Effect of Marine Polyunsaturated Fatty Acids on Biofilm Formation of Candida albicans and Candida dubliniensis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
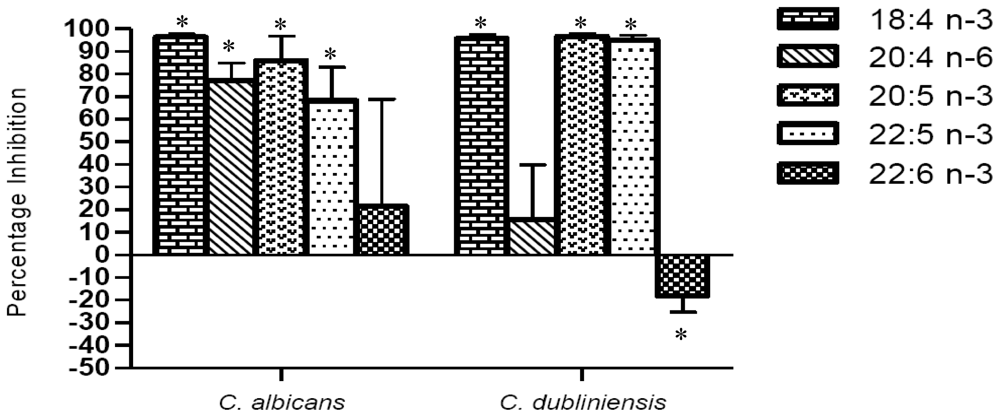
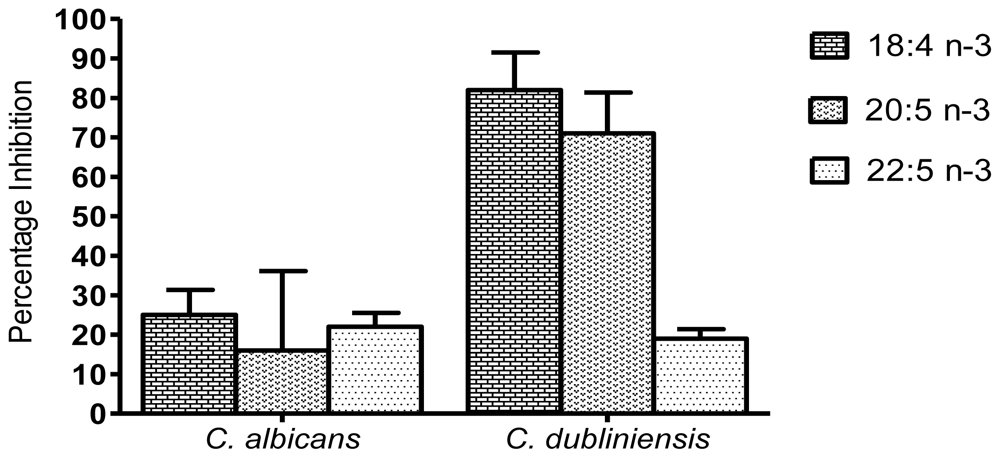
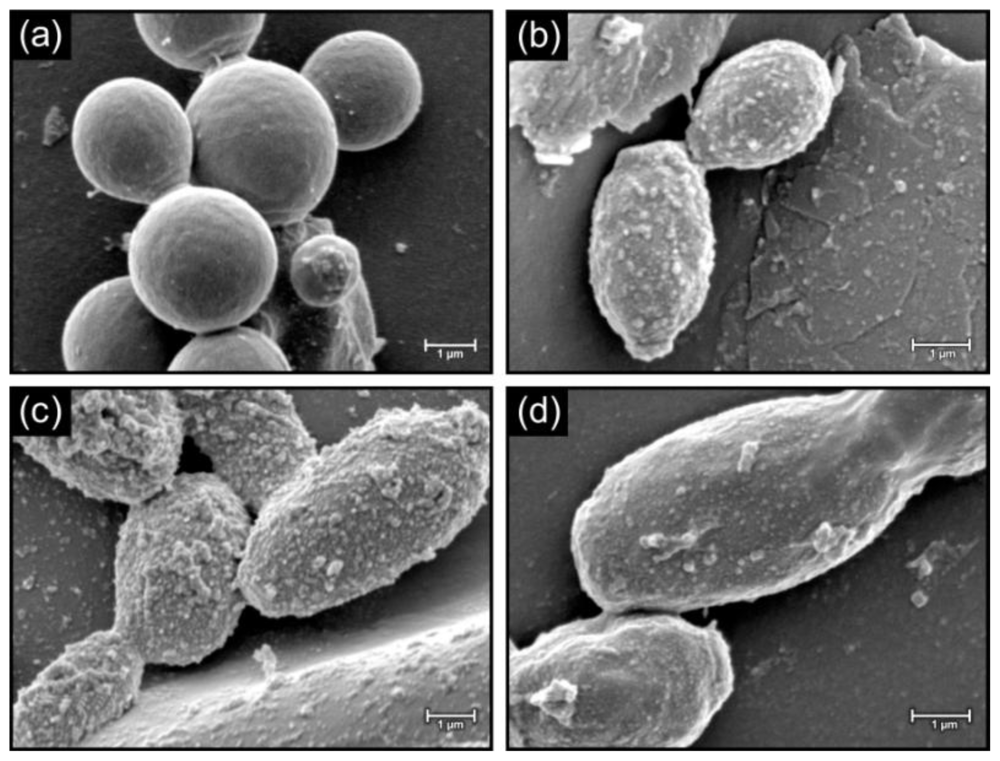
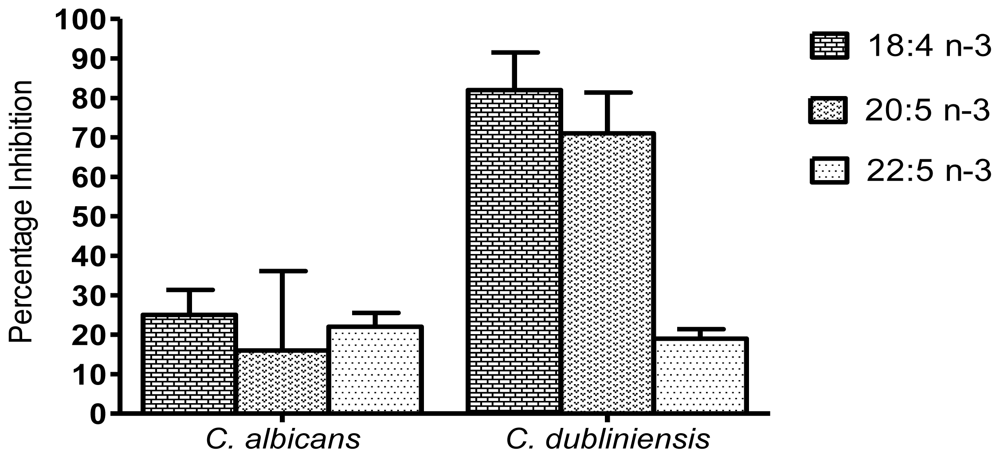
2. Results and Discussion
2.1. Inhibition of mitochondrial metabolism
2.2. Inhibition of biomass production
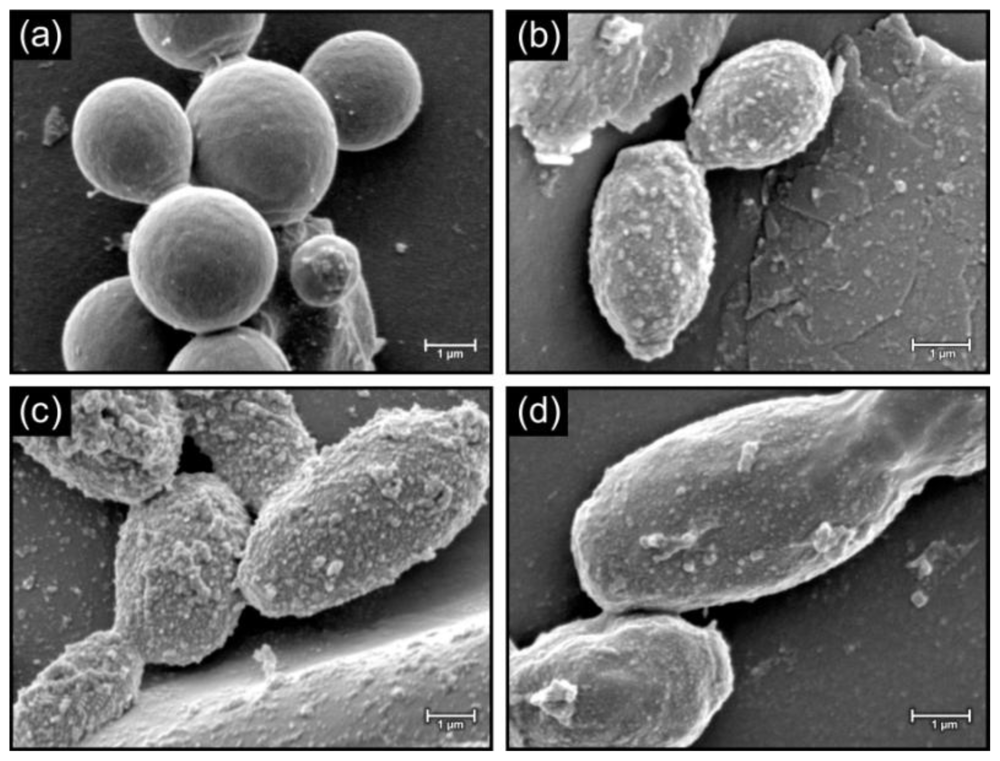
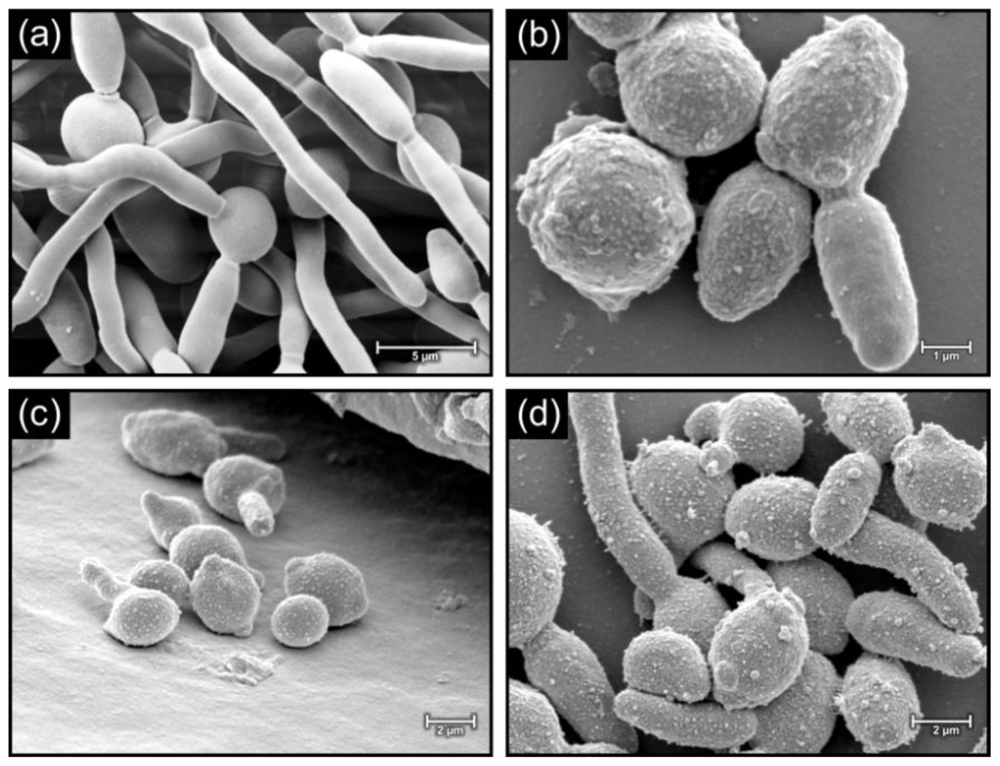
2.3. Morphology of cells in biofilms
3. Experimental
3.1. Strains used
3.2. XTT assay of biofilms
3.3. Biomass determination
3.4. Morphological examination
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Molero, G; Díez-Orejas, R; Navarro-García, F; Monteoliva, L; Pla, J; Gil, C; Sánchez-Pérez, M; Nombela, C. Candida albicans: genetics, dimorphism and pathogenicity. Int. Microbiol 1998, 1, 95–106. [Google Scholar]
- Ramage, G; Vande Walle, K; Wickes, BL; Lopez-Ribot, JL. Biofilm formation by Candida dubliniensis. J. Clin. Microbiol 2001, 39, 3234–3240. [Google Scholar]
- Berman, J; Sudbery, PE. Candida albicans: a molecular revolution built on lessons from budding yeast. Nat. Rev. Genet 2002, 3, 918–930. [Google Scholar]
- Neofytos, D; Fishman, JA; Horn, D; Anaissie, E; Chang, C-H; Olyaei, A; Pfaller, M; Steinbach, WJ; Webster, KM; Marr, KA. Epidemiology and outcome of invasive fungal infections in solid organ transplant recipients. Transpl. Infect. Dis 2010, 12, 220–229. [Google Scholar]
- Coleman, D; Sullivan, D; Harrington, B; Haynes, K; Henman, M; Shanley, D; Bennett, D; Moran, G; McCreary, C; O’Neill, L. Molecular and phenotypic analysis of Candida dubliniensis: a recently identified species linked with oral candidasis in HIV-infected and AIDS patients. Oral Dis 1997, 36(Suppl 1), S96–S101. [Google Scholar]
- Jabra-Rizk, MA; Falkler, WA, Jr; Merz, WG; Baqui, AA; Kelley, JI; Meiller, TF. Retrospective identification and characterization of Candida dubliniensis isolates among Candida albicans clinical laboratory isolates from human immunodeficiency virus (HIV)-infected and non-HIV-infected individuals. J. Clin. Microbiol 2000, 38, 2423–2426. [Google Scholar]
- Coleman, DC; Rinaldi, MG; Haynes, KA; Rex, JH; Summerbel, RC; Anaissie, EJ; Li, A; Sullivan, DJ. Importance of Candida species other than Candida albicans as opportunistic pathogens. Med. Mycol 1998, 36(Suppl 1), 156–165. [Google Scholar]
- Ballie, GS; Douglas, LJ. Candida biofilms and their susceptibility to antifungal agents. Meth. Enzymol 1999, 310, 644–656. [Google Scholar]
- Sylvain, LS; Lucia, VM; Elisabetta, G. Effect of α-linolenic, capric and lauric acid on the fatty acid biosynthesis in Staphylococcus aureus. Int. J. Food Microbiol 2009, 129, 288–294. [Google Scholar]
- Bergusson, G; Arnfinnsson, J; Steingrimsson, O; Thormar, H. In vitro killing of Candida albicans by fatty acids and monoglycerides. Antimicrob. Agents Chemother 2001, 45, 3209–3212. [Google Scholar]
- Noverr, MC; Huffnagle, GB. Regulation of Candida albicans morphogenesis by fatty acid metabolites. Infect. Immun 2004, 72, 6206–6210. [Google Scholar]
- Sahena, F; Zaidul, ISM; Jinap, S; Yazid, AM; Khatib, A; Norulaini, NAN. Fatty acid composition of fish oil extracted from different parts of Indian mackerel (Rastrelliger kanagurta) using various techniques of supercritical CO2 extraction. Food Chem 2010, 120, 879–885. [Google Scholar]
- Saito, H; Xue, C; Yamashiro, R; Moromizato, S; Itabashi, Y. High polyunsaturated fatty acid levels in two subtropical macroalgae, Cladosiphon okamuranus and Caulerpa lentillifera. J. Phycol 2010, 46, 665–673. [Google Scholar] [CrossRef]
- Çelik, M; Diler, A; Küçükgülmez, A. A comparison of the proximate compositions and fatty acid profiles of zander (Sander lucioperca) from two different regions and climatic conditions. Food Chem 2005, 92, 637–641. [Google Scholar]
- De Angelis, L; Risè, P; Giavarini, F; Galli, C; Bolis, CL; Colombo, ML. Marine macroalgae analyzed by mass spectrometry are rich sources of polyunsaturated fatty acids. J. Mass Spectrom 2005, 40, 1605–1608. [Google Scholar]
- Kris-Etherton, PM; Harris, WS; Appel, LJ. Fish oil. Omega-3 fatty acids and cardiovasicular disease. Circulation 2002, 103, 2747–2757. [Google Scholar]
- Singh, RB; Niaz, MA; Sharma, JP; Kumar, R; Rastogi, V; Moshiri, M. Randomised double-blind, placebo-controlled trial of fish and mustard oil in patients with suspected acute mycordial infarction: The Indian experiment of infarct survival. Cardiovasc. Drugs Ther 1997, 11, 485–491. [Google Scholar]
- Von Schacky, C; Angerer, P; Kothny, W; Mudra, H. The effect of dietary omega-3 fatty acids on coronary atherosclerosis. A randomized double-blind, placebo-controlled trial. Ann. Intern. Med 1999, 130, 554–562. [Google Scholar]
- Kuhn, DM; Balkis, M; Chandra, J; Mukherjee, PK; Ghannoum, MA. Uses and limitations of the XTT in the studies of Candida growth and metabolism. J. Clin. Microbiol 2003, 41, 506–508. [Google Scholar]
- Nollin, SD; Borgers, M. Scanning electron microscopy of Candida albicans after in vitro treatment with miconazole. Antimicrob. Agents Chemother 1975, 7, 704–711. [Google Scholar]
- Vandenbosch, D; Braeckmans, K; Nelis, HJ; Coenye, T. Fungicidal activity of miconazole against Candida spp. biofilms. J. Antimicrob. Chemother 2010, 65, 694–700. [Google Scholar]
- Lemar, KM; Passa, O; Aon, MA; Cortassa, S; Müller, CT; Plummer, S; O’Rourke, B; Lloyd, D. Allyl alcohol and garlic (Allium sativum) extract produce oxidative stress in Candida albicans. Microbiology 2005, 151, 3257–3265. [Google Scholar]
- Leeuw, NJ. The influence of oxidized oils on fungal growth and lipid utilization. Ph.D. Thesis, University of the Free State, Bloemfontein, South Africa, 2010. [Google Scholar]
- Samaranayake, YM; Wu, PC; Samaranayake, LP. Relationship between the cell surface hydrophobicity and adherence of Candida krusei and Candida albicans to epithelial and denture acrylic surfaces. APMIS 1995, 103, 707–713. [Google Scholar]
- Van Wyk, PWJ; Wingfield, MJ. Ascospores ultrastructure and development in Ophiostoma cucullatum. Mycologia 1991, 83, 698–707. [Google Scholar]

© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Thibane, V.S.; Kock, J.L.F.; Ells, R.; Wyk, P.W.J.v.; Pohl, C.H. Effect of Marine Polyunsaturated Fatty Acids on Biofilm Formation of Candida albicans and Candida dubliniensis. Mar. Drugs 2010, 8, 2597-2604. https://doi.org/10.3390/md8102597
Thibane VS, Kock JLF, Ells R, Wyk PWJv, Pohl CH. Effect of Marine Polyunsaturated Fatty Acids on Biofilm Formation of Candida albicans and Candida dubliniensis. Marine Drugs. 2010; 8(10):2597-2604. https://doi.org/10.3390/md8102597
Chicago/Turabian StyleThibane, Vuyisile S., Johan L. F. Kock, Ruan Ells, Pieter W. J. van Wyk, and Carolina H. Pohl. 2010. "Effect of Marine Polyunsaturated Fatty Acids on Biofilm Formation of Candida albicans and Candida dubliniensis" Marine Drugs 8, no. 10: 2597-2604. https://doi.org/10.3390/md8102597