Cloning and Characterization of Glutamate Receptors in Californian Sea Lions (Zalophus californianus)

Abstract
:1. Introduction
2. Results
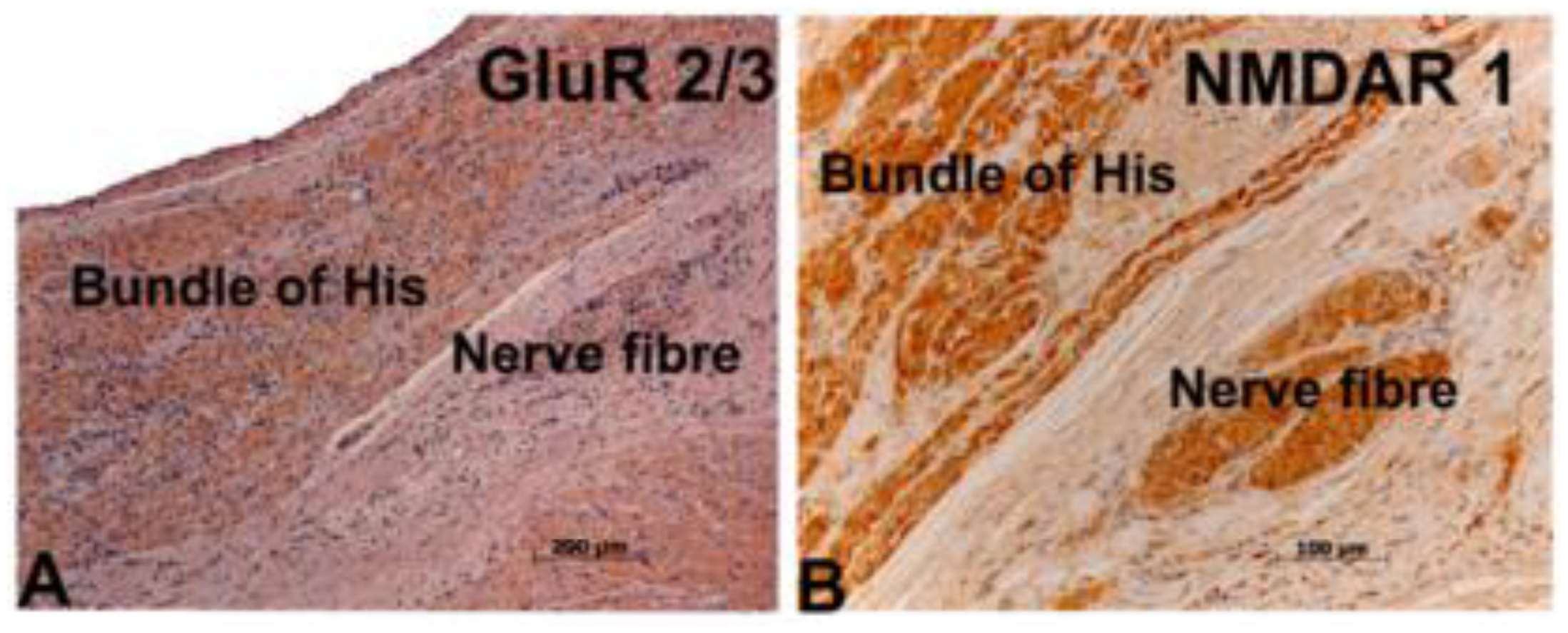
3. Expression Analysis
4. Discussion
5. Experimental Section
5.1. Materials and Methods
5.2. Tissue Preparation and Immunohistochemistry
5.3. Primer Design
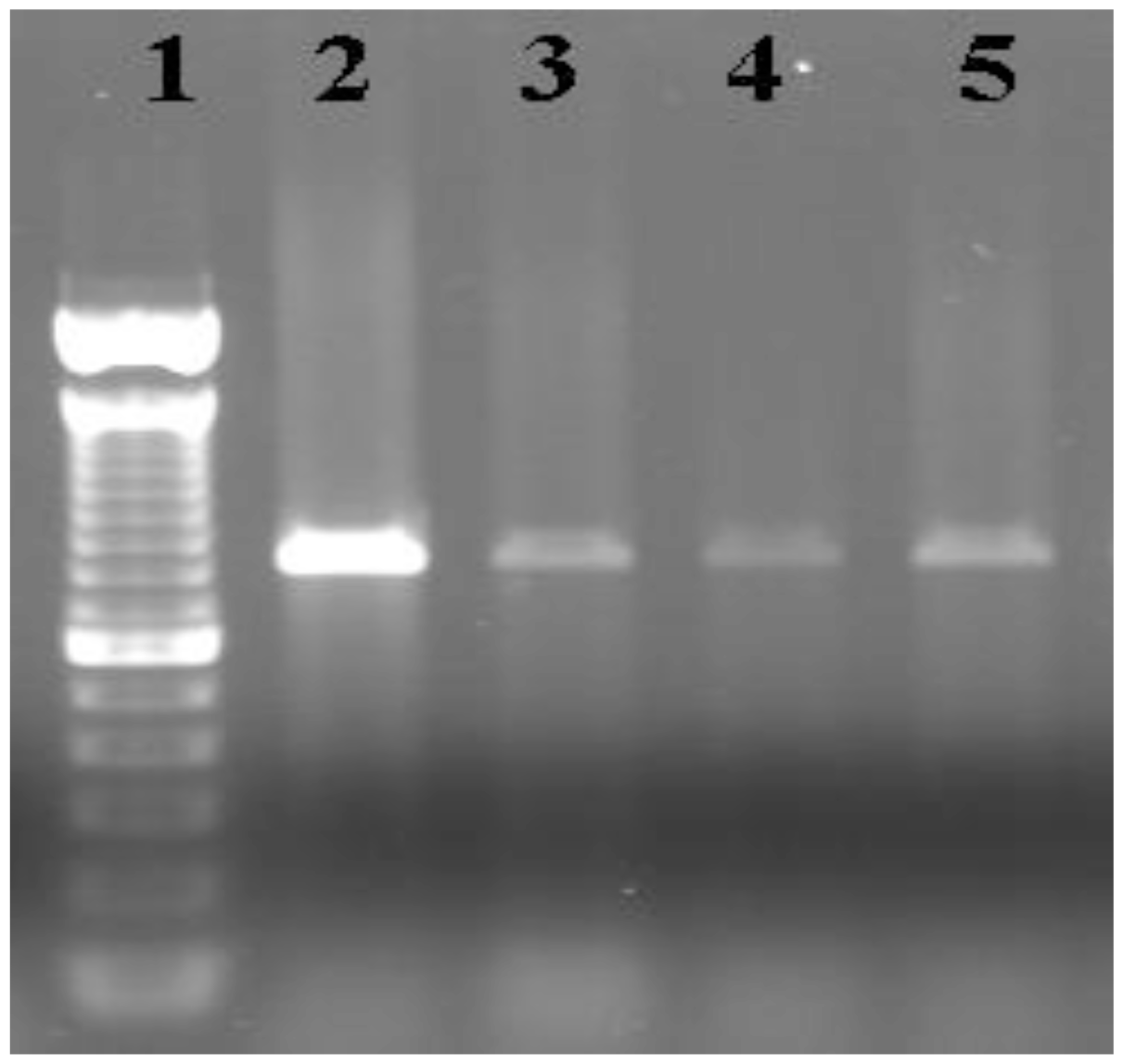
5.4. PCR and Cloning
5.5. Sequencing of Clones
6. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Iverson, F; Truelove, J; Tryphonas, L; Nera, EA. The toxicology of domoic acid administered systemically to rodents and primates. Can Dis Wkly Rep 1990, 16, 15–19. [Google Scholar]
- Perl, TM; Bedard, L; Kosatsky, T; Hockin, JC; Todd, EC; McNutt, L; Remis, RS. An outbreak of toxic encephalopathy caused by eating mussels contaminated with domoic acid. N Engl J Med 1990, 322, 1775–1780. [Google Scholar]
- Pulido, O. Domoic acid toxicologic pathology. Mar Drugs 2008, 6, 180–219. [Google Scholar]
- Teitelbaum, J; Zatorre, RJ; Carpenter, S; Gendron, D; Cashman, NR. Neurological sequelae of domoic acid intoxication. Can Dis Wkly Rep 1990, 16, 9–12. [Google Scholar]
- Teitelbaum, JS; Zatorre, RJ; Carpenter, S; Gendron, D; Evans, AC; Gjedde, A; Cashman, NR. Neurologic sequelae of domoic acid intoxication due to the ingestion of contaminated mussels. N Engl J Med 1990, 322, 1781–1787. [Google Scholar]
- Tryphonas, L; Truelove, J; Iverson, F; Todd;, ECD; Nera, EA. Neuropathology of experimental domoic acid poisoning in non-human primates and rats. Can Dis Wkly Rep 1990, 16, 75–81. [Google Scholar]
- Bakke, MJ; Horsberg, TE. Effects of algal-produced neurotoxins on metabolic activity in telencephalon, optic tectum and cerebellum of Atlantic salmon (Salmo salar). Aquat Toxicol 2007, 85, 96–103. [Google Scholar]
- Goldstein, T; Mazet, JA; Zabka, TS; Langlois, G; Colegrove, KM; Silver, M; Bargu-Astes, S; Van Dolah, F; Leighfield, T; Conrad, PA; Barakos, J; Williams, DC; Dennison, S; Haulena, MA; Gulland, FMD. Novel symptomatology and changing epidemiology of domoic acid toxicosis in California sea lions (Zalophus californianus): An increasing risk to marine mammal health. Proc Roy Soc B 2008, 275, 267–276. [Google Scholar]
- Greig, DJ; Gulland, FMD; Kreuder, C. A decade of live California sea lion (Zalophus californianus) strandings along the central California coast: Causes and trends, 1991–2000. Aquat Mammals 2005, 3, 40–51. [Google Scholar]
- Gulland, FMD; Haulena, MA; Fauquier, D; Langlois, G; Lander, ME; Zabka, T; Duerr, R. Domoic acid toxicity in Californian sea lions (Zalophus californianus): Clinical signs, treatment and survival. Vet Rec 2002, 150, 475–480. [Google Scholar]
- Kreuder, C; Miller, MA; Lowenstine, LJ; Conrad, PA; Carpenter, TE; Jessup, DA; Mazet, JAK. Evaluation of cardiac lesions and risk factors associated with myocarditis and dilated cardiomyopathy in southern sea otters (Enhydra lutris nereis). Am J Vet Res 2005, 66, 289–299. [Google Scholar]
- Lowenstine, LJ; Spraker, T; Lipscomb, TP; Gulland, FMD. Pathology of domoic acid toxicity in California sea lions (Zalophus californianus). Vet Pathol 2005, 42, 184–191. [Google Scholar]
- Scholin, CA; Gulland, F; Doucette, GJ; Benson, S; Busman, M; Chavez, FP; Cordaro, J; DeLong, R; DeVogelaere, A; Harvey, J; Haulena, M; Lefebvre, K; Lipscomb, T; Loscutoff, S; Lowenstine, LJ; Marin, R; Miller, PE; McLellan, WA; Moeller, PD; Powell, CL; Rowles, T; Silvagni, P; Silver, M; Spraker, T; Trainer, V; Van Dolah, FM. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 403, 80–84. [Google Scholar]
- Silvagni, PA; Lowenstine, LJ; Spraker, T; Lipscomb, TP; Gulland, FM. Pathology of domoic acid toxicity in California sea lions (Zalophus californianus). Vet Pathol 2005, 42, 184–191. [Google Scholar]
- Work, TM; Barr, B; Beale, AM; Fritz, L; Quiallam, MA; Wright, LJC. Epidemiology of domoic acid poisoning in brown pelicans (Pelecanus occidentalis) and Brandt’s cormorants (Phalacrocorax penicillatus) in California. J Zoo Wild Med 1993, 24, 54–62. [Google Scholar]
- Zabka, TS; Goldstein, T; Cross, C; Mueller, R; Kreuder-Johnson, C; Gill, S; Gulland, FMD. Characterization of a degenerative cardiomyopathy associated with exposure to domoic acid toxin in California sea lions (Zalophus californianus). Vet Pathol 2009, 45, 105–119. [Google Scholar]
- Gill, S; Pulido, O. Gill, S, Pulido, O, Eds.; Glutamate receptors in peripheral tissues. Distribution, and implications for toxicology. In Glutamate Receptors in Peripheral Tissue: Excitatory Transmission Outside the CNS; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2005. [Google Scholar]
- Gill, S; Veinot, J; Kavanagh, M; Pulido, O. Human heart glutamate receptors—implications for toxicology, food safety, and drug discovery. Toxicol Pathol 2007, 35, 411–417. [Google Scholar]
- Mueller, R; Gill, S; Pulido, O. Neural biomarkers in the monkey (Macaca fascicularis) heart: An immunohistochemical study-Potential target sites for neurotoxins and pharmacological interventions. Toxicol Pathol 2003, 31, 227–234. [Google Scholar]
- Cunningham, MD; Ferkany, JW; Enna, SJ. Excitatory amino acid receptors: A gallery of new targets for pharmacological intervention. Life Sci 1994, 4, 135–148. [Google Scholar]
- Hollmann, M; Heinemann, S. Cloned glutamate receptors. Ann Rev Neurosci 1994, 17, 31–108. [Google Scholar]
- Pinheiro, P; Mulle, C. Kainate Receptor. Cell Tissue Res 2006, 326, 457–482. [Google Scholar]
- Gao, X; Xu, X; Pang, J; Zhang, C; Ding, JM; Peng, X; Liu, Y; Cao, JM. NMDA receptor activation induces mitochondrial dysfunction, oxidative stress and apoptosis in cultured neonatal rat cardiomyocytes. Physiol Rev 2007, 56, 559–569. [Google Scholar]
- Gill, S; Pulido, O. Glutamate receptors in peripheral tissues: Current knowledge, future research, and implications for toxicology. Toxicol Pathol 2001, 20, 208–223. [Google Scholar]
- Van Deutekom, JCT; Wljmenga, C; Van Tlenhoven, EAE; Gruter, AM; Hewitt, JE; Padberg, GW; Van Ommen, GJV; Hofker, MH; Fronts, RR. FSHD associated DNA rearrangements are due to deletions of integral copies of a 3.2 kb tandemly repeated unit. Hum Mol Genet 1993, 2, 2037–2042. [Google Scholar]
- Nijjar, MS; Pierce, GN; Nijjar, SS; Dhalla, NS. Domoic acid attenuates the adenosine-5′-triphosphate-induced increase. J Cardiovasc Pharmacol Ther 1999, 4, 159–166. [Google Scholar]
- Perl, TM; Bedard, L; Kosatsky, T; Hockin, JC; Todd, EC; McNutt, L; Remis, RS. An outbreak of toxic encephalopathy caused by eating mussels contaminated with domoic acid. N Engl J Med 1990, 322, 1775–1780. [Google Scholar]
- Reyes, A; Gissi, C; Catzeflis, F; Nevo, E; Pesole, G; Saccone, C. Congruent mammalian trees from mitochondrial and nuclear genes using Bayesian methods. Mol Biol Evol 2004, 21, 397–403. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene of interest | Forward Primer | Reverse Primer | Expected fragment size |
|---|---|---|---|
| GluR 6 | GGACTAAACCTGGCTATGAC | CATGCAGCAAGGTTCTGAGC | 840 bp |
| Kainate Ka 2 | GCACATGGGCCGCAAGC | CAGCAATGATGAGGCCACAG | 875 bp |
| GluR 1 | GTTCCCAGTTCTCCAAAGGAG | R1-CAT TCC AGT AAC CGA TCT TTC GG R2-CAT TCC AGT AAC CAA TCT TTC GG | 927 bp |
| mGluR 5 | CTTCAGGCGAAGCATGAAG | GGTGGCGGCAGCGGATG | 947 bp |
| NMDAR 1 | CTAGCCAGGTCTACGCTATC | CTGGGAATCTCCTTCTTGACC | 1393 bp |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gill, S.; Goldstein, T.; Situ, D.; Zabka, T.S.; Gulland, F.M.D.; Mueller, R.W. Cloning and Characterization of Glutamate Receptors in Californian Sea Lions (Zalophus californianus). Mar. Drugs 2010, 8, 1637-1649. https://doi.org/10.3390/md8051637
Gill S, Goldstein T, Situ D, Zabka TS, Gulland FMD, Mueller RW. Cloning and Characterization of Glutamate Receptors in Californian Sea Lions (Zalophus californianus). Marine Drugs. 2010; 8(5):1637-1649. https://doi.org/10.3390/md8051637
Chicago/Turabian StyleGill, Santokh, Tracey Goldstein, Donna Situ, Tanja S. Zabka, Frances M. D. Gulland, and Rudi W. Mueller. 2010. "Cloning and Characterization of Glutamate Receptors in Californian Sea Lions (Zalophus californianus)" Marine Drugs 8, no. 5: 1637-1649. https://doi.org/10.3390/md8051637