Analysis of the Toll-Like Receptor 2-2 (TLR2-2) and TLR4 mRNA Expression in the Intestinal Mucosal Immunity of Broilers Fed on Diets Supplemented with Nickel Chloride


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chickens and Diets
2.2. Detection of TLR2 and TLR4 mRNA Expression Levels in the Intestinal Mucosa and the Cecal Tonsil by qRT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Accession Number | Primer | Primer Sequence (5′ → 3′) |
|---|---|---|---|
| TLR2-2 | AB046533 | F | AGGCACTTGAGATGGAGCAC |
| R | CCTGTTATGGGCCAGGTTTA | ||
| TLR4 | AY064697 | F | AGTCTGAAATTGCTGAGCTCAAAT |
| R | GCGACGTTAAGCCATGGAAG | ||
| β-Actin | L08165 | F | TGCTGTGTTCCCATCTATCG |
| R | TTGGTGACAATACCGTGTTCA |
2.3. Statistical Analysis
3. Results
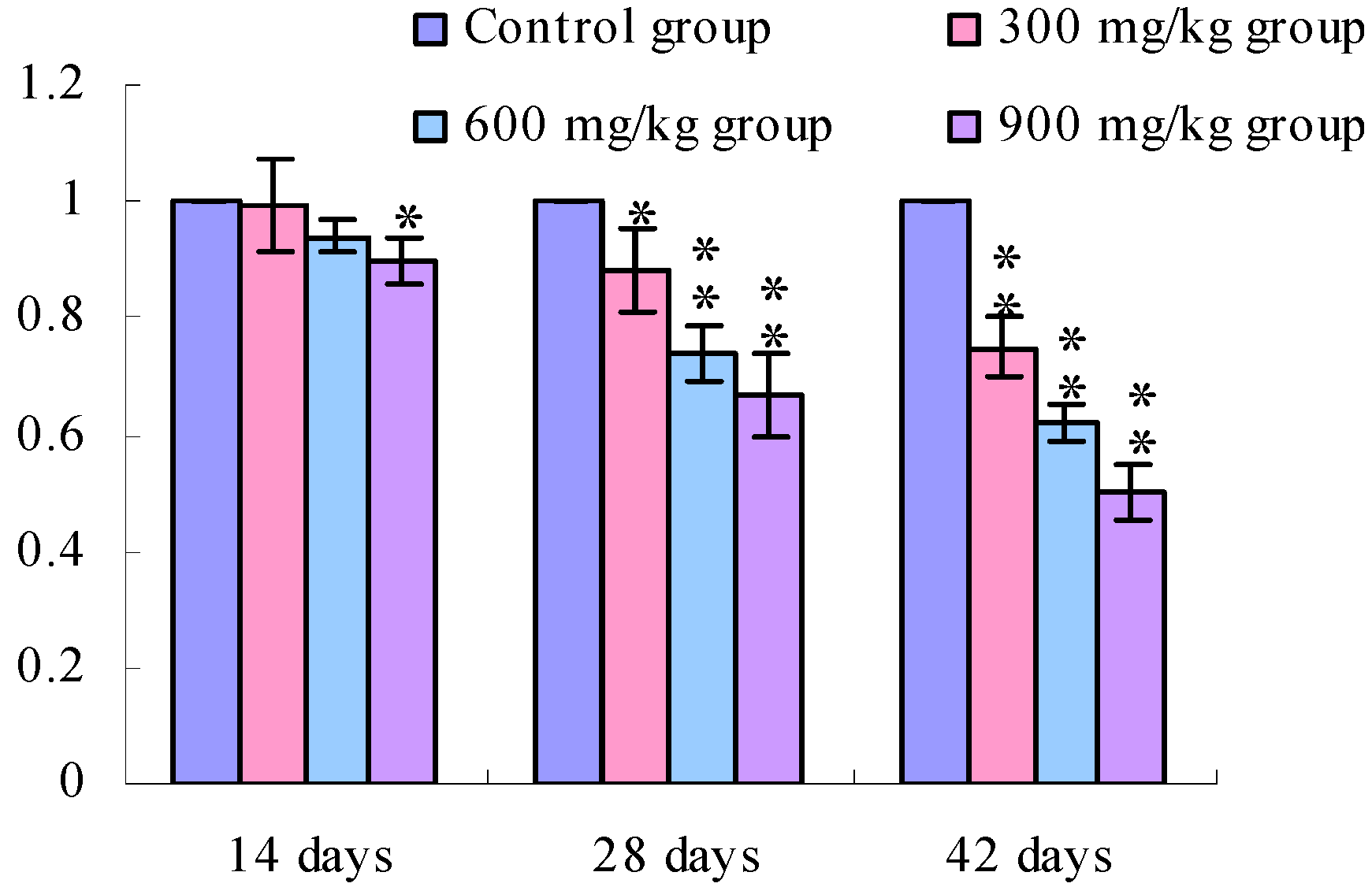
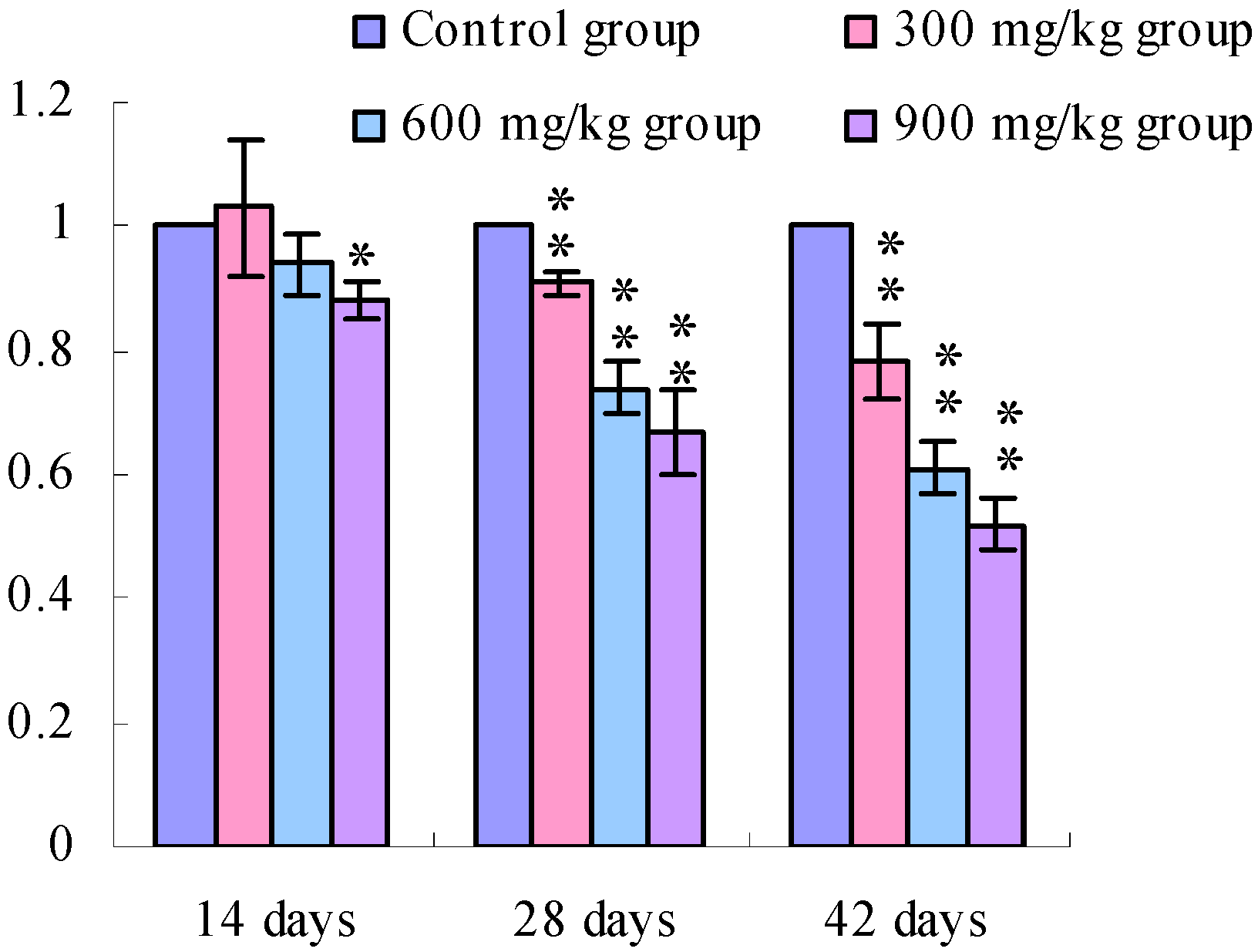
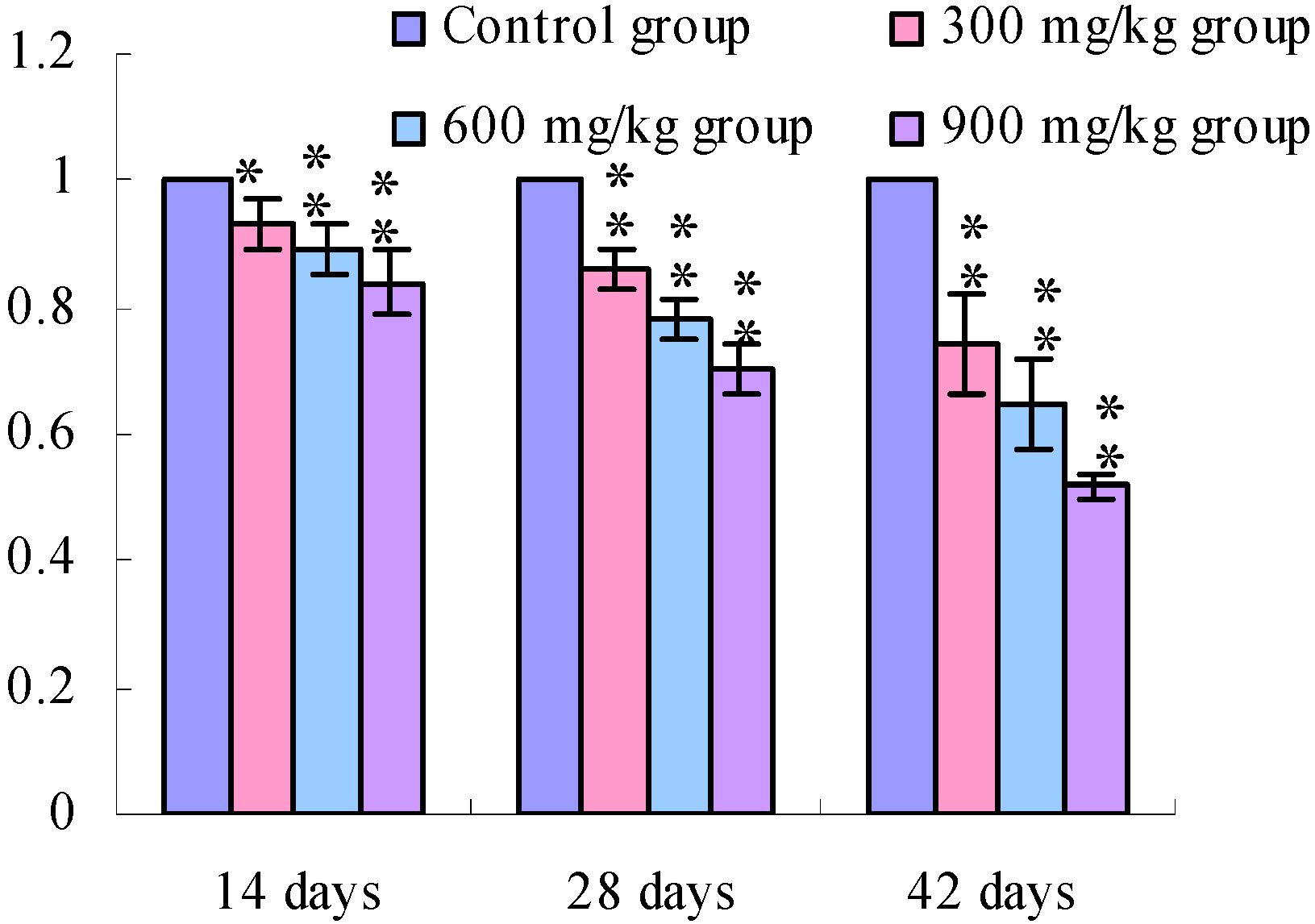
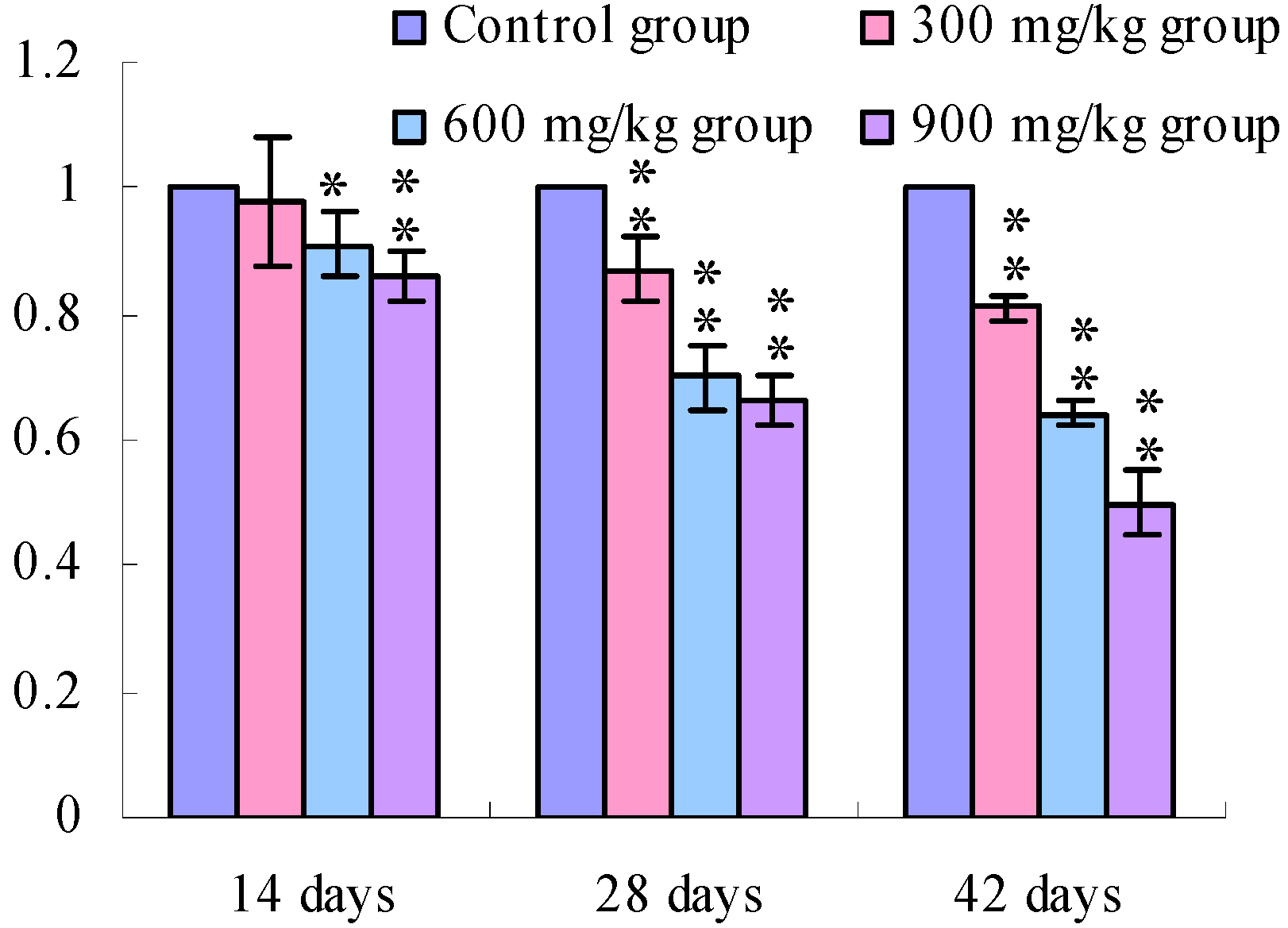
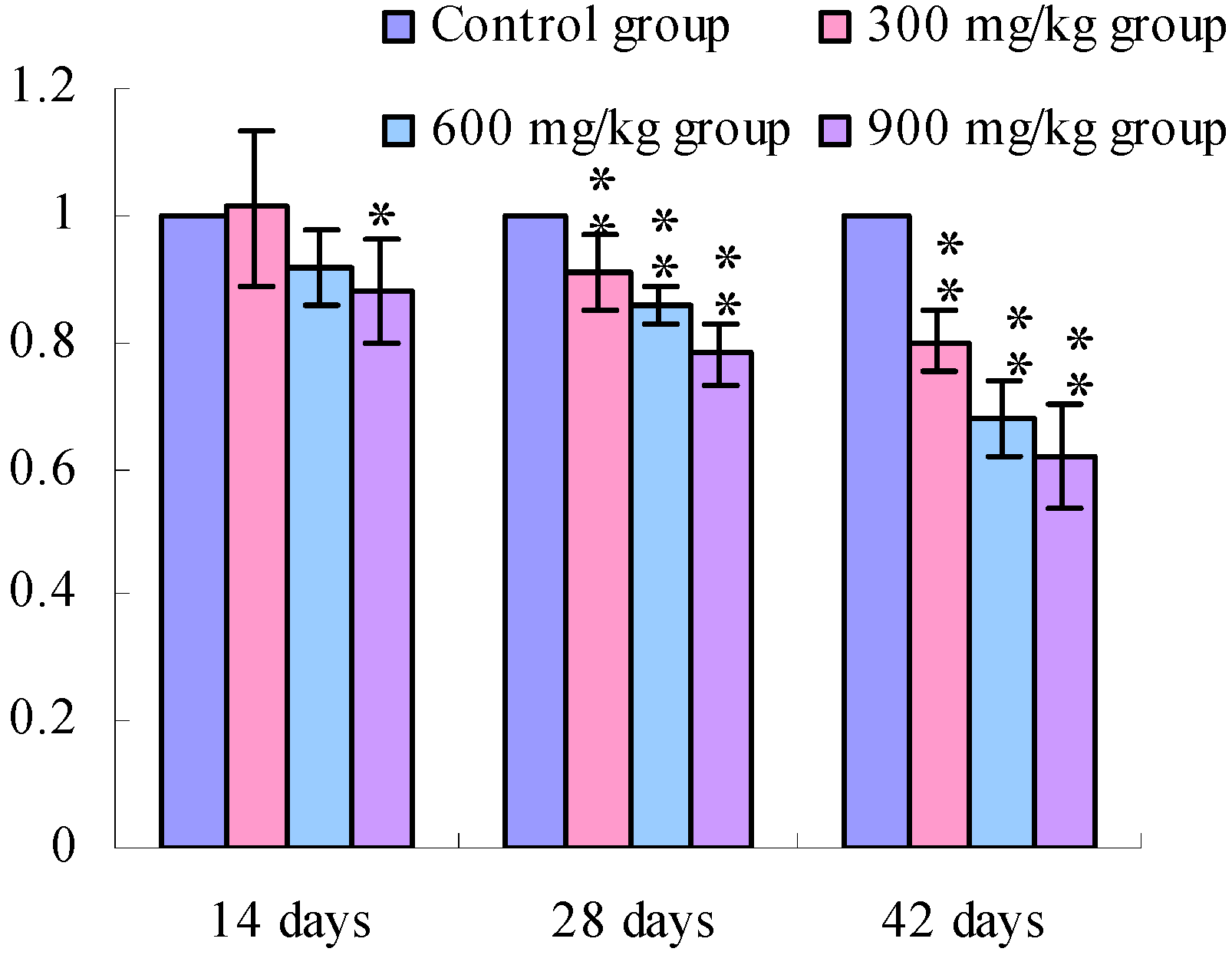
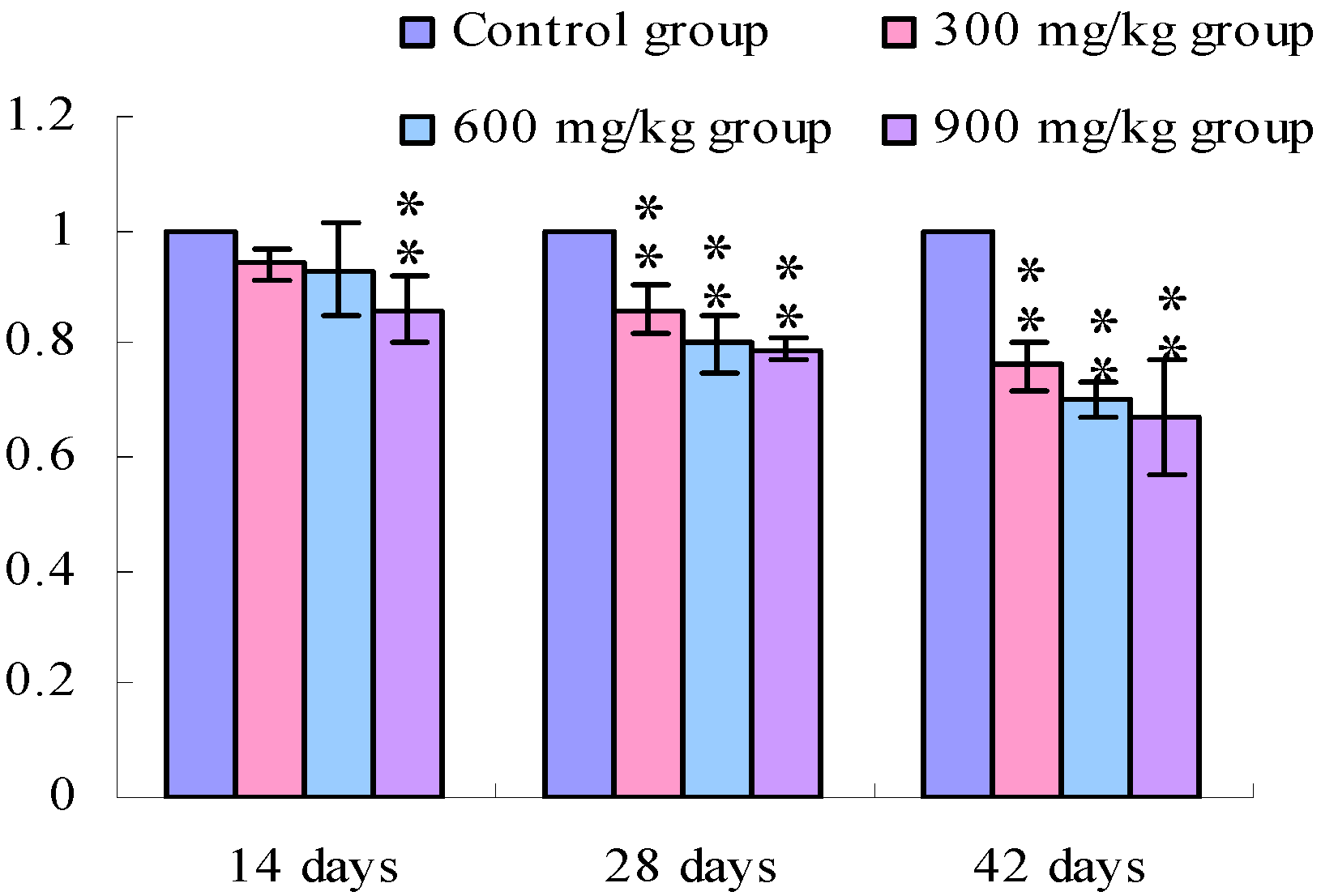
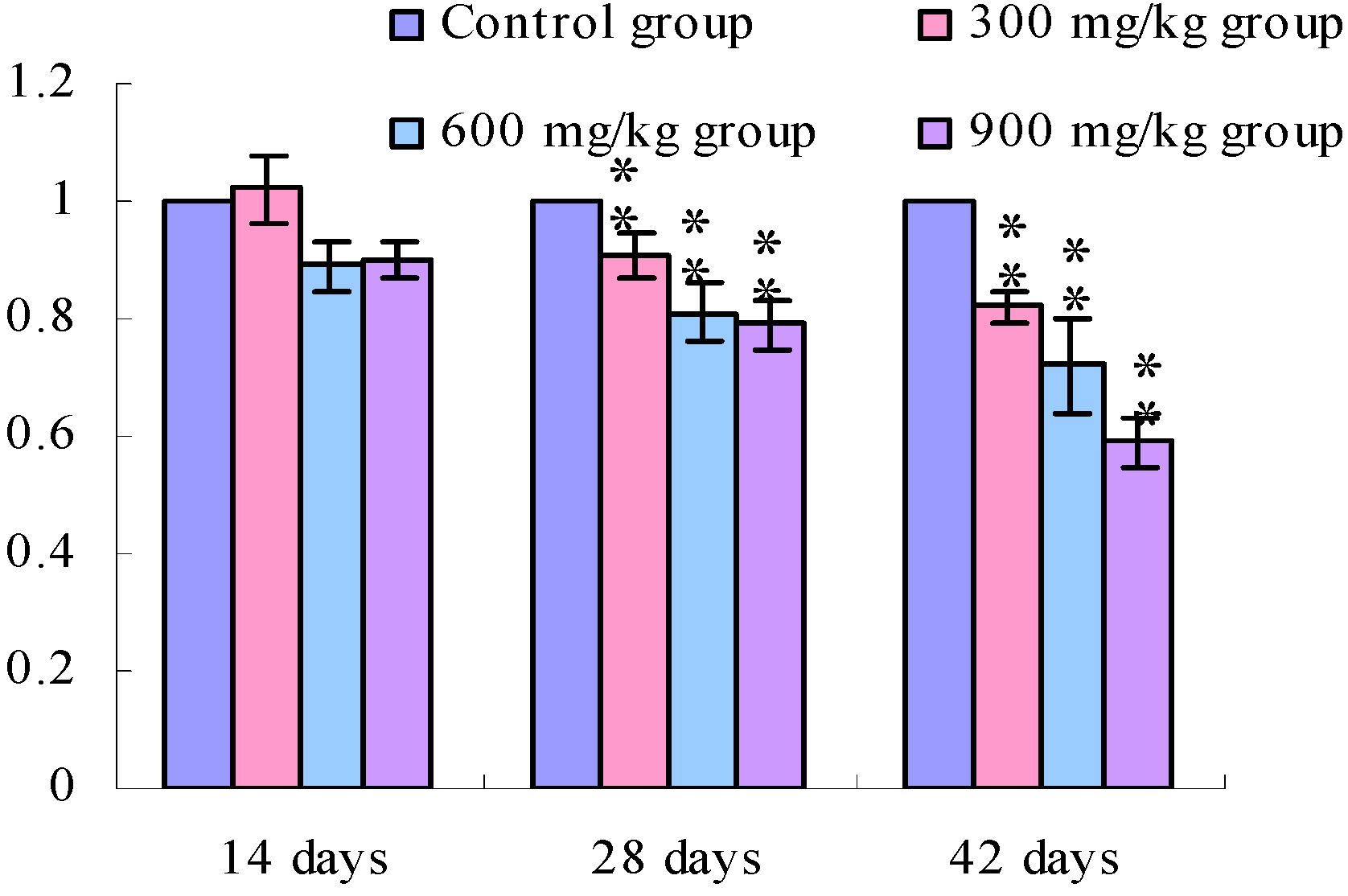
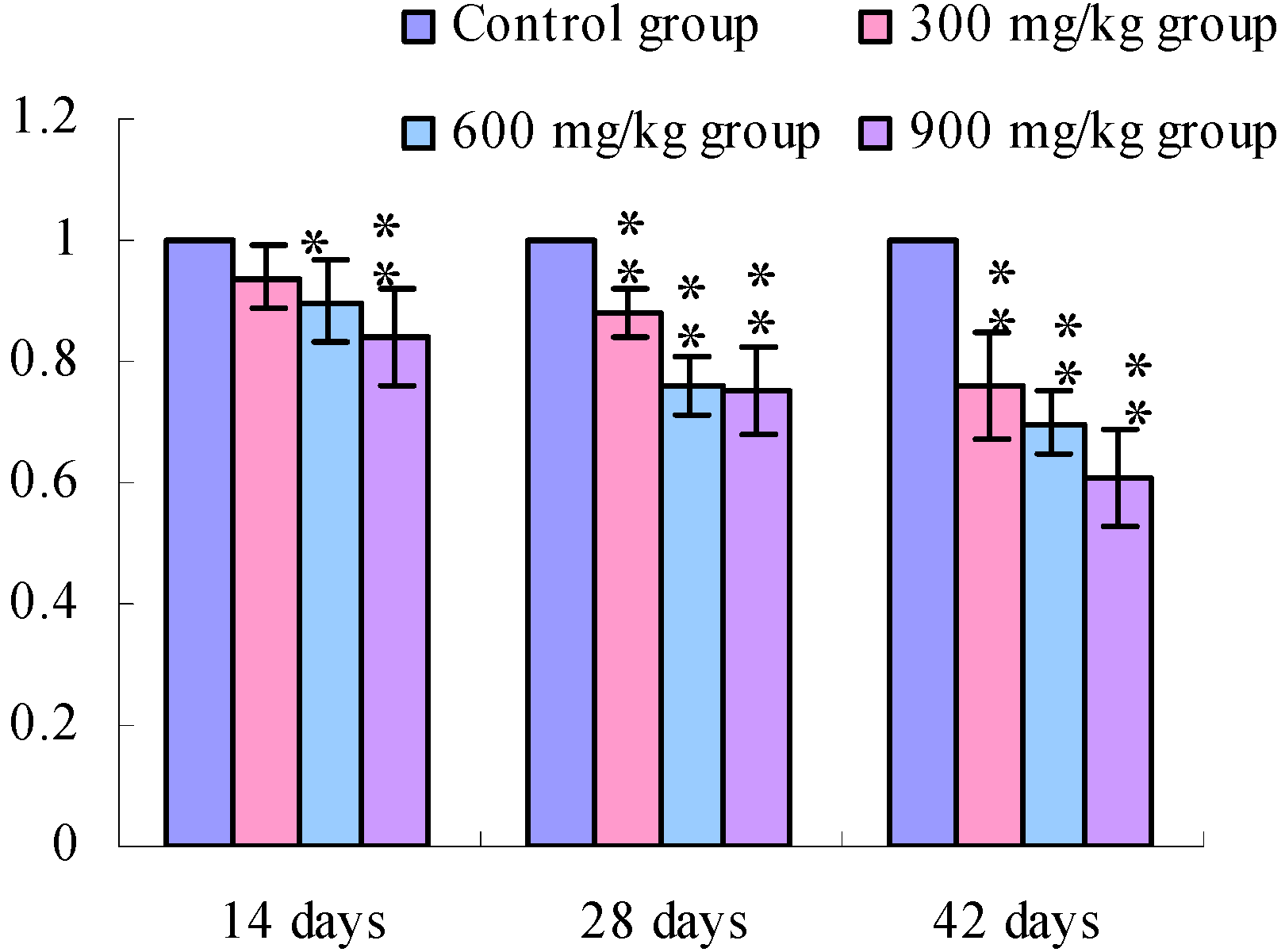
3.1. Changes of the TLR2-2 mRNA Expression Levels

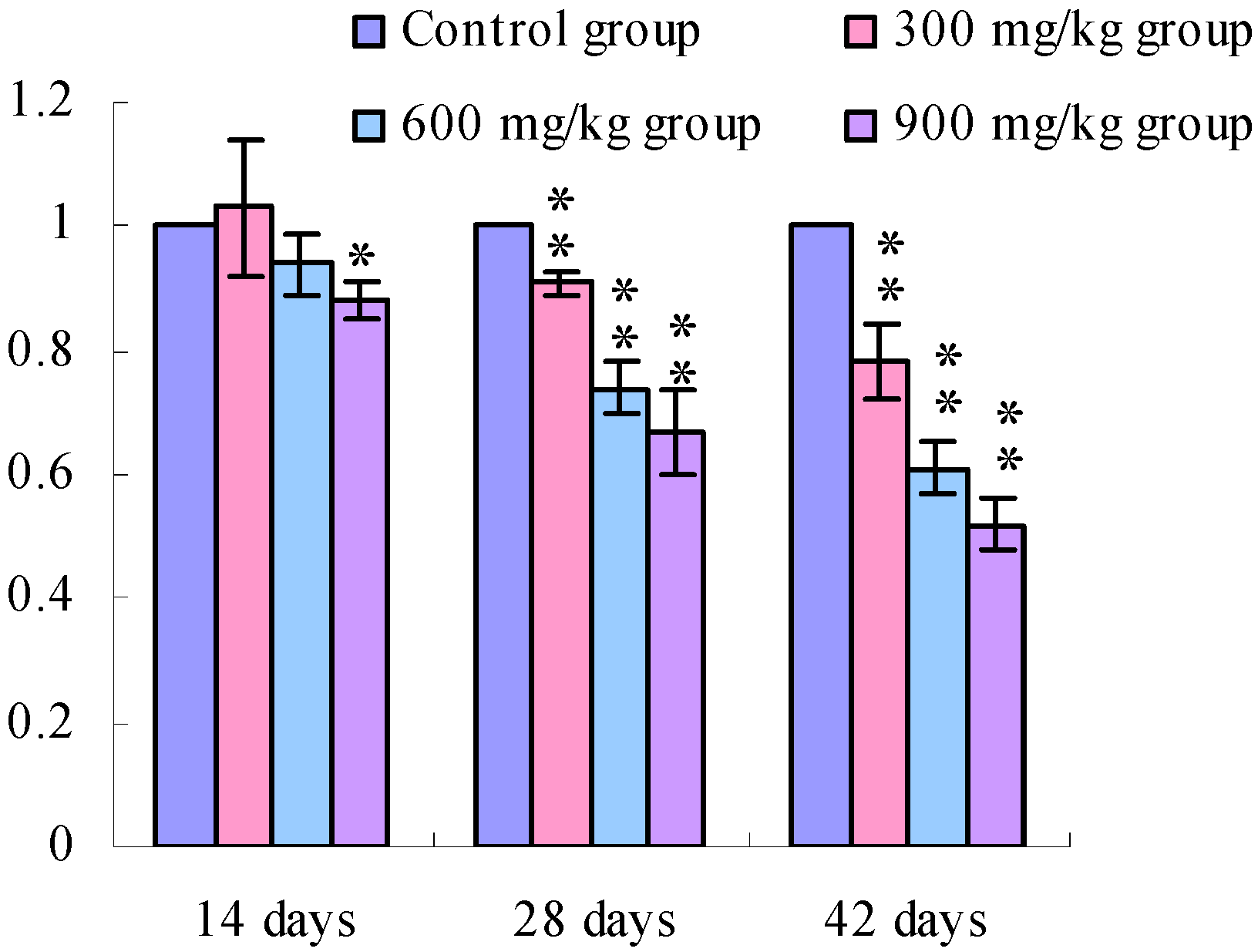
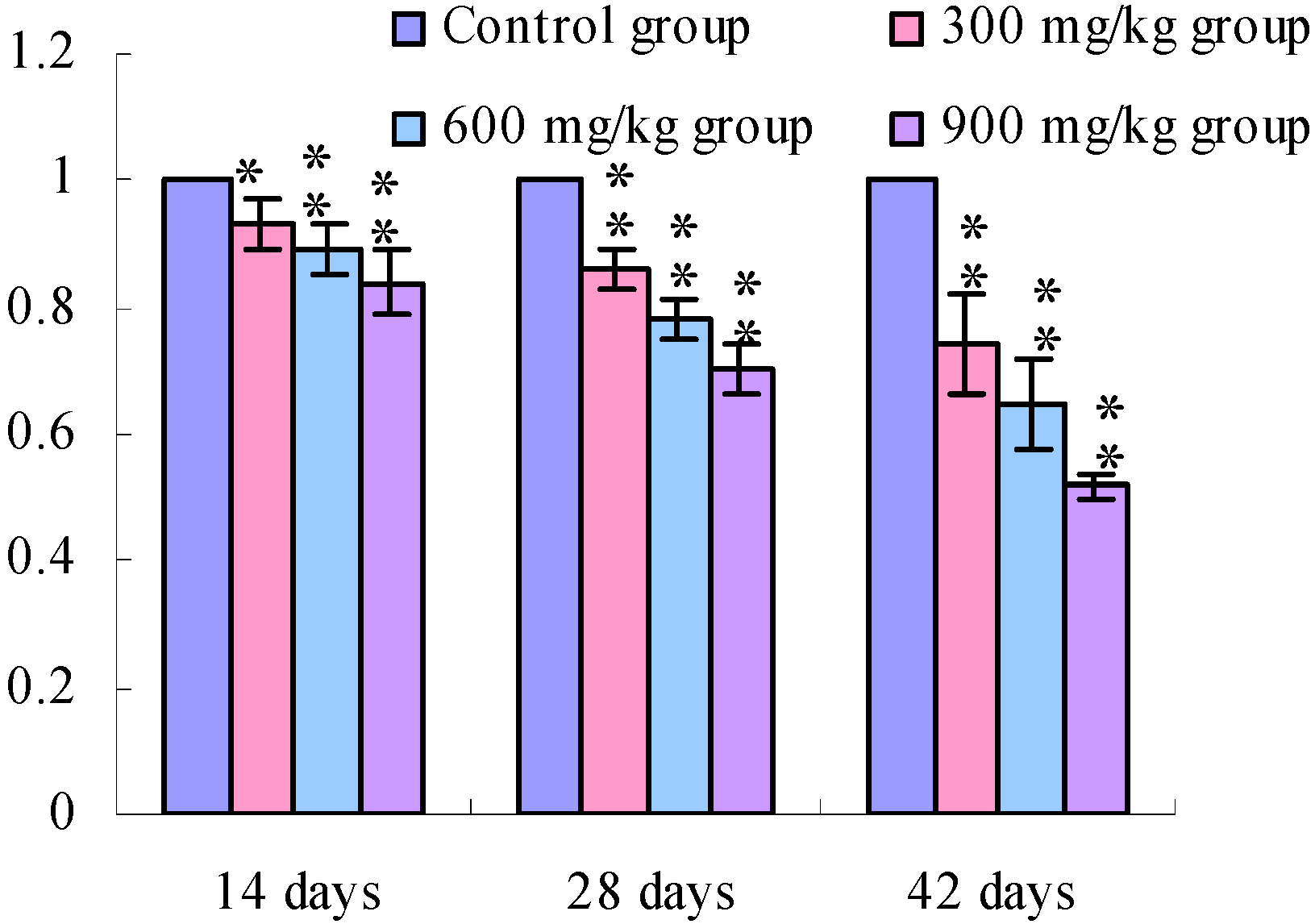
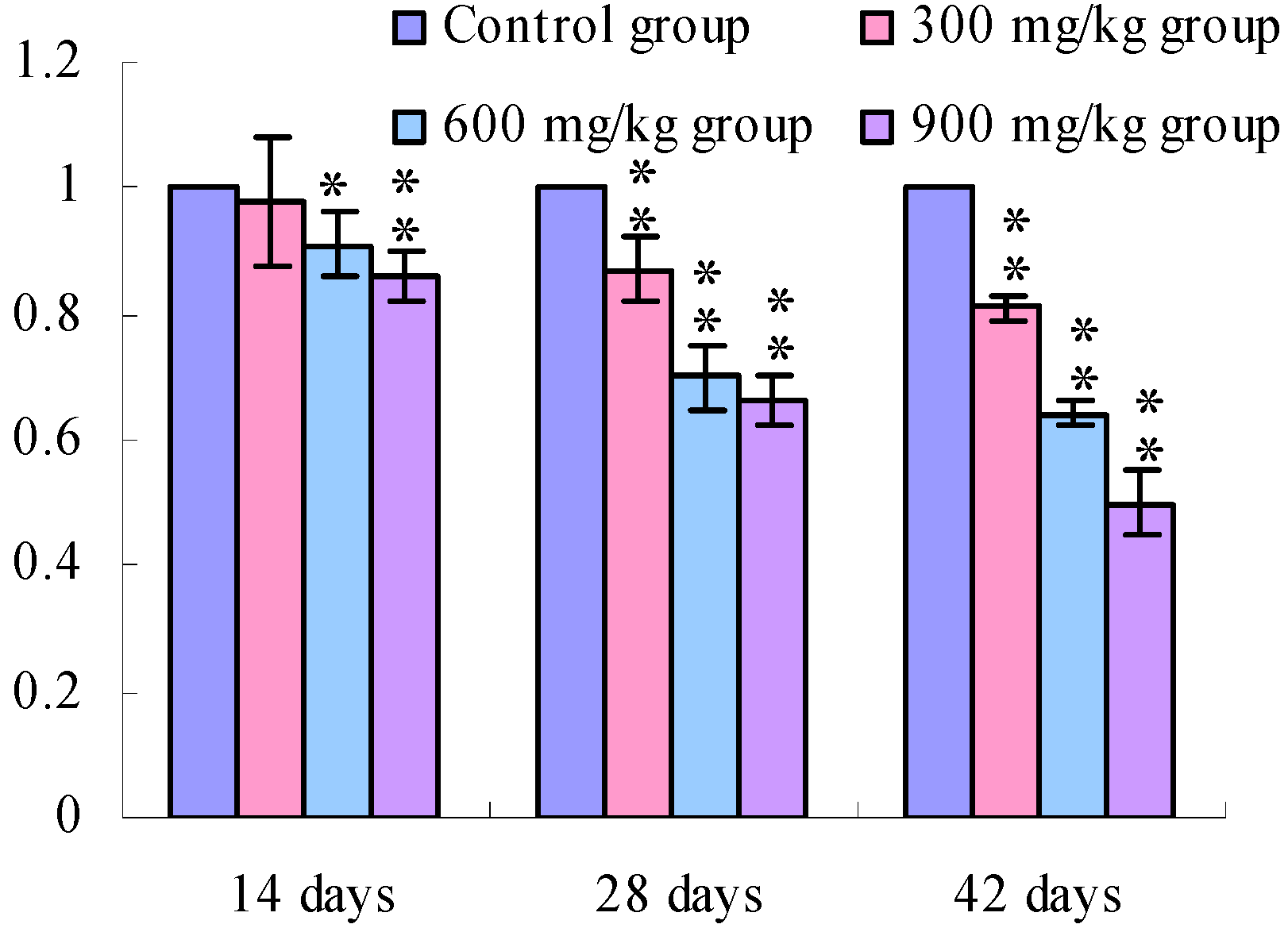
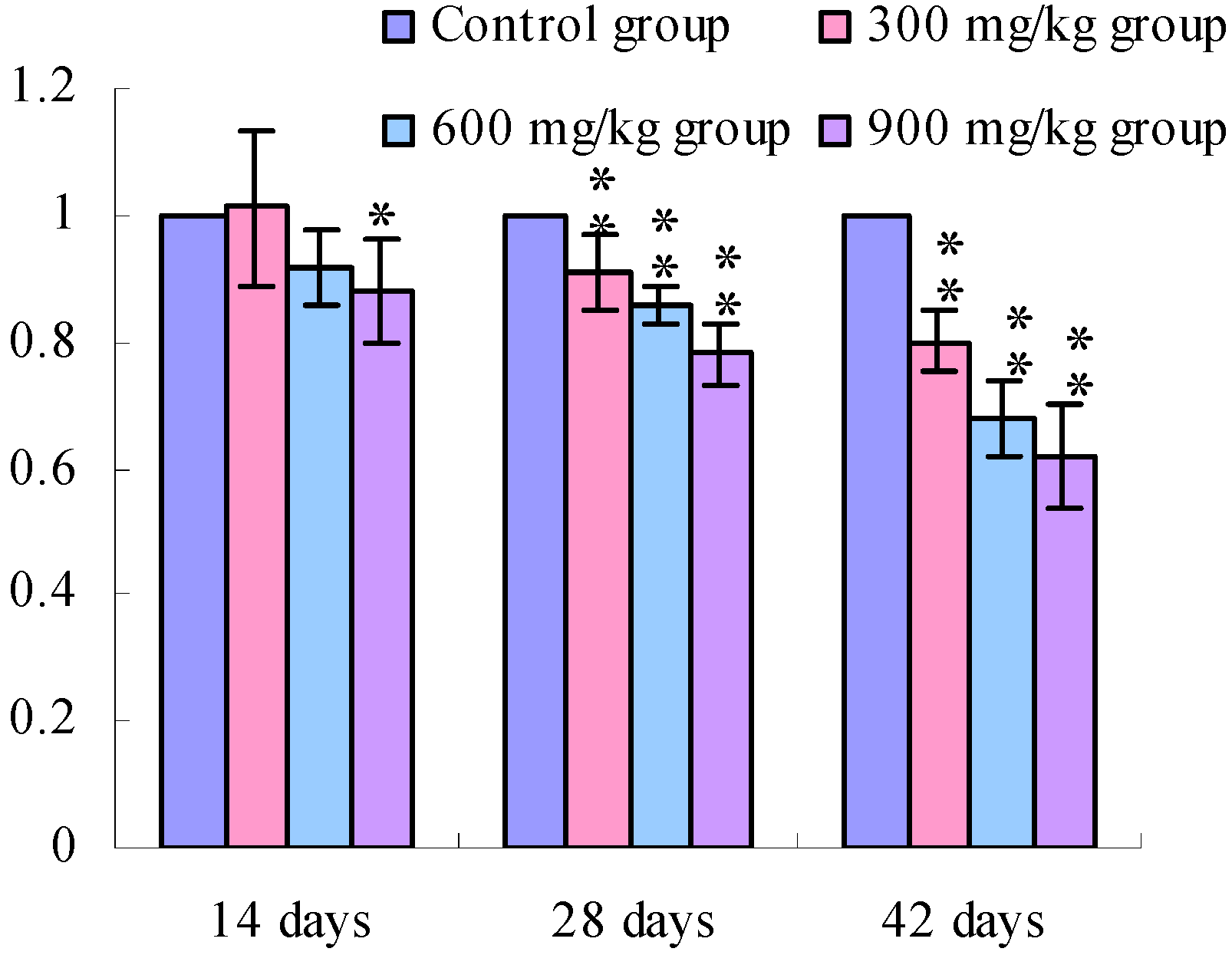
3.2. Changes of the TLR4 mRNA Expression Levels
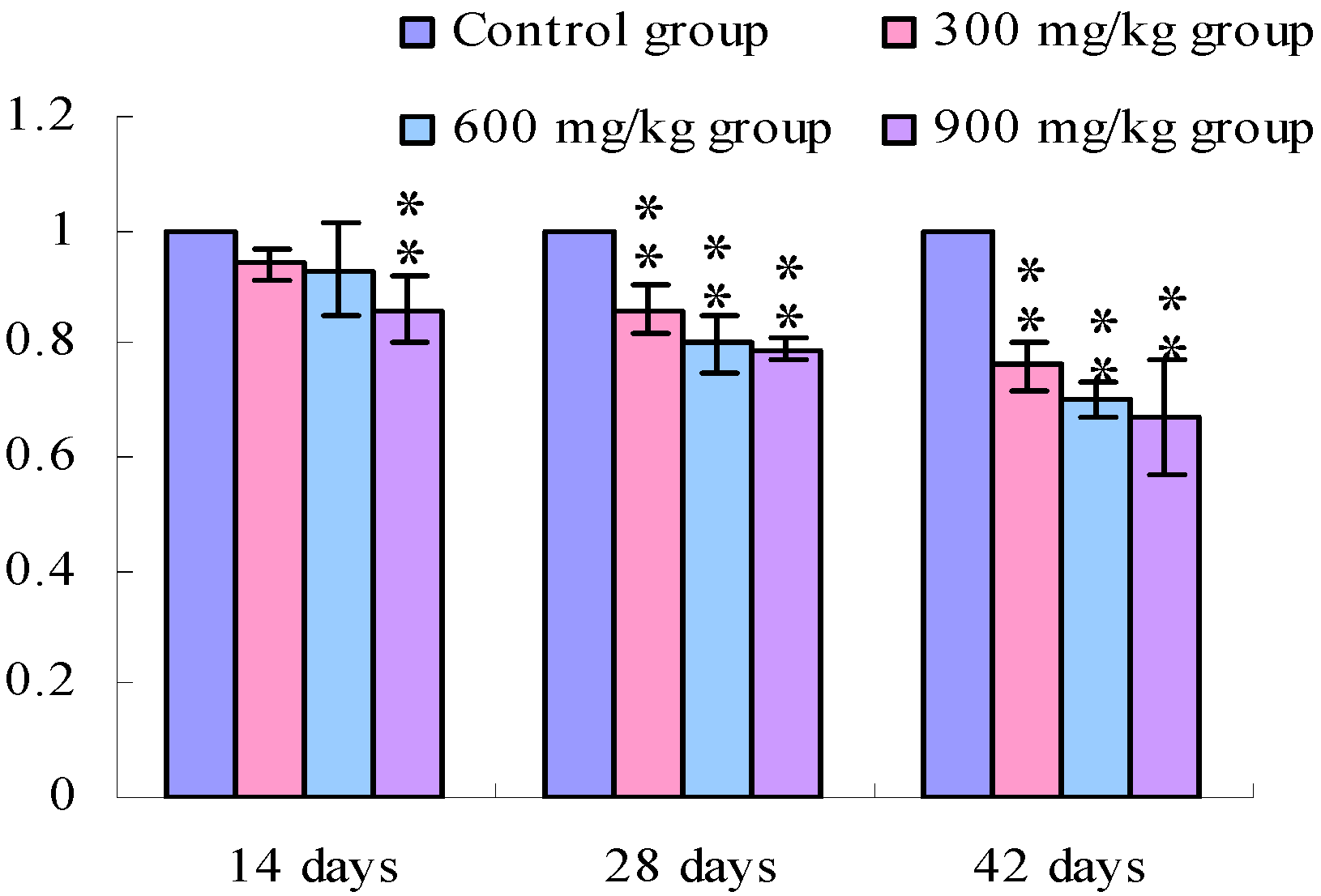
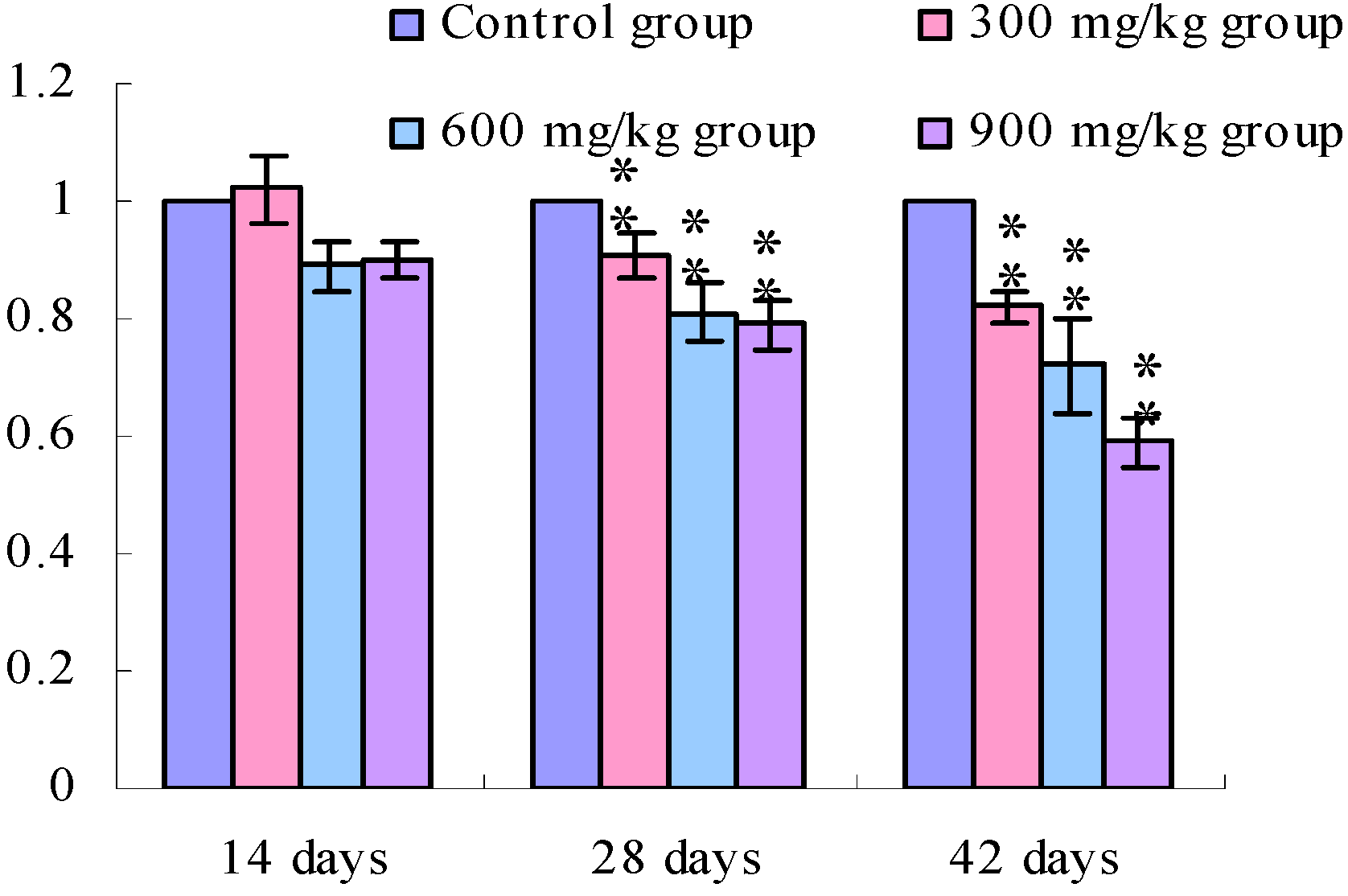
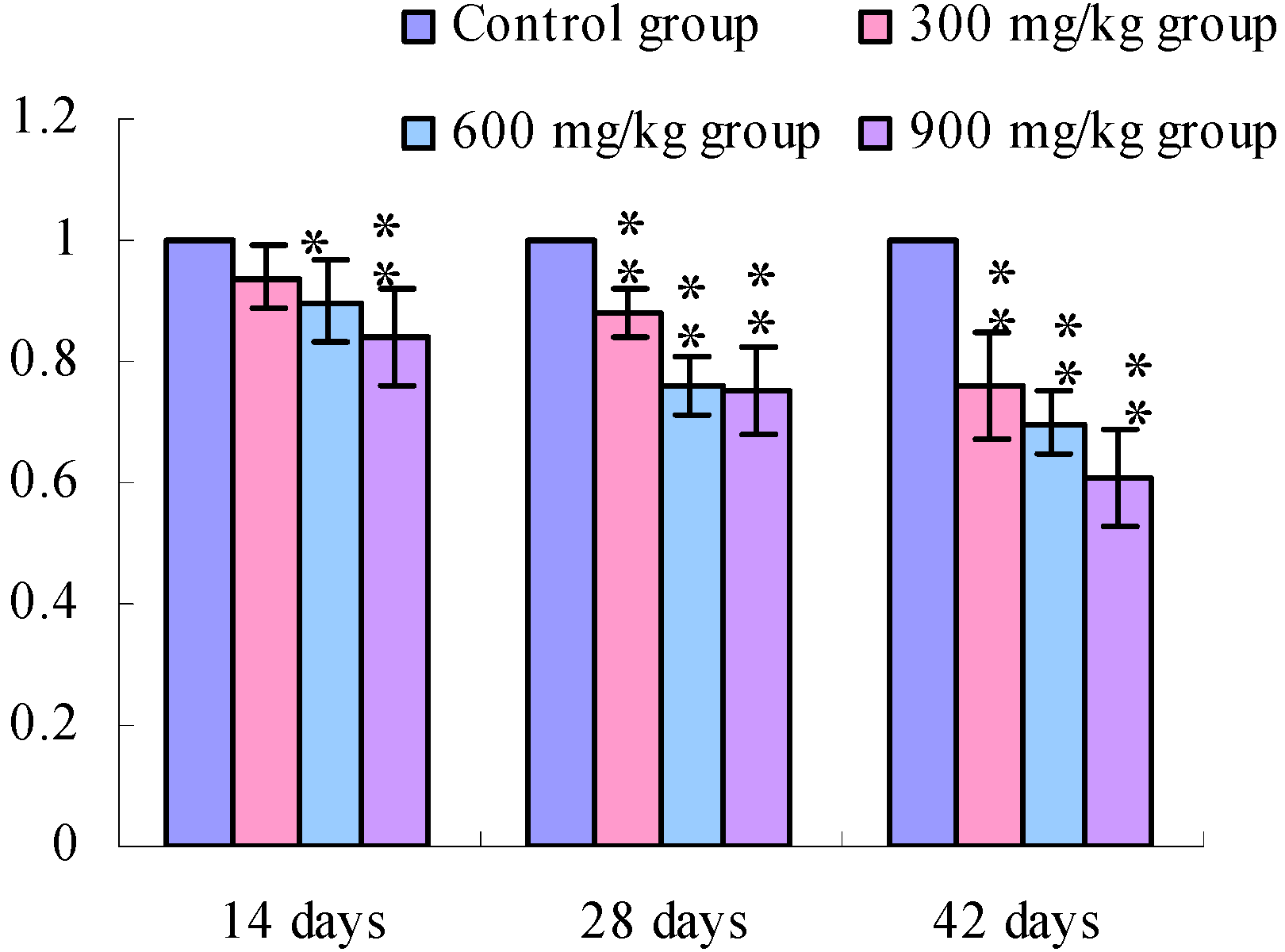
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Grandjean, P. Human exposure to Ni. IARC Sci. Publ. 1984, 53, 469–485. [Google Scholar]
- Marzec, Z. Alimentary chromium, Ni, and selenium intake of adults in Poland estimated by analysis and calculations using the duplicate portion technique. Nahrung 2004, 48, 47–52. [Google Scholar] [CrossRef]
- Spears, J.W. Ni is a “newer trace element” in the nutrition of domestic animals. J. Anim. Sci. 1984, 59, 823–835. [Google Scholar]
- Samal, L.; Mishra, C. Significance of Ni in livestock health and production. IJAVMS 2011, 5, 349–361. [Google Scholar] [CrossRef]
- Kasprzak, K.S.; Sunderman, F.W., Jr.; Salnikow, K. Ni carcinogenesis. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2003, 533, 67–97. [Google Scholar] [CrossRef]
- Lidén, C.; Carter, S. Ni release from coins. Contact Dermatitis 2001, 44, 160–165. [Google Scholar] [CrossRef]
- Lidén, C.; Skare, L.; Vahter, M. Release of Ni from coins and deposition onto skin from coin handling-comparing euro coins and SEK. Contact Dermatitis 2008, 59, 31–37. [Google Scholar] [CrossRef]
- Shen, H.M.; Zhang, Q.F. Risk assessment of Ni carcinogenicity and occupational lung cancer. Environ.Health Perspect. 1994, 102, 275–282. [Google Scholar] [CrossRef]
- Costa, M.; Zhuang, Z.X.; Huang, X.; Cosentino, S.; Klein, C.B.; Salnikow, K. Molecular mechanisms of Ni carcinogenesis. Sci. Total Environ. 1994, 148, 191–199. [Google Scholar] [CrossRef]
- Oller, A.R.; Costa, M.; Oberdorster, G. Carcinogenicity assessment of selected Ni compounds. Toxicol. Appl. Pharm. 1997, 143, 152–166. [Google Scholar] [CrossRef]
- Schaumloffel, D. Ni species: Analysis and toxic effects. J. Trace Elem. Med. Biol. 2012, 26, 1–6. [Google Scholar] [CrossRef]
- Anke, M.; Groppel, B.; Krause, U.; Langer, M. Further Data on the Biological Essentiality of Nickel. In Trace Elements in Man and Animals 6; Hurley, L.S., Keen, C.L., Lonnerdal, B., Rucker, R.B., Eds.; Plenum: New York, NY, USA, 1988. [Google Scholar]
- Ling, J.R.; Leach, M., Jr. Studies on nickel metabolism: Interaction with other elements. Poult. Sci. 1979, 58, 591–596. [Google Scholar] [CrossRef]
- Schmidt, M.; Raghavan, B.; Müller, V.; Vogl, T.; Fejer, G.; Tchaptchet, S.; Keck, S.; Kalis, C.; Nielsen, P.J.; Galanos, C.; et al. Crucial role for human Toll-like receptor 4 in the development of contact allergy to nickel. Nat. Immunol. 2010, 11, 814–819. [Google Scholar] [CrossRef]
- Kopp, E.B.; Medzhitov, R. The toll-receptor family and control of innate immunity. Curr. Opin. Immunol. 1999, 11, 13–18. [Google Scholar] [CrossRef]
- Zarember, K.A.; Godowski, P.J. Tissue expression of human toll-like receptors and differential regulation of toll-like receptor mRNAs in leukocytes in response to microbes, their products, and cytokine. J. Immunol. 2002, 168, 554–561. [Google Scholar]
- Humberd, J.; Boyd, K.; Webster, R.G. Emergence of influenza A virus variants after prolonged shedding from pheasants. J. Virol. 2007, 81, 4044–4051. [Google Scholar] [CrossRef]
- Chung, C.S.; Song, G.Y.; Moldawer, L.L.; Chaudry, I.H.; Ayala, A. Neither Fas ligand nor endotoxin is responsible for inducible peritoneal phagocyte apoptosis during sepsis/peritonitis. J. Surg. Res. 2000, 91, 147–153. [Google Scholar] [CrossRef]
- Medvedev, A.E.; Kopydlowski, K.M.; Vogel, S.N. Inhibition of lipopolysaccharide-induced signal transduction in endotoxin-tolerized mouse macrophages: Dysregulation of cytokine, chemokine, and toll-like receptor 2 and 4 gene expression. J. Immunol. 2000, 164, 5564–5574. [Google Scholar]
- Sato, S.; Nomura, F.; Kawai, T.; Takeuchi, O.; Mühlradt, P.F.; Takeda, K.; Akira, S. Synergy and cross-tolerance between toll-like receptor (TLR) 2-and TLR4-mediated signaling pathways. J. Immunol. 2000, 165, 7096–7101. [Google Scholar]
- Thoma-Uszynski, S.; Kiertscher, S.M.; Ochoa, M.T.; Bouis, D.A.; Norgard, M.V.; Miyake, K.; Godowski, P.J.; Roth, M.D.; Modlin, R.L. Activation of toll-like receptor 2 on human dendritic cells triggers induction of IL-12, but not IL-10. J. Immunol. 2000, 165, 3804–3810. [Google Scholar]
- Wang, Q.; Dziarski, R.; Kirschning, C.J.; Muzio, M.; Gupta, D. Micrococci and peptidoglycan activate TLR2→MyD88→IRAK→TRAF→NIK→IKK→NF-κB signal transduction pathway that induces transcription of interleukin-8. Infect. Immun. 2001, 69, 2270–2276. [Google Scholar] [CrossRef]
- Anderson, K.V.; Jurgens, G.; Nusslein-Volhard, C. Establishment of dorsal-ventral polarity in the Drosophila embryo: Genetic studies on the role of the Toll gene product. Cell 1985, 42, 779–789. [Google Scholar] [CrossRef]
- Halfon, M.S.; Hashimoto, C.; Keshishian, H. The Drosophila toll gene functions zygotically and is necessary for proper motoneuron and muscle development. Dev. Biol. 1995, 169, 151–167. [Google Scholar] [CrossRef]
- Keith, F.J.; Gay, N.J. The Drosophila membrane receptor Toll can function to promote cellular adhesion. EMBO J. 1990, 9, 4299–4306. [Google Scholar]
- Hajjar, A.M.; O’Mahony, D.S.; Ozinsky, A.; Underhill, D.M.; Aderem, A.; Klebanoff, S.J.; Wilson, C.B. Cutting edge: Functional interactions between Toll-like receptor (TLR) 2 and TLR1 or TLR6 in response to phenolsoluble modulin. J. Immunol. 2001, 166, 15–19. [Google Scholar]
- Ozinsky, A.; Underhill, D.M.; Fontenot, J.D.; Hajjar, A.M.; Smith, K.D.; Wilson, C.B.; Schroeder, L.; Aderem, A. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between Toll-like receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 13766–13771. [Google Scholar] [CrossRef]
- Ulevitch, R.J. Immunology: Toll gates for pathogen selection. Nature 1999, 401, 755–756. [Google Scholar] [CrossRef]
- Akter, S.; Khan, M.Z.I.; Jahan, M.R.; Karim, M.R.; Islam, M.R. Histomorphological study of the lymphoid tissues of broiler chickens. Bangladesh J. Vet. Med. 2006, 4, 87–92. [Google Scholar]
- Jeurissen, S.H.M.; Veldman, B. The Interactions between Feed (Components) and Eimeria Infections in Poultry Health. In Nutrion and Health of the Gastrointestinal Tract; Blok, M.C., Vahl, H.A., de Lange, L., van de Braak, A.E., Hemke, G., Hessing, M., Eds.; Wageningen Academic Publishers: Haarlem, Netherlands, 2002; pp. 159–182. [Google Scholar]
- Rietschel, E.T.; Brade, H. Bacterial endotoxins. Sci. Amer. 1992, 267, 54–61. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janaway, C.A. Innate immunity: The virtues of nonclonal system of recognition. Cell 1997, 91, 295–298. [Google Scholar] [CrossRef]
- Cario, E.; Podolsky, D.K. Differential alteration in intestinal epithelial cell expression of toll-like receptor 3 (TLR3) and TLR4 in inflammatory bowel disease. Infect. Immun. 2000, 68, 7010–7017. [Google Scholar] [CrossRef]
- Mayer, L.; Eisenhardt, D.; Salomon, P.; Bauer, W.; Plous, R.; Piccini, I. Expression of class II molecules on intestinal epithelial cells in humans: Differences between normal and inflammatory bowel disease. Gastroenterology 1991, 100, 3–12. [Google Scholar]
- Hecht, G.A. Microbial Pathogenesis and the Intestinal Epithelial Cell; ASM American Society for Microbiology: Washington, DC, USA, 2003; pp. 61–72. [Google Scholar]
- Cario, E.; Rosenberg, I.M.; Brandwein, S.L.; Beck, P.L.; Reinecker, H.C.; Podolsky, D.K. Lipopolysaccharide activates distinct signaling pathways in intestinal epithelial cell lines expressing Toll-like receptors. J. Immunol. 2000, 164, 966–972. [Google Scholar]
- Kostial, K.; Kargačin, B.; Landeka, M. Gut retention of metals in rats. Biol. Trace Elem. Res. 1989, 21, 213–218. [Google Scholar] [CrossRef]
- Denkhaus, E.; Salnikow, K. Ni essentiality, toxicity, and carcinogenicit. Crit. Rev. Oncol. Hematol. 2002, 42, 35–56. [Google Scholar] [CrossRef]
- Wu, B.Y.; Cui, H.M.; Peng, X.; Fang, J.; Zuo, Z.C.; Deng, J.L.; Huang, J.Y. Dietary Ni chloride induces oxidative intestinal damage in broilers. Int. J. Environ. Res. Public Health 2013, 10, 2109–2119. [Google Scholar] [CrossRef]
- Wu, B.Y.; Cui, H.M.; Peng, X.; Fang, J.; Zuo, Z.C.; Deng, J.L.; Huang, J.Y. Dietary nickel chloride restrains the development of small intestine in broilers. Biol. Trace Elem. Res. 2013, 155, 236–246. [Google Scholar] [CrossRef]
- Wu, B.Y.; Cui, H.M.; Peng, X.; Fang, J.; Zuo, Z.C.; Deng, J.L.; Huang, J.Y. Changes of the serum cytokine contents in broilers fed on diets supplemented with nickel chloride. Biol. Trace Elem. Res. 2013, 151, 234–239. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Iqbal, M.; Philbin, V.J.; Smith, A.L. Expression patterns of the Toll-like receptor mRNA in tissues, immune cell subsets and cell lines. Vet. Immunol. Immunopathol. 2005, 104, 117–127. [Google Scholar] [CrossRef]
- Schmidt, M.; Goebeler, M. Ni allergies: Paying the toll for innate immunity. J. Mol. Med. 2011, 89, 961–970. [Google Scholar] [CrossRef]
- Roediger, B.; Weninger, W. How Ni turns on innate immune cells. Immunol. Cell Biol. 2010, 89, 1–2. [Google Scholar] [CrossRef]
- Campbell, K.; Perkins, N. Regulation of NF-κB function. Biochem. Soc. Symp. 2006, 73, 165–180. [Google Scholar]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Boyd, Y.; Goodchild, M.; Morroll, S.; Bumstead, N. Mapping of the chicken and mouse genes for toll-like receptor 2 (TLR2) to an evolutionarily conserved chromosomal segment. Immunogenetics 2001, 52, 294–298. [Google Scholar] [CrossRef]
- Fukui, A.; Inoue, N.; Matsumoto, M.; Nomura, M.; Yamada, K.; Matsuda, Y.; Toyoshima, K.; Seya, T. Molecular cloning and functional characterization of chicken toll-like receptors. A single chicken toll covers multiple molecular patterns. J. Biol. Chem. 2001, 276, 47143–47149. [Google Scholar]
- Leveque, G.; Forgetta, V.; Morroll, S.; Smith, A.L.; Bumstead, N.; Barrow, P.; Loredo-Osti, J.C.; Morgan, K.; Malo, D. Allelic variation in TLR4 is linked to susceptibility to Salmonella enterica serovar Typhimurium infection in chickens. Infect. Immun. 2003, 71, 1116–1124. [Google Scholar] [CrossRef]
- Cario, E.; Gerken, G.; Podolsky, D.K. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 2007, 132, 1359–1374. [Google Scholar] [CrossRef]
- Hausmann, M.; Kiessling, S.; Mestermann, S.; Webb, G.; Spöttl, T.; Andus, T.; Schölmerich, J.; Herfarth, H.; Ray, K.; Falk, W.; et al. Toll-like receptors 2 and 4 are up-regulated during intestinal inflammation. Gastroenterology 2002, 122, 1987–2000. [Google Scholar] [CrossRef]
- Brant, K.A.; Fabisiak, J.P. Ni Alterations of TLR2-dependent chemokine profiles in lung fibroblasts are mediated by COX-2. Am. J. Respir. Cell Mol. Biol. 2008, 38, 591–599. [Google Scholar] [CrossRef]
- Aliprants, A.O.; Yang, R.B.; Weiss, D.S.; Godowski, P.; Zychlinsky, A. The apoptotic signaling pathway activated by Toll-like receptor-2. EMBO J. 2000, 19, 3325–3336. [Google Scholar] [CrossRef]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol. 1999, 162, 3749–3752. [Google Scholar]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in the Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef]
- Chow, J.C.; Young, D.W.; Golenbock, D.T.; Christ, W.J.; Gusovsky, F. Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J. Biol. Chem. 1999, 274, 10689–10692. [Google Scholar]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Muzio, M.; Natoli, G.; Saccani, S.; Levrero, M.; Mantovani, A. The human Toll signaling pathway: Divergence of nuclear factor kB and JNK/SAPK activation upstream of tumor necrosis factor receptor-associated factor 6 (TRAF6). J. Exp. Med. 1998, 187, 2097–2101. [Google Scholar] [CrossRef]
- Zhang, F.X.; Kirscning, C.; Mancinelli, R.; Jin, Y.; Mantovani, A.; Faure, E.; Rothe, M.; Muzio, M.; Arditi, M. Bacterial lipopolysaccharide activates nuclear factor-kB through interleukin-1 signaling mediators in cultured human dermal endothelial cells and human mononuclear phagocytes. J. Biol. Chem. 1999, 274, 7611–7614. [Google Scholar] [CrossRef]
- Rothenberg, M.E. Innate sensing of Ni. Nat. Immunol. 2010, 11, 781–782. [Google Scholar] [CrossRef]
- Medzhitov, R.; Preston-Hurlburt, P.; Kopp, E.; Stadlen, A.; Chen, C.; Ghosh, S.; Janeway, C.A., Jr. MyD88 is an adaptor protein in the hToll/IL-1 receptor family signaling pathways. Mol. Cell 1998, 2, 253–258. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janeway, C. Innate immunity. N. Engl. J. Med. 2000, 343, 338–344. [Google Scholar] [CrossRef]
- Anderson, K.V. Toll signaling pathways in the innate immune response. Curr. Opin. Immunol. 2000, 12, 13–19. [Google Scholar] [CrossRef]
- Wu, B.Y.; Cui, H.M.; Peng, X.; Fang, J.; Zuo, Z.C.; Deng, J.L.; Huang, J.Y. Dietary nickel chloride induces oxidative stress, apoptosis and alters Bax/Bcl-2 and caspase-3 mRNA expression in the cecal tonsil of broilers. Food Chem. Toxicol. 2014, 63, 18–29. [Google Scholar] [CrossRef]
- Fas, S.C.; Fritzsching, B.; Suri-Payer, E.; Krammer, P.H. Death receptor signaling and its function in the immune system. Curr. Dir. Autoimmun. 2006, 9, 1–17. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, B.; Cui, H.; Peng, X.; Fang, J.; Zuo, Z.; Deng, J.; Huang, J. Analysis of the Toll-Like Receptor 2-2 (TLR2-2) and TLR4 mRNA Expression in the Intestinal Mucosal Immunity of Broilers Fed on Diets Supplemented with Nickel Chloride. Int. J. Environ. Res. Public Health 2014, 11, 657-670. https://doi.org/10.3390/ijerph110100657
Wu B, Cui H, Peng X, Fang J, Zuo Z, Deng J, Huang J. Analysis of the Toll-Like Receptor 2-2 (TLR2-2) and TLR4 mRNA Expression in the Intestinal Mucosal Immunity of Broilers Fed on Diets Supplemented with Nickel Chloride. International Journal of Environmental Research and Public Health. 2014; 11(1):657-670. https://doi.org/10.3390/ijerph110100657
Chicago/Turabian StyleWu, Bangyuan, Hengmin Cui, Xi Peng, Jing Fang, Zhicai Zuo, Junliang Deng, and Jianying Huang. 2014. "Analysis of the Toll-Like Receptor 2-2 (TLR2-2) and TLR4 mRNA Expression in the Intestinal Mucosal Immunity of Broilers Fed on Diets Supplemented with Nickel Chloride" International Journal of Environmental Research and Public Health 11, no. 1: 657-670. https://doi.org/10.3390/ijerph110100657