Tasco®, a Product of Ascophyllum nodosum, Imparts Thermal Stress Tolerance in Caenorhabditis elegans

Abstract
:1. Introduction
2. Results
2.1. Tasco® Water Extract (TWE)-Treated Nematodes Exhibited Enhanced Thermal Tolerance
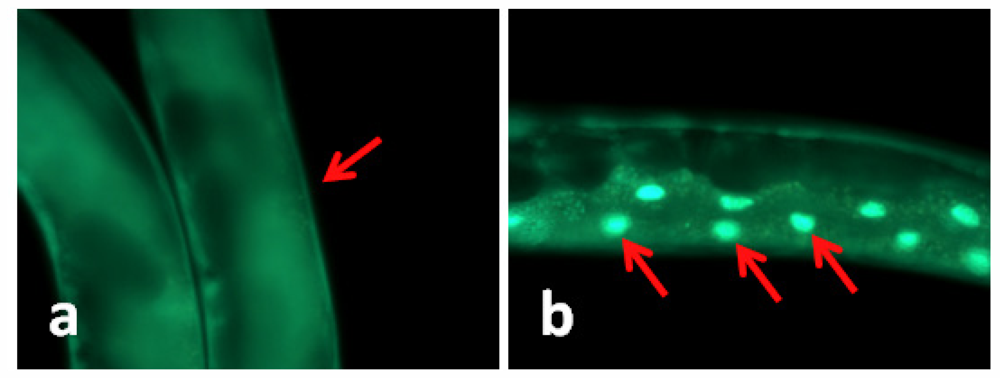
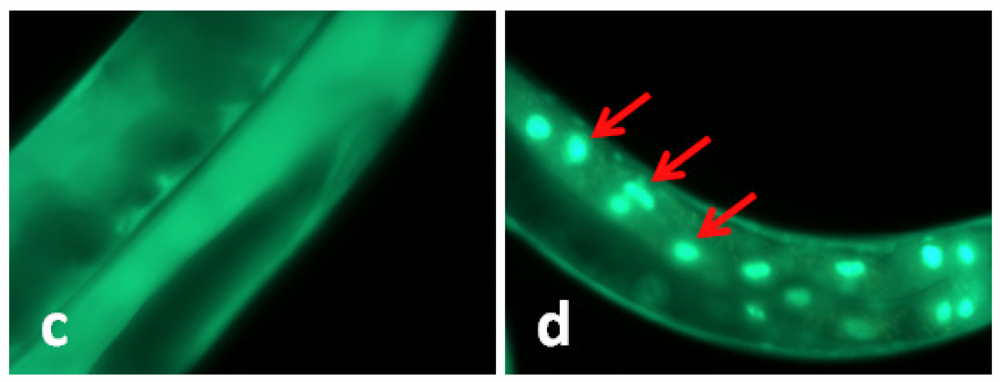
2.2. TWE-Treatment Does Not Affect Cellular Localization of daf-16 in C. elegans
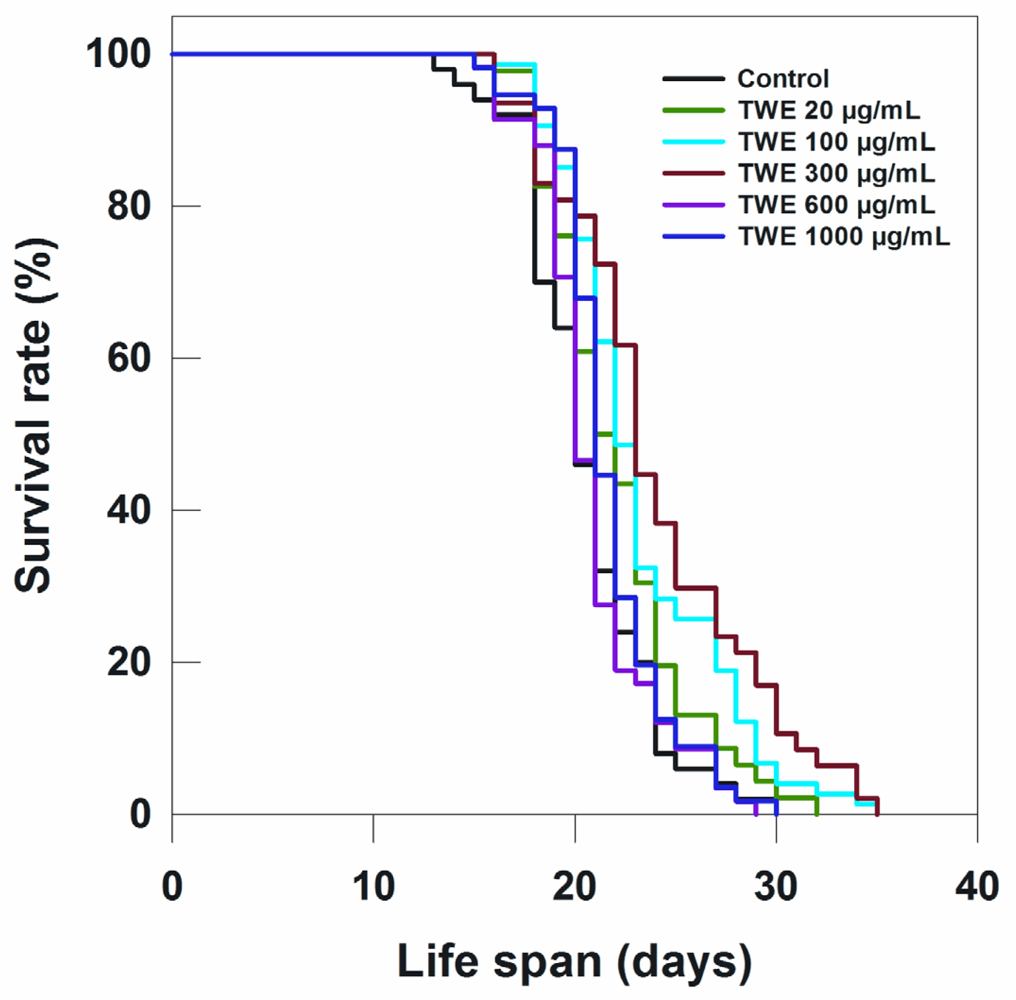
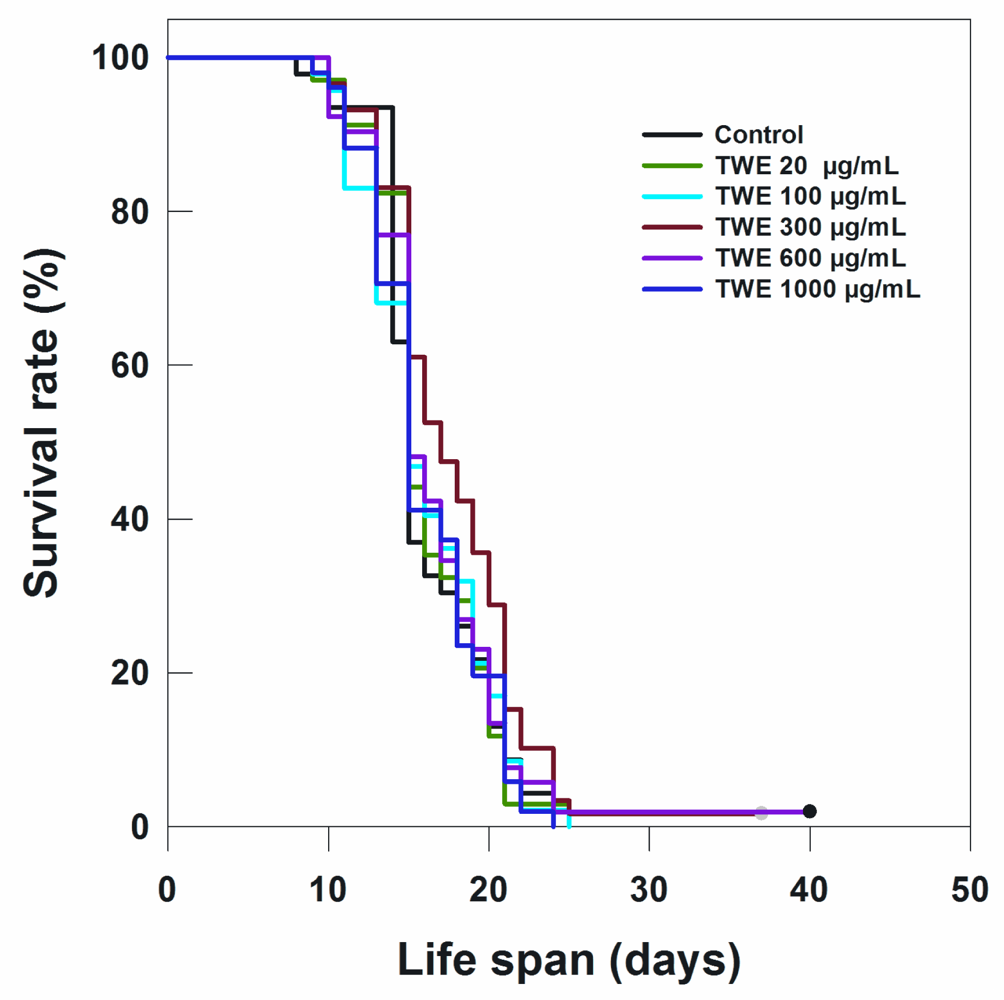
2.3. TWE Treatment Prolongs Life Span in Wild Type C. elegans
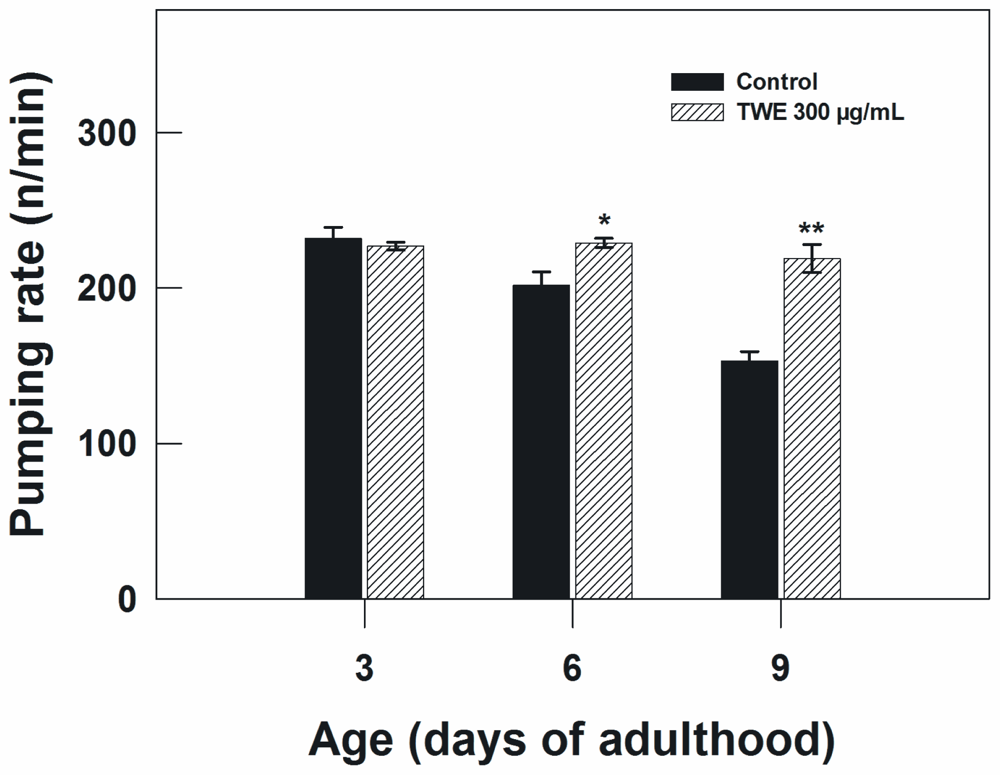
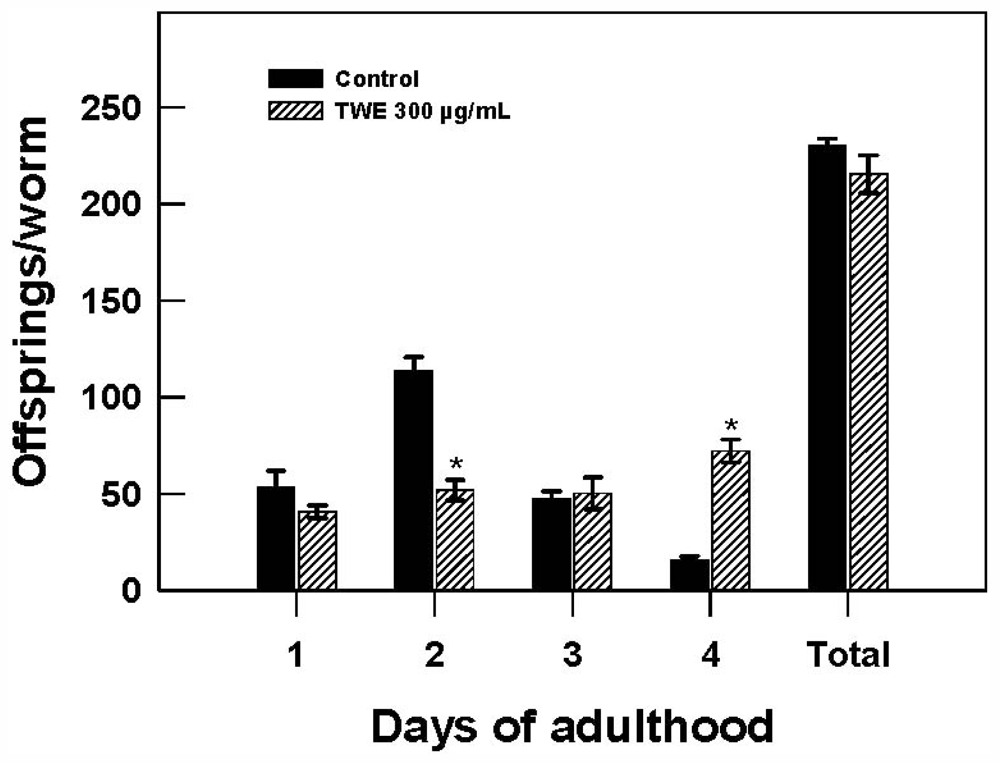
2.4. Effect of TWE Treatment on Body Size, Pharyngeal Pumping Rate, and Fecundity in C. elegans
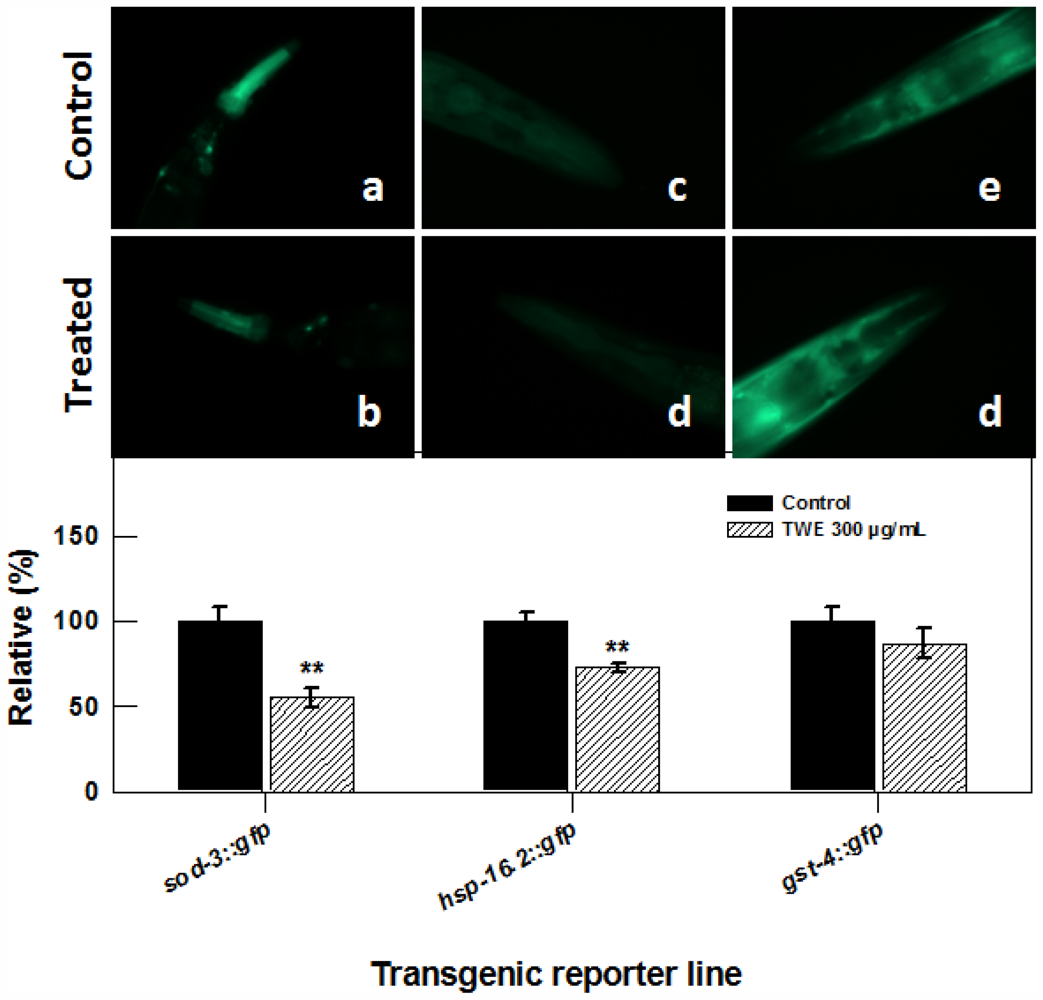
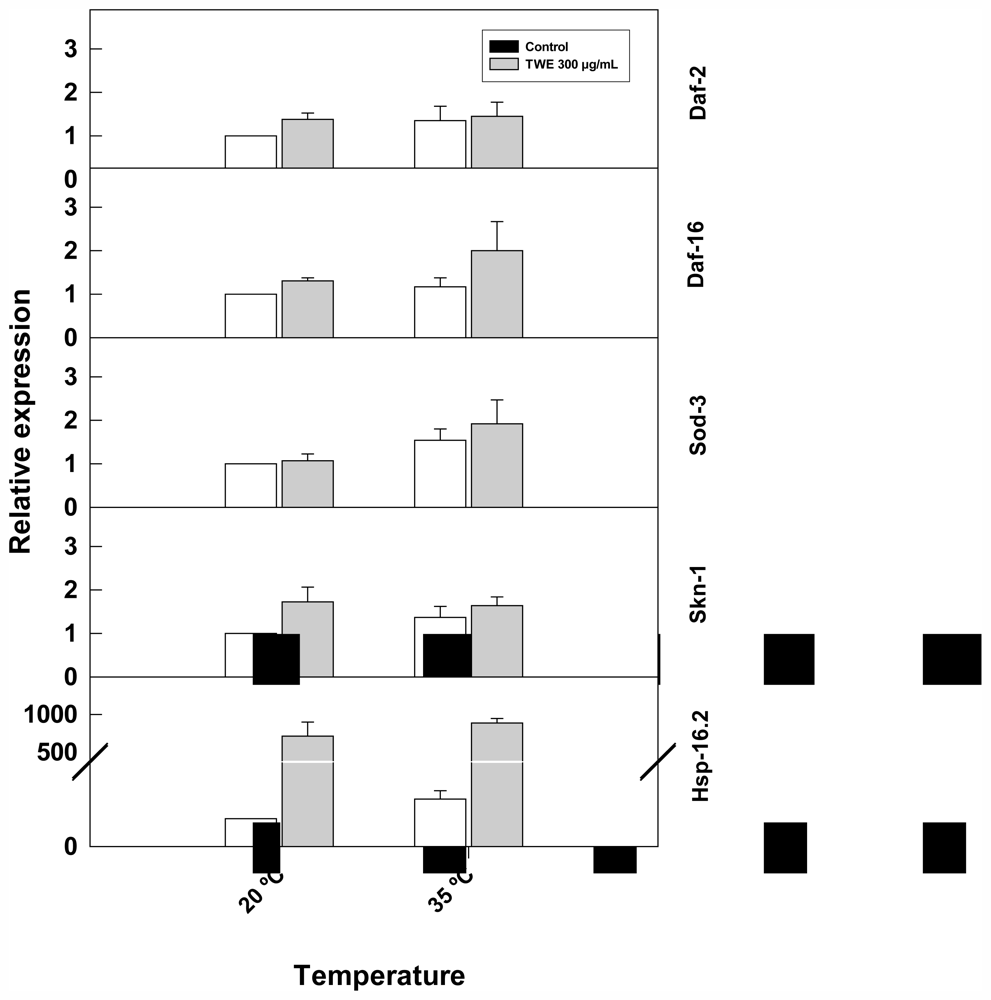
2.5. TWE Affects the Expression of a Number of Stress Related Genes in C. elegans
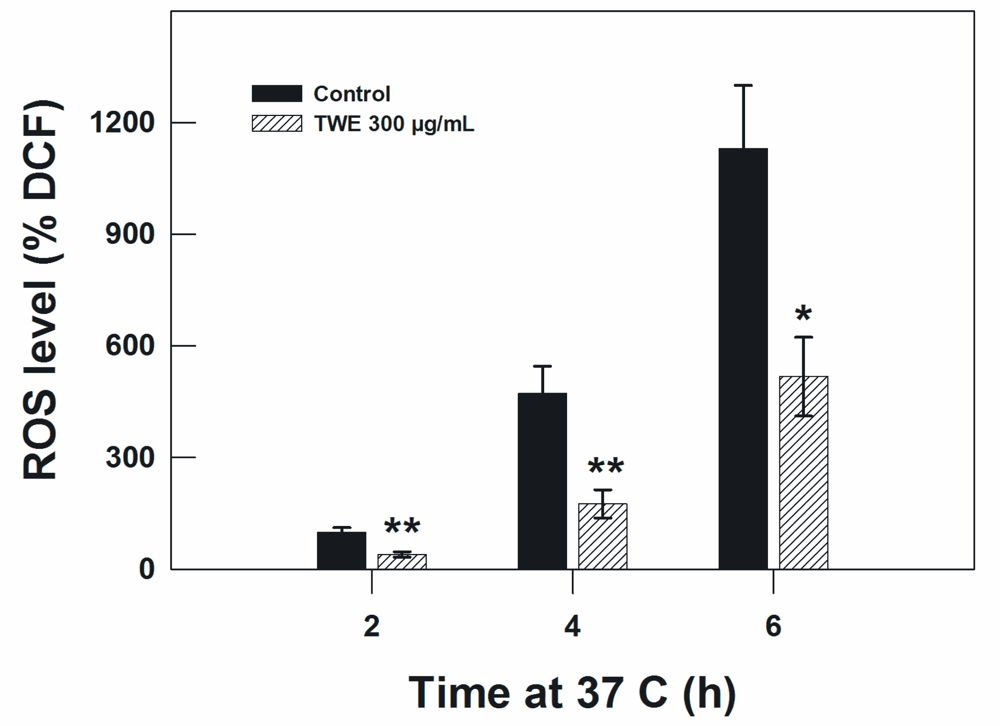
2.6. Measurement of Reactive Oxygen Species (ROS)
2.7. Effect of TWE Treatment on the Expression of Stress Related Genes in C. elegans
2.8. Effect of TWE on C. elegans Proteome under Thermal Stress
2.9. Functional Prediction of Hypothetical Proteins
2.9.1. Signal Transduction
2.9.2. Temperature Specific Marker Proteins
3. Discussion
4. Experimental Section
4.1. Chemicals and C. elegans Strains
4.2. Preparation of Tasco® Water Extract (TWE)
4.3. Thermal-Tolerance Assay
4.4. Stress Response Assay with Mutants and Transgenic Strains of C. elegans
4.5. Life Span Assay
4.6. Body Size Analysis
4.7. Brood Size Assay
4.8. Feeding Rate Assay
4.9. Measurement of Reactive Oxygen Species (ROS)
4.10. Expression Analysis of Stress Induced Genes in C. elegans
4.11. Proteome Analysis of TWE Treated C. elegans
4.11.1. Preparation of Synchronized Nematode Populations
4.11.2. Treatment Conditions
4.11.3. Protein Extraction
4.11.4. 2D SDS-PAGE
4.12. Protein Identification
4.12.1. Protein Digestion
4.12.2. Protein Identification and Sequencing by 2D Nano LC-MS/MS
4.12.3. Database Searching with MS/MS Spectra
4.13. Statistical Analysis
5. Conclusions
Acknowledgements
References
- Kim, S.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: a review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive proteins, peptides, and amino acids from Macroalgae. J. Phycol 2011, 47, 218–232. [Google Scholar]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development (review). J. Plant Growth Regul 2009, 28, 386–399. [Google Scholar]
- Ugarte, R.; Craigie, J.S.; Critchley, A.T. Fucoid Flora of the Rocky Intertidal of the Canadian Maritimes: Implications for the Future with Rapid Climate Change. In Seaweeds and Their Role in Globally Changing Environments; Israel, A., Einav, R., Seckbach, J., Eds.; Springer: Berlin, Germany, 2010. [Google Scholar]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol 2011, 23, 371–393. [Google Scholar]
- Allen, V.G.; Pond, K.R.; Saker, K.E.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Schmidt, R.E.; Fike, J.H.; Zhang, X.; et al. Tasco: Influence of a brown seaweed on antioxidants in forages and livestock—A review. J. Anim. Sci 2001, 79, E21–E31. [Google Scholar]
- Bach, A.; Iglesias, C.; Devant, M. Daily rumen pH pattern of loose-housed dairy cattle as affected by feeding pattern and live yeast supplementation. Anim. Feed Sci. Technol 2007, 136, 146–153. [Google Scholar]
- Hoshino, T.; Hayashi, T.; Hayashi, K.; Hamada, J.; Lee, J.B. An antiviral active sulfated polysaccharide from Sargassum horneri (Tuner) C. Agardh. Biol. Pharm. Bull 1998, 21, 730–734. [Google Scholar]
- Allen, V.G.; Pond, K.R.; Saker, K.E.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Brown, C.P.; Miller, M.F.; Montgomery, J.L.; et al. Tasco-Forage: III. Influence of a seaweed extract on performance, monocyte immune cell response, and carcass characteristics in feedlot-finished steers. J. Anim. Sci 2001, 79, 1032–1040. [Google Scholar]
- Gruber, J.; Ng, L.F.; Poovathingal, S.K.; Halliwell, B. Deceptively simple but simply deceptive—Caenorhabditis elegans lifespan studies: Considerations for aging and antioxidant effects. FEBS Lett 2009, 583, 3377–3387. [Google Scholar]
- Kampkotter, A.; Gombitang, N.C.; Zurawski, R.F.; Timpel, C.; Chovolou, Y. Effects of the flavonoids kaempferol and fisetin on thermotolerance, oxidative stress and FoxO transcription factor DAF-16 in the model organism Caenorhabditis elegans. Arch. Toxicol 2007, 81, 849–858. [Google Scholar]
- Henderson, S.T.; Johnson, T.E. daf-16 integrates developmental and environmental inputs to mediate aging in the nematode Caenorhabditis elegans. Curr. Biol 2001, 11, 1975–1980. [Google Scholar]
- NCBI protein database. Available online: http://www.ncbi.nlm.nih.gov/blast/html (accessed on 14 November 2010).
- Heschl, M.F.; Baillie, D.L. The HSP70 multigene family of Caenorhabditis elegans. Comp. Biochem. Physiol 1990, 96, 633–637. [Google Scholar]
- Saker, K.E.; Allen, V.G.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Wester, D.B. Tasco-Forage: II. Monocyte immune cell response and performance of beef steers grazing tall fescue treated with a seaweed extract. J. Anim. Sci 2001, 79, 1022–1031. [Google Scholar]
- Fike, J.H.; Allen, V.G.; Schmidt, R.E.; Zhang, X.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Coelho, R.W.; Wester, D.B. Tasco-Forage: I. Influence of a seaweed extract on antioxidant activity in tall fescue and in ruminants. J. Anim. Sci 2001, 79, 1011–1021. [Google Scholar]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Dis 2006, 5, 387–399. [Google Scholar]
- Yu, Y.B.; Dosanjh, L.; Lao, L.X.; Tan, M.; Shim, B.S.; Luo, Y. Cinnamomum cassia bark in two herbal formulas increases life span in Caenorhabditis elegans via insulin signaling and stress response pathways. PLoS One 2010, 5. [Google Scholar] [CrossRef] [Green Version]
- Prahlad, V.; Cornelius, T.; Morimoto, R.I. Regulation of the cellular heat shock response in Caenorhabditis elegans by thermosensory neurons. Science 2008, 320, 811–814. [Google Scholar]
- Herndon, L.A.; Schmeissner, P.J.; Dudaronek, J.M.; Brown, P.A.; Listner, K.M.; Sakano, Y.; Paupard, M.C.; Hall, D.H.; Driscoll, M. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature 2002, 419, 808–814. [Google Scholar]
- Link, C.D. C. elegans models of age-associated neurodegenerative diseases: Lessons from transgenic worm models of Alzheimer’s disease. Exp. Gerontol 2006, 41, 1007–1013. [Google Scholar]
- Moseley, P.L. Heat shock proteins and heat adaptation of the whole organism. J. Appl. Physiol 1997, 83, 1413–1417. [Google Scholar]
- Samali, A.; Holmberg, C.I.; Sistonen, L.; Orrenius, S. Thermotolerance and cell death are distinct cellular responses to stress: Dependence on heat shock proteins. FEBS Lett 1999, 461, 306–310. [Google Scholar]
- Horikawa, M.; Sakamoto, K. Fatty-acid metabolism is involved in stress-resistance mechanisms of Caenorhabditis elegans. Biochem. Biophys. Res. Commun 2009, 390, 1402–1407. [Google Scholar]
- Baumeister, R.; Schaffitzel, E.; Hertweck, M. Endocrine signaling in Caenorhabditis elegans controls stress response and longevity. J. Endocrinol 2006, 190, 191–202. [Google Scholar]
- Lamitina, S.T.; Strange, K. Transcriptional targets of DAF-16 insulin signaling pathway protect C. elegans from extreme hypertonic stress. Am. J. Physiol. Cell Physiol 2005, 288, 467–474. [Google Scholar]
- Lin, K.; Dorman, J.B.; Rodan, A.; Kenyon, C. daf-16: An HNF-3/forkhead family member that can function to double the life-span of Caenorhabditis elegans. Science 1997, 278, 1319–1322. [Google Scholar]
- Hsu, A.L.; Murphy, C.T.; Kenyon, C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 2003, 300, 1142–1145. [Google Scholar]
- Libina, N.; Berman, J.R.; Kenyon, C. Tissue-specific activities of C. elegans DAF-16 in the regulation of life span. Cell 2003, 115, 489–502. [Google Scholar]
- Morley, J.F.; Morimoto, R.I. Regulation of longevity in Caenorhabditis elegans by heat shock factor and molecular chaperones. Mol. Biol. Cell 2004, 15, 657–664. [Google Scholar]
- Westerheide, S.D.; Morimoto, R.I. Heat shock response modulators as therapeutic tools for diseases of protein conformation. J. Biol. Chem 2005, 280, 33097–33100. [Google Scholar]
- de Thonel, A.; Mezger, V.; Garrido, C. Implication of heat shock factors in tumorigenesis: therapeutical potential. Cancers 2011, 3, 1158–1181. [Google Scholar]
- Morimoto, R.I. Regulation of the heat shock transcriptional response: Cross talk between a family of heat shock factors, molecular chaperones, and negative regulators. Genes Dev 1998, 12, 3788–3796. [Google Scholar]
- Kampkotter, A.; Pielarski, T.; Rohrig, R.; Timpel, C.; Chovolou, Y.; Watjen, W.; Kahl, R. The Ginkgo biloba extract EGb761 reduces stress sensitivity, ROS accumulation and expression of catalase and glutathione S-transferase 4 in Caenorhabditis elegans. Pharmacol. Res 2007, 55, 139–147. [Google Scholar]
- Strayer, A.; Wu, Z.; Christen, Y.; Link, C.D.; Luo, Y. Expression of the small heat-shock protein Hsp16-2 in Caenorhabditis elegans is suppressed by Ginkgo biloba extract EGb 761. FASEB J 2003, 17, 2305–2307. [Google Scholar]
- Rea, S.L.; Wu, D.; Cypser, J.R.; Vaupel, J.W.; Johnson, T.E. A stress-sensitive reporter predicts longevity in isogenic populations of Caenorhabditis elegans. Nat. Genet 2005, 37, 894–898. [Google Scholar]
- Zhang, L.; Jie, G.; Zhang, J.; Zhao, B. Significant longevity-extending effects of EGCG on Caenorhabditis elegans under stress. Free Rad. Biol. Med 2009, 46, 414–421. [Google Scholar]
- Fernandez-Ayala, D.J.; Chen, S.; Kemppainen, E.; O’Dell, K.M.; Jacobs, H.T. Gene expression in a Drosophila model of mitochondrial disease. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Anestia, V.; Scorrano, L. The relationship between mitochondrial shape and function and the cytoskeleton. Biochim. Biophys. Acta (BBA) Bioenerg 2006, 1757, 692–699. [Google Scholar]
- Kim, D.H.; Ausubel, F.M. Evolutionary perspectives on innate immunity from the study of Caenorhabditis elegans. Curr. Opin. Immunol 2005, 17, 4–10. [Google Scholar]
- Li, B.; Lu, F.; Wei, X.J.; Zhao, R.X. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar]
- Stiernagle, T. Maintenance ofC. elegans. WormBook. 2006. Available online: http://wormbook.org/chapters/www_strainmaintain/strainmaintain.html (accessed on 1 November 2010). [CrossRef]
- Java image processing program ImageJ; US National Institute of Health: Bethesda, MD, USA. Available online: http://rsb.info.nih.gov/ij/index.html (accessed on 30 August 2011).
- Hosono, R. Sterilization and growth inhibition of Caenorhabditis elegans by 5-fluorodeoxyuridine (FdUrd). Exp. Geron 1978, 13, 369–374. [Google Scholar]
- Raizen, D.M.; Lee, R.Y.N.; Avery, L. Interacting genes required for pharyngeal excitation by motor neuron MC in Caenorhabditis elegans. Genetics 1995, 141, 1365–1382. [Google Scholar]
- Kim, S.T.; Kim, S.G.; Hwang, D.H.; Kang, S.Y.; Kim, H.J.; Lee, B.H.; Lee, J.J.; Kang, K.Y. Proteomic analysis of pathogen-responsive proteins from rice leaves induced by rice blast fungus, Magnaporthe grisea. Proteomics 2004, 4, 3569–3578. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Salekdeh, G.H.; Siopongco, J.; Wade, L.J.; Ghareyazie, B.; Bennett, J. A proteomic approach to analyzing drought and salt-responsive in rice. Field Crop. Res 2002, 76, 1999–2199. [Google Scholar]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal. Chem 1996, 68, 850–858. [Google Scholar]
- Protein or DNA sequence data base. Available online: http://www.matrixscience.com/search_form_select.html (accessed on 24 October 2010).
- MASCOT Help. Available online: http://www.matrixscience.com/help/scoring_help.html#PBM (accessed on 24 December 2010).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (20 °C) | Total (N) | Censored (N) | Adult Life Span, Days (Mean ± SE) | P vs. Control (log-Rank) |
|---|---|---|---|---|
| Control | 125 | 0 | 20.56 ± 0.47 | |
| TWE 20 μg/mL | 115 | 0 | 22.09 ± 0.52 | 0.055 |
| TWE 100 μg/mL | 185 | 0 | 23.24 ± 0.47 | 0.0003 |
| TWE 300 μg/mL | 118 | 0 | 24.02 ± 0.72 | 0.00004 |
| TWE 600 μg/mL | 145 | 0 | 20.85 ± 0.39 | 0.899 |
| TWE 1000 μg/mL | 140 | 0 | 21.66 ± 0.37 | 0.175 |
| ID Number a | Name of Protein | Increase Fold in Treated Larva b | Accession Number | Molecular Mass (Da)/pI | Coverage (%) | Matched Peaks | Putative Functions |
|---|---|---|---|---|---|---|---|
| Up regulated proteins following TWE treatment | |||||||
| 1 | O62040_CAEEL Putative uncharacterized protein OS = Caenorhabditis elegans | D | O62040 | 37,708 | 2.37% | 1 | Strictosidine synthase (a key enzyme in alkaloid biosynthesis) activity |
| 2 | Q9N5D3_CAEEL Drosophila sos homolog protein 1 OS = Caenorhabditis elegans GN = SOS-1 | D | Q9N5D3 | 169,902 | 0.47% | 1 | DNA binding, Rho guanyl-nucleotide exchange factor activity, calmodulin binding |
| 3 | AL7A1_CAEEL Putative aldehyde dehydrogenase family 7 member A1 homolog OS = Caenorhabditis elegans | D | P46562 | 56,994.8 | 4.52% | 3 | Detoxification of aldehydes generated by alcohol metabolism and lipid peroxidation, Protects cells from oxidative stress. |
| 4 | HSP17_CAEEL Heat shock protein HSP-16.48/HSP-16.49 OS = Caenorhabditis elegans GN | D | P02513 | 16,282 | 4.9% | 1 | Determination of adult lifespan, endoplasmic reticulum unfolded protein response, response to heat |
| 5 | RSSA_CAEEL 40S ribosomal protein SA OS = Caenorhabditis elegansCaenorhabditis elegans GN = RPS-0 PE = 1 SV = 3 | D | P46769 | 30,685.1 | 6.16% | 2 | Assembly and/or stability of the 40S ribosomal subunit. and processing of the 20S rRNA-precursor to mature 18S rRNA in a late step of the maturation of 40S ribosomal subunits |
| 6 | Q9XW04_CAEEL Protein Y18D10A.21, partially confirmed by transcript evidence OS = C | D | Q9XW04 | 32,897.3 | 2.8% | 1 | Catalytic activity, cation binding, carbohydrate metabolic process |
| 7 | Q94255_CAEEL Saposin-like protein family protein 20 | D | Q94255 | 24,484.9 | 2.76% | 1 | Lipid metabolism |
| 8 | Q17849_CAEEL Heat shock protein protein 25, isoform a OS=Caenorhabditis elegans | D | Q17849 | 25,238.6 | 4.57% | 1 | Response to heat, protein binding, Stress response |
| 9 | D0LGS3_HALO1 Carbamoyl-phosphate synthase L chain ATP-binding protein OS = Haliang | 1.89 | D0LGS3 | 211,022 | 0.47% | 1 | ATP binding, biotin binding, ligase activity |
| 10 | C3JZ86_PSEFS Superoxide dismutase OS = Pseudomonas fluorescens (strain SBW25) GN = P | D | A0KJL0 | 22,743.6 | 4.57% | 1 | superoxide dismutase activity, oxidation reduction, superoxide metabolic process |
| 11 | Q18100_CAEEL Sensory axon guidance protein 7 | D | Q18100 | 127,605 | 0.61% | 1 | Involved in specific pathways by attractive and repulsive cues in the extracellular environment |
| 12 | Q6EUT7_CAEEL Protein ZK1151.1g, partially confirmed by transcript evidence OS = Caenorhabditis elegans | 1.88 | Q6EUT7 | 561,703 | 0.2% | 1 | Cell cycle arrest, Actin binding and Calcium ion binding |
| 13 | 14331_CAEEL 14-3-3-like protein 1 OS = Caenorhabditis elegans GN = PAR-5 PE = 1 SV = 2|6 | 1.52 | P41932 | 28,173 | 10.1% | 3 | Adapter protein involved in regulation of general and specialized signaling pathway |
| 14 | C5IWV5_PIG Trypsinogen OS = Sus scrofa PE = 2 SV = 1 | 1.57 | C5IWV5 | 24,391.3 | 7.79% | 2 | Proteolysis, serine-type endopeptidase activity |
| 15 | MLR1_CAEEL Myosin regulatory light chain 1 OS = Caenorhabditis elegans GN = MLC-1 PE | 4.9 | P19625 | 8586.4 | 31.2% | 15 | Calcium ion binding, motor activity |
| 16 | MLR1_CAEEL Myosin regulatory light chain 1 OS = Caenorhabditis elegans GN = MLC-1 PE | D | P19625 | 8586.4 | 31.2% | 6 | Increases myosin filament stability |
| 17 | C5IWV5_PIG Trypsinogen OS = Sus scrofa PE = 2 SV = 1 | D | C5IWV5 | 24,391.3 | 7.79% | 2 | Proteolysis, serine-type endopeptidase activity |
| 18 | HSP11_CAEEL Heat shock protein HSP-16.1/HSP-16.11 OS = Caenorhabditis elegans GN = h | D | P34696 | 16,235.5 | 18.6% | 1 | Defense response, determination of adult lifespan, positive regulation of growth rate, response to heat |
| 19 | COX5A_CAEEL Cytochrome c oxidase subunit 5A, mitochondrial OS = Caenorhabditis elegans | 4.31 | P55954 | 20,093.3 | 5.17% | 1 | Cytochrome-c oxidase activity, metal ion binding |
| 20 | O45509_CAEEL Protein F41D3.10, partially confirmed by transcript evidence OS = Caenorhabditis elegans | D | O45509 | 76,674 | 1.19% | 1 | Sequence-specific DNA binding, steroid hormone receptor activity, zinc ion binding |
| 21 | Q9UAX1_CAEEL Putative uncharacterized protein T12B3.4 OS = Caenorhabditis elegans | D | Q9UAX1 | 33,698.7 | 2.74% | 2 | Protein binding |
| 22 | HSP17_CAEEL Heat shock protein HSP-16.48/HSP-16.49 OS = Caenorhabditis elegans GN | D | P02513 | 16,282 | 11.2% | 5 | Determination of adult lifespan, endoplasmic reticulum unfolded protein response, response to heat |
| 23 | HSP11_CAEEL Heat shock protein HSP-16.1/HSP-16.11 OS = Caenorhabditis elegans GN = h | 1.2 | P34696 | 16,235.5 | 33.8% | 6 | Defense response, determination of adult lifespan, response to heat |
| 24 | O18180_CAEEL Protein W09D10.3, confirmed by transcript evidence OS = Caenorhabditis elegans | 2.36 | O18180 | 18,421.4 | 16.8% | 5 | Determination of adult lifespan, positive regulation of growth rate, structural constituent of ribosome |
| 25 | O44751_CAEEL Putative uncharacterized protein OS = Caenorhabditis elegans | 2.02 | O44751 | 55,504.4 | 1.23% | 1 | ATP binding, protein kinase activity |
| 26 | Q9XUT0_CAEEL Protein K08E3.4, confirmed by transcript evidence OS = Caenorhabditis elegans | 11.14 | Q9XUT0 | 71,922.8 | 3.11% | 2 | Actin binding |
| 27 | O45177_CAEEL Putative uncharacterized protein OS = Caenorhabditis elegans GN = K07H8 | 10.63 | O45177 | 40,387 | 2.31% | 1 | RNA binding |
| 28 | C5IWV5_PIG Trypsinogen OS = Sus scrofa PE = 2 SV = 1 | 1.0 | C3JZ86 | 21,959.7 | 4.55% | 1 | Proteolysis, serine-type endopeptidase activity |
| 29 | ATPB_CAEEL ATP synthase subunit beta, mitochondrial OS = Caenorhabditis elegans GN | 1.59 | P46561 | 57,509.3 | 1.86% | 1 | ATP binding, hydrogen ion transporting ATP synthase activity |
| 30 | GPX2_CAEEL Probable glutathione peroxidase R05H10.5 OS = Caenorhabditis elegans GN | 3.91 | O62327 | 18,134.3 | 18.4% | 3 | GSH-Px -an antioxidant, Important role in the metabolism of certain hydroperoxides |
| 31 | Q21057_CAEEL Galectin OS = Caenorhabditis elegans PE = 2 SV = 1|6239 | 3.79 | Q21057 | 15,911.4 | 26% | 4 | Sugar binding |
| 32 | Q20804_CAEEL CNB-1 OS = Caenorhabditis elegans GN = CNB-1 PE = 2 SV = 3|6239 | 1.01 | Q20804 | 19,654.9 | 5.85% | 1 | Calcium ion binding |
| 33 | 14331_CAEEL 14-3-3-like protein 1 OS = Caenorhabditis elegans GN = PAR-5 PE = 1 SV = 2|6 | D | P41932 | 28,173 | 7.26% | 2 | Protein domain specific binding, dauer entry, determination of adult lifespan |
| 34 | HSP17_CAEEL Heat shock protein HSP-16.48/HSP-16.49 OS = Caenorhabditis elegans GN | 8.43 | P02513 | 16,282 | 11.2% | 2 | Determination of adult lifespan, endoplasmic reticulum unfolded protein response to heat |
| Down regulated proteins due to treatment | |||||||
| 35 | IF5A1_CAEEL Eukaryotic translation initiation factor 5A-1 OS = Caenorhabditis eleg | D | P34563 | 17,849.3 | 6.83% | 2 | RNA binding, ribosome binding, translation elongation factor activity |
| 36 | Q20644_CAEEL Protein F52B5.3, partially confirmed by transcript evidence OS = Caenorhabditis elegans | D | Q20644 | 162,454 | 0.7% | 1 | Inorganic anion exchanger activity |
| 37 | EIF3C_CAEEL Eukaryotic translation initiation factor 3 subunit C OS = Caenorhabditis elegans | D | O02328 | 103,827 | 0.67% | 1 | Protein synthesis, translation initiation factor activity |
| 38 | RL18_CAEEL 60S ribosomal protein L18 OS = Caenorhabditis elegans GN = RPL-18 PE = 3 SV | 0.30 | O45946 | 51,385.2 | 3.72% | 1 | RNA binding, Structural constituent of ribosome |
| 39 | Q23440_CAEEL Protein ZK1307.8, confirmed by transcript evidence OS = Caenorhabditis elegans | 0.46 | Q23440 | 58,045.8 | 1.38% | 1 | Calcium ion binding, protein binding |
| 40 | A8DYR6_CAEEL Peroxiredoxin protein 2, isoform b OS = Caenorhabditis elegans GN | 0.80 | A8DYR6 | 21,767 | 14.9% | 3 | Thioredoxin peroxidase activity, determination of adult lifespan, hydrogen peroxide catabolic process |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kandasamy, S.; Fan, D.; Sangha, J.S.; Khan, W.; Evans, F.; Critchley, A.T.; Prithiviraj, B. Tasco®, a Product of Ascophyllum nodosum, Imparts Thermal Stress Tolerance in Caenorhabditis elegans. Mar. Drugs 2011, 9, 2256-2282. https://doi.org/10.3390/md9112256
Kandasamy S, Fan D, Sangha JS, Khan W, Evans F, Critchley AT, Prithiviraj B. Tasco®, a Product of Ascophyllum nodosum, Imparts Thermal Stress Tolerance in Caenorhabditis elegans. Marine Drugs. 2011; 9(11):2256-2282. https://doi.org/10.3390/md9112256
Chicago/Turabian StyleKandasamy, Saveetha, Di Fan, Jatinder Singh Sangha, Wajahatullah Khan, Franklin Evans, Alan T. Critchley, and Balakrishnan Prithiviraj. 2011. "Tasco®, a Product of Ascophyllum nodosum, Imparts Thermal Stress Tolerance in Caenorhabditis elegans" Marine Drugs 9, no. 11: 2256-2282. https://doi.org/10.3390/md9112256