Cloning and Comparative Studies of Seaweed Trehalose-6-Phosphate Synthase Genes

Abstract
:Introduction
2. Materials and Methods
2.1. Seaweed materials
2.2. Generation of the TPS gene from P. yezoensis
2.3. DNA extraction
2.4. Total RNA extraction and cDNA syntheses
2.5. PCR amplification of TPS genes from other nine seaweed species
2.6. Analyses and comparison of TPS genes
2.7. Cloning the PyTPS gene into vector pET22b
3. Results
3.1. Generation of full-length PyTPS Cdna
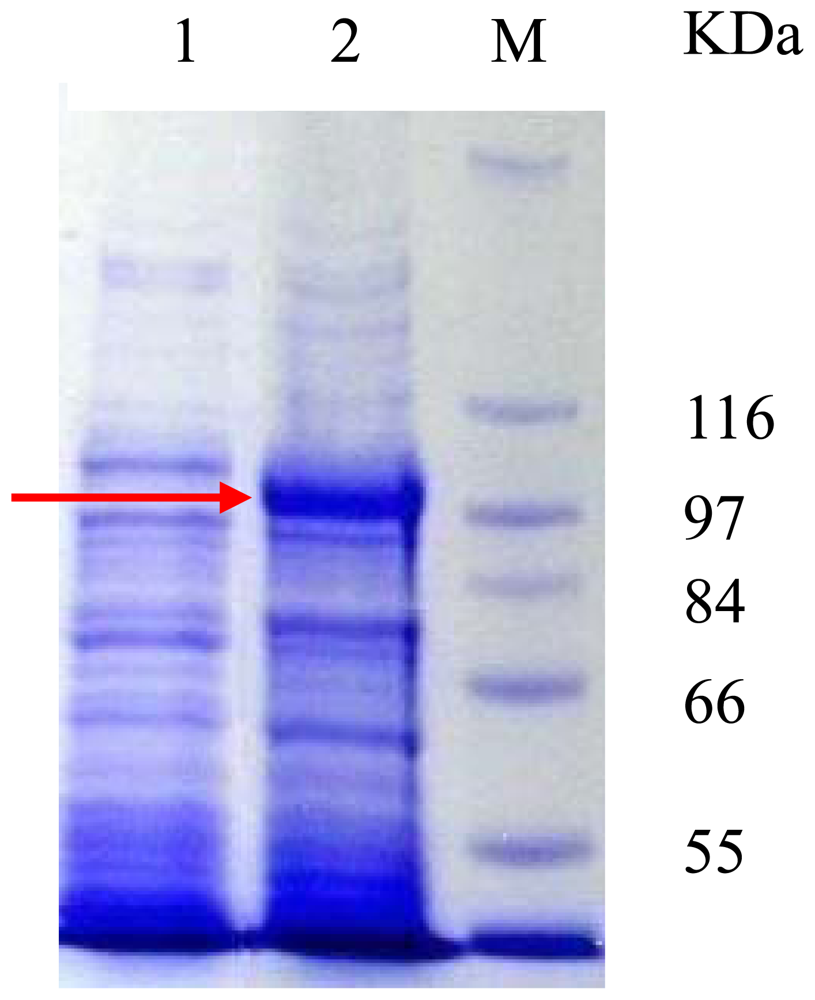
3.2. Expression of the PyTPS gene in E. coli
3.3. Domain analysis of the PyTPS protein
3.4. Comparison of PyTPS with TPS proteins from other organisms
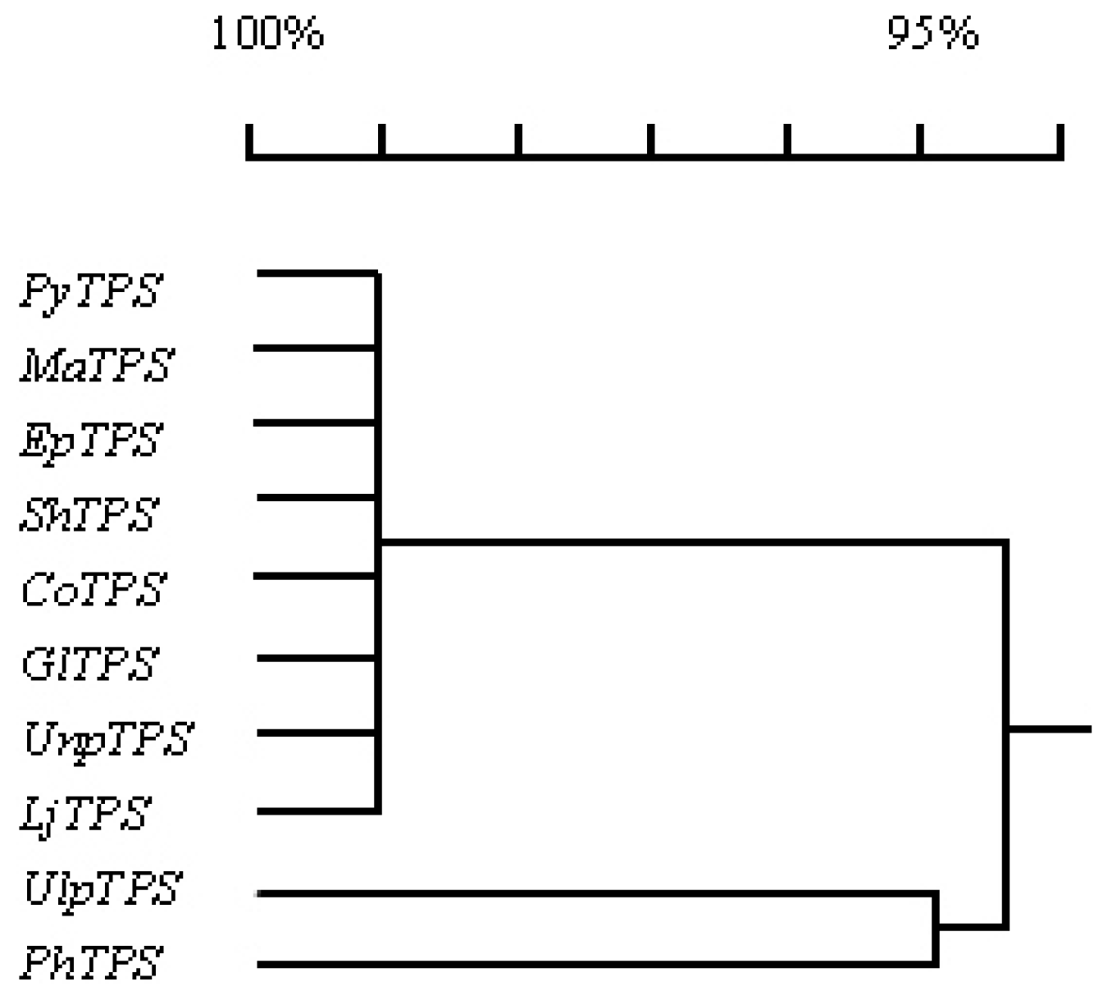
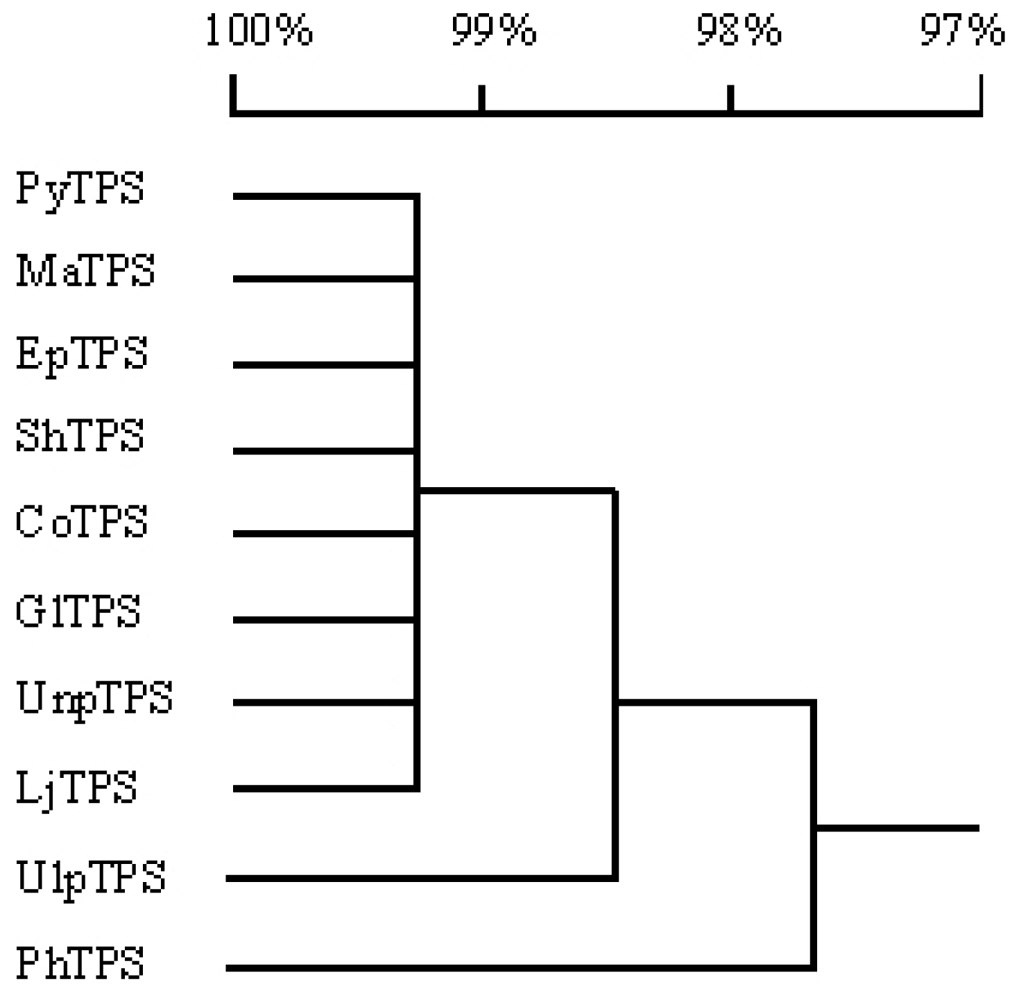
3.5. Comparison of PyTPS with TPS genes from other nine seaweeds
3.6. Comparison of the TPS gene sequences from cDNA and genomic DNA
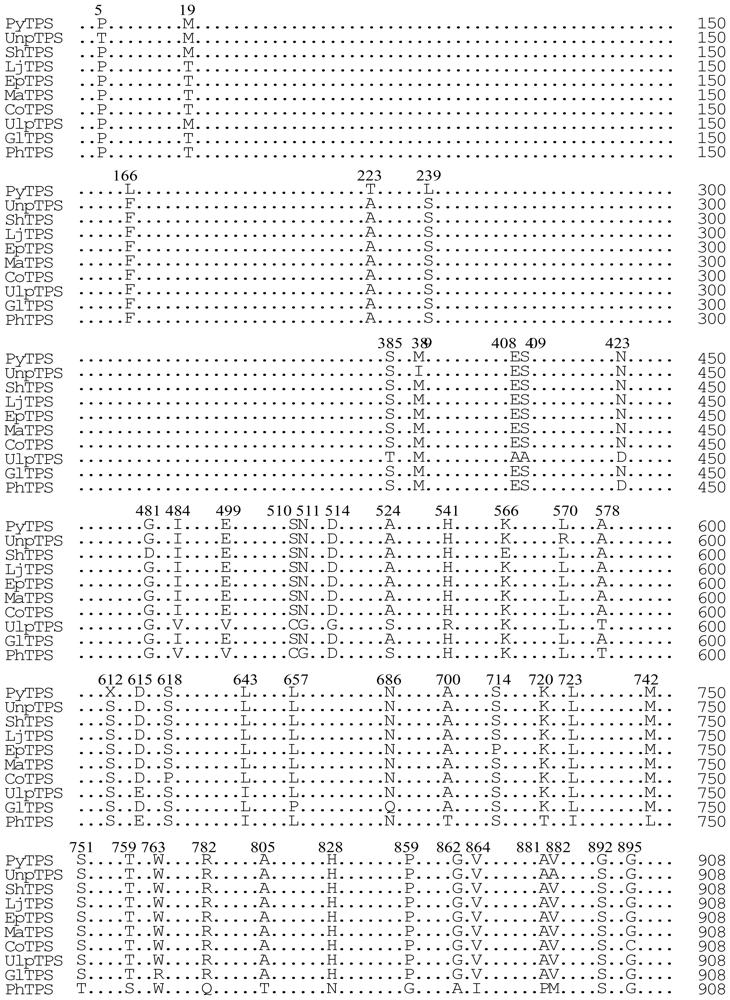
3.7. Comparison of the TPS proteins from the 10 seaweed species
4. Discussion
Acknowledgements
References
- Zhang, XC. Zhang, XC, Qin, S, Ma, JH, Xu, P, Eds.; Introduction. In The Genetics of Marine Algae; Chinese Agriculture Press: Beijing, China, 2005; pp. 1–14, (In Chinese with English title and content list). [Google Scholar]
- Chan, CX; Ho, CL; Phang, SM. Trends in seaweed research. Trends Plant Sci 2006, 11, 165–166. [Google Scholar]
- Goeme, A. A small introduction on bioenergy and alage: the new biofuel? Available online: http://algaetobioenergy.wordpress.com (accessed on 14 June 2008).
- Reith, H; Huijgen, W; Hal, JV; Lenstra, J. Seaweed potential in the Netherlands. ECN report, 2 June 2009. Available online: http://www.supergen-bioenergy.net/Resources/user/docs/new/Hans%20Reith-Seaweed%potertial%20in%20NL-%20Macroalgae-%20Seaweed%20Bioenergy%20Research%20Forum%202nd%20June%2009%Plymouth.pdf (accessed on 1 October 2009).
- Qin, S; Jiang, P; Tseng, CK. Molecular biotechnology of marine algae in China. Hydrobiologia 2004, 512, 21–26. [Google Scholar]
- Sahoo, D; Tang, XR; Yarish, C. Porphyra—the economic seaweed as a new experimental system. Curr Sci 2002, 83, 1313–1316. [Google Scholar]
- Waaland, JR; Stiller, JW; Cheney, DP. Macroalgal candidates for genomics. J. Physiol 2004, 40, 26–33. [Google Scholar]
- Goddijn, OJM; Smeekens, S. Sensing trehalose biosynthesis in plants. Plant J 1998, 14, 143–146. [Google Scholar]
- Lowe, RGT; Lord, M; Rybak, K; Trengove, RD; Oliver, RP; Solomon, PS. Trehalose biosynthesis is involved in sporulation of Stagonospora nodorum. Fungal Genet. Biol 2009, 46, 381–389. [Google Scholar]
- Goddijn, OJM; van Dun, K. Trehalose metabolism in plants. Trends Plant Sci 1999, 4, 315–319. [Google Scholar]
- Eastmond, PJ; Van Dijken, AJH; Spielman, M; Kerr, A; Tissier, AF; Dickinson, HG; Jones, JDG; Smeekens, SC; Graham, IA. Trehalose-6-phosphate synthase 1, which catalyses the first step in trehalose synthesis, is essential for Arabidopsis embryo maturation. Plant J 2002, 29, 225–235. [Google Scholar]
- Schluepmann, H; Pellny, T; van Dijken, A; Smeekens, S; Paul, M. Trehalose-6-phosphate is indispensable for carbohydrate utilization and growth in Arabidopsis thaliana. Proc. Natal. Acad. Sci. USA 2003, 100, 6849–6854. [Google Scholar]
- Zhang, Y; Primavesi, LF; Jhurreea, D; Andralojc, PJ; Mitchell, RAC; Powers, SJ; Schluepmann, H; Delatte, T; Wingler, A; Paul, MJ. Inhibition of SNF1-related protein kinase1 activity and regulation of metabolic pathways by trehalose-6-phosphate. Plant Physiol 2009, 149, 1860–1871. [Google Scholar]
- Kaasen, I; McDougall, J; Strom, AR. Analysis of the otsBA operon for osmoregulatory trehalose synthesis in Escherichia coli and homology of the OtsA and OtsB proteins to the yeast trehalose-6-phosphate synthasephosphatase complex. Gene 1994, 145, 9–15. [Google Scholar]
- Cai, ZJ; Peng, GX; Cao, YQ; Liu, YC; Jin, K; Xia, YX. Trehalose-6-phosphate synthase 1 from Metarhizium anisopliae: Clone, expression and properties of recombinants. J. Biosci. Bioengineering 2009, 107, 499–505, (In Chinese with English abstract). [Google Scholar]
- Vuorio, OE; Kalkkinen, N; Londesborough, J. Cloning of two related genes encoding the 56-kDa and 123-kDa subunits of trehalose synthase from the yeast Saccharomyces cerevisiae. Eur. J. Biochem 1993, 216, 849–861. [Google Scholar]
- De Virgilio, C; Burckert, N; Bell, W; Jeno, P; Boller, T; Wiemken, A. The role of trehalose synthesis for the acquisition of thermotolerance in yeast. I. Genetic evidence that trehalose is a thermoprotectant. Eur. J. Biochem 1993, 212, 315–323. [Google Scholar]
- Schluepmann, H; van Dijken, A; Aghdasi, M; Wobbes, B; Paul, M; Smeekens, S. Trehalose mediated growth inhibition of Arabidopsis seedlings is due to trehalose-6-phosphate accumulation. Plant Physiol 2004, 135, 879–890. [Google Scholar]
- Smeekens, S. Sugar signals and molecular networks controling plant growth. Curr. Opin. Plant Biol 2009, 13, 1–6. [Google Scholar]
- Vandesteene, L; Ramon, M; Roy, KL; Van Dijck, P; Rolland, F. A single active trehalose-6-phosphate synthase (TPS) and a family of putative regulatory TPS-like protein in Arabidopsis thaliana. Mol. Plant 2010, 3, 406–419. [Google Scholar]
- Blazque, MA; Santos, E; Flores, CL; Martinez-Zapater, JM; Salinas, J; Gancedo, C. Isolation and molecular characterization of the Arabidopsis TPS1 gene, encoding trehalose-6-phosphate synthase. Plant J 1998, 13, 685–689. [Google Scholar]
- Vogel, G; Aeschbacher, RA; Muller, J; Boller, T; Wiemken, A. Trehalose-6-phosphate phosphatases from Arabidopsis thaliana: identification by functional complementation of the yeast tps2 mutant. Plant J 1998, 13, 673–683. [Google Scholar]
- Zentella, R; Mascorro-Gallardo, JO; Van Dijck, P; Folch-Mallol, J; Bonini, B; Van Vaeck, C; Gaxiola, R; Covarrubias, AA; Nieto-Sotelo, J; Thevelein, JM; Iturriaga, GA. Selaginella lepidophylla trehalose-6-phosphate synthase complements growth and stress-tolerance defects in a yeast tps1 mutant. Plant Physiol 1999, 119, 1473–1482. [Google Scholar]
- Eastmond, P; Li, Y; Graham, IA. Is trehalose-6-phosphate a regulator of sugar metabolism in plants? J. Exp. Bot 2003, 54, 533–537. [Google Scholar]
- Holmstrom, KO; Mantyla, E; Welin, B; Mananal; Palva, ET; Tunnela, OE; Londesborough, J. Drought tolerance in tobacco. Nature 1996, 379, 683–684. [Google Scholar]
- Pilon-Smits, EAM; Terry, N; Sears, T; Kim, H; Zayed, A; Hwang, S; van Dun, K; Voogd, E; Verwoerd, TC; Krutwagen, RW. Trehalose producing transgenic tobacco plants show improved growth performance under drought stress. J. Plant Physiol 1998, 152, 525–532. [Google Scholar]
- Garg, AK; Kim, JK; Owens, TG; Ranwala, AP; Choi, YD; Kochian, LV; Wu, RJ. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. USA 2002, 99, 15898–15903. [Google Scholar]
- Jeong, SC; Pack, IS; Cho, EY; Youk, ES; Park, S; Yoon, WK; Kim, CG; Choi, YD; Kim, JK; Kim, HM. Molecular analysis and quantitative detection of a transgenic rice line expressing a bifunctional fusion TPSP. Food Control 2007, 18, 1434–1442. [Google Scholar]
- Mu, Y; He, J; Fu, FL; Li, WC. Transformation of maize with trehalose synthase gene (TPS1) cloning from Saccharomyces cerevisiae. J. Nucl. Agric. Sci 2007, 21, 430–435, (In Chinese with English abstract). [Google Scholar]
- Jia, WL; Hu, YL; Zhang, YQ; Yang, LL; Lin, ZP; Wu, Q. Transformation of trehalose synthase gene (TPS gene) into Perennial Ryegrass and identification of drought tolerance. Mol. Plant Breed 2007, 5, 27–31, (In Chinese with English abstract). [Google Scholar]
- Provasoli, L. Media and prospects for the cultivation of marine algae. Culture and Collections of Algae, Proc US-Japan Conf. Hakone, September 1966; Watanabe, A, Hattori, A, Eds.; Japonica Society of Plant Physiology: Hakone, Japan, 1968; pp. 63–75. [Google Scholar]
- Chomczynski, P; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem 1987, 162, 156–159. [Google Scholar]
- Deng, DY; Xuan, JS; Yang, JL; Duan, DL; Weng, ML; Wang, B. Construction and characterization of a bacterial artificial chromosome (BAC) library of marine macroalga Porphyra yezoensis (Rhodophyta). Plant Mol. Biol. Rep 2004, 22, 375–386. [Google Scholar]
- Sun, JW; Jin, DM; Zhou, CJ; Yang, QK; Weng, ML; Duan, DL; Xu, P; Ma, JH; Wang, B. Identification of Porphyra lines (Rhodophyta) by AFLP-DNA fingerprinting and molecular markers. Plant Mol. Biol. Rep 2005, 23, 251–262. [Google Scholar]
- Ga, JW; Liu, JZ; Li, B; Li, ZS. Isolation and purification of functional total RNA from blue-grained wheat endosperm tissues containing high levels of starches and flavonoids. Plant Mol. Biol. Rep 2001, 19, 185a–185i. [Google Scholar]
- Tabor, S; Richardson, CC. A bacteriophage T7 RNA polymerase/promoter system for controlled exclusive expression of specific genes. Proc. Natl. Acad. Sci. USA 1985, 82, 1074–1078. [Google Scholar]
- Thaller, MC; Schippa, S; Rossolini, GM. Conserved sequence motifs among bacterial, eukaryotic and archaeal phosphatases that define a new phosphohydrolase superfamily. Protein Sci 1998, 7, 1647–1652. [Google Scholar]
- Kosmas, SA; Argyrokastritis, A; Loukas, MG; Eliopoulos, E; Tsakas, S; Kaltsikes, PJ. Isolation and characterization of drought-related trehalose 6-phosphate-synthase gene from cultivated cotton (Gossypium hirsutum L.). Planta 2006, 223, 329–339. [Google Scholar]
- Muller, J; Wiemken, A; Aeschbacher, R. Trehalose metabolism in sugar sensing and plant development. Plant Sci 1999, 147, 37–47. [Google Scholar]
- Vogel, G; Fiehn, O; Jean-Richaro-dit-Bresse, L; Boller, T; Wiemken, A; Aeschbacher, RA; Wingler, A. Trehalose metabolism Arabidopsis: occurrence of trehalose and molecular cloning and characterization of Trehalose-6-phosphate synthase homologs. J. Exp. Bot 2001, 52, 1817–1826. [Google Scholar]
- Leyman, B; Van Dijck, P; Thevelein, JM. An unexpected plethora of trehalose biosynthesis genes in Arabidopsis thaliana. Trends Plant Sci 2001, 6, 510–513. [Google Scholar]
- Geelen, D; Royackers, K; Vanstraelen, M; De Bus, M; Inze, D; Van Dijck, P; Thevelein, JM; Leyman, B. Trehalose-6-phosphate synthase AtTPS1 high molecular weight complexes in yeast and Arabidopsis. Plant Sci 2007, 173, 426–437. [Google Scholar]
- Wang, B. Institute of Genetics and Developmental Biology: Beijing, China, Unpublished work; 2010.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer in pair | Sequence (5′→3′) | Product size (kb) | Orientation and position |
|---|---|---|---|
| 1.TPSR1 | GACTCATATGaACCCCCGGGCCTATCACTA | 1.36 | 5′→3′ (1~22 nt) |
| 3KpnI | CATGATGCTGTACAGCGCAAG | 3′→5′ (1339 ~1359 nt) | |
| 2.Tre1 | CTACGCGCGTCACTTTCTCTC | 1.2 | 5′→3′ (861~881 nt) |
| TPSa2 | CACTCCTTCGAATTCTTCTTG | 3′→5′ (2034~2054 nt) | |
| 3.TPSb1 | CAAGAAGAATTCGAAGGAGTG | 0.7 | 5′→3′ (2034~2054 nt) |
| TPSb2 | GACTAAGCTTbCTACTGGCTCGGCAACGAGGAC | 3′→5′ (2706~2727 nt) |
| Isolated TPS gene | From the seaweed species | GenBank accession number (NCBI) |
|---|---|---|
| PyTPS | Porphyra yezoensis | AY729671 |
| MaTPS | Monostroma angicava | DQ666324 |
| LjTPS | Laminaria japonica | DQ666325 |
| PhTPS | Porphyra haitanensis | DQ666326 |
| GlTPS | Gracilariopsis lemaneiformis | DQ666327 |
| CoTPS | Chondrus ocellatus | DQ666328 |
| UlpTPS | Ulva pertusa | DQ666329 |
| EpTPS | Enteromorpha prolifera | DQ666330 |
| UnpTPS | Undaria pinnatifida | GQ352535 |
| ShTPS | Sargassum henslowianum | GQ352536 |
| TPS gene | Numbers of nucleotide variations | Numbers of amino acid substitutions | Homology of nucleotide sequences (%) | Homology of deduced amino acid sequences (%) |
|---|---|---|---|---|
| PyTPS | - | - | 100 | 100 |
| MaTPS | 8 | 6 | 99.7 | 99.3 |
| EpTPS | 9 | 7 | 99.7 | 99.3 |
| ShTPS | 10 | 7 | 99.6 | 99.2 |
| CoTPS | 11 | 8 | 99.6 | 99.2 |
| GlTPS | 11 | 8 | 99.6 | 99.1 |
| UnpTPS | 14 | 9 | 99.5 | 99.1 |
| LjTPS | 28 | 6 | 99.0 | 99.0 |
| UlpTPS | 95 | 19 | 96.5 | 97.9 |
| PhTPS | 152 | 30 | 94.4 | 96.7 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, G.; Zhao, G.; Feng, Y.; Xuan, J.; Sun, J.; Guo, B.; Jiang, G.; Weng, M.; Yao, J.; Wang, B.; et al. Cloning and Comparative Studies of Seaweed Trehalose-6-Phosphate Synthase Genes. Mar. Drugs 2010, 8, 2065-2079. https://doi.org/10.3390/md8072065
Wang G, Zhao G, Feng Y, Xuan J, Sun J, Guo B, Jiang G, Weng M, Yao J, Wang B, et al. Cloning and Comparative Studies of Seaweed Trehalose-6-Phosphate Synthase Genes. Marine Drugs. 2010; 8(7):2065-2079. https://doi.org/10.3390/md8072065
Chicago/Turabian StyleWang, Guoliang, Ge Zhao, Yanbin Feng, Jinsong Xuan, Jianwei Sun, Baotai Guo, Guoyong Jiang, Manli Weng, Jianting Yao, Bin Wang, and et al. 2010. "Cloning and Comparative Studies of Seaweed Trehalose-6-Phosphate Synthase Genes" Marine Drugs 8, no. 7: 2065-2079. https://doi.org/10.3390/md8072065