Prebiotics from Marine Macroalgae for Human and Animal Health Applications

 ,
,
Abstract
:1. Introduction
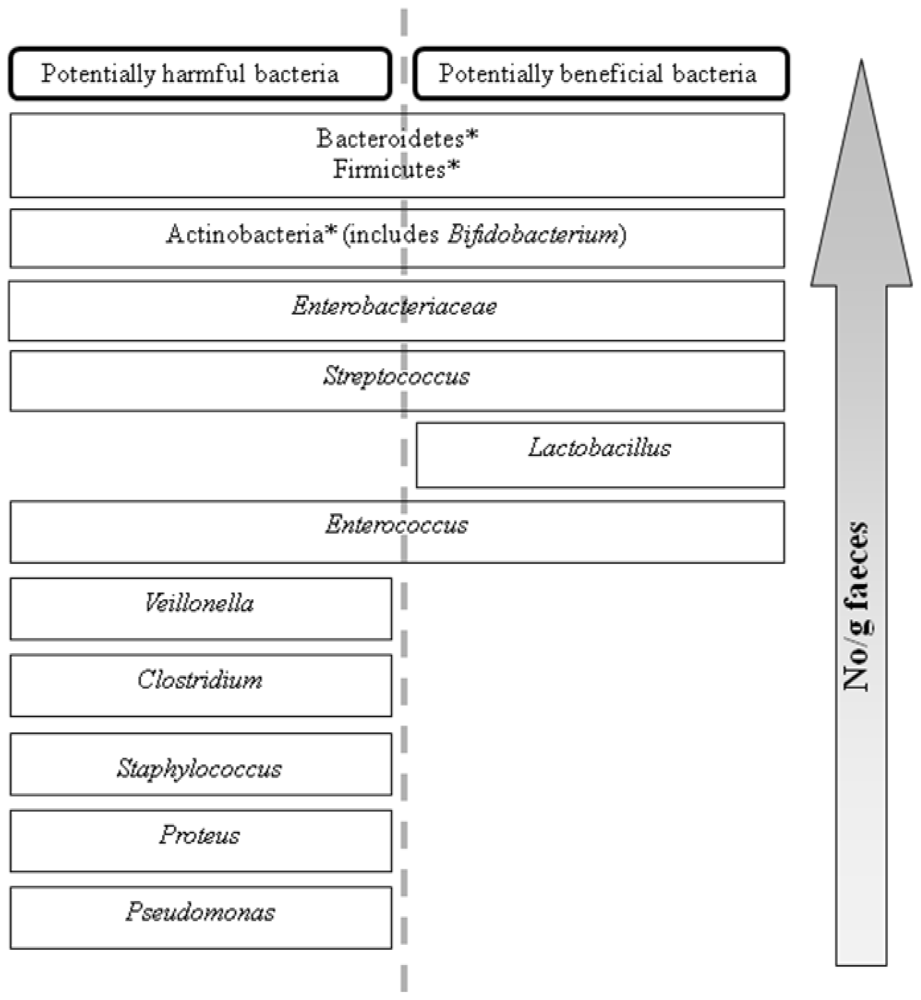
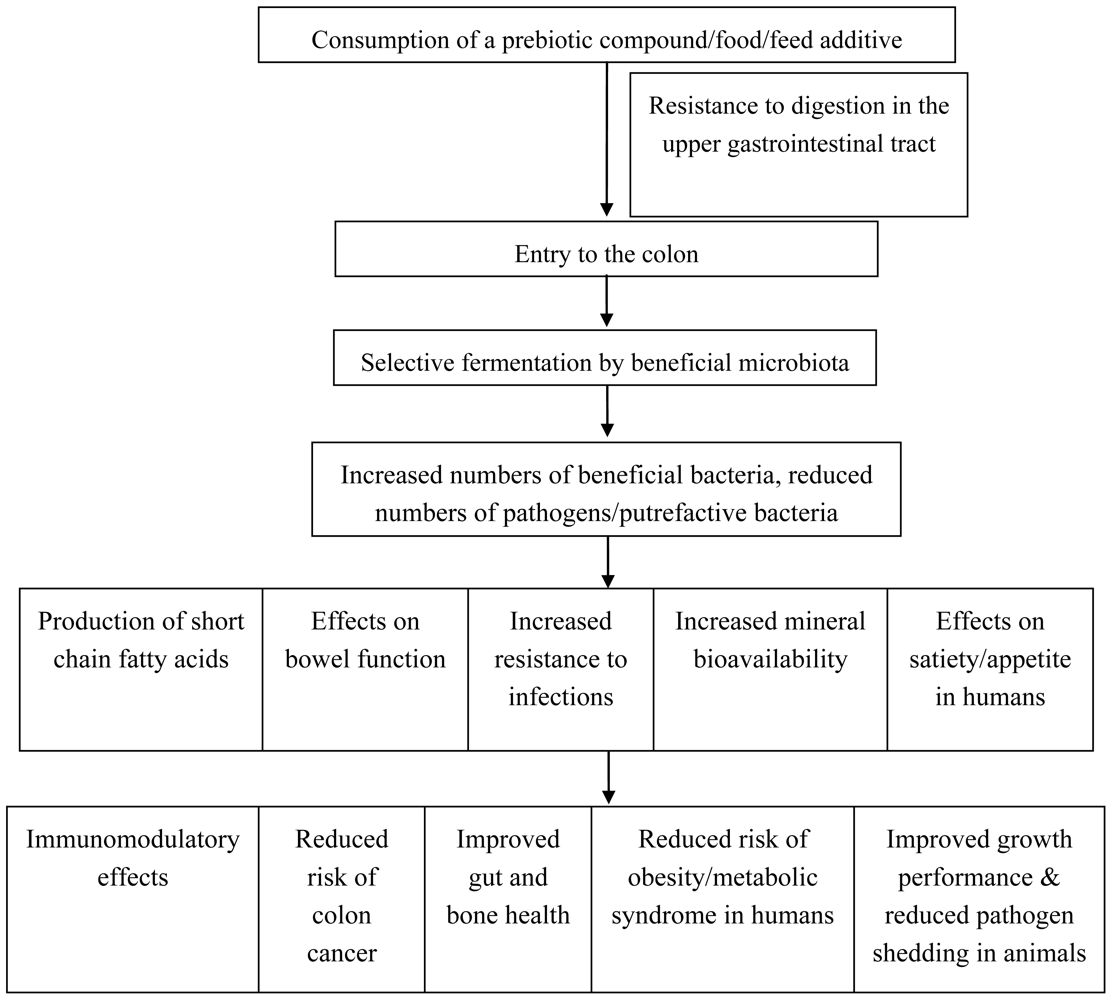
2. The Prebiotic Concept
Applications of prebiotics
3. Chemistry of Seaweed Polysaccharides
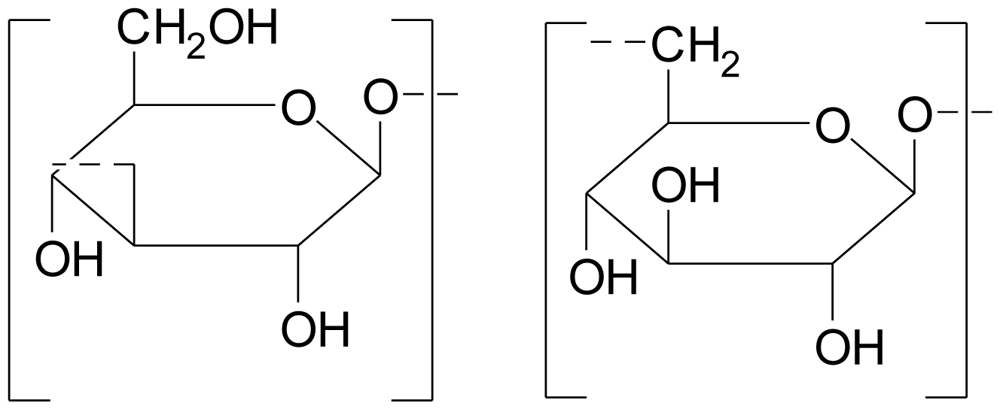
3.1. Polysaccharides from chlorophyta
Complex sulfated hetero-polysaccharides
3.2. Polysaccharides from phaeophyta
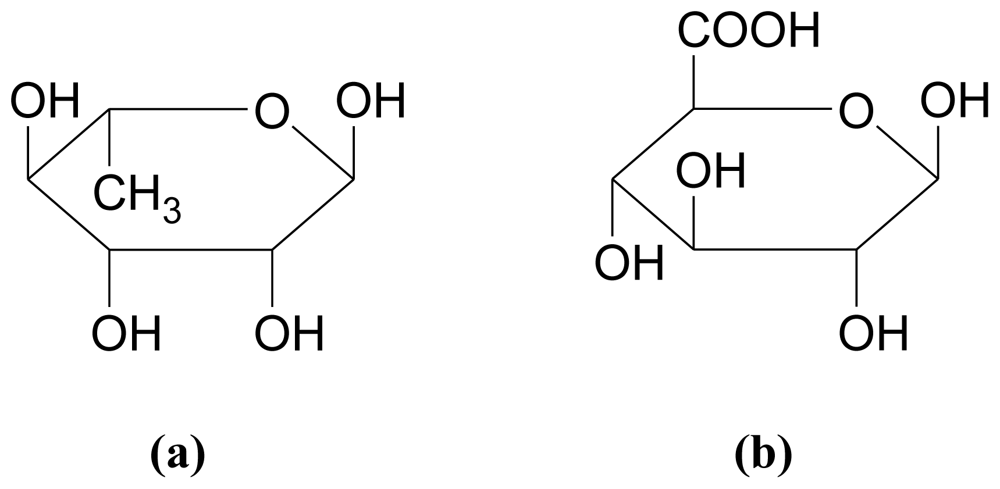
3.2.1. Alginates (also called alginic acid or algin)
- The seaweed is washed, macerated, extracted with sodium carbonate and filtered
- Sodium/calcium chloride is added to the filtrate and a fibrous precipitate of sodium/calcium alginate is formed
- The alginate salt is transformed to alginic acid by treatment with hydrochloric acid
- The alginate is purified, dried and powdered
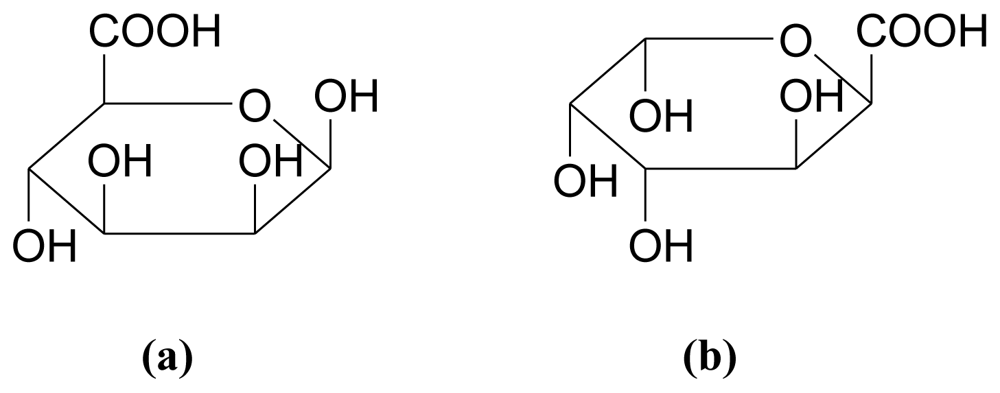
3.2.2. Fucans/Fucoidans
3.2.3. Laminarin
3.3. Polysaccharides from rhodophyta
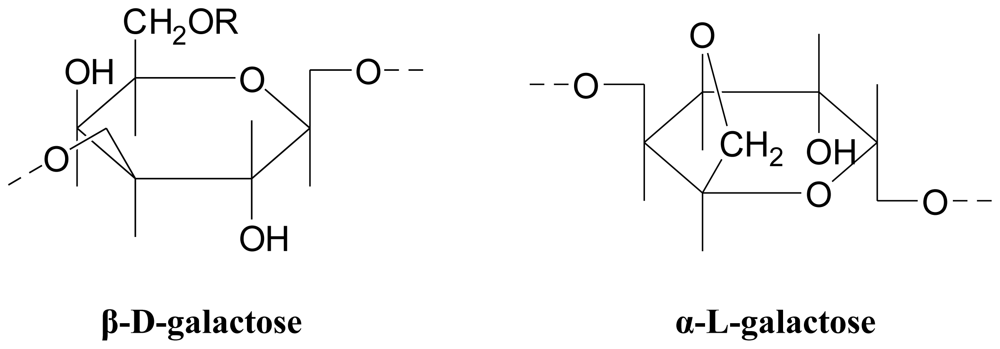
3.3.1. Agar
- Agarose (the gelling fraction)—A neutral linear molecule, free of sulfates
- Agaropectin (the non-gelling fraction)—Contains all the charged polysaccharide components, with some galactose residues substituted with pyruvic acid ketal, 4,6-O-(1-carboxyethylidene)D-galactopyranose, or methylated or sulfated sugar units [58]. Agaropectin is a slightly branched heterogeneous mixture of smaller molecules.
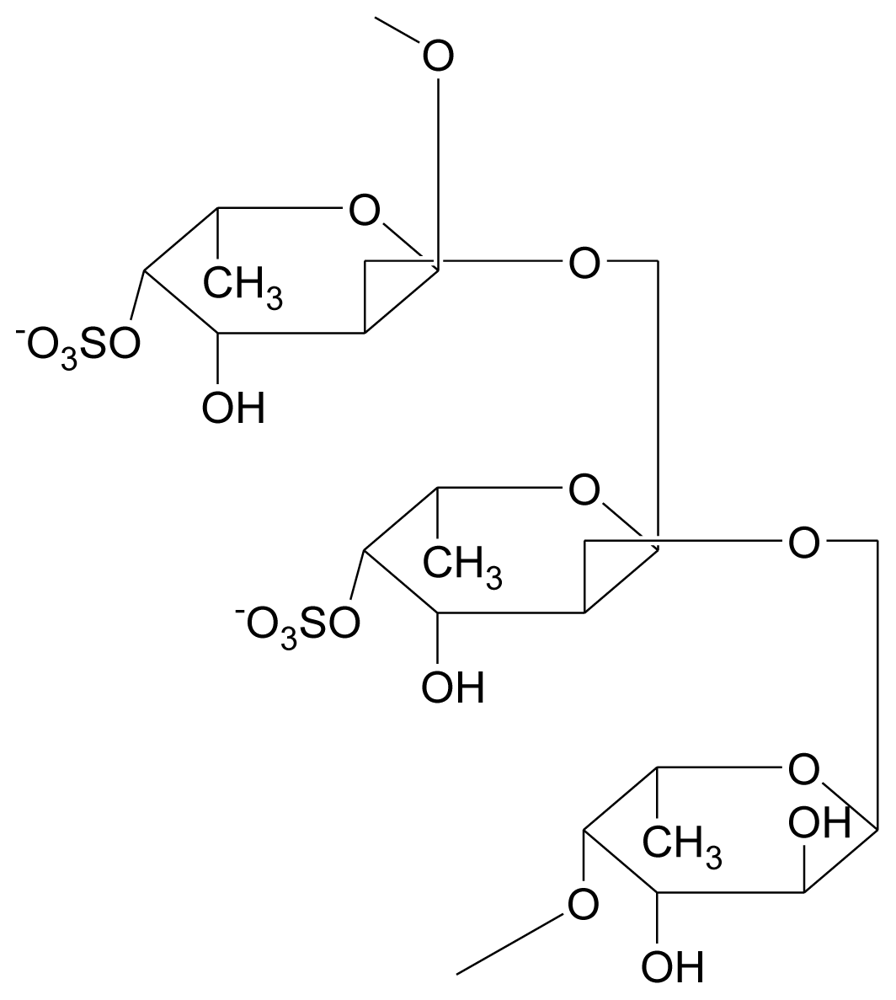
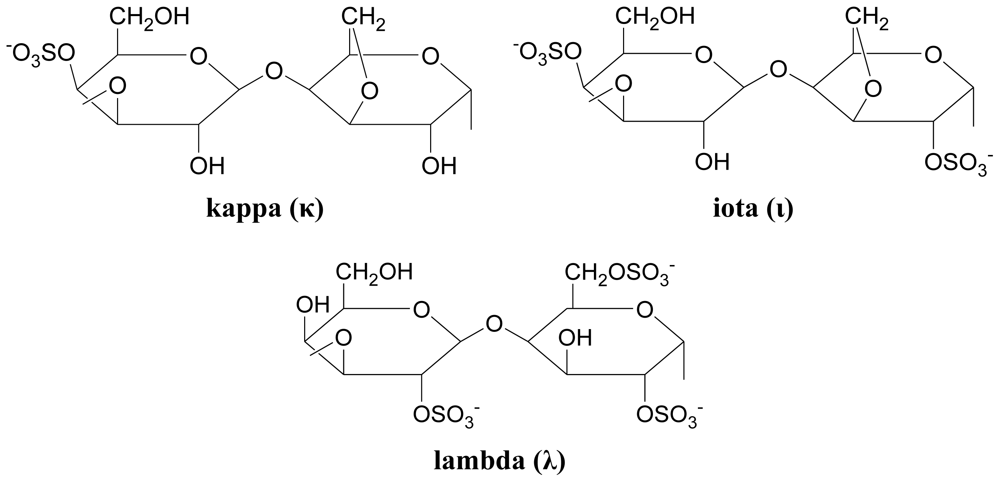
3.3.2. Carrageenans (sulfated polysaccharides)
3.3.3. Other polysaccharides and polysaccharide derivatives
- Hypneans are extracted from the Hypnea spp. Structurally, hypneans are similar to agar and carrageenan, but with a higher percentage of 3,6-anhydrogalactose. They are primarily used as gelling agents in food applications and as fertilizers in dry arid soils.
- Porphyran is a highly substituted polysaccharide extracted from the Porphyra genus. It is used as a gelling agent, a nutritional supplement (e.g., to help cope with stress) and an antioxidant.
- Funorans, extracted from species such as Gloiopeltis complanata, are composed of a heterogeneous series of polysaccharides and sulfated galactans. Funoran has been shown to inhibit the adherence and colonization of oral bacteria, reducing dental caries in rat studies [62]. It is also reported to reduce blood pressure, lower cholesterol and exhibit anti-tumor properties.
4. In Vitro Studies Examining the Prebiotic Potential of Seaweed Polysaccharides and Oligosaccharide Derivatives
4.1. Resistance to digestive enzymes
4.2. Selective fermentation by pure cultures
4.3. Fermenter studies to determine effects on intestinal microbiota
5. In Vivo Studies Examining the Prebiotic Potential of Macroalgal Polysaccharides and Oligosaccharide Derivatives
5.1. Studies in laboratory animals
5.2. Studies in farm animals
5.2.1. Effects of marine polysaccharides on growth performance
5.2.2. Effects on intestinal microflora
5.2.3. Additional benefits of prebiotics in swine husbandry
6. Conclusions
References
- Gonzalez del Val, A; Platas, G; Basilio, A; Cabello, A; Gorrochategui, J; Suay, I; Vicente, F; Portillo, E; Jimenez del Rio, M; Reina, GG; Pelaez, F. Screening of antimicrobial activities in red, green and brown macroalgae from Gran Canaria (Canary Islands, Spain). Int. Microbiol 2001, 4, 35–40. [Google Scholar]
- Yuan, YV; Walsh, NA. Antioxidant and antiproliferative activities of extracts from a variety of edible seaweeds. Food Chem. Toxicol 2006, 44, 1144–1150. [Google Scholar]
- Chandini, SK; Ganesan, P; Bhaskar, N. In vitro antioxidant activities of three selected brown seaweeds of India. Food Chem 2008, 107, 707–713. [Google Scholar]
- Kang, JY; Khan, MNA; Park, NH; Cho, JY; Lee, MC; Fujii, H; Hong, YK. Antipyretic, analgesic, and anti-inflammatory activities of the seaweed Sargassum fulvellum and Sargassum thunbergii in mice. J. Ethnopharmacol 2008, 116, 187–190. [Google Scholar]
- Pushpamali, WA; Nikapitiya, C; De Zoysa, M; Whang, I; Kim, SJ; Lee, J. Isolation and purification of an anticoagulant from fermented red seaweed Lomentaria catenata. Carbohydr. Polym 2008, 73, 274–279. [Google Scholar]
- Sinha, S; Astani, A; Ghosh, T; Schnitzler, P; Ray, B. Polysaccharides from Sargassum tenerrimum: structural features, chemical modification and anti-viral activity. Phytochemistry 2009, 71, 235–242. [Google Scholar]
- Kwon, MJ; Nam, TJ. Porphyran induces apoptosis related signal pathway in AGS gastric cancer cell lines. Life Sci 2006, 79, 1956–1962. [Google Scholar]
- d’Ayala, GG; Malinconico, M; Laurienzo, P. Marine derived polysaccharides for biomedical applications: Chemical modification approaches. Molecules 2008, 13, 2069–2106. [Google Scholar]
- Guo, JH; Skinner, GW; Harcum, WW; Barnum, PE. Pharmaceutical applications of naturally occurring water-soluble polymers. Pharm. Sci. Technol. Today 1998, 1, 254–261. [Google Scholar]
- Dhargalkar, VK; Verlecar, XN. Southern Ocean seaweeds: A resource for exploration in food and drugs. Aquaculture 2009, 287, 229–242. [Google Scholar]
- Jimenez-Escrig, A; Sanchez-Muniz, FJ. Dietary fiber from edible seaweeds: chemical structure, physicochemical properties and effects on cholesterol metabolism. Nut. Res 2000, 20, 585–598. [Google Scholar]
- Goni, I; Valdivieso, L; Gudiel-Urbano, M. Capacity of edible seaweeds to modify in vitro starch digestibility of wheat bread. Nahrung 2002, 46, 18–20. [Google Scholar]
- Haugan, JA; Liaaenjensen, S. Algal Carotenoids 54. Carotenoids of Brown-Algae (Phaeophyceae). Biochem. Syst. Ecol 1994, 22, 31–41. [Google Scholar]
- Robic, A; Gaillard, C; Sassi, JF; Lerat, Y; Lahaye, M. Ultrastructure of Ulvan: A Polysaccharide from Green Seaweeds. Biopolymers 2009, 91, 652–664. [Google Scholar]
- McHugh, DJ. A guide to seaweed industry. FAO Fish. Tech. Pap. 2003, T441, 118. [Google Scholar]
- Roberfroid, MB. Gibson, GR, Roberfroid, MB, Eds.; Prebiotics in Nutrition. In Handbook of Prebiotics, 1st ed; CRC Group: Boca Raton, FL, USA, 2008; pp. 1–11. [Google Scholar]
- Gibson, GR; Roberfroid, MB. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr 1995, 125, 1401–1412. [Google Scholar]
- Roberfroid, M. Gibson, GR, Roberfroid, M, Eds.; Prebiotics: Concept, definition, criteria, methodologies, and products. In Handbook of Prebiotics; CRC Press: Boca Raton, FL, USA, 2008; pp. 39–69. [Google Scholar]
- Courtois, J. Oligosaccharides from land plants and algae: production and applications in therapeutics and biotechnology. Curr. Opin. Microbiol 2009, 12, 261–273. [Google Scholar]
- Kolida, S; Gibson, GR. Gibson, GR, Roberfroid, M, Eds.; The prebiotic effect: Review of experimental and human data. In Handbook of Prebiotics; CRC Press: Boca Raton, FL, USA, 2008; pp. 69–93. [Google Scholar]
- Manning, TS; Gibson, GR. Prebiotics. Best Pract. Res. Clin. Gastroenterol 2004, 18, 287–298. [Google Scholar]
- Kelly, G. Inulin-Type Prebiotics: A Review (Part 2). Altern. Med. Rev 2009, 14, 36–55. [Google Scholar]
- Spiller, R. Review article: probiotics and prebiotics in irritable bowel syndrome. Aliment. Pharm. Therap 2008, 28, 385–396. [Google Scholar]
- Davis, CD; Milner, JA. Gastrointestinal microflora, food components and colon cancer prevention. J. Nut. Biochem 2009, 20, 743–752. [Google Scholar]
- Lomax, AR; Calder, PC. Prebiotics, immune function, infection and inflammation: a review of the evidence. Br. J. Nutr 2009, 101, 633–658. [Google Scholar]
- Tuohy, KM; Probert, HM; Smejkal, CW; Gibson, GR. Using probiotics and prebiotics to improve gut health. Drug Discov. Today 2003, 8, 692–700. [Google Scholar]
- Sanders, ME. Overview of Functional Foods: Emphasis on Probiotic Bacteria. Int. Dairy J 1998, 8, 341–347. [Google Scholar]
- Saulnier, DMA; Spinler, JK; Gibson, GR; Versalovic, J. Mechanisms of probiosis and prebiosis: considerations for enhanced functional foods. Curr. Opin. Biotechnol 2009, 20, 135–141. [Google Scholar]
- Wierup, M. The Swedish experience of the 1986 year ban of antimicrobial growth promoters, with special reference to animal health, disease prevention, productivity, and usage of antimicrobials. Microb. Drug Res 2001, 7, 183–190. [Google Scholar]
- Gaggia, F; Mattarelli, P; Biavati, B. Probiotics and prebiotics in animal feeding for safe foods production. Int. J. Food Microbiol 2010. [Google Scholar] [CrossRef]
- Percival, E; McDowell, RH. Chemistry and Enzymology of Marine Algal Polysaccharides; Academic Press: New York, NY, USA, 1967. [Google Scholar]
- Percival, E. Polysaccharides of Green, Red and Brown Seaweeds: Their Basic Structure, Biosynthesis and Function. Br. Phycol. J 1979, 14, 103–117. [Google Scholar]
- Rioux, LE; Turgeon, SL; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym 2007, 69, 530–537. [Google Scholar]
- Rinaudo, M. Kamerling, JP, Ed.; Comprehensive Glycoscience. In Seaweed Polysaccharides; Elsevier: Amsterdam, The Netherland, 2007; Volume 2. [Google Scholar]
- Pereira, L; Amado, AM; Critchley, AT; van de Velde, F; Ribeiro-Claro, PJA. Identification of selected seaweed polysaccharides (phycocolloids) by vibrational spectroscopy (FTIR-ATR and FT-Raman). Food Hydrocolloid 2009, 23, 1903–1909. [Google Scholar]
- Graham, LE; Wilcox, LW. Algae, 1st ed; Prentice Hall: Upper Saddle River, NJ, USA, 2000. [Google Scholar]
- Murphy, V. An investigation into the mechanisms of heavy metal binding by selected seaweed species. Ph.D. Thesis, Waterford Institute of Technology, Waterford, Ireland, 2007. [Google Scholar]
- Mckinnell, JP; Percival, E. Acid polysaccharide from green seaweed. Ulva-lactuca. J. Chem. Soc 1962, 2082–2083. [Google Scholar]
- Lahaye, M; Axelos, MAV. Gelling properties of water-soluble polysaccharides from proliferating marine green seaweeds (Ulva Spp). Carbohydr. Polym 1993, 22, 261–265. [Google Scholar]
- Paradossi, G; Cavalieri, F; Chiessi, E. A conformational study on the algal polysaccharide ulvan. Macromolecules 2002, 35, 6404–6411. [Google Scholar]
- Ray, B; Lahaye, M. Cell-wall polysaccharides from the marine green-alga Ulva-rigida (Ulvales, Chlorophyta) - chemical-structure of ulvan. Carbohydr. Res 1995, 274, 313–318. [Google Scholar]
- Andrieux, C; Hibert, A; Houari, AM; Bensaada, M; Popot, F; Szylit, O. Ulva lactuca is poorly fermented but alters bacterial metabolism in rats inoculated with human fecal flora from methane and non-methane producers. J. Sci. Food Agric 1998, 77, 25–30. [Google Scholar]
- Davis, TA; Volesky, B; Mucci, A. A review of the biochemistry of heavy metal biosorption by brown algae. Water Res 2003, 37, 4311–4330. [Google Scholar]
- Chapman, VJ; Chapman, DJ. Seaweeds and Their Uses; Chapman & Hall: London, UK, 1980. [Google Scholar]
- Fourest, E; Volesky, B. Contribution of sulfonate groups and alginate to heavy metal biosorption by the dry biomass of Sargassum fluitans. Environ. Sci. Technol 1996, 30, 277–282. [Google Scholar]
- Haug, A; Larsen, B; Smidsrod, O. Studies on sequence of uronic acid residues in alginic acid. Acta Chem. Scand 1967, 21, 691–704. [Google Scholar]
- Haug, A. Affinity of some divalent metals to different types of alginates. Acta Chem. Scand 1961, 15, 1794–1795. [Google Scholar]
- Mackie, W; Preston, RD. Stewart, WDP, Ed.; Algal physiology and biochemistry. In Cell Wall and Intercellular Region Polysaccharides; Blackwell Scientific Publications: Oxford, UK, 1974. [Google Scholar]
- Li, B; Lu, F; Wei, XJ; Zhao, RX. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar]
- Shibata, H; Iimuro, M; Uchiya, N; Kawamori, T; Nagaoka, M; Ueyama, S; Hashimoto, S; Yokokura, T; Sugimura, T; Wakabayashi, K. Preventive effects of Cladosiphon fucoidan against Helicobacter pylori infection in Mongolian gerbils. Helicobacter 2003, 8, 59–65. [Google Scholar]
- Dunstan, DE; Goodall, DG. Terraced self assembled nano-structures from laminarin. Int. J. Biol. Macromol 2007, 40, 362–366. [Google Scholar]
- Chizhov, AO; Dell, A; Morris, HR; Reason, AJ; Haslam, SM; McDowell, RA; Chizhov, OS; Usov, AI. Structural analysis of laminarans by MALDI and FAB mass spectrometry. Carbohydr. Res 1998, 310, 203–210. [Google Scholar]
- Neyrinck, AM; Mouson, A; Delzenne, NM. Dietary supplementation with laminarin, a fermentable marine beta (1–3) glucan, protects against hepatotoxicity induced by LPS in rat by modulating immune response in the hepatic tissue. Int. Immunopharmacol 2007, 7, 1497–1506. [Google Scholar]
- Deville, C; Gharbi, M; Dandrifosse, G; Peulen, O. Study on the effects of laminarin, a polysaccharide from seaweed, on gut characteristics. J. Sci. Food Agric 2007, 87, 1717–1725. [Google Scholar]
- Deville, C; Damas, J; Forget, P; Dandrifosse, G; Peulen, O. Laminarin in the dietary fiber concept. J. Sci. Food Agric 2004, 84, 1030–1038. [Google Scholar]
- Marinho-Soriano, E. Agar polysaccharides from Gracilaria species (Rhodophyta, Gracilariaceae). J. Biotech 2001, 89, 81–84. [Google Scholar]
- Falshaw, R; Furneaux, RH; Stevenson, DE. Agars from nine species of red seaweed in the genus Curdiea (Gracilariaceae, Rhodophyta). Carbohydr. Res 1998, 308, 107–115. [Google Scholar]
- Labropoulos, KC; Niesz, DE; Danforth, SC; Kevrekidis, PG. Dynamic rheology of agar gels: theory and experiments. Part I. Development of a rheological model. Carbohydr. Polym 2002, 50, 393–406. [Google Scholar]
- Hu, B; Gong, QH; Wang, Y; Ma, YM; Li, JB; Yu, WG. Prebiotic effects of neoagaro-oligosaccharides prepared by enzymatic hydrolysis of agarose. Anaerobe 2006, 12, 260–266. [Google Scholar]
- Falshaw, R; Bixler, HJ; Johndro, K. Structure and performance of commercial kappa-2 carrageenan extracts I. Structure analysis. Food Hydrocollid 2001, 15, 441–452. [Google Scholar]
- Errea, MI; Matulewicz, MC. Unusual structures in the polysaccharides from the red seaweed Pterocladiella capillacea (Gelidiaceae, Gelidiales). Carbohydr. Res 2003, 338, 943–953. [Google Scholar]
- Saeki, Y; Kato, T; Naito, Y; Takazoe, I; Okuda, K. Inhibitory effects of funoran on the adherence and colonization of mutans streptococci. Caries Res 1996, 30, 119–125. [Google Scholar]
- Muraoka, T; Ishihara, K; Oyamada, C; Kunitake, H; Hirayama, I; Kimura, T. Fermentation properties of low-quality red alga susabinori Porphyra yezoensis by intestinal bacteria. Biosci. Biotechnol. Biochem 2008, 72, 1731–1739. [Google Scholar]
- Salyers, AA; Vercellotti, JR; West, SEH; Wilkins, TD. Fermentation of mucin and plant polysaccharides by strains of bacteroides from the human colon. Appl. Environ. Microbiol 1977, 33, 319–322. [Google Scholar]
- Salyers, AA; West, SEH; Vercellotti, JR; Wilkins, TD. Fermentation of mucins and plant polysaccharides by anaerobic bacteria from the human colon. Appl. Environ. Microbiol 1977, 34, 529–533. [Google Scholar]
- Coles, LT; Moughan, PJ; Darragh, AJ. In vitro digestion and fermentation methods, including gas production techniques, as applied to nutritive evaluation of foods in the hindgut of humans and other simple-stomached animals. Anim. Feed Sci. Tech 2005, 123, 421–444. [Google Scholar]
- Michel, C; Macfarlane, GT. Digestive fates of soluble polysaccharides from marine macroalgae: Involvement of the colonic microflora and physiological consequences for the host. J. Appl. Bacteriol 1996, 80, 349–369. [Google Scholar]
- Kuda, T; Yano, T; Matsuda, N; Nishizawa, M. Inhibitory effects of laminaran and low molecular alginate against the putrefactive compounds produced by intestinal microflora in vitro and in rats. Food Chem 2005, 91, 745–749. [Google Scholar]
- Dierick, N; Ovyn, A; De Smet, S. Effect of feeding intact brown seaweed Ascophyllum nodosum on some digestive parameters and on iodine content in edible tissues in pigs. J. Sci. Food Agric 2009, 89, 584–594. [Google Scholar]
- Michel, C; Benard, C; Lahaye, M; Formaglio, D; Kaeffer, B; Quemener, B; Berot, S; Yvin, JC; Blottiere, HM; Cherbut, C. Algal oligosaccharides as functional foods: in vitro study of their cellular and fermentative effects. Food Sci 1999, 19, 311–332. [Google Scholar]
- Marzorati, M; Verhelst, A; Luta, G; Sinnott, R; Verstraete, W; Van de Wiele, T; Possemiers, S. In vitro modulation of the human gastrointestinal microbial community by plant-derived polysaccharide-rich dietary supplements. Int. J. Food Microbiol 2010, 139, 168–176. [Google Scholar]
- Wang, Y; Han, F; Hu, B; Li, JB; Yu, WG. In vivo prebiotic properties of alginate oligosaccharides prepared through enzymatic hydrolysis of alginate. Nut. Res 2006, 26, 597–603. [Google Scholar]
- Gudiel-Urbano, M; Goni, I. Effect of edible seaweeds (Undaria pinnatifida and Porphyra ternera) on the metabolic activities of intestinal microflora in rats. Nut. Res 2002, 22, 323–331. [Google Scholar]
- Pluske, JR; Hampson, DJ; Williams, IH. Factors influencing the structure and function of the small intestine in the weaned pig: a review. Livestock Prod. Sci 1997, 51, 215–236. [Google Scholar]
- Estrada, A; Drew, MD; Van Kessel, A. Effect of the dietary supplementation of fructooligosaccharides and Bifidobacterium longum to early-weaned pigs on performance and fecal bacterial populations. Can. J. Anim. Sci 2001, 81, 141–148. [Google Scholar]
- Reilly, P; O’Doherty, JV; Pierce, KM; Callan, JJ; O’Sullivan, JT; Sweeney, T. The effects of seaweed extract inclusion on gut morphology, selected intestinal microbiota, nutrient digestibility, volatile fatty acid concentrations and the immune status of the weaned pig. Animal 2008, 2, 1465–1473. [Google Scholar]
- Bach, SJ; Wang, Y; McAllister, TA. Effect of feeding sun-dried seaweed (Ascophyllum nodosum) on fecal shedding of Escherichia coli O157: H7 by feedlot cattle and on growth performance of lambs. Anim. Feed Sci. Tech 2008, 142, 17–32. [Google Scholar]
- Gardiner, GE; Campbell, AJ; O’Doherty, JV; Pierce, E; Lynch, PB; Leonard, FC; Stanton, C; Ross, RP; Lawlor, PG. Effect of Ascophyllum nodosum extract on growth performance, digestibility, carcass characteristics and selected intestinal microflora populations of grower-finisher pigs. Anim. Feed Sci. Tech 2008, 141, 259–273. [Google Scholar]
- Gahan, DA; Lynch, MB; Callan, JJ; O’Sullivan, JT; O’Doherty, JV. Performance of weanling piglets offered low-, medium- or high-lactose diets supplemented with a seaweed extract from Laminaria spp. Animal 2009, 3, 24–31. [Google Scholar]
- McDonnell, P; Figat, S; O’Doherty, JV. The effect of dietary laminarin and fucoidan in the diet of the weanling piglet on performance, selected fecal microbial populations and volatile fatty acid concentrations. Animal 2010, 4, 579–585. [Google Scholar]
- O’Doherty, JV; Dillon, S; Figat, S; Callan, JJ; Sweeney, T. The effects of lactose inclusion and seaweed extract derived from Laminaria spp. on performance, digestibility of diet components and microbial populations in newly weaned pigs. Anim. Feed Sci. Tech 2010, 157, 173–180. [Google Scholar]
- Turner, JL; Dritz, SS; Higgins, JJ; Minton, JE. Effects of Ascophyllum nodosum extract on growth performance and immune function of young pigs challenged with Salmonella typhimurium. J. Anim. Sci 2002, 80, 1947–1953. [Google Scholar]
- Lynch, MB; Sweeney, T; Callan, JJ; O’Sullivan, JT; O’Doherty, JV. The effect of dietary Laminaria-derived laminarin and fucoidan on nutrient digestibility, nitrogen utilization, intestinal microflora and volatile fatty acid concentration in pigs. J. Sci. Food Agric 2010, 90, 430–437. [Google Scholar]
- Janczyk, P; Pieper, R; Smidt, H; Souffrant, WB. Effect of alginate and inulin on intestinal microbial ecology of weanling pigs reared under different husbandry conditions. FEMS Microbiol. Ecol 2010, 72, 132–142. [Google Scholar]
- Braden, KW; Blanton, JR; Allen, VG; Pond, KR; Miller, MF. Ascophyllum nodosum supplementation: A preharvest intervention for reducing Escherichia coli O157:H7 and Salmonella spp. in feedlot stears. J. Food Prot 2004, 67, 1824–1828. [Google Scholar]
- Awati, A; Williams, BA; Bosch, MW; Gerrits, WJJ; Verstegen, MWA. Effect of inclusion of fermentable carbohydrates in the diet on fermentation end-product profile in feces of weanling piglets. J. Anim. Sci 2006, 84, 2133–2140. [Google Scholar]
- Yokoyama, MT; Carlson, JR. Microbial metabolites of tryptophan in the intestinal-tract with special reference to skatole. Am. J. Clin. Nutr 1979, 32, 173–178. [Google Scholar]
- Lanthier, F; Lou, Y; Terner, MA; Squires, EJ. Characterizing developmental changes in plasma and tissue skatole concentrations in the prepubescent intact male pig. J. Anim. Sci 2006, 84, 1699–1708. [Google Scholar]
- Byrne, DV; Thamsborg, SM; Hansen, LL. A sensory description of boar taint and the effects of crude and dried chicory roots (Cichorium intybus L.) and inulin feeding in male and female pork. Meat Sci 2008, 79, 252–269. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seaweed | Structure of Laminarin |
|---|---|
| Several species of Laminaria | Linear β-(1,3) linked D-glucose |
| Laminaria digitata | Linear backbone of β-(1,3) linked D-glucose, with β-(1,6) linked side chains |
| Eisenia bicyclis | Linear chain of (1–3) and (1–6) links, in the ratio of 2:1 |
| Algal supplement | Age and heath status of pigs | Dose | Effect on gut microbiota | Effect on growth performance and health | Ref |
|---|---|---|---|---|---|
| A. nodosum extract (ANE) | Healthy grower-finisher pigs | 0, 3, 6 or 9 g/kg basal feed | Animals supplemented with 6 or 9 g ANE/kg had lower (P < 0.05) ileal coliform counts than animals that received 3 g/kg Linear reduction (P < 0.05) in coliform counts in the ileal contents as ANE increased Reduction (P < 0.05) in cecal Bifidobacterium counts with increasing ANE supplementation | Linear decrease (P < 0.05) in weight gain with increasing levels of extract | [78] |
| A. nodosum meal (ANM) | Healthy, weanling piglets | 10 or 20 g/kg basal feed | Reduced (P < 0.05)E. coli in the small intestine and increased (P< 0.05) Lactobacillus/E. coli ratio in animals fed 10 g/kg | No effects on final weight No effects on intestinal histology or intestinal immune cells | [69] |
| Laminaria spp. extract containing a combination of laminarin & fucoidan (ranging from 0.112–0.446 and 0.890–0.356 g/kg, respectively) | Healthy, weanling piglets | 0, 1, 2, 4 g/kg basal feed containing increasing levels of lactose (60–250 g/kg) | Effects on gut microbiota were not determined | Weight gain and feed intake increased (P < 0.05) as the level of seaweed extract increased; however, this was only observed when fed in combination with low and medium levels of lactose | [79] |
| Laminaria spp. extract containing either laminarin or fucoidan or a combination of both (0.3 and 0.24 g/kg, respectively) | Healthy, weanling piglets fed a nutritionally-challenged diet (high protein, low lactose) | Basal feed + 0.3 g/kg laminarin; basal feed + 0.24 g/kg fucoidan; basal feed + 0.3 g/kg laminarin and 0.24 g/kg fucoidan | Laminarin supplementation resulted in lower (P < 0.05) fecal E. coli populations compared to control group Interaction (P < 0.01) between laminarin and fucoidan with respect to fecal lactobacilli populations | Laminarin supplementation resulted in increased (P < 0.01) daily weight gain Pigs offered combination of laminarin and fucoidan had reduced (P < 0.05) diarrhoea | [80] |
| Alginate | Healthy, weanling piglets | 1 g/kg starter feed | Higher enterococci counts in distal small intestine, cecum and proximal colon (P < 0.001) compared with inulin or control group. Reduced (P < 0.05) lactobacilli in all intestinal segments but only after 6 days of alginate supplementation, and not before or thereafter Increased microbial diversity | Animals were in good health throughout the study | [84] |
| Exp 1: L. hyperborea extract (112 g/kg laminarin & 89 g/kg fucoidan) Exp 2: Purified laminarin (0.30 g/kg), fucoidan (0.24 g/kg) and a combination of both laminarin and fucoidan (0.30 and 0.24 g/kg, respectively) | Healthy finishing boars | Exp 1:0.7, 1.4, 2.8, 5.6 g/kg extract Exp 2: Basal diet + 0.30 g/kg laminarin; basal diet + 0.24 g/kg fucoidan; basal diet + 0.30 g/kg laminarin and 0.24 g/kg fucoidan | Exp 1: Quadratic response (P < 0.05) to seaweed extract on cecal (P < 0.05) Enterobacterium spp., colonic (P < 0.05) Enterobacterium spp. and (P < 0.001) Bifidobacterium spp. Linear decrease in cecal Bifidobacterium spp and colonic Lactobacillus spp. with increasing seaweed extract (P < 0.01, P < 0.05, respectively) Exp 2: Fucoidan diet resulted in increases in colonic Lactobacillus spp. (P < 0.001). Combination diet resulted in increase in Enterobacterium spp. (P < 0.05). | Growth performance was not evaluated | [83] |
| L. hyperborea extract (LHE), containing laminarin and fucoidan (0.17 and 0.13 g/kg, respectively) L. digitata extract (LDE), containing laminarin and fucoidan (0.17 and 0.14 g/kg, respectively) Combination of LHE and LDE containing laminarin and fucoidan (0.17 and 0.13 g/kg, respectively) | Healthy, weanling piglets | Basal feed + 1.5 g/kg LHE Basal feed + 1.5 g/kg LDE Basal feed + 1.5 g/kg LHE & LDE | Animals offered LHE diet had lower (P < 0.05) numbers of colonic Bifidobacterium and lower populations of cecal and colonic (P < 0.05, P < 0.001, respectively) lactobacilli compared to control diet Supplementation with LDE resulted in lower populations of cecal and colonic (P < 0.05) Enterobacterium, cecal (P < 0.05) Bifidobacterium and cecal and colonic (P < 0.05, P < 0.001, respectively) Lactobacillus compared to control diet Animals offered combination diet had lower (P < 0.05) populations of colonic and cecal Enterobacterium and Lactobacillus (P < 0.01) compared to control diet | No effects on animal performance Marginal differences in systemic immune response reported in animals fed combination diet | [76] |
| L. digitata extract containing laminarin (0.11 g/kg), and fucoidan (0.89 g/kg) | Healthy, weanling piglets fed a nutritionally-challenged diet | Diet 1: 150g lactose (L)/kg Diet 2: 150 g/kg lactose + 2.8 g/kg seaweed extract (SE) Diet 3: 250g lactose/kg Diet 4: 250g lactose/kg + 2.8 g/kg SE | The inclusion of SE decreased (P < 0.05) fecal E. coli counts compared to non-SWE diets Dietary inclusion of SE increased (P < 0.001) Lactobacillus counts in pigs fed high L diets | Animals offered seaweed diets had higher (P < 0.01) average daily gain and gain to feed ratio (P < 0.05) Fecal score not affected by dietary inclusion of SE | [81] |
Abbreviations
| GIT | Gastrointestinal tract |
| SCFA | short-chain fatty acids |
| NAOS | Neoagaro-oligosaccharides |
| FOS | Fructo-oligosaccharides |
| FISH | Fluorescence in situ hybridization |
| PCR | Polymerase chain reaction |
| DGGE | Denaturing gradient gel electrophoresis |
| qPCR | Quantitative PCR |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
O’Sullivan, L.; Murphy, B.; McLoughlin, P.; Duggan, P.; Lawlor, P.G.; Hughes, H.; Gardiner, G.E. Prebiotics from Marine Macroalgae for Human and Animal Health Applications. Mar. Drugs 2010, 8, 2038-2064. https://doi.org/10.3390/md8072038
O’Sullivan L, Murphy B, McLoughlin P, Duggan P, Lawlor PG, Hughes H, Gardiner GE. Prebiotics from Marine Macroalgae for Human and Animal Health Applications. Marine Drugs. 2010; 8(7):2038-2064. https://doi.org/10.3390/md8072038
Chicago/Turabian StyleO’Sullivan, Laurie, Brian Murphy, Peter McLoughlin, Patrick Duggan, Peadar G. Lawlor, Helen Hughes, and Gillian E. Gardiner. 2010. "Prebiotics from Marine Macroalgae for Human and Animal Health Applications" Marine Drugs 8, no. 7: 2038-2064. https://doi.org/10.3390/md8072038