Pentachlorophenol-Induced Cytotoxic, Mitogenic, and Endocrine-Disrupting Activities in Channel Catfish, Ictalurus punctatus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction
Materials and Methods
Chemicals and Reagents
Preparation and Culture of Catfish Hepatocytes
Cytotoxicity and Cell Proliferation Experiments
Sample Collection and Protein Determination
Western Blot Analysis for identification of Specific Cellular Proteins
Results
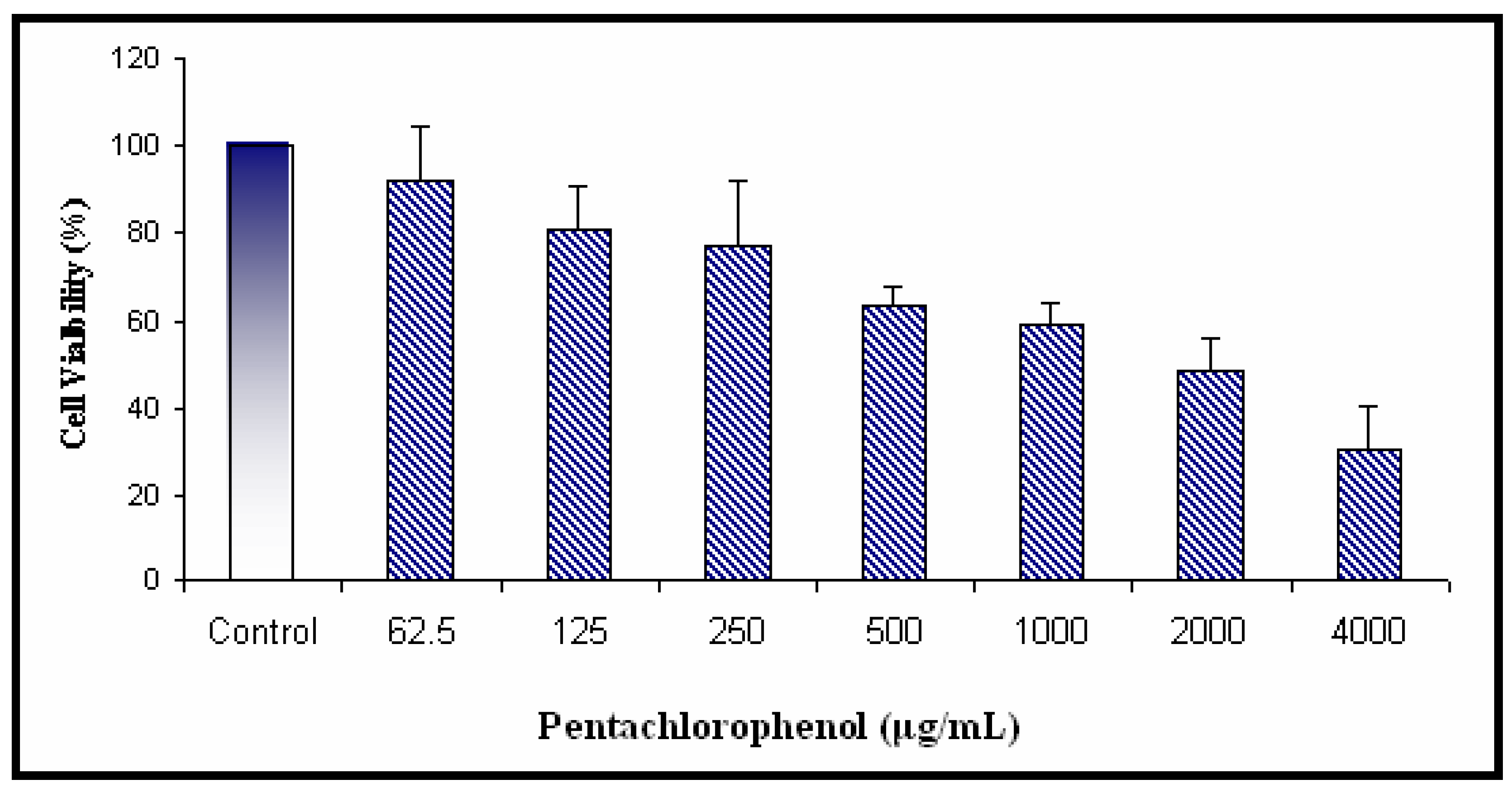
Cytotoxicity of PCP
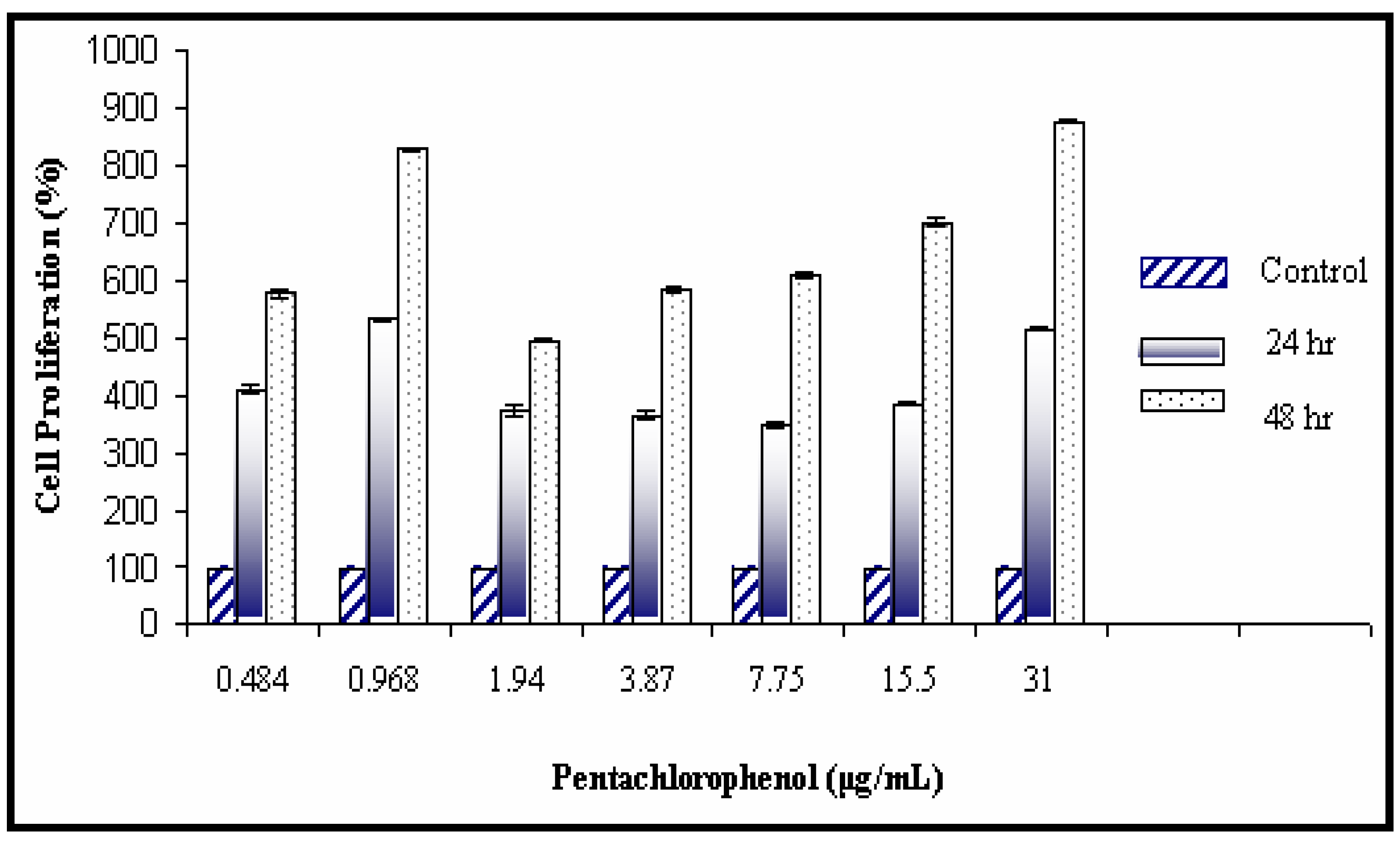
Mitogenic Activity of PCP
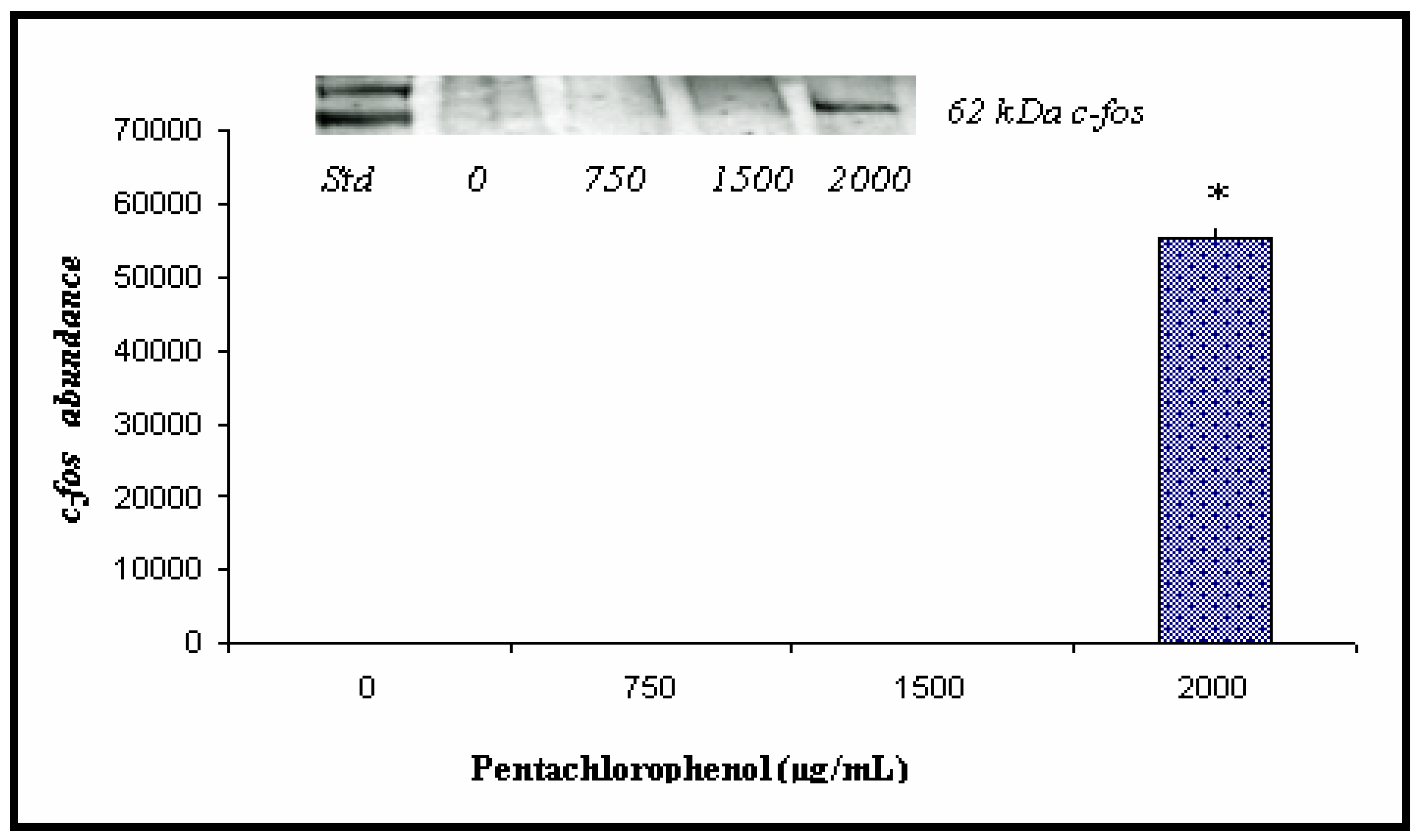
Western and Densitometric Analyses for c-fos Expression
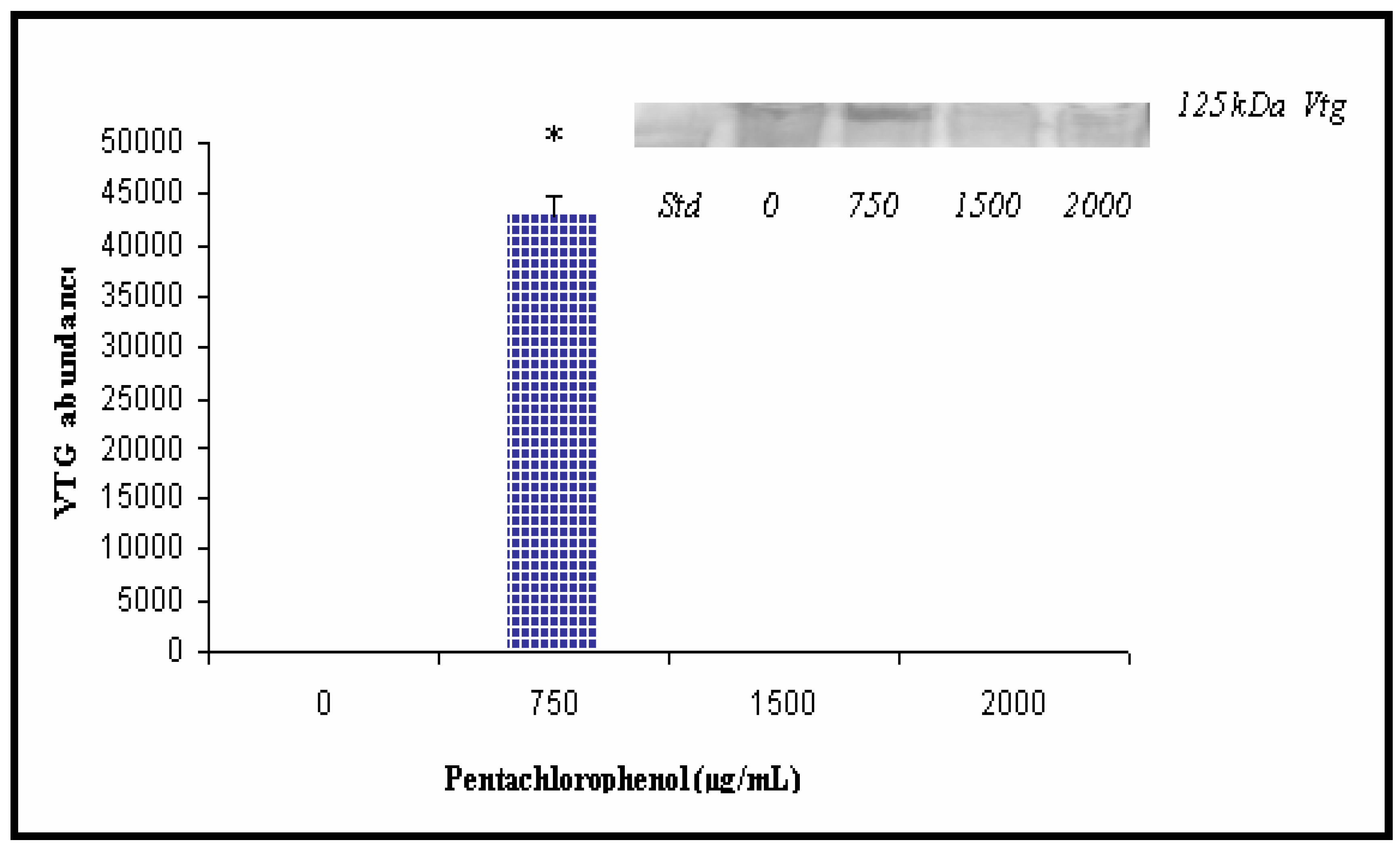
Western and Densitometric Analyses for Vtg Expression




Discussion
Cytotoxicity Studies
Mitogenic Activity of PCP in Catfish Hepatocytes
c-fos Expression in Catfish Hepatocytes
Vitellogenin Studies
Conclusions
Acknowledgments
References
- Dorsey, W.C.; Tchounwou, P.B.; Ishaque, A.B.; Shen, E. Transcriptional activation of stress genes and cytotoxicity in human liver carcinoma (HepG2) cells exposed to pentachlorophenol. Int. J. Mol. Sci. 2002, 3, 992–1007. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Special report on environmental endocrine disruption: an effects assessment and analysis. EPA630R96012. Prepared or the Risk Assessment forum; Washington, D.C., 1997. [Google Scholar]
- Burow, M.E.; Yang, T.; Collins-Burow, B.M.; Krajewski, S; Reed, J.C.; McLachlan, J.A.; Beckman, B.S. Effects of environmental estrogens on tumor necrosis factor α-mediated apoptosis in MCF-7 cells. Carcinogenesis 1999, 20(11), 2057–2061. [Google Scholar]
- Yasuhara, A.; Katami, T.; Shibamoto, T. Formation of PCDDs, PCDFs, and coplanar PCBs from incineration of various woods in the presence of chlorides. Environ. Sci Technol. 2003, 37, 1563–1567. [Google Scholar] [CrossRef]
- Huff, J.E.; Boyd, J.A.; Barrett, J.C. Cellular and molecular mechanisms of hormonal carcinogenesis: environmental influences. Wiley-Liss: New York, 1996. [Google Scholar]
- Cocco, P.; Kaxerouni, N.; Zahm, S.K. Cancer mortality and environmental exposure to DDE in United States. Environ. Health Perspectives. 2000, 108, 1–4. [Google Scholar] [CrossRef]
- Alexander, R. A developing toxic tort: lumber mills, log cabins, leukaemia, lymphomas and soft tissue sarcomas: the case against pentachlorophenol. The Consumer Law Page: Articles, 1999. [Google Scholar]
- Colborn, T.; vom Saal, F.S.; Soto, A.M. Developmental effects of endocrine-disrupting chemicals in wildlife and humans. Environ. Health Perspectives. 1993, 101(5), 256–257. [Google Scholar]
- Guillette, L.J., Jr.; Gross, T.S.; Masson, G.R.; Matter, J.M.; Percival, H.F.; Woodward, A.R. Developmental abnormalities of the gonad and abnormal sex hormone concentrations in juvenile alligators from contaminated and control lakes in Florida. Environmental Health Perspectives 1994, 102, 608–688. [Google Scholar]
- Kavlock, R.; Ankley, G.; Francis, E.; Gray, E.; McMaster, S.; Reese, D.; Sayles, G.; Sergeant, A.; Vallero, D. Research plan for endocrine disruptors: a report of the U.S. EPA; EPA600R98087; 1998. [Google Scholar]
- Beard, A.P.; Barthlewski, P.M.; Chandolia, R.K.; Honaramooz, A.; Rawlings, N.C. Reproductive and endocrine function in rams exposed to the organochlorine pesticides lindane and PCP from conception. J. Reproductive Fertility 1999, 115, 303–314. [Google Scholar] [CrossRef]
- Beard, A.P.; Bartlewski, P.M.; Rawlings, N.C. Endocrine and reproductive function in ewes exposed to the organochlorine pesticides lindane or pentachlorophenol. J. Toxicol. Environ. Health. 1999, 56(1), 23–46. [Google Scholar]
- Colosio, C.; Maroni, M.; Barcellini, W.; Meroni, P.; Alcini, D.; Colombi, A.; Cavallo, D.; Foa, V. Toxicological and immune findings in workers exposed to pentachlorophenol (PCP). Archives of Environmental Health 1993, 48(2), 81–88. [Google Scholar]
- Daniel, V.; Wolfgang, H.; Klausdieter, B. and in patients with elevated pentachlorophenol (PCP) blood levels. Archives of Environmental Health 1995, 50(4), 287–292. [Google Scholar]
- Miller, W.R. Sharpe: Environmental oestrogens and human reproductive cancers. Endocrine-Related Cancer 1998, 5, 69–96. [Google Scholar] [CrossRef]
- Peper, M.; Ertl, M.; Gerhard, I. Long-term exposure to wood-preserving chemicals containing pentachlorophenol and lindane is related to neurobehavioral performance in women. American Journal of Industrial Medicine 1999, 35, 632–641. [Google Scholar] [CrossRef]
- Rawlings, N.C.; Cook, S.J.; Waldbillig, D. Effects of the pesticides carbofuran, chlorpyrifos, dimehoate, lindane, triallte, triluralin, 2,4-D, and pentachlorophenol on the metabolic endocrine and reproductive endocrine system in ewes. J. Toxicol. Environ. Health 1998, 54, 21–36. [Google Scholar]
- Danzo, B.J. Environmental xenobiotics may disrupt normal endocrine function by interfering with the binding of physiological ligands to steroid receptor and binding proteins. Environmental Health Perspectives 1997, 105, 294–301. [Google Scholar] [CrossRef]
- Illinois Environmental Protection Agency. Illinois EPA Endocrine Disruptors Strategy Document; 1997. [Google Scholar]
- Fry, D.M. Reproductive effects in birds exposed to pesticides and industrial chemicals. Environmental Health Perspectives 1995, 103 (supplement 7), 165–171. [Google Scholar] [CrossRef]
- Sumpter, J.P.; Jobling, S. Vitellogenesis as a biomarker for estrogenic contamination of the aquatic environment. Environmental Health Perspectives 1995, 103, 173–178. [Google Scholar] [CrossRef]
- Christiansen, T.; Korsgaard, B.; Jespersen, A. Effects of nonylphenol and 17β-oestradiol on vitellogenin synthesis, testicular structure and cytology in male eelpout Zoarces viviparus. J. Experimental Bio. 1998, 201, 179–192. [Google Scholar]
- Hennies, M.; Wiesmann, M.; Allner, B.; Sauerwein, H. Vitellogenin in carp (Cyprinus carpio) and (Perca fluviatilis: purification, characterization and development of an ELISA for the detection of estrogenic effects. Sci. Total Environ 2003, 309, 93–103. [Google Scholar] [CrossRef]
- Wallace, R.A. Vitellogenesis and oocyte growth in nonmammlian vertebrates. In Developmental biology: a comprehensive synthesis vol 1: Oogenesis; Browder, L.W., Ed.; New York, NY, 1985; pp. 127–177. [Google Scholar]
- Palmer, B.D.; Palmer, S.K. Vitellogenin induction by xenobiotic estrogens in the red-eared turtle and the African clawed frog. Environmental Health Perspectives 1995, 103, 19–25. [Google Scholar] [CrossRef]
- Palmer, B.D.; Selcer, K.W. Vitellogenin as a biomarker for xenobiotic estrogens: a review. In Environmental toxicology and risk assessment: biomarkers and risk assessment; Volume 5, Bengton, D.A., Henshel, D.S., Eds.; ASTM Special Technical Publication nr 1306; American Society for Testing and Materials: Philadelphia (PA), 1996; pp. 3–22. [Google Scholar]
- Kinzell, J.H.; Ames, N.K.; Sleight, S.D.; Krehbiel, J.D.; Kuo, C.; Zabik, M.J.; Shull, L.R. Sub chronic administration of technical pentachlorophenol to lactating dairy cattle: performance, general health, and pathologic changes. J. Dairy Sci. 1981, 64, 42–51. [Google Scholar] [CrossRef]
- Wood, S.; Rom, V.N.; White, G.L.; Logan, D.C. Pentachlorophenol poisoning. J. Occup. Med. 1983, 25, 527–530. [Google Scholar]
- Menzer, R.E.; Nelson, J.O. Water and soil pollutants. In Casarett and Doull’s Toxicology, 3rd Ed.; Klaassen, C.D., Amdur, M.O., Doull, J., Eds.; Macmillan Publishing Co.: New York, 1986; pp. 825–853. [Google Scholar]
- Bevenue, A.; Beckman, H. Pentachlorophenol: a discussion of its properties and its occurrence as a residue in human and animal tissues. Residue Review 1967, 19, 83–134. [Google Scholar]
- Cirelli, D.P. Patterns of pentachlorophenol usage in the United States of America: an overview. In pentachlorophenol: chemistry, pharmacology, and environmental toxicology; Rao, K.r., Ed.; Plenum Press: New York, 1978; pp. 13–18. [Google Scholar]
- U.S. National Toxicology Program (NTP 1991). Pentachlorophenol, NTP Chemical Repository. Report. Radian Corporation. 1991.
- World Health Organization (WHO). Pentachlorophenol. Environmental Health Criteria 71. Geneva, 1987. [Google Scholar]
- Office of Ground Water and Drinking Water (OGWDW). Technical Drinking Water and Health Contaminant Specific Fact Sheet. Technical fact sheet on pentachlorophenol. 1998. URL: http://www/epa.gov/OGWDW/dwh/tsoc/pentach.
- Pignatello, J.J.; Martinson, M.M.; Steiert, J.G.; Carlson, R.E.; Crawford, R.L. Biodegradation and photolysis of pentachlorophenol in artificial freshwater streams. Applied Environmental Microbiology. 1983, 46, 1024–1031. [Google Scholar]
- U.S. Department of Health and Human Services (DHHS). Toxicological Profile for Pentachlorophenol. Agency for Toxic Substance and Disease Registry; Draft; 1992. [Google Scholar]
- Extension Toxicology Network (Extoxnet). Pentachlorophenol. Pesticide Information Profile. A Pesticide Information Project of Cooperative Extension Offices of Cornell University, Michigan State University, Oregon State University, and University of California at Davis. 1998. URL: http://pmep.cce.cornell.edu./profiles/extoxnet/metiram-propoxur/ pentachlorophenol-ext.html.
- Agency for Toxic Substances and Disease Registry (ATSDR). Toxicological Profile for Pentachlorophenol (Draft); Public Health Service, U.S. Department of Health and Human Services: Atlanta, GA; p. 1992.
- U.S. Environmental Protection Agency. Integrated Risk. Information System (IRIS) on Pentachlorophenol. Environmental Criteria and Assessment Office, Office of Health and Environmental Assessment, Office of Research and Development: Cincinnati, OH, 1993. [Google Scholar]
- Wagner, S.L. Clinical Toxicology of Agricultural Chemicals. Noyes Data Corporation. 1983.
- Welsh, J.J.; Collins, T.F.; Black, T.N.; Graham, S.L.; O’Donnell, M.W., Jr. Teratogenic potential of purified pentachlorophenol and pentachloroanisole in sub chronically exposed Spague-Dawley rats. Food Chemical Toxicology. 1987, 25, 163–172. [Google Scholar] [CrossRef]
- Beard, A.P.; McRae, A.C.; Rawlings, N.C. Reproductive efficiency in mink (Mustela vison) treated with the pesticides lindane, carofuran and pentachlorophenol. Journal of Reproductive Fertility 1997, 111, 21–28. [Google Scholar] [CrossRef]
- Owens, K.D.; Baer, K.N. Modifications of the topical Japanese medaka (Oryzias latipes) embryo larval assay for assessing developmental toxicity of pentachlorophenol and p, p′ -Dichlorodiphenyltrichloroethane. Ecotoxicology Environmental Safety 2000, 47, 87–95. [Google Scholar] [CrossRef]
- Gerhard, I.; Frick, A.; Monga, B.; Runnebaum, B. Pentachlorophenol exposure in women with gynaecological and endocrine dysfunction. Environmental Research 1999, 80, 383–388. [Google Scholar] [CrossRef]
- Dorsey, W.C.; Tchounwou, P.B. CYP1A1, HSP70, p53, and c-fos expression in human liver carcinoma cells (HepG2) exposed to pentachlorophenol. The Instrumentation, Systems, and Automation Society, ISA. 2003, 437, 389–396. [Google Scholar]
- Pierce, R.H., Jr.; Victor, D.M. The fate of pentachlorophenol in an aquatic ecosystem. In pentachlorophenol: chemistry, pharmacology, and environmental toxicology; Rao, K.R., Ed.; Plenum Press: New York, 1978; pp. 41–52. [Google Scholar]
- Marino, M.; Acconcia, F.; Bresciana, R.; Weiz, A.; Trentalance, A. Distinct nongenomic signal transduction pathways controlled by 17β-estradiol regulate DNA synthesis and Cyclin D1 gene transcription in HepG2 cells. Molecular Biology of the Cell 2002, 13, 3720–3729. [Google Scholar] [CrossRef]
- Schreiber, M.; Kolbus, A.; Piu, F.; Szabowski, A.; Mohle-Steinlein, U.; Tian, J.; Karin, M.; Angel, P.; Wagner, E.F. Control of cell cycle progression by c-jun is p53 dependent. Genes & Development 1999, 13, 617–619. [Google Scholar]
- Wang, Z.; Templeton, D.M. Induction of c-fos proto-oncogene in mesangial cells by cadmium. J. Bio. Chem. 1998, 273, 73–79. [Google Scholar] [CrossRef]
- Gronen, S.; Denslow, N.; Manning, S.; Barnes, S.; Barnes, D.; Brouwer, M. Serum vitellogenin levels and reproductive impairment of male Japanese Medaka (oryzias latipes) exposed to 4-tert-octylphenol. Environmental Health Perspectives 1999, 107, 385–390. [Google Scholar] [CrossRef]
- Heppell, S.A.; Denslow, N.D.; Folmar, L.C.; Sullivan, C.V. Universal assay of vitellogenin as a biomarker for environmental estrogens. Environmental Health Perspectives 1995, 103 (Supplement 7), 9–15. [Google Scholar] [CrossRef]
- Monteverdi, G.H.; Di Giulio, R.T. An enzyme-linked immunosorbent assay for estrogenicity using primary hepatocyte cultures from the channel catfish (Ictalurus punctatus). Archives of Environmental Contamination and Toxicology 1999, 37, 62–69. [Google Scholar] [CrossRef]
- Soto, A.M.; Chung, K.L.; Sonnenschein, C. The pesticides endosulfan, toxaphene and dieldrin have estrogenic effects on human estrogen-sensitive cells. Environmental Health Perspective 1994, 102, 380–383. [Google Scholar] [CrossRef]
- Dewailly, E.; Ayotte, P.; Dodin, S. Could the rising levels of estrogen receptor in breast cancer be due to estrogenic pollutants? Journal of the National Cancer Institute 1997, 89, 888. [Google Scholar] [CrossRef]
- Cooper, R.L.; Kavlock, R.J. Endocrine disruptors and reproductive development: a weight-of-evidence overview. Journal of Endocrinology 1997, 152, 159–166. [Google Scholar] [CrossRef]
- Kunkel, J.G. Serum and egg vitellogenin measurement in the Atlantic cod, Gadus morhua, and its relationship to ovarian development. Proposal response to CMER NOAA/NMFS Research Topic. Biochemical indices of maturity and egg auality in Atlantic cod. 1996. [Google Scholar]
- Folmar, L.C.; Denslow, N.D.; Rao, V.; Chow, M.; Crain, A.; Enblom, J.; Marcino, J.; Guillette, L.J. Vitellogenin induction and reduced serum testosterone concentration in Feral Male Carp (Cyprinus carpio) captured near a major metropolitan sewage treatment plant. Environmental Health Perspectives 1996, 104, 1096. [Google Scholar] [CrossRef]
- Lye, C.M.; Frid, C.L.J.; Gill, M.E.; McCormick, D. Abnormalities in the reproductive health of Flounder Platichthys flesus exposed to effluent from a sewage treatment works. Marine Pollution Bulletin 1997, 34, 34–41. [Google Scholar]
© 2004 MDPI. All rights reserved.
Share and Cite
Dorsey, W.C.; Tchounwou, P.B. Pentachlorophenol-Induced Cytotoxic, Mitogenic, and Endocrine-Disrupting Activities in Channel Catfish, Ictalurus punctatus. Int. J. Environ. Res. Public Health 2004, 1, 90-99. https://doi.org/10.3390/ijerph2004020090
Dorsey WC, Tchounwou PB. Pentachlorophenol-Induced Cytotoxic, Mitogenic, and Endocrine-Disrupting Activities in Channel Catfish, Ictalurus punctatus. International Journal of Environmental Research and Public Health. 2004; 1(2):90-99. https://doi.org/10.3390/ijerph2004020090
Chicago/Turabian StyleDorsey, Waneene C., and Paul B. Tchounwou. 2004. "Pentachlorophenol-Induced Cytotoxic, Mitogenic, and Endocrine-Disrupting Activities in Channel Catfish, Ictalurus punctatus" International Journal of Environmental Research and Public Health 1, no. 2: 90-99. https://doi.org/10.3390/ijerph2004020090