Search for Hydrophilic Marine Fungal Metabolites: A Rational Approach for Their Production and Extraction in a Bioactivity Screening Context

Abstract
:1. Introduction
2. Results and Discussion
2.1. Choice of the extraction process: organic or aqueous process?
2.2. Choice of the fermentation method: SmF or ASF?
2.3. Choice of the raw material to extract: mycelium and/or medium?
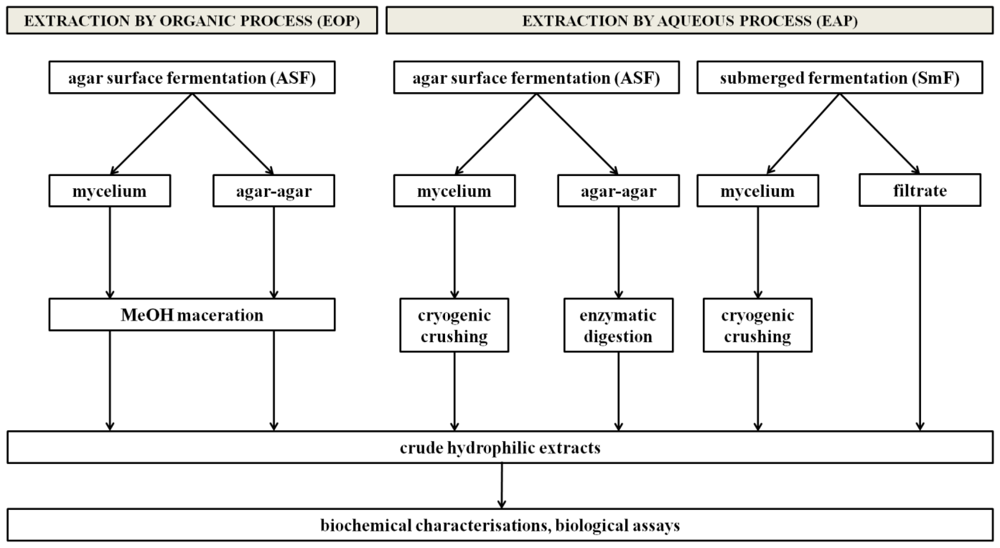
2.4. The proposed strategy
2.5. Marine fungi, a promising source of bioactive hydrophilic substances
3. Experimental Section
3.1. Fungal strains and culture medium
3.2. Agar surface fermentation: culture and extraction
3.2.1. Agar surface fermentation
3.2.2. Extraction of agar-agar
3.2.2.1 Extraction of agar-agar by aqueous process (EAP)
3.2.2.2. Extraction of agar-agar by organic process (EOP)
3.2.3. Extraction of mycelium
3.3. Submerged fermentation: culture and extraction
3.3.1. Submerged fermentation
3.3.2. Extraction from SmF
3.4. Biochemical and bioactivity evaluations
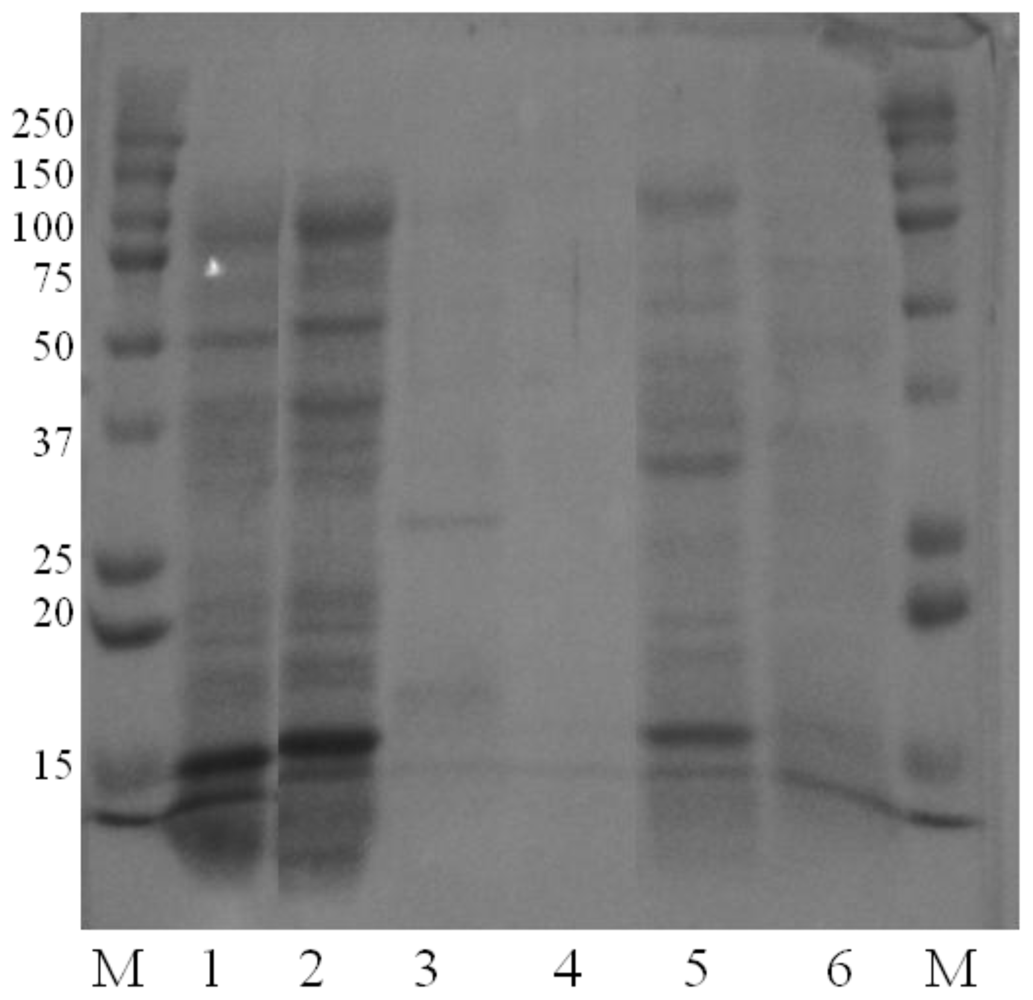
3.4.1. Characterization and measurement
3.4.2. Bioactivity assays
3.4.2.1. Neuroactivity assay on blowfly larvae
3.4.2.2. Cytotoxicity assay
4. Conclusion
Acknowledgements
- Sample Availability: Available from the authors.
References
- Bugni, TS; Ireland, CM. Marine-derived fungi: a chemically and biologically diverse group of microorganisms. Nat Prod Rep 2004, 21, 143–163. [Google Scholar]
- Kerzaon, I; Grovel, O; Robiou du Pont, T; Le Pape, P; Pouchus, YF. Effects of seawater on growth and gliotoxin excretion of marine strains of Aspergillus fumigatus Fres. Toxicon 2008, 51, 398–405. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, WP; Murray, HG; Munro, MH; Northcote, PT; Prinsep, MR. Marine natural products. Nat Prod Rep 2009, 26, 170–244. [Google Scholar]
- Yang, XB; Gao, XD; Han, F; Xu, BS; Song, YC; Tan, RX. Purification, characterization and enzymatic degradation of YCP, a polysaccharide from marine filamentous fungus Phoma herbarum YS4108. Biochimie 2005, 87, 747–754. [Google Scholar]
- Sun, HH; Mao, WJ; Chen, Y; Guo, SD; Li, HY; Qi, XH; Chen, YL; Xu, J. Isolation, chemical characteristics and antioxidant properties of the polysaccharides from marine fungus Penicilliums. F23–2. Carbohydr Polym 2009, 78, 117–124. [Google Scholar]
- Sun, C; Wang, JW; Lang, L; Gao, XD; Tan, RX. Free radical scavenging and antioxidant activities of EPS2, an exopolysaccharide produced by a marine filamentous fungus Keissleriella sp. YS4108. Life Sci 2004, 75, 1063–1073. [Google Scholar]
- Arias, HR. Marine toxins targeting ion channels. Mar Drugs 2006, 4, 37–69. [Google Scholar]
- Ng, TB. Peptides and proteins from fungi. Peptides 2004, 25, 1055–1073. [Google Scholar]
- Elias, BC; Said, S; de Albuquerque, S; Pupo, MT. The influence of culture conditions on the biosynthesis of secondary metabolites by Penicillium verrucosum Dierck. Microbiol Res 2006, 161, 273–280. [Google Scholar]
- Shimizu, Y; Li, B. Purification of Water-Soluble Natural Products. Natural Products Isolation Methods in Biotechnology 2005, 20, 415–438. [Google Scholar]
- Singhania, RR; Patel, AK; Soccol, CR; Pandey, A. Recent advances in solid state fermentation. Biochem Eng J 2009, 44, 13–18. [Google Scholar]
- Perdue, RE. KB cell culture. I. Role in discovery of antitumor agents from higher plants. J Nat Prod 1982. [Google Scholar]
- Iwashita, K. Recent studies of protein secretion by filamentous fungi. J Biosci Bioeng 2002, 94, 530–535. [Google Scholar]
- Wolski, E; Menusi, E; Remonatto, D; Vardanega, R; Arbter, F; Rigo, E; Ninow, J; Mazutti, MA; Di Luccio, M; Oliviera, D; Treichel, H. Partial characterization of lipases produced by a newly isolated Penicillium sp. in solid state and submerged fermentation: A comparative study. Food Sci Technol 2009, 42, 1557–1560. [Google Scholar]
- Saqib, AAN; Hassan, M; Khan, NF; Baig, S. Thermostability of crude endoglucanase from Aspergillus fumigatus grown under solid state fermentation (SSF) and submerged fermentation (SmF). Process Biochem 2010, 45, 641–646. [Google Scholar]
- Ruiz, N; Dubois, N; Wielgosz-Collin, G; Robiou du Pont, T; Bergé, JP; Pouchus, YF; Barnathan, G. Lipid content and fatty acid composition of a marine-derived Trichoderma longibrachiatum strain cultured by agar surface and submerged fermentations. Process Biochem 2007, 42, 676–680. [Google Scholar]
- Hölker, U; Lenz, J. Solid-state fermentation - are there any biotechnological advantages. Curr Opin Microbiol 2005, 8, 301–306. [Google Scholar]
- Caldwell, RB; Lattemann, CT. Simple and reliable method to precipitate proteins from bacterial culture supernatant. Appl Environ Microbiol 2004, 70, 610–612. [Google Scholar]
- Wilson, LM; Idnurm, A; Howlett, BJ. Characterization of a gene (sp1) encoding a secreted protein from Leptosphaeria maculans, the blackleg pathogen of Brassica napus. J Plant Pathol 2002, 3, 487–493. [Google Scholar]
- Fogliano, V; Marchese, A; Scalonis, A; Ritieni, A; Viscontis, A; Randazzo, G; Graniti, A. Characterization of a 60 kDa phytotoxic glycoprotein produced by Phoma tracheiphila and its relation to malseccin. Physiol Mol Plant Pathol 1998, 53, 149–161. [Google Scholar]
- Nachmias, A; Barash, I; Buchner, V; Solel, Z; Strobel, GA. A phytotoxic glycopeptide from lemon leaves infected with Phoma tracheiphila. Physiol Plant Pathol 1979, 14, 139–140. [Google Scholar]
- Narayanan, S; Surendranath, K; Bora, N; Surolia, A; Karande, AA. Ribosome inactivating proteins and apoptosis. FEBS Lett 2005, 579, 1324–1331. [Google Scholar]
- Mody, R; Joshi, S; Chaney, W. Use of lectins as diagnostic and therapeutic tools for cancer. J Pharmacol Toxicol Methods 1995, 33, 1–10. [Google Scholar]
- Martinez-Ruiz, A; Kao, R; Davies, J; Martinez del Pozo, A. Ribotoxins are a more widespread group of proteins within the filamentous fungi than previously believed. Toxicon 1999, 37, 1549–1563. [Google Scholar]
- Singh, RS; Sharma, S; Kaur, G; Bhari, R. Screening of Penicillium species for occurrence of lectins and their characterization. J Basic Microbiol 2009, 49, 471–476. [Google Scholar]
- Bartnicki-Garcia, S. Cell wall chemistry, morphogenesis and taxonomy of fungi. Annu Rev Microbiol 1968, 22, 87–108. [Google Scholar]
- Bradford, MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal Biochem 1976, 72, 248–250. [Google Scholar]
- Laemmli, UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]
- Dubois, M; Gilles, KA; Hamilton, JK; Rebers, PA; Smith, F. Colorimetric method for determination of sugars and related substances. Anal Chem 1956, 28, 350–356. [Google Scholar]
- Zlotkin, E; Fraenkel, G; Miranda, F; Lissitzky, S. The effect of scorpion venom on blowfly larvae; a new method for evaluation of scorpion venom potency. Toxicon 1971, 9, 1–8. [Google Scholar]
- Ruiz, N; Petit, K; Vansteelandt, M; Kerzaon, I; Baudet, J; Amzil, Z; Biard, JF; Grovel, O; Pouchus, YF. Enhancement of domoic acid neurotoxicity on Diptera larvae bioassay by marine fungal metabolites. Toxicon 2010, 55, 805–810. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 1983, 16, 55–63. [Google Scholar]
Abbreviations
| ASF | agar surface fermentation |
| EAP | extraction by aqueous process |
| EOP | extraction by organic process |
| MED | minimal effective dose |
| SmF | submerged fermentation |
| SSF | solid state fermentation |




{kind=link}
{kind=link}
{kind=link}
| EOP | EAP | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ASF | ASF | SmF | ||||||||||
| mycelium | agar-agar | mycelium | agar-agar | mycelium | filtratea | |||||||
| protein yield | sugar yield | protein yield | sugar yield | protein yield | sugar yield | protein yield | sugar yield | protein yield | sugar yield | protein yield | sugar yield | |
| (mg/g dw) | (mg/g dw) | (mg/mL) | (mg/mL) | (mg/g dw) | (mg/g dw) | (mg/mL) | (mg/mL) | (mg/g dw) | (mg/g dw) | (mg/mL) | (mg/mL) | |
| Penicillium citreonigrum | 0.78 | 14.25 | 0.07 | 0.25 | 10.67 | 34.19 | 0.55 | 2.13 | 9.52 | 35.41 | 0.36 | 2.27 |
| Phoma sp. | 1.11 | 11.11 | 0.09 | 0.65 | 14.45 | 33.32 | 0.60 | 3.56 | 13.87 | 32.15 | 0.37 | 4.13 |
| Phoma exigua var exigua | 2.48 | 20.23 | 0.05 | 0.31 | 10.50 | 44.25 | 0.22 | 0.85 | 11.19 | 42.63 | 0.19 | 0.77 |
| Scopulariopsis sp. | 4.01 | 36.43 | 0.12 | 0.21 | 16.04 | 51.35 | 0.42 | 1.33 | 16.54 | 50.86 | 0.28 | 1.19 |
| Chrysosporium queenslandicum | 5.70 | 37.66 | 0.08 | 0.45 | 21.03 | 55.28 | 0.15 | 2.43 | 23.17 | 53.77 | 0.13 | 2.25 |
| Pycnidiophora dispersa | 2.43 | 20.30 | 0.09 | 0.42 | 12.44 | 46.66 | 0.20 | 2.97 | 11.35 | 45.34 | 0.18 | 2.08 |
| average | 2.75 | 23.33 | 0.08 | 0.38 | 14.19 | 44.18 | 0.36 | 2.21 | 14.27 | 43.36 | 0.25 | 2.12 |
| EOP | EAP | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ASF | ASF | SmF | ||||||||||
| mycelium | agar-agar | mycelium | agar-agar | mycelium | filtrate | |||||||
| neuroactivity | cytotoxicity | neuroactivity | cytotoxicity | neuroactivity | cytotoxicity | neuroactivity | cytotoxicity | neuroactivity | cytotoxicity | neuroactivity | cytotoxicity | |
| MED | IC50 | MED | IC50 | MED | IC50 | MED | IC50 | MED | IC50 | MED | IC50 | |
| mg/mL | μg/mL | mg/mL | μg/mL | mg/mL | μg/mL | mg/mL | μg/mL | mg/mL | μg/mL | mg/mL | μg/mL | |
| Penicillium citreonigrum | 100 | - | 100 | - | 12.5 | - | 100 | - | 50 | - | - | - |
| Phoma sp. | 50 | - | - | - | 25 | 68 | 100 | 130 | 50 | 62 | 100 | 85 |
| Phoma exigua var exigua | 50 | - | 100 | - | - | - | - | 57 | - | - | - | 97 |
| Scopulariopsis sp. | 100 | - | 100 | - | 100 | - | - | 44 | - | - | - | 48 |
| Chrysosporium queenslandicum | 100 | - | 100 | - | 100 | - | - | - | 100 | - | - | - |
| Pycnidiophora dispersa | 100 | - | - | - | - | 70 | - | - | - | 94 | - | - |
| inert medium | - | - | - | - | - | - | ||||||
Biomassa (g dw/L) | ||
|---|---|---|
| ASF | SmF | |
| Penicillium citreonigrum | 50.0 ± 2.1 | 35.6 ± 1.8 |
| Phoma sp. | 33.3 ± 1.7 | 7.1 ± 0.2 |
| Phoma exigua var exigua | 53.4 ± 2.4 | 6.2 ± 0.3 |
| Scopulariopsis sp. | 57.9 ± 2.6 | 2.0 ± 0.1 |
| Chrysosporium queenslandicum | 5.8 ± 0.3 | 0.8 ± 0.3 |
| Pycnidiophora dispersa | 54.2 ± 1.9 | 31.7 ± 1.5 |
| average | 42.4 | 13.9 |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ker, C.L.; Petit, K.-E.; Biard, J.-F.; Fleurence, J. Search for Hydrophilic Marine Fungal Metabolites: A Rational Approach for Their Production and Extraction in a Bioactivity Screening Context. Mar. Drugs 2011, 9, 82-97. https://doi.org/10.3390/md9010082
Ker CL, Petit K-E, Biard J-F, Fleurence J. Search for Hydrophilic Marine Fungal Metabolites: A Rational Approach for Their Production and Extraction in a Bioactivity Screening Context. Marine Drugs. 2011; 9(1):82-97. https://doi.org/10.3390/md9010082
Chicago/Turabian StyleKer, Carine Le, Karina-Ethel Petit, Jean-François Biard, and Joël Fleurence. 2011. "Search for Hydrophilic Marine Fungal Metabolites: A Rational Approach for Their Production and Extraction in a Bioactivity Screening Context" Marine Drugs 9, no. 1: 82-97. https://doi.org/10.3390/md9010082