The Tropical Brown Alga Lobophora variegata: A Source of Antiprotozoal Compounds

Abstract
:
1. Introduction
2. Results and Discussion
3. Experimental Section
3.1. General experimental procedures
3.2. Algal material
3.3. Extraction and isolation of 1–3
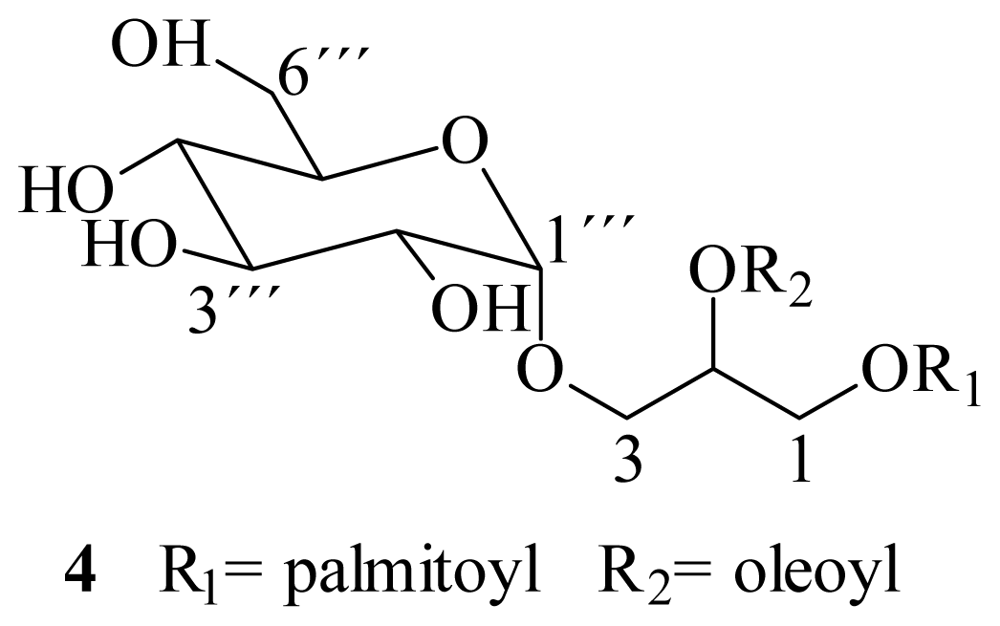
3.4. Alkaline hydrolysis
3.5. Enzymatic hydrolysis
3.6. Antiprotozoal activity
3.7. Cytotoxicity assay
4. Conclusions
Acknowledgements
- Sample Availability: Available from the corresponding author.
References
- Vanacova, S; Liston, D; Tachezy, J; Johnson, P. Molecular biology of the amitochondriate parasites, Giardia intestinalis, Entamoeba histolytica and Trichomonas vaginalis. Int J Parasitol 2003, 33, 235–255. [Google Scholar]
- WHO/PAHO/UNESCO. Report of a consultation of experts on amoebiasis. Weekly Epidemiol Rep 1997, 97–99.
- WHO, The World Health Report 1996; World Health Organization: Geneva, Switzerland, 1996.
- Ali, V; Nozaki, T. Current therapeutics, their problems, and sulfur-containing-amino-acid metabolism as a novel target against infections by “amitochondriate” protozoan parasites. Clin Microbiol Rev 2007, 20, 164–87. [Google Scholar]
- Nanda, N; Michel, RG; Kurdgelashvili, G; Wendel, KA. Trichomoniasis and its treatment. Expert Rev Anti Infect Ther 2006, 4, 125–135. [Google Scholar]
- Rossignol, JF; Maisonneuve, H; Cho, YW. Nitroimidazoles in the treatment of trichomoniasis, giardiasis, and amebiasis. Int J Clin Pharmacol Ther Toxicol 1984, 22, 63–72. [Google Scholar]
- Upcroft, P; Upcroft, JA. Drug targets and mechanism of resistance in the anaerobic protozoa. Clin Microbiol Rev 2001, 14, 150–164. [Google Scholar]
- Gardner, TB; Hill, DR. Treatment of Giardiasis. Clin Microbiol Rev 2001, 14, 114–128. [Google Scholar]
- Lopez, MM; Palermo, AM; Mudry, MD; Carballo, MA. Cytogenetic evaluation of two nitroimidazole derivatives. Toxicol in Vitro 2003, 17, 35–40. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat Prod Rep 2002, 19, 1–49. [Google Scholar]
- Blunt, JW; Copp, BR; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat Prod Rep 2006, 23, 26–78. [Google Scholar]
- Moo-Puc, R; Robledo, D; Freile-Pelegrín, Y. Evaluation of selected tropical seaweeds for in vitro anti-trichomonal activity. J Ethnopharmacol 2008, 120, 92–97. [Google Scholar]
- León-Deniz, LV; Dumonteil, E; Moo-Puc, R; Freile-Pelegrin, Y. Antitrypanosomal activity of tropical marine algae extracts. Pharm Biol 2009, 47, 864–871. [Google Scholar]
- Zubia, M; Robledo, D; Freile-Pelegrin, Y. Antioxidant activities in tropical marine macroalgae from the Yucatan Peninsula, Mexico. J Appl Phycol 2007, 19, 449–458. [Google Scholar]
- Kupchan, SM; Britton, RW; Lacadie, JA; Ziegler, MF; Sigel, CW. The isolation and structural elucidation of Bruceantin and Bruceantinol, new potent antileukemic Quassinoids from Brucea antidysenterica. J Org Chem 1975, 40, 648–654. [Google Scholar]
- Wang, H; Li, YL; Shen, WZ; Rui, W; Ma, XJ; Cen, YZ. Antiviral activity of a sulfoquinovosyldiacylglycerol (SQDG) compound isolated from the green alga Caulerpa racemosa. Bot Mar 2007, 50, 185–190. [Google Scholar]
- Fischer, W; Heinz, E; Zeus, M. The Suitability of lipase from Rhizopus arrhizus delemar for analysis of fatty acid distribution in dihexosyl diglycerides, phospholipids and plant sulfolipids. Hoppe-Seyler’s Z Physiol Chem 1973, 354, 1115–1123. [Google Scholar]
- Kim, YH; Yoo, JS; Kim, MS. Structural characterization of sulfoquinovosyl, monogalactosyl and digalactosyl diacylglycerols by FAB-CID-MS/MS. J Mass Spectrom 1997, 32, 968–977. [Google Scholar]
- Costello, CE; Vath, JE. McCloskey, JA, Ed.; Mass spectrometry. In Methods in Enzimology; Academic Press: San Diego, CA, USA, 1990; Volume 193, p. 738. [Google Scholar]
- Domon, B; Costello, CE. Structure elucidation of glycosphingolipids and gangliosides using high-performance tandem mass spectrometry. Biochemistry 1988, 27, 1534–1543. [Google Scholar]
- Rastrelli, L; De Tommassi, N; Berger, I; Caceres, A. Glycolipids from Byrsonima crassifolia. Phytochemistry 1997, 45, 647–650. [Google Scholar]
- Takahashi, Y; Itabashi, Y; Suzuki, M; Kuksis, A. Determination of stereochemical configuration of the glicerol moieties in Glycoglycerolipids by chiral phase High-Performance Liquid Chromatography. Lipids 2001, 36, 741–748. [Google Scholar]
- Zhou, B; Tang, S; Johnson, R; Mattern, M; Lazo, J; Sharlow, E; Harich, K; Kingston, D. New glycolipid inhibitors of Myt1 kinase. Tetrahedron 2005, 61, 883–887. [Google Scholar]
- Shao, ZY; Cai, JN; Ye, QZ; Guo, YW. Crassicaulisine, a new sulphonoglycolipid from the red alga Chondria crassicaulis Harv. J Asian Nat Prod Res 2002, 4, 205–209. [Google Scholar]
- Tang, HF; Yi, YH; Yao, XS; Wu, JH; Zhang, SY; Xu, QZ. Studies on the chemical constituents from marine brown alga Ishige okamurai. Zhongguo Zhong Yao Za Zhi 2002, 27, 269–273. [Google Scholar]
- Amarquaye, A; Che, CT; Bejar, E; Malone, MH; Fong, HHS. A New Glycolipid from Byrsonima crassifolia. Planta Med 1994, 60, 85–86. [Google Scholar]
- Qi, SH; Zhang, S; Huang, JS; Xiao, ZH; Wu, J; Long, LJ. Glycerol derivatives and sterols from Sargassum parvivesiculosum. Chem Pharm Bull 2004, 52, 986–988. [Google Scholar]
- Golik, J; Dickey, JK; Todderud, G; Lee, D; Alford, J; Huang, S; Klohr, S; Eustice, D; Aruffo, A; Agler, ML. Isolation and structure determination of sulfoquinovosyl dipalmitoyl glyceride, a P-selectin receptor inhibitor from the alga Dictyochloris fragrans. J Nat Prod 1997, 60, 387–389. [Google Scholar]
- Wu, W; Hasumi, K; Peng, H; Hu, X; Wang, X; Bao, B. Fibrinolytic compounds isolated from a brown alga Sargassum fulvellum. Mar Drugs 2009, 7, 85–94. [Google Scholar]
- Mizushina, Y; Kasai, N; Iijima, H; Sugawara, F; Yoshida, H; Sakaguchi, K. Sulfo-quinovosyl-acyl-glycerol (SQAG), a eukaryotic DNA polymerase inhibitor and anti-cancer agent. Curr Med Chem Anticancer Agents 2005, 5, 613–625. [Google Scholar]
- Murakami, C; Yamazaki, T; Hanashima, S; Takahashi, S; Takemura, M; Yoshida, S; Ohta, K; Yoshida, H; Sugawara, F; Sakaguchi, K; Mizushina, Y. A novel DNA polymerase inhibitor and a potent apoptosis inducer: 2-Mono-O-acyl-3-O-(α-d-sulfoquinovosyl)-glyceride with stearic acid. BBA Proteins Proteomics 2003, 1645, 72–80. [Google Scholar]
- Yamazaki, T; Aoki, S; Ohta, K; Hyuma, S; Sakaguchi, K; Sugawara, F. Synthesis of an immunosuppressant SQAG9 and determination of the binding peptide by T7 phage display. Bioorg Med Chem Lett 2004, 14, 4343–4346. [Google Scholar]
- Loya, S; Reshef, V; Mizrachi, E; Silberstein, C; Rachamim, Y; Carmeli, S; Hizi, A. The inhibition of the reverse transcriptase of HIV-1 by the natural sulfoglycolipids from cyanobacteria: Contribution of different moieties to their high potency. J Nat Prod 1998, 61, 891–895. [Google Scholar]
- Dai, JQ; Zhu, QX; Zhao, CY; Yang, L; Li, Y. Glyceroglycolipids from Serratula strangulata. Phytochemistry 2001, 58, 1305–1309. [Google Scholar]
- Al-Fadhli, A; Wahidulla, S; D’Souza, L. Glycolipids from the red alga Chondria armata (Kütz.) Okamura. Glycobiology 2006, 16, 902–915. [Google Scholar]
- Vonthron-Sénécheau, C; Weniger, B; Ouattara, M; Fezan, TB; Kamenan, A; Lobstein, A; Brun, R; Anton, R. In vitro antiplasmodial activity and cytotoxicity of ethnobotanically selected Ivorian plants. J Ethnopharmacol 2003, 87, 221–225. [Google Scholar]
- Littler, DS; Littler, MM. Caribbean Reef Plants, 2nd ed; Offshore Graphics, Inc: Washington, DC, USA, 2000; p. 300. [Google Scholar]
- Cedillo-Rivera, R; Chávez, B; González-Robles, A; Tapia-Contreras, A; Yépez-Mulia, L. In vitro effect of nitazoxanide against Entamoeba histolytica, Giardia lamblia and Trichomonas vaginalis trophozoites. J Eukaryot Microbiol 2002, 49, 201–208. [Google Scholar]
- Rahman, A; Choudhary, MI; Thomsen, WJ. Bioassay techniques for drug development. In Manual of Bioassay Techniques for Natural Products Research; Harword Academic Publishers: Amsterdam, The Netherlands, 2001; pp. 34–35. [Google Scholar]




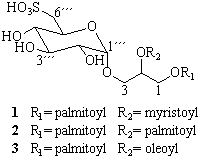{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds 1 and 2 | Compound 3 | |||
|---|---|---|---|---|
| position | δH/mult., (J in Hz) | δC | δH/mult., (J in Hz) | δC |
| 1a | 4.34 dd (12.0, 2.4) | 62.7 | 4.34 dd (12.0, 2.4) | 62.7 |
| 1b | 4.13 dd (12.0, 7.5) | 4.13 dd (12.0, 7.5) | ||
| 2 | 5.13 m | 69.8 | 5.13 m | 69.8 |
| 3a | 3.89 dd (10.5, 6.0) | 64.6 | 3.89 dd (10.5, 6.0) | 64.6 |
| 3b | 3.39 | 3.39 | ||
| 1‴ | 4.57 d (3.5) | 98.3 | 4.57 d (3.5) | 98.3 |
| 2‴ | 3.18 dd (3.5, 9.6) | 71.7 | 3.18 dd (3.5, 9.6) | 71.7 |
| 3‴ | 3.35 | 72.9 | 3.35 | 72.9 |
| 4‴ | 2.92 dd (9.6) | 74.2 | 2.92 dd (9.6) | 74.2 |
| 5‴ | 3.77 ddd (9.6, 6.8, 4.3) | 68.6 | 3.77 ddd (9.6, 6.8, 4.3) | 68.6 |
| 6‴a | 2.90 dd (14.0, 4.3) | 54.5 | 2.90 dd (14.0, 4.3) | 54.5 |
| 6‴b | 2.54 dd (14.0, 6.9) | 2.54 dd (14.0, 6.9) | ||
| 1′, 1″ | 172.4, 172.6 | 172.4, 172.6 | ||
| 2′, 2″ | 2.28 m | 33.6 | 2.28 m | 33.6 |
| 3′, 3″ | 1.49 m | 31.4 | 1.49 m | 31.4 |
| 8″, 11″ | 1.97 m | 26.7 | ||
| 9″, 10″ | 5.3 t (4.7) | 129.61, 129.64 | ||
| -CH2 | 1.22 bs | 29.2–22.2 | 1.22 bs | 29.2–22.2 |
| CH3 | 0.84 t (6.9) | 14.0 | 0.84 t (6.9) | 14.0 |
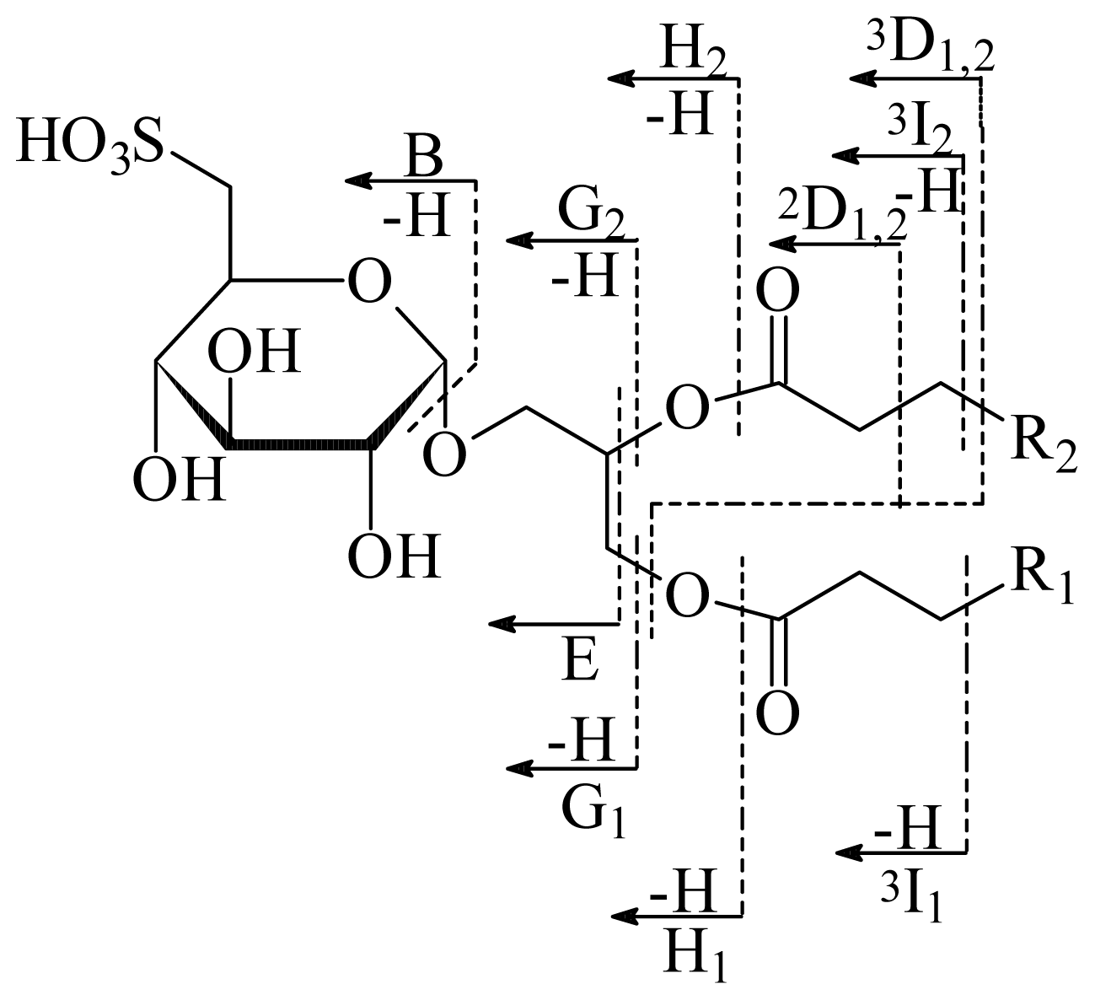
Product ions, m/z | |||
|---|---|---|---|
| Assignment | SQDG-1 [M-H+2Na]+ = 811 | SQDG-2 [M-H+2Na]+ = 839 | SQDG-3 [M-H+2Na]+ = 865 |
| B | 271 | 271 | 271 |
| E | 329 | 329 | 329 |
| 2D1,2 | 387 | 387 | 387 |
| 3D1,2 | 401 | 401 | 401 |
| G1 | 555 | 583 | 609 |
| G2 | 583 | 583 | 583 |
| H1 | 571 | 599 | 625 |
| H2 | 599 | 599 | 599 |
| 3I1 | 627 | 655 | 681 |
| 3I2 | 655 | 655 | 655 |
| SO3Na2+ | 126 | 126 | 126 |
| Extract/fraction/compound (weight) | Antiprotozoal activity IC50 μg/mL | Cytotoxicity CC50 μg/mL | Selectivity index SI | ||||
|---|---|---|---|---|---|---|---|
| Gi | Eh | Tv | MDCK | MDCK/Gi | MDCK/Eh | MDCK/Tv | |
| Whole extract (35.5 g) | 10.5 ± 0.45 | 10.8± 0.09 | 3.2± 0.09 | 78.0 ± 2.45 | 7.4 | 7.2 | 24.4 |
| Hexane fraction (10 g) | 1.0 ± 0.06 | 2.8 ± 0.08 | 13.4 ± 0.61 | 65.0 ± 1,23 | 65 | 23.2 | 4.9 |
| Chloroform fraction (3.6 g) | 0.5 ± 0.02 | 6.2 ± 0.32 | 3.7 ± 0.11 | 87.5 ± 0.89 | 175 | 14.1 | 23.6 |
| Ethyl acetate fraction (16.7 g) | 0.8 ± 0.03 | 1.7 ± 0.04 | 11.7 ± 0.22 | 500.1 ± 3.4 | 625.1 | 294.2 | 42.7 |
| n-butanol fraction (3.4 g) | - | 15.5 ± 0.73 | - | 90.8 ± 1.78 | nt | 5.9 | nt |
| Aqueous residue (1.8 g) | 8.8 ± 0.71 | 15.6 ± 0.41 | - | 789.9 ± 2.67 | 89.8 | 50.6 | nt |
| SQDG’s 1–3 (276.3 mg) | 20.9 ± 0.89 | 3.9 ± 0.03 | 8.0 ± 0.42 | 85.3 ± 1.26 | 4.1 | 21.9 | 10.7 |
| Metronidazole | 0.22 ± 0.0005 | 0.13 ± 0.0005 | 0.04 ± 0.02 | 68 ± 1.2 | 309 | 523 | 1700 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cantillo-Ciau, Z.; Moo-Puc, R.; Quijano, L.; Freile-Pelegrín, Y. The Tropical Brown Alga Lobophora variegata: A Source of Antiprotozoal Compounds. Mar. Drugs 2010, 8, 1292-1304. https://doi.org/10.3390/md8041292
Cantillo-Ciau Z, Moo-Puc R, Quijano L, Freile-Pelegrín Y. The Tropical Brown Alga Lobophora variegata: A Source of Antiprotozoal Compounds. Marine Drugs. 2010; 8(4):1292-1304. https://doi.org/10.3390/md8041292
Chicago/Turabian StyleCantillo-Ciau, Zulema, Rosa Moo-Puc, Leovigildo Quijano, and Yolanda Freile-Pelegrín. 2010. "The Tropical Brown Alga Lobophora variegata: A Source of Antiprotozoal Compounds" Marine Drugs 8, no. 4: 1292-1304. https://doi.org/10.3390/md8041292