilus. J Biol Chem 2005, 280, 30073–30082. [Google Scholar]
- Palagiano, E; Demarino, S; Minale, L; Riccio, R; Zollo, F; Iorizzi, M; Carre, JB; Debitus, C; Lucarain, L; Provost, J. Ptilomycalin-A, Crambescidin-800 and Related New Highly Cytotoxic Guanidine Alkaloids from the Starfishes Fromia-Monilis and Celerina-Heffernani. Tetrahedron 1995, 51, 3675–3682. [Google Scholar]
- Black, GP; Coles, SJ; Hizi, A; Howard-Jones, AG; Hursthouse, MB; McGown, AT; Loya, S; Moore, CG; Murphy, PJ; Smith, NK; Walshe, NDA. Synthesis and Biological Activity of Analogues of Ptilomycalin A. Tetrahedron Lett 2001, 42, 3377–3381. [Google Scholar]
- Lazaro, JEH; Nitcheu, J; Mahmoudi, N; Ibana, JA; Mangalindan, GC; Black, GP; Howard-Jones, AG; Moore, CG; Thomas, DA; Mazier, D; Ireland, CM; Concepcion, GP; Murphy, PJ; Diquet, B. Antimalarial Activity of Crambescidin 800 and Synthetic Analogues against Liver and Blood Stage of Plasmodium Sp. J Antibiot 2006, 59, 583–590. [Google Scholar]
- Kashman, Y; Hirsh, S; McConnell, OJ; Ohtani, I; Kusumi, T; Kakisawa, H. Ptilomycalin-A - a Novel Polycyclic Guanidine Alkaloid of Marine Origin. J Am Chem Soc 1989, 111, 8925–8926. [Google Scholar]
- Gassner, NC; Tamble, CM; Bock, JE; Cotton, N; White, KN; Tenney, K; St Onge, RP; Proctor, MJ; Giaever, G; Nislow, C; Davis, RW; Crews, P; Holman, TR; Lokey, RS. Accelerating the Discovery of Biologically Active Small Molecules Using a High-Throughput Yeast Halo Assay. J Nat Prod 2007, 70, 383–390. [Google Scholar]
- Carter, GT; Rinehart, KL. Acarnidines, Novel Anti-Viral and Anti-Microbial Compounds from Sponge Acarnus-Erithacus-(De Laubenfels). J Am Chem Soc 1978, 100, 4302–4304. [Google Scholar]
- Schmitz, FJ; Hollenbeak, KH; Prasad, RS. Marine Natural-Products - Cytotoxic Spermidine Derivatives from the Soft Coral Sinularia-Brongersmai. Tetrahedron Lett 1979, 3387–3390. [Google Scholar]
- Karigiannis, G; Papaioannou, D. Structure, Biological Activity and Synthesis of Polyamine Analogues and Conjugates. Eur J Org Chem 2000, 1841–1863. [Google Scholar]
- Ushiosata, N; Matsunaga, S; Fusetani, N; Honda, K; Yasumuro, K. Penaramides, Which Inhibit Binding of Omega-Conotoxin Gvia to N-Type Ca2+ Channels, from the Marine Sponge Penares Aff Incrustans. Tetrahedron Lett 1996, 37, 225–228. [Google Scholar]
- Coffey, DS; McDonald, AI; Overman, LE; Rabinowitz, MH; Renhowe, PA. A Practical Entry to the Crambescidin Family of Guanidine Alkaloids. Enantioselective Total Syntheses of Ptilomycalin a, Crambescidin 657 and Its Methyl Ester (Neofolitispates 2), and Crambescidin 800. J Am Chem Soc 2000, 122, 4893–4903. [Google Scholar]
- Tsukamoto, S; Kato, H; Hirota, H; Fusetani, N. Pseudoceratidine: A New Antifouling Spermidine Derivative from the Marine Sponge Pseudoceratina Purpurea. Tetrahedron Lett 1996, 37, 1439–1440. [Google Scholar]
- Buchanan, MS; Carroll, AR; Fechner, GA; Boyle, A; Simpson, MM; Addepalli, R; Avery, VM; Hooper, JNA; Su, N; Chen, HW; Quinn, RJ. Spermatinamine, the First Natural Product Inhibitor of Isoprenylcysteine Carboxyl Methyltransferase, a New Cancer Target. Bioorg Med Chem Lett 2007, 17, 6860–6863. [Google Scholar]
- Koppisch, AT; Browder, CC; Moe, AL; Shelley, JT; Kinke, BA; Hersman, LE; Iyer, S; Ruggiero, CE. Petrobactin is the Primary Siderophore Synthesized by Bacillus Anthracis Str. Sterne under Conditions of Iron Starvation. BioMetals 2005, 18, 577–585. [Google Scholar]
- Sata, NU; Sugano, M; Matsunaga, S; Fusetani, N. Bioactive Marine Metabolites - Part 88 - Sinulamide: An H,K-ATPase Inhibitor from a Soft Coral Sinularia sp. Tetrahedron Lett 1999, 40, 719–722. [Google Scholar]
- Wright, LP; Philips, MR. Caax Modification and Membrane Targeting of Ras. J Lipid Res 2006, 47, 883–891. [Google Scholar]
- Barbeau, K; Zhang, GP; Live, DH; Butler, A. Petrobactin, a Photoreactive Siderophore Produced by the Oil-Degrading Marine Bacterium Marinobacter Hydrocarbonoclasticus. J Am Chem Soc 2002, 124, 378–379. [Google Scholar]
- Koppisch, AT; Dhungana, S; Hill, KK; Boukhalfa, H; Heine, HS; Colip, LA; Romero, RB; Shou, YL; Ticknor, LO; Marrone, BL; Hersman, LE; Iyer, S; Ruggiero, CE. Petrobactin is Produced by Both Pathogenic and Non-Pathogenic Isolates of the Bacillus Cereus Group of Bacteria. BioMetals 2008, 21, 581–589. [Google Scholar]
- Bergeron, RJ; Huang, GF; Smith, RE; Bharti, N; McManis, JS; Butler, A. Total Synthesis and Structure Revision of Petrobactin. Tetrahedron 2003, 59, 2007–2014. [Google Scholar]
- Gardner, RA; Kinkade, R; Wang, CJ; Phanstiel, O. Total Synthesis of Petrobactin and Its Homologues as Potential Growth Stimuli for Marinobacter Hydrocarbonoclasticus, an Oil-Degrading Bacteria. J Org Chem 2004, 69, 3530–3537. [Google Scholar]
- Abergel, RJ; Wilson, MK; Arceneaux, JEL; Hoette, TM; Strong, RK; Byers, BR; Raymond, KN. Anthrax Pathogen Evades the Mammalian Immune System through Stealth Siderophore Production. Proc Natl Acad Sci USA 2006, 103, 18499–18503. [Google Scholar]
- Gardner, RA; Kinkade, R; Wang, CJ; Phanstiel, O. Total Synthesis of Petrobactin and Its Homologues as Potential Growth Stimuli for Marinobacter Hydrocarbonoclasticus, an Oil-Degrading Bacteria. (Correction). J Org Chem 2007, 72, 3158–3158. [Google Scholar]
- Lee, JY; Janes, BK; Passalacqua, KD; Pfleger, BF; Bergman, NH; Liu, HC; Hakansson, K; Somu, RV; Aldrich, CC; Cendrowski, S; Hanna, PC; Sherman, DH. Biosynthetic Analysis of the Petrobactin Siderophore Pathway from Bacillus Anthracis. J Bacteriol 2007, 189, 1698–1710. [Google Scholar]
- Oves-Costales, D; Kadi, N; Fogg, MJ; Song, L; Wilson, KS; Challis, GL. Enzymatic Logic of Anthrax Stealth Siderophore Biosynthesis: AsbA Catalyzes ATP-Dependent Condensation of Citric Acid and Spermidine. J Am Chem Soc 2007, 129, 8416–8417. [Google Scholar]
- Pfleger, BF; Lee, JY; Somu, RV; Aldrich, CC; Hanna, PC; Sherman, DH. Characterization and Analysis of Early Enzymes for Petrobactin Biosynthesis in Bacillus Anthracis. Biochemistry (Mosc) 2007, 46, 4147–4157. [Google Scholar]
- Fox, DT; Hotta, K; Kim, CY; Koppisch, AT. The Missing Link in Petrobactin Biosynthesis: AsbF Encodes a (-)-3-Dehydroshikimate Dehydratase. Biochemistry (Mosc) 2008, 47, 12251–12253. [Google Scholar]
- Koppisch, AT; Hotta, K; Fox, DT; Ruggiero, CE; Kim, CY; Sanchez, T; Iyer, S; Browder, CC; Unkefer, PJ; Unkefer, CJ. Biosynthesis of the 3,4-Dihydroxybenzoate Moieties of Petrobactin by Bacillus Anthracis. J Org Chem 2008, 73, 5759–5765. [Google Scholar]
- Pfleger, BF; Kim, YC; Nusca, TD; Maltseva, N; Lee, JY; Rath, CM; Scaglione, JB; Janes, BK; Anderson, EC; Bergman, NH; Hanna, PC; Joachimiak, A; Sherman, DH. Structural and Functional Analysis of AsbF: Origin of the Stealth 3,4-Dihydroxybenzoic Acid Subunit for Petrobactin Biosynthesis. Proc Natl Acad Sci U S A 2008, 105, 17133–17138. [Google Scholar]
- Lee, J; Sperandio, V; Frantz, DE; Longgood, J; Camilli, A; Phillips, MA; Michael, AJ. An Alternative Polyamine Biosynthetic Pathway Is Widespread in Bacteria and Essential for Biofilm Formation in Vibrio Cholerae. J Biol Chem 2009, 284, 9899–9907. [Google Scholar]
- Oves-Costales, D; Song, L; Challis, GL. Enantioselective Desymmetrisation of Citric Acid Catalysed by the Substrate-Tolerant Petrobactin Biosynthetic Enzyme AsbA. Chem Commun 2009, 1389–1391. [Google Scholar]
- Ponasik, JA; Kassab, DJ; Ganem, B. Synthesis of the Antifouling Polyamine Pseudoceratidine and Its Analogs: Factors Influencing Biocidal Activity. Tetrahedron Lett 1996, 37, 6041–6044. [Google Scholar]
- Ponasik, JA; Conova, S; Kinghorn, D; Kinney, WA; Rittschof, D; Ganem, B. Pseudoceratidine, a Marine Natural Product with Antifouling Activity: Synthetic and Biological Studies. Tetrahedron 1998, 54, 6977–6986. [Google Scholar]
- Katz, JE; Dlakic, M; Clarke, S. Automated Identification of Putative Methyltransferases from Genomic Open Reading Frames. Mol Cell Proteomics 2003, 2, 525–540. [Google Scholar]
- Fujioka, M. Mammalian Small Molecule Methyltransferases - Their Structural and Functional Features. Int J Biochem 1992, 24, 1917–1924. [Google Scholar]
- Cheng, XD; Roberts, RJ. AdoMet-Dependent Methylation, DNA Methyltransferases and Base Flipping. Nucleic Acids Res 2001, 29, 3784–3795. [Google Scholar]
- Yeates, TO. Structures of Set Domain Proteins: Protein Lysine Methyltransferases Make Their Mark. Cell 2002, 111, 5–7. [Google Scholar]
- Romano, JD; Michaelis, S. Topological and Mutational Analysis of Saccharomyces Cerevisiae Ste14p, Founding Member of the Isoprenylcysteine Carboxyl Methyltransferase Family. Mol Biol Cell 2001, 12, 1957–1971. [Google Scholar]
- Farooqi, AHA; Shukla, YN; Shukla, A; Bhakuni, DS. Cytokinins from Marine Organisms. Phytochemistry 1990, 29, 2061–2063. [Google Scholar]
- Ashour, M; Edrada-Ebel, R; Ebel, R; Wray, V; Van Soest, RWM; Proksch, P. New Purine Derivatives from the Marine Sponge Petrosia Nigricans. Nat Prod Commun 2008, 3, 1889–1894. [Google Scholar]
- Lindsay, BS; Battershill, CN; Copp, BR. 1,3-Dimethylguanine, a New Purine from the New Zealand Ascidian Botrylloides Leachi. J Nat Prod 1999, 62, 638–639. [Google Scholar]
- Tasdemir, D; Mangalindan, GC; Concepcion, GP; Harper, MK; Irelanda, CM. 3,7-Dimethylguanine, a New Purine from a Philippine Sponge Zyzzya Fuliginosa. Chem Pharm Bull (Tokyo) 2001, 49, 1628–1630. [Google Scholar]
- Perry, NB; Blunt, JW; Munro, MHG. 1,3,7-Trimethylguanine from the Sponge Latrunculia-Brevis. J Nat Prod 1987, 50, 307–308. [Google Scholar]
- Berry, Y; Bremner, JB; Davis, A; Samosorn, S. Isolation and NMR Spectroscopic Clarification of the Alkaloid 1,3,7-Trimethylguanine from the Ascidian Eudistoma Maculosum. Nat Prod Res 2006, 20, 479–483. [Google Scholar]
- Pearce, AN; Babcock, RC; Lambert, G; Copp, BR. N-2,N-2,7-Trimethylguanine, a New Trimethylated Guanine Natural Product from the New Zealand Ascidian, Lissoclinum Notti. Nat Prod Lett 2001, 15, 237–241. [Google Scholar]
- Lindsay, BS; Almeida, AMP; Smith, CJ; Berlinck, RGS; Da Rocha, RM; Ireland, CM. 6-Methoxy-7-Methyl-8-Oxoguanine, an Unusual Purine from the Ascidian Symplegma Rubra. J Nat Prod 1999, 62, 1573–1575. [Google Scholar]
- Chehade, CC; Dias, RLA; Berlinck, RGS; Ferreira, AG; Costa, LV; Rangel, M; Malpezzi, ELA; Defreitas, JC; Hajdu, E. 1,3-Dimethylisoguanine, a New Purine from the Marine Sponge Amphimedon Viridis. J Nat Prod 1997, 60, 729–731. [Google Scholar]
- Cafieri, F; Fattorusso, E; Mangoni, A; Taglialatelascafati, O. Longamide and 3,7-Dimethylisoguanine, 2 Novel Alkaloids from the Marine Sponge Agelas-Longissima. Tetrahedron Lett 1995, 36, 7893–7896. [Google Scholar]
- Copp, BR; Wassvik, CM; Lambert, G; Page, MJ. Isolation and Characterization of the New Purine 1,3,7-Trimethylisoguanine from the New Zealand Ascidian Pseudodistoma Cereum. J Nat Prod 2000, 63, 1168–1169. [Google Scholar]
- Appleton, DR; Page, MJ; Lambert, G; Copp, BR. 1,3-Dimethyl-8-Oxoisoguanine, a New Purine from the New Zealand Ascidian Pseudodistoma Cereum. Nat Prod Res 2004, 18, 39–42. [Google Scholar]
- Derosa, S; Destefano, S; Puliti, R; Mattia, CA; Mazzarella, L. Isolation and X-Ray Crystal-Structure of a Derivative of 2,6-Diaminopurine from a Sea-Anemone. J Nat Prod 1987, 50, 876–880. [Google Scholar]
- Cimino, G; Degiulio, A; Derosa, S; Destefano, S; Puliti, R; Mattia, CA; Mazzarella, L. Isolation and X-Ray Crystal-Structure of a Novel 8-Oxopurine Compound from a Marine Sponge. J Nat Prod 1985, 48, 523–528. [Google Scholar]
- Cooper, RA; Defreitas, JC; Porreca, F; Eisenhour, CM; Lukas, R; Huxtable, RJ. The Sea-Anemone Purine, Caissarone - Adenosine Receptor Antagonism. Toxicon 1995, 33, 1025–1031. [Google Scholar]
- Yagi, H; Matsunaga, S; Fusetani, N. Isolation of 1-Methylherbipoline, a Purine Base, from a Marine Sponge, Jaspis Sp. J Nat Prod 1994, 57, 837–838. [Google Scholar]
- Bourguet-Kondracki, ML; Martin, MT; Vacelet, J; Guyot, M. Mucronatine, a New N-Methyl Purine from the French Mediterranean Marine Sponge Stryphnus Mucronatus. Tetrahedron Lett 2001, 42, 7257–7259. [Google Scholar]
- Rosemeyer, H. The Chemodiversity of Purine as a Constituent of Natural Products. Chem Biodivers 2004, 1, 361–401. [Google Scholar]
- Kerr, RG; Baker, BJ. Marine Sterols. Nat Prod Rep 1991, 8, 465–497. [Google Scholar]
- Giner, JL; Faraldos, JA; Boyer, GL. Novel Sterols of the Toxic Dinoflagellate Karenia Brevis (Dinophyceae): A Defensive Function for Unusual Marine Sterols? J Phycol 2003, 39, 315–319. [Google Scholar]
- Nechev, J; Christie, WW; Robaina, R; De Diego, F; Popov, S; Stefanov, K. Chemical Composition of the Sponge Hymeniacidon Sanguinea from the Canary Islands. Comp Biochem Physiol A: Mol Integr Physiol 2004, 137, 365–374. [Google Scholar]
- Djerassi, C; Lam, WK. Phospholipid Studies of Marine Organisms .25. Sponge Phospholipids. Acc Chem Res 1991, 24, 69–75. [Google Scholar]
- Giner, JL. Biosynthesis of Marine Sterol Side-Chains. Chem Rev 1993, 93, 1735–1752. [Google Scholar]
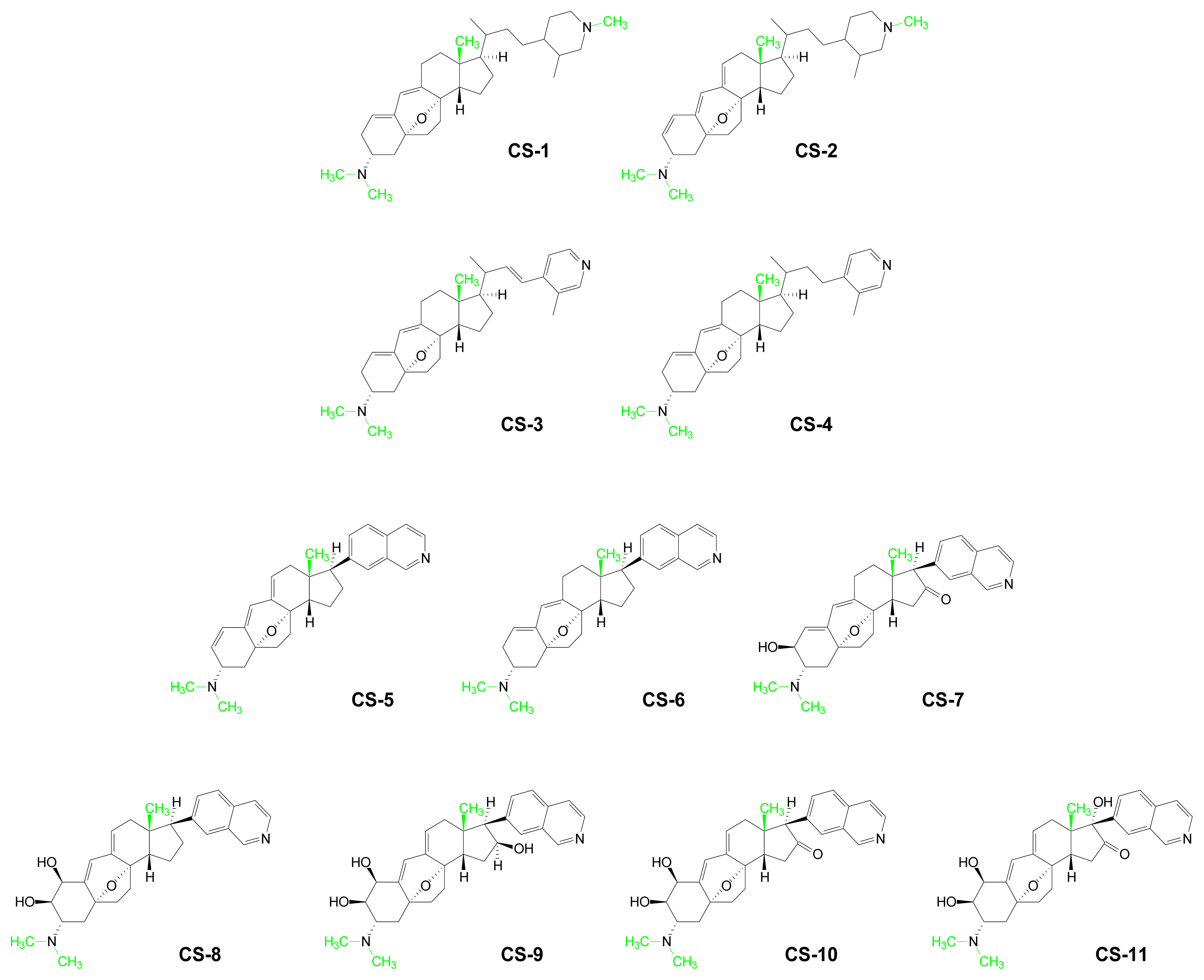
- Aoki, S; Watanabe, Y; Tanabe, D; Arai, M; Suna, H; Miyamoto, K; Tsujibo, H; Tsujikawa, K; Yamamoto, H; Kobayashi, M. Structure-Activity Relationship and Biological Property of Cortistatins, Anti-Angiogenic Spongean Steroidal Alkaloids. Bioorg Med Chem 2007, 15, 6758–6762. [Google Scholar]
- Guo, HW; Ecker, JR. The Ethylene Signaling Pathway: New Insights. Curr Opin Plant Biol 2004, 7, 40–49. [Google Scholar]
- Pandey, S; Ranade, SA; Nagar, PK; Kumar, N. Role of Polyamines and Ethylene as Modulators of Plant Senescence. J Biosci (Bangalore) 2000, 25, 291–299. [Google Scholar]
- Alonso, JM; Stepanova, AN. The Ethylene Signaling Pathway. Science 2004, 306, 1513–1515. [Google Scholar]
- Bleecker, AB; Kende, H. Ethylene: A Gaseous Signal Molecule in Plants. Ann Rev Cell Dev Biol 2000, 16, 1–18. [Google Scholar]
- Roje, S. S-Adenosyl-l-Methionine: Beyond the Universal Methyl Group Donor. Phytochem 2006, 67, 1686–1698. [Google Scholar]
- Wang, KLC; Li, H; Ecker, JR. Ethylene Biosynthesis and Signaling Networks. Plant Cell 2002, 14, S131–S151. [Google Scholar]
- Plettner, I; Steinke, M; Malin, G. Ethene (Ethylene) Production in the Marine Macroalga Ulva (Enteromorpha) Intestinalis L. (Chlorophyta, Ulvophyceae): Effect of Light-Stress and Co-Production with Dimethyl Sulphide. Plant Cell Environ 2005, 28, 1136–1145. [Google Scholar]
- Watanabe, T; Kondo, N. Ethylene Evolution in Marine-Algae and a Proteinaceous Inhibitor of Ethylene Biosynthesis from Red Alga. Plant Cell Physiol 1976, 17, 1159–1166. [Google Scholar]
- Watanabe, T; Kondo, N; Fujii, T; Noguchi, T. Affinity Chromatography of an Ethylene-Synthesizing Enzyme from Red Alga Porphyra-Tenera on an Immobilized Inhibitor of Ethylene Evolution. Plant Cell Physiol 1977, 18, 387–392. [Google Scholar]
- Krasko, A; Schroder, HC; Perovic, S; Steffen, R; Kruse, M; Reichert, W; Muller, IM; Muller, WEG. Ethylene Modulates Gene Expression in Cells of the Marine Sponge Suberites Domuncula and Reduces the Degree of Apoptosis. J Biol Chem 1999, 274, 31524–31530. [Google Scholar]
- O'Hagan, D. Recent Developments on the Fluorinase from Streptomyces Cattleya. J Fluorine Chem 2006, 127, 1479–1483. [Google Scholar]
- Fujimori, DG; Walsh, CT. What's New in Enzymatic Halogenations. Curr Opin Chem Biol 2007, 11, 553–560. [Google Scholar]
- Blasiak, LC; Drennan, CL. Structural Perspective on Enzymatic Halogenation. Acc Chem Res 2009, 42, 147–155. [Google Scholar]
- Deng, H; Cobb, SL; McEwan, AR; McGlinchey, RP; Naismith, JH; O'Hagan, D; Robinson, DA; Spencer, JB. The Fluorinase from Streptomyces Cattleya is also a Chlorinase. Angew Chem Int Ed (Engl) 2006, 45, 759–762. [Google Scholar]
- Layer, G; Heinz, DW; Jahn, D; Schubert, WD. Structure and Function of Radical SAM Enzymes. Curr Opin Chem Biol 2004, 8, 468–476. [Google Scholar]
- Wang, SC; Frey, PA. S-Adenosylmethionine as an Oxidant: The Radical SAM Superfamily. Trends Biochem Sci 2007, 32, 101–110. [Google Scholar]
- Booker, SJ; Cicchillo, RM; Grove, TL. Self-Sacrifice in Radical S-Adenosylmethionine Proteins. Curr Opin Chem Biol 2007, 11, 543–552. [Google Scholar]
- Duschene, KS; Veneziano, SE; Silver, SC; Broderick, JB. Control of Radical Chemistry in the AdoMet Radical Enzymes. Curr Opin Chem Biol 2009, 13, 74–83. [Google Scholar]
- Chew, AGM; Frigaard, NU; Bryant, DA. Bacteriochlorophyllide c C-8(2) and C-12(1) Methyltransferases Are Essential for Adaptation to Low Light in Chlorobaculum Tepidum. J Bacteriol 2007, 189, 6176–6184. [Google Scholar]
- Huster, MS; Smith, KM. Biosynthetic Studies of Substituent Homologation in Bacteriochlorophylls c and d. Biochemistry (Mosc) 1990, 29, 4348–4355. [Google Scholar]
- Swingley, WD; Chen, M; Cheung, PC; Conrad, AL; Dejesa, LC; Hao, J; Honchak, BM; Karbach, LE; Kurdoglu, A; Lahiri, S; Mastrian, SD; Miyashita, H; Page, L; Ramakrishna, P; Satoh, S; Sattley, WM; Shimada, Y; Taylor, HL; Tomo, T; Tsuchiya, T; Wang, ZT; Raymond, J; Mimuro, M; Blankenship, RE; Touchman, JW. Niche Adaptation and Genome Expansion in the Chlorophyll d-Producing Cyanobacterium Acarylochloris Marina. Proc Natl Acad Sci USA 2008, 105, 2005–2010. [Google Scholar]
- Fuqua, WC; Winans, SC; Greenberg, EP. Quorum Sensing in Bacteria - the LuxR-LuxI Family of Cell Density-Responsive Transcriptional Regulators. JBacteriol 1994, 176, 269–275. [Google Scholar]
- Hastings, JW; Greenberg, EP. Quorum Sensing: The Explanation of a Curious Phenomenon Reveals a Common Characteristic of Bacteria. J Bacteriol 1999, 181, 2667–2668. [Google Scholar]
- Fuqua, C; Greenberg, EP. Self Perception in Bacteria: Quorum Sensing with Acylated Homoserine Lactones. Curr Opin Microbiol 1998, 1, 183–189. [Google Scholar]
- Fuqua, C; Greenberg, EP. Listening in on Bacteria: Acyl-Homoserine Lactone Signalling. Nat Rev Mol Cell Biol 2002, 3, 685–695. [Google Scholar]
- Schaefer, AL; Val, DL; Hanzelka, BL; Cronan, JE; Greenberg, EP. Generation of Cell-to-Cell Signals in Quorum Sensing: Acyl Homoserine Lactone Synthase Activity of a Purified Vibrio Fescheri LuxI Protein. Proc Natl Acad Sci USA 1996, 93, 9505–9509. [Google Scholar]
- Milton, DL. Quorum Sensing in Vibrios: Complexity for Diversification. Int J Med Microbiol 2006, 296, 61–71. [Google Scholar]
- Krick, A; Kehraus, S; Eberl, L; Riedel, K; Anke, H; Kaesler, I; Graeber, I; Szewzyk, U; Konig, GM. A Marine Mesorhizobium Sp Produces Structurally Novel Long-Chain N-Acyl-l-Homoserine Lactones. Appl Environ Microbiol 2007, 73, 3587–3594. [Google Scholar]
- Brinkhoff, T; Giebel, HA; Simon, M. Diversity, Ecology, and Genomics of the Roseobacter Clade: A Short Overview. Arch Microbiol 2008, 189, 531–539. [Google Scholar]
- Wagner-Dobler, I; Biebl, H. Environmental Biology of the Marine Roseobacter Lineage. Annu Rev Microbiol 2006, 60, 255–280. [Google Scholar]
- Wagner-Dobler, I; Thiel, V; Eberl, L; Allgaier, M; Bodor, A; Meyer, S; Ebner, S; Hennig, A; Pukall, R; Schulz, S. Discovery of Complex Mixtures of Novel Long-Chain Quorum Sensing Signals in Free-Living and Host-Associated Marine Alphaproteobacteria. ChemBioChem 2005, 6, 2195–2206. [Google Scholar]
- Buchan, A; Gonzalez, JM; Moran, MA. Overview of the Marine Roseobacter Lineage. Appl Environ Microbiol 2005, 71, 5665–5677. [Google Scholar]
- Rickard, AH; Palmer, RJ; Blehert, DS; Campagna, SR; Semmelhack, MF; Egland, PG; Bassler, BL; Kolenbrander, PE. Autoinducer 2: A Concentration-Dependent Signal for Mutualistic Bacterial Biofilm Growth. Mol Microbiol 2006, 60, 1446–1456. [Google Scholar]
- Sturme, MHJ; Kleerebezem, M; Nakayama, J; Akkermans, ADL; Vaughan, EE; de Vos, WM. Cell to Cell Communication by Autoinducing Peptides in Gram-Positive Bacteria. Anton Leeuw Int J G Mol Microbiol 2002, 81, 233–243. [Google Scholar]
- Federle, MJ; Bassler, BL. Interspecies Communication in Bacteria. J Clin Invest 2003, 112, 1291–1299. [Google Scholar]
- Uroz, S; Dessaux, Y; Oger, P. Quorum Sensing and Quorum Quenching: The Yin and Yang of Bacterial Communication. ChemBioChem 2009, 10, 205–216. [Google Scholar]
- Kaufmann, GF; Sartorio, R; Lee, SH; Rogers, CJ; Meijler, MM; Moss, JA; Clapham, B; Brogan, AP; Dickerson, TJ; Janda, KD. Revisiting Quorum Sensing: Discovery of Additional Chemical and Biological Functions for 3-Oxo-N-Acylhomoserine Lactones. Proc Natl Acad Sci USA 2005, 102, 309–314. [Google Scholar]
- Roggo, BE; Petersen, F; Delmendo, R; Jenny, HB; Peter, HH; Roesel, J. 3-Alkanoyl-5-Hydroxymethyl Tetronic Acid Homologs and Resistomycin - New Inhibitors of HIV-1 Protease. 1. Fermentation, Isolation and Biological-Activity. J Antibiot 1994, 47, 136–142. [Google Scholar]
- Schobert, R. Domino Syntheses of Bioactive Tetronic and Tetramic Acids. Naturwissenschaften 2007, 94, 1–11. [Google Scholar]
- Roggo, BE; Hug, P; Moss, S; Raschdorf, F; Peter, HH. 3-Alkanoyl-5-Hydroxymethyl- Tetronic Acid Homologs - New Inhibitors of HIV-1 Protease. 2. Structure Determination. J Antibiot 1994, 47, 143–147. [Google Scholar]
- Denys, R; Wright, AD; Konig, GM; Sticher, O. New Halogenated Furanones from the Marine Alga Delisea-Pulchra (cf. Fimbriata). Tetrahedron 1993, 49, 11213–11220. [Google Scholar]
- Persson, T; Givskov, M; Nielsen, J. Quorum Sensing Inhibition: Targeting Chemical Communication in Gram-Negative Bacteria. Curr Med Chem 2005, 12, 3103–3115. [Google Scholar]
- Givskov, M; Denys, R; Manefield, M; Gram, L; Maximilien, R; Eberl, L; Molin, S; Steinberg, PD; Kjelleberg, S. Eukaryotic Interference with Homoserine Lactone-Mediated Prokaryotic Signaling. J Bacteriol 1996, 178, 6618–6622. [Google Scholar]
- Manefield, M; De Nys, R; Kumar, N; Read, R; Givskov, M; Steinberg, P; Kjelleberg, SA. Evidence that Halogenated Furanones from Delisea Pulchra Inhibit Acylated Homoserine Lactone (AHL)-Mediated Gene Expression by Displacing the AHL Signal from Its Receptor Protein. Microbiol-UK 1999, 145, 283–291. [Google Scholar]
- Williams, DE; Lassota, P; Andersen, RJ. Motuporamines A-C, Cytotoxic Alkaloids Isolated from the Marine Sponge Xestospongia Exigua (Kirkpatrick). J Org Chem 1998, 63, 4838–4841. [Google Scholar]
- Williams, DE; Craig, KS; Patrick, B; McHardy, LM; Van Soest, R; Roberge, M; Andersen, RJ. Motuporamines, Anti-Invasion and Anti-Angiogenic Alkaloids from the Marine Sponge Xestospongia Exigua (Kirkpatrick): Isolation, Structure Elucidation, Analogue Synthesis, and Conformational Analysis. J Org Chem 2002, 67, 245–258. [Google Scholar]
- Roskelley, CD; Williams, DE; McHardy, LM; Leong, KG; Troussard, A; Karsan, A; Andersen, RJ; Dedhar, S; Roberge, M. Inhibition of Tumor Cell Invasion and Angiogenesis by Motuporamines. Cancer Res 2001, 61, 6788–6794. [Google Scholar]
- To, KCW; Loh, KT; Roskelley, CD; Andersen, RJ; O'Connor, TP. The Anti-Invasive Compound Motuporamine C is a Robust Stimulator of Neuronal Growth Cone Collapse. Neurosci 2006, 139, 1263–1274. [Google Scholar]
- To, KCW; Church, J; O'Connor, TP. Growth Cone Collapse Stimulated by both Calpain- and Rho-Mediated Pathways. Neurosci 2008, 153, 645–653. [Google Scholar]
- Luibrand, RT; Erdman, TR; Vollmer, JJ; Scheuer, PJ; Finer, J; Clardy, J. Ilimaquinone, a Sesquiterpenoid Quinone from a Marine Sponge. Tetrahedron 1979, 35, 609–612. [Google Scholar]
- Takizawa, PA; Yucel, JK; Veit, B; Faulkner, DJ; Deerinck, T; Soto, G; Ellisman, M; Malhotra, V. Complete Vesiculation of Golgi Membranes and Inhibition of Protein-Transport by a Novel Sea Sponge Metabolite, Ilimaquinone. Cell 1993, 73, 1079–1090. [Google Scholar]
- Dinter, A; Berger, EG. Golgi-Disturbing Agents. Histochem Cell Biol 1998, 109, 571–590. [Google Scholar]
- Mayer, TU. Chemical Genetics: Tailoring Tools for Cell Biology. Trends Cell Biol 2003, 13, 270–277. [Google Scholar]
- Radeke, HS; Digits, CA; Casaubon, RL; Snapper, ML. Interactions of (-)-Ilimaquinone with Methylation Enzymes: Implications for Vesicular-Mediated Secretion. Chem Biol 1999, 6, 639–647. [Google Scholar]
- Moore, KS; Wehrli, S; Roder, H; Rogers, M; Forrest, JN; McCrimmon, D; Zasloff, M. Squalamine - an Aminosterol Antibiotic from the Shark. Proc Natl Acad Sci USA 1993, 90, 1354–1358. [Google Scholar]
- Rao, MN; Shinnar, AE; Noecker, LA; Chao, TL; Feibush, B; Snyder, B; Sharkansky, I; Sarkahian, A; Zhang, XH; Jones, SR; Kinney, WA; Zasloff, M. Aminosterols from the Dogfish Shark Squalus-Acanthias. J Nat Prod 2000, 63, 631–635. [Google Scholar]
- Singh, R; Sharma, M; Joshi, P; Rawat, DS. Clinical Status of Anti-Cancer Agents Derived from Marine Sources. Anti-Cancer Agents Med Chem 2008, 8, 603–617. [Google Scholar]
- Pontius, A; Krick, A; Mesry, R; Kehraus, S; Foegen, SE; Muller, M; Klimo, K; Gerhauser, C; Konig, GM. Monodictyochromes A and B, Dimeric Xanthone Derivatives from the Marine Algicolous Fungus Monodictys Putredinis. J Nat Prod 2008, 71, 1793–1799. [Google Scholar]
- Marquez, BL; Watts, KS; Yokochi, A; Roberts, MA; Verdier-Pinard, P; Jimenez, JI; Hamel, E; Scheuer, PJ; Gerwick, WH. Structure and Absolute Stereochemistry of Hectochlorin, a Potent Stimulator of Actin Assembly. J Nat Prod 2002, 65, 866–871. [Google Scholar]
- Suntornchashwej, S; Chaichit, N; Isobe, M; Suwanborirux, K. Hectochlorin and Morpholine Derivatives from the Thai Sea Hare, Bursatella Leachii. J Nat Prod 2005, 68, 951–955. [Google Scholar]
- Ramaswamy, AV; Sorrels, CM; Gerwick, WH. Cloning and Biochemical Characterization of the Hectochlorin Biosynthetic Gene Cluster from the Marine Cyanobacterium Lyngbya Majuscula. J Nat Prod 2007, 70, 1977–1986. [Google Scholar]
- Edwards, DJ; Marquez, BL; Nogle, LM; McPhail, K; Goeger, DE; Roberts, MA; Gerwick, WH. Structure and Biosynthesis of the Jamaicamides, New Mixed Polyketide-Peptide Neurotoxins from the Marine Cyanobacterium Lyngbya Majuscula. Chem Biol 2004, 11, 817–833. [Google Scholar]
- Kehraus, S; Gorzalka, S; Hallmen, C; Iqbal, J; Muller, CE; Wright, AD; Wiese, M; Konig, GM. Novel Amino Acid Derived Natural Products from the Ascidian Atriolum Robustum: Identification and Pharmacological Characterization of a Unique Adenosine Derivative. J Med Chem 2004, 47, 2243–2255. [Google Scholar]
- Schlenk, F. Methylthioadenosine. Adv Enzymol Relat Areas Mol Biol 1983, 54, 195–265. [Google Scholar]
- Grillo, MA; Colombatto, S. S-Adenosylmethionine and Its Products. Amino Acids 2008, 34, 187–193. [Google Scholar]
- Kamatani, N; Kubota, M; Willis, EH; Frincke, LA; Carson, DA. 5'-Methylthioadenosine Is the Major Source of Adenine in Human-Cells. Adv Exp Med Biol 1984, 165, 83–88. [Google Scholar]
- Walker, J; Barrett, J. Parasite Sulphur Amino Acid Metabolism. Int J Parasitol 1997, 27, 883–897. [Google Scholar]
- Sekowska, A; Kung, HF; Danchin, A. Sulfur Metabolism in Escherichia coli and Related Bacteria: Facts and Fiction. J Mol Microbiol Biotechnol 2000, 2, 145–177. [Google Scholar]
- Pirkov, I; Norbeck, J; Gustafsson, L; Albers, E. A Complete Inventory of All Enzymes in the Eukaryotic Methionine Salvage Pathway. FEBS J 2008, 275, 4111–4120. [Google Scholar]
- Gibbs, NK; Tye, J; Norval, M. Recent Advances in Urocanic Acid Photochemistry, Photobiology and Photoimmunology. Photochem Photobiol Sci 2008, 7, 655–667. [Google Scholar]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | AdoMet Metabolite | Marine Source | Reference |
|---|---|---|---|
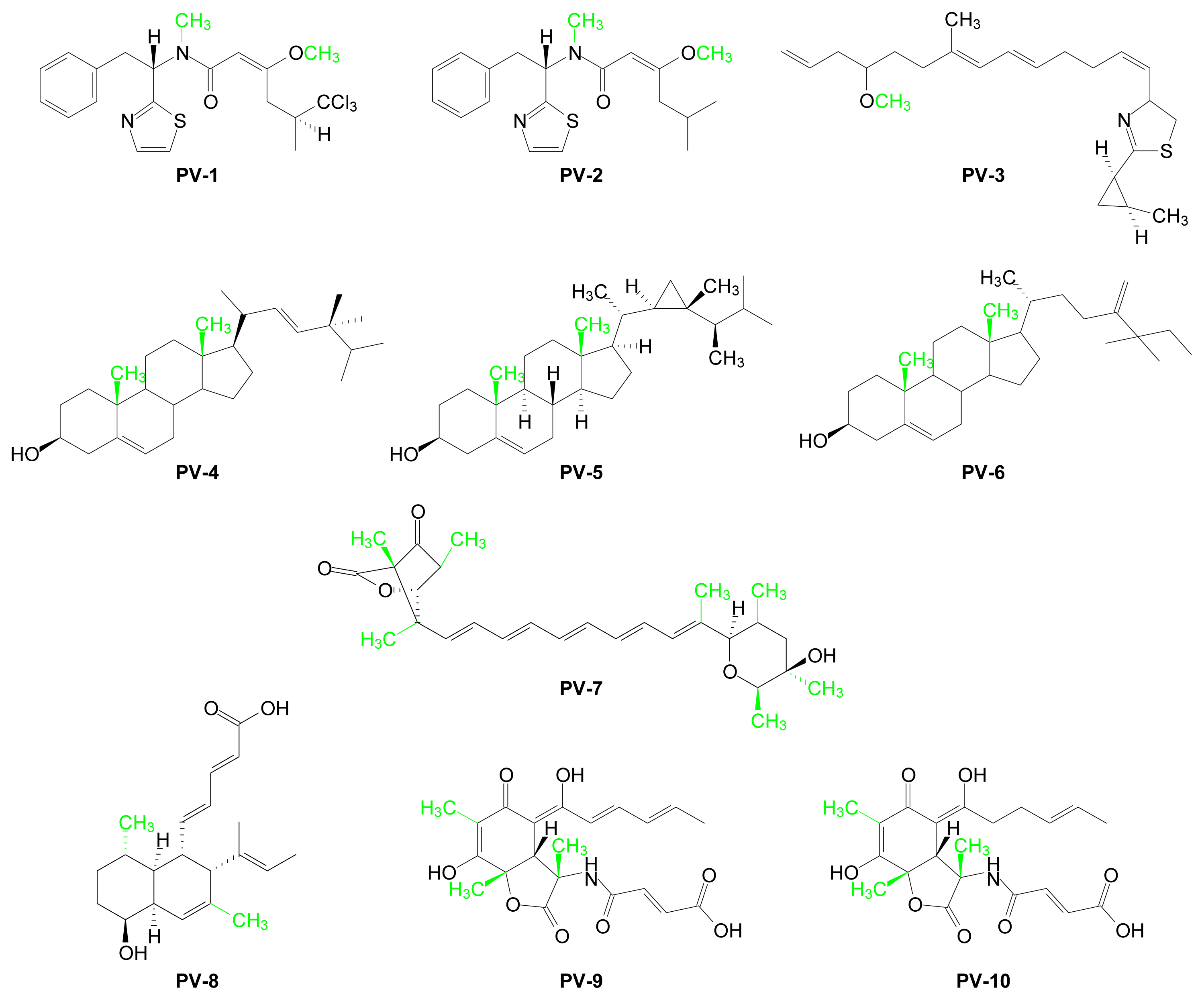
| PV-1 | Barbamide | Lyngbya majuscula (cyanobacterium) | [26–28] |
| PV-2 | Dechlorobarbaramide | Lyngbya majuscula (cyanobacterium) | [26] |
| PV-3 | Curacin A | Lyngbya majuscula (cyanobacterium) | [29] |
| PV-4 | Brassicasterol | Bugula neritina (bryozoan) | [30] |
| PV-5 | Gorgosterol | Isis hippuris (coral) | [31] |
| PV-6 | Mutasterol | Xestspongia muta (sponge) | [32] |
| PV-7 | Prugosene A1 | Penicillium rugulosum (sponge-derived fungus) | [33] |
| PV-8 | Phomopsidin | Phomopsis sp (fungus) | [34] |
| PV-9 | Sorbicillactone A | Penicillium chrysogenum (sponge) | [35] |
| PV-10 | Sorbicillactone B | Penicillium chrysogenum (sponge) | [35] |
| Compound | Polyamine | Structure |
|---|---|---|
| PA-1 | Diaminopropane | H2N(CH2)3NH2 |
| PA-2 | Putrescine | H2N(CH2)4NH2 |
| PA-3 | Cadaverine | H2N(CH2)5NH2 |
| PA-4 | Norspermidine | H2N(CH2)3NH(CH2)3NH2 |
| PA-5 | Spermidine | H2N(CH2)3NH(CH2)4NH2 |
| PA-6 | Homospermidine | H2N(CH2)4NH(CH2)4NH2 |
| PA-7 | Thermine | H2N(CH2)3NH(CH2)3NH (CH2)3NH2 |
| PA-8 | Spermine | H2N(CH2)3NH(CH2)4NH (CH2)3NH2 |
| PA-9 | Thermospermine | H2N(CH2)3NH(CH2)3NH(CH2)4 NH2 |
| PA-10 | Homospermine | H2N(CH2)3NH(CH2)4NH(CH2)4 NH2 |
| PA-11 | Caldopentamine | H2N(CH2)3NH(CH2)3NH(CH2)3NH (CH2)3NH2 |
| PA-12 | Thermopentamine | H2N(CH2)3NH(CH2)3NH(CH2)4NH (CH2)3NH2 |
| PA-13 | Homocaldopentamine | H2N(CH2)3NH(CH)3NH(CH2)3NH(CH2)3NH(CH2)4NH2 |
| PA-14 | Caldohexamine | H2N(CH2)3NH(CH2)3NH(CH2)3NH(CH2)3NH (CH2)3NH2 |
| PA-15 | Homocaldohexamine | H2N(CH2)3NH(CH2)3NH(CH2)3NH(CH2)3NH(CH2)4NH2 |
| Compound | Polyamine Conjugate | Marine Source | Reference |
|---|---|---|---|
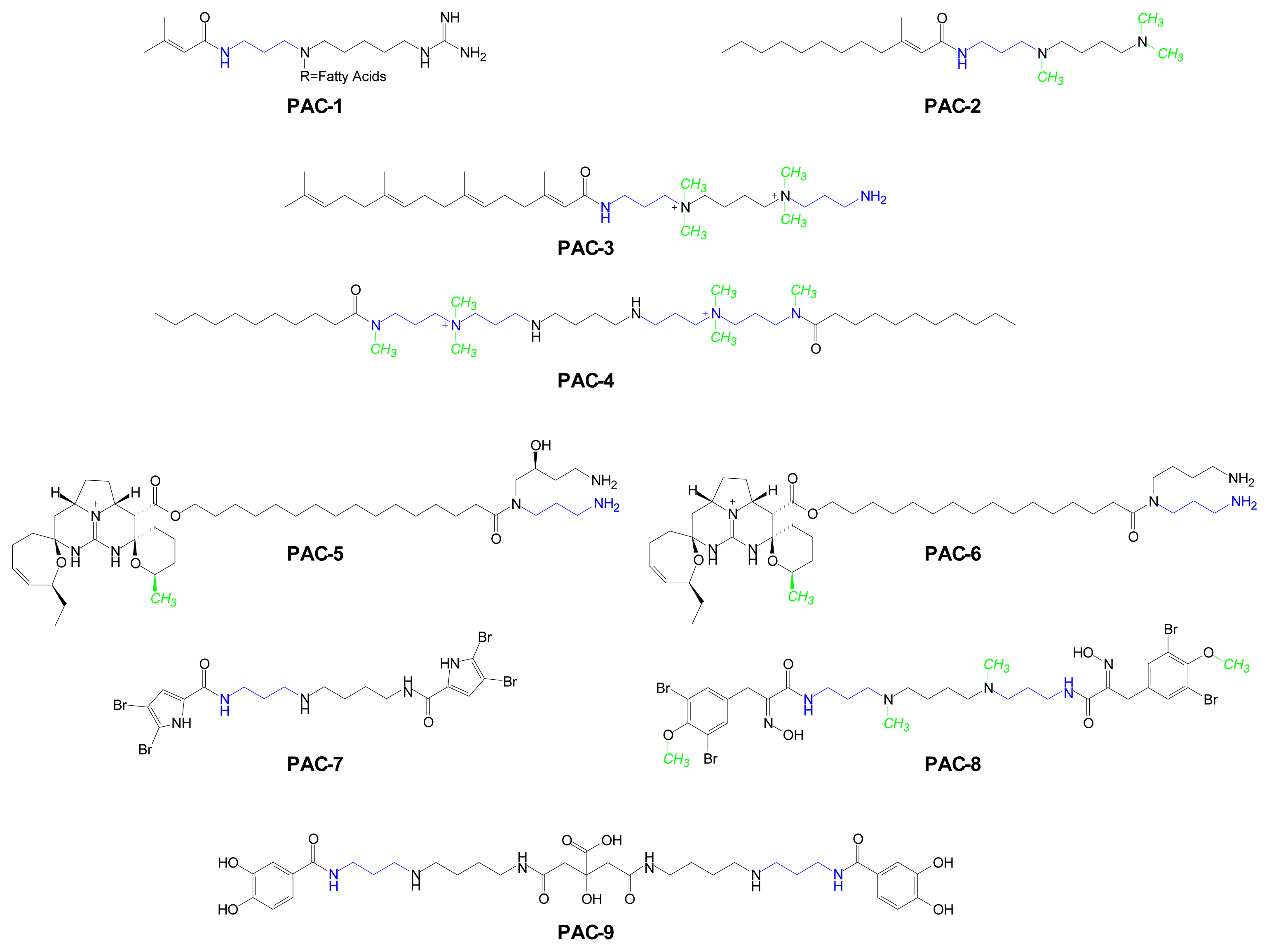
| PAC-1 | Acarnidines | Acarnus erithacus (sponge) | [58] |
| PAC-2 | N-trimethylSpd FAE* | Sinularia brongersmai (coral) | [59] |
| PAC-3 | Sinulamide | Sinularia sp. 1 (coral) | [59,60] |
| PAC-4 | Penaramide A | Penares aff. Incrustans (sponge) | [61] |
| PAC-5 | Crambescidin 800 | Crambe crambe (sponge) | [53,62] |
| PAC-6 | Ptilomycalin A | Hemimycale sp (sponge) | [53,62] |
| PAC-7 | Pseudoceratidine | Pseudoceratina purpurea (sponge) | [63] |
| PAC-8 | Spermatinamine | Pseudoceratina sp. (sponge) | [64] |
| PAC-9 | Petrobactin 1 | Bacillus anthracis str. Sterne (bacterium) | [65] |
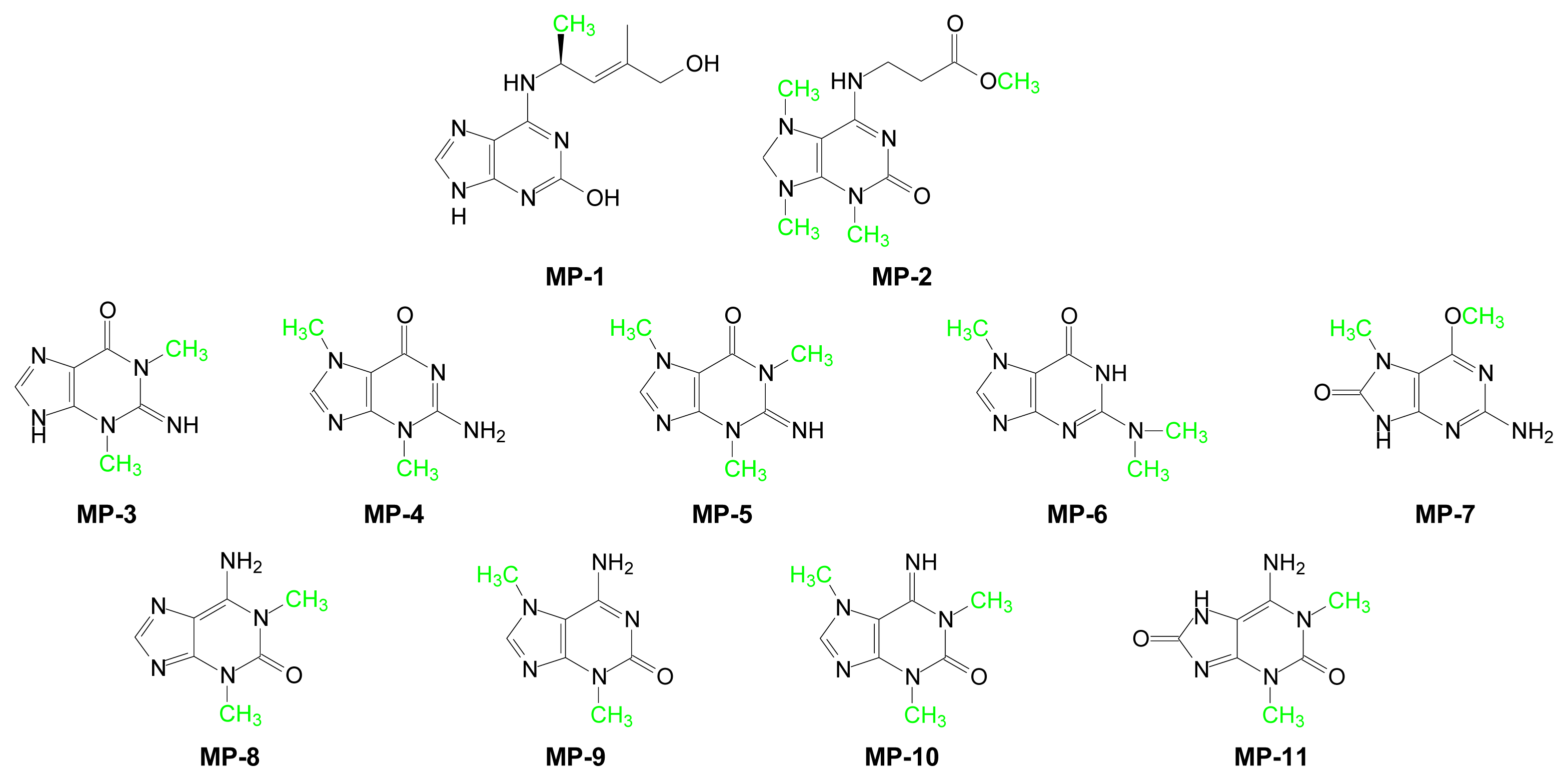
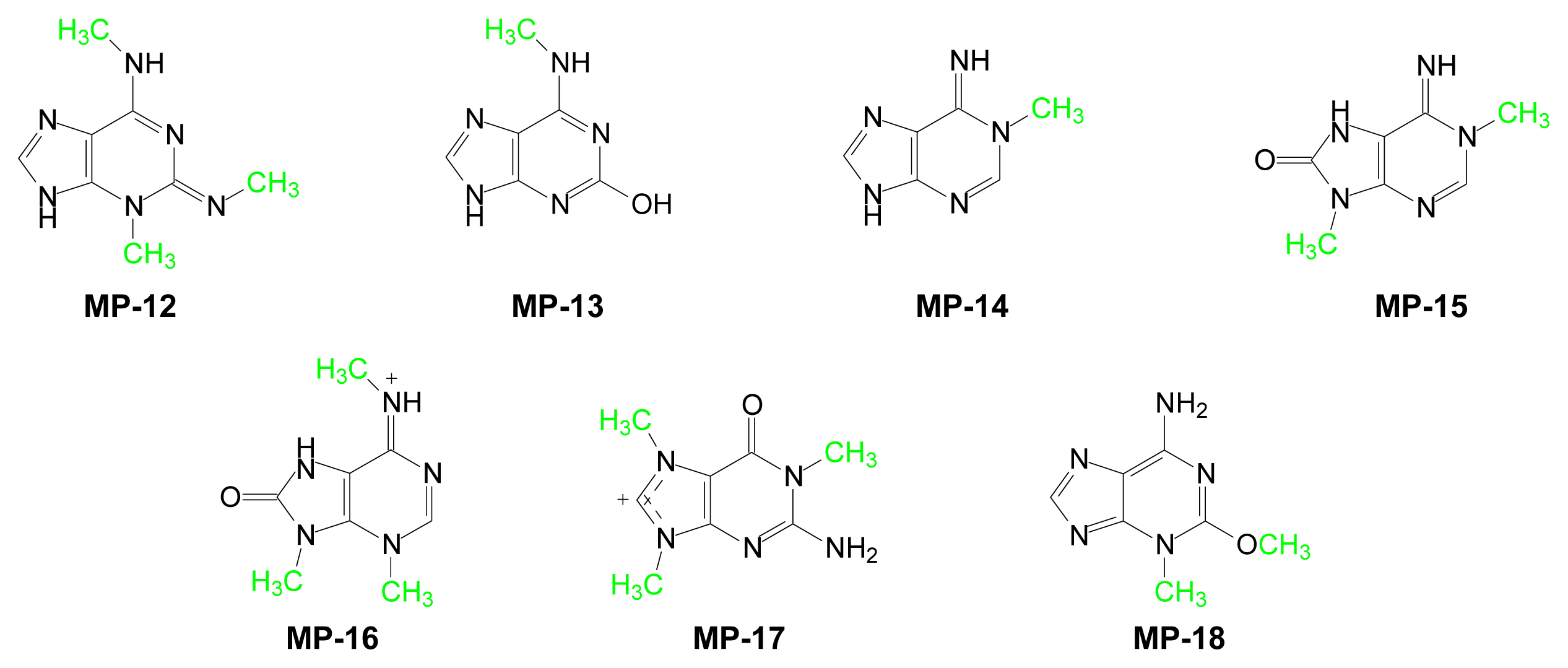
| Compound | Purine | Marine Source | Ref. |
|---|---|---|---|
| MP-1 | 2-Hydroxy-1’-methylzeatin | Green algae and blue coral | [89] |
| MP-2 | Nigricine 4 | Petrosia nigricans (sponge) | [90] |
| MP-3 | 1,3-Dimethylguanine | Botrylloides leachi (acidian) | [91] |
| MP-4 | 3,7-Dimethylguanine | Zyzzya fuliginosa (sponge) | [92] |
| MP-5 | 1,3,7-Trimethylguanine | Latrunculia brevis (sponge) Eudistoma maculosum (ascidian) | [93] [94] |
| MP-6 | N2,N2,N7-Trimethylguanine | Lissoclinum notti (ascidian) | [95] |
| MP-7 | 6-Methoxy-7-methyl-8-oxoguanine | Symplegma rubra (ascidian) | [96] |
| MP-8 | 1,3-Dimethylisoguanine | Amphimedon viridis (sponge) | [97] |
| MP-9 | 3,7-Dimethylisoguanine | Agelas longissima (sponge) | [98] |
| MP-10 | 1,3,7-Trimethylisoguanine | Pseudodistoma cereum (ascidian) | [99] |
| MP-11 | 1,3-Dimethyl-8-oxoisoguanine | Pseudodistoma cereum (ascidian) | [100] |
| MP-12 | 3-Methyl-6-methylamino-2-methylimino-9H-purine | Sagartia troglodytes Price (sea anemone) | [101] |
| MP-13 | 2-Hydroxy-6-methylaminopurine | Green algae and blue coral | [89] |
| MP-14 | 1-Methyl-6-iminopurine | Hymeniacidon Grant (sponge) | [102] |
| MP-15 | 1,9-Dimethyl-6-imino-8-oxopurine | Hymeniacidon sanguinea Grant (sponge) | [102] |
| MP-16 | Caissarone | Bunodosoma-Caissasum (sea-anemone) | [103] |
| MP-17 | 1-Methylherbipoline | Jaspis sp (sponge) | [104] |
| MP-18 | Mucronatine | Stryphnus mucronatus (sponge) | [105] |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sufrin, J.R.; Finckbeiner, S.; Oliver, C.M. Marine-Derived Metabolites of S-Adenosylmethionine as Templates for New Anti-Infectives. Mar. Drugs 2009, 7, 401-434. https://doi.org/10.3390/md7030401
Sufrin JR, Finckbeiner S, Oliver CM. Marine-Derived Metabolites of S-Adenosylmethionine as Templates for New Anti-Infectives. Marine Drugs. 2009; 7(3):401-434. https://doi.org/10.3390/md7030401
Chicago/Turabian StyleSufrin, Janice R., Steven Finckbeiner, and Colin M. Oliver. 2009. "Marine-Derived Metabolites of S-Adenosylmethionine as Templates for New Anti-Infectives" Marine Drugs 7, no. 3: 401-434. https://doi.org/10.3390/md7030401