A Tale of Two City Blocks: Differences in Immature and Adult Mosquito Abundances between Socioeconomically Different Urban Blocks in Baltimore (Maryland, USA)


Abstract
:1. Introduction

2. Experimental Section


{kind=link}
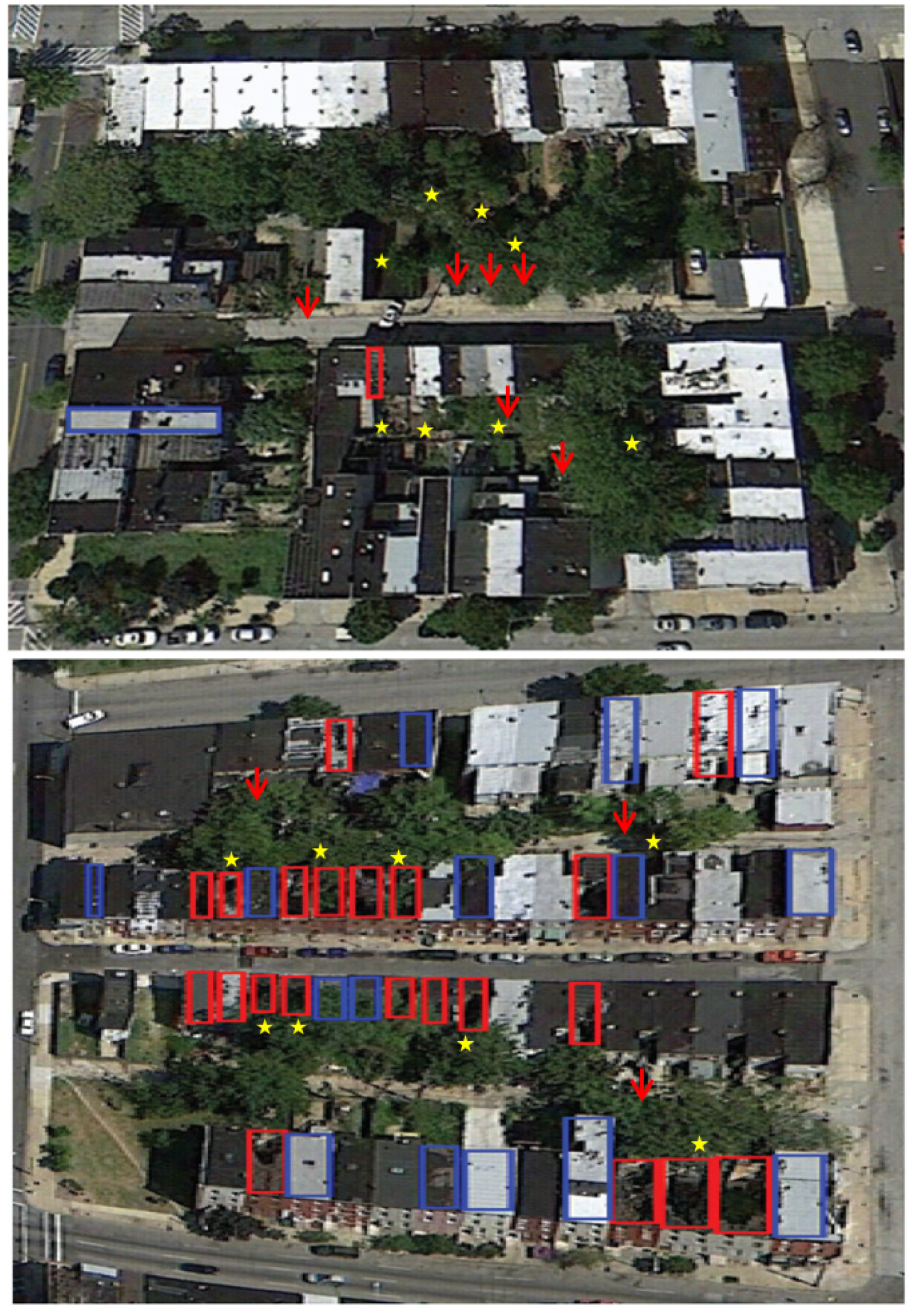{kind=link}
| Block Characteristic | Low-decay | High-decay |
|---|---|---|
| Area (hectares) | 1.31 | 1.28 |
| Parcels (includes empty lots) | 58 | 77 |
| Total Buildings | 54 | 70 |
| Vacant Buildings | 2 | 36 |
| Vacant Buildings with missing roof | 1 | 21 |
2.1. Adult Mosquito Samples
2.2. Immature Mosquito Samples
3. Results and Discussion
3.1. Adult Sampling
3.2. Immature Sampling
| Block Metric | Low-decay | High-decay |
|---|---|---|
| Parcels surveyed (#) | 17.24% (10) | 58.33% (42) |
| Parcels with accessible water , % surveyed | 100.00% | 16.67% |
| Parcels with mosquito larvae, % surveyed | 60.00% | 7.14% |
| Containers per parcel, mean (se) | 2.57 (0.26) | 0.26 (0.08) |
| Containers with larvae, % | 50.00% | 22.22% |
| Containers with pupae, % | 27.78% | 22.22% |
| Mean larvae per surveyed parcel, Ae. albopictus (se) | 14.78 (6.36) | 2.69 (1.92) |
| Mean larvae per surveyed parcel, Cx. pipiens (se) | 26.22 (14.12) | 8.57 (6.30) |
| Mean adult per trap night, Ae. albopictus (se) | 70.57 (13.45) | 11.00 (3.40) |
| Mean adult per trap night, Cx. pipiens (se) | 3.29 (1.32) | 0.63 (0.50) |
3.3. Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brown, D.G.; Johnson, K.M.; Loveland, T.R.; Theobald, D.M. Rural land-use trends in the conterminous United States, 1950–2000. Ecol. Appl. 2005, 15, 1851–1863. [Google Scholar] [CrossRef]
- Glaeser, E.L.; Gyourko, J. Urban decline and durable housing. J. Polit. Econ. 2005, 113, 345–375. [Google Scholar] [CrossRef]
- Grossmann, K.; Bontje, M.; Haase, A. Shrinking cities: Notes for the further research agenda. Cities 2013, 35, 221–225. [Google Scholar] [CrossRef]
- Wilson, W.J. When Work Disappears: The World of the New Urban Poor; Vintage: New York, NY, USA, 1996. [Google Scholar]
- Beauregard, R.A. Federal policy and postwar urban decline: A case of government complicity? Hous. Policy Debate 2001, 12, 129–151. [Google Scholar] [CrossRef]
- Semyonov, M.; Haberfeld, Y.; Cohen, Y.; Lewin-Epstein, N. Racial composition and occupational segregation and inequality across American cities. Soc. Sci. Res. 2000, 29, 175–187. [Google Scholar] [CrossRef]
- Cohen, P.N. Race, class and labor markets: The White working class and racial composition of US metropolitan areas. Soc. Sci. Res. 2001, 30, 146–169. [Google Scholar] [CrossRef]
- Cohen, J.E. Human population grows up. Sci. Am. 2005, 293, 48–55. [Google Scholar] [CrossRef]
- Haase, D. Urban ecology of shrinking cities: An unrecognized opportunity? Nat. Cult. 2008, 3, 1–8. [Google Scholar]
- Pickett, S.T.A.; Boone, C.G.; McGrath, B.P.; Cadenasso, M.L.; Childers, D.L.; Ogden, L.A.; McHale, M.; Grove, J.M. Ecological science and transformation to the sustainable city. Cities 2013, 32, S10–S20. [Google Scholar] [CrossRef]
- Kelling, G.L.; Coles, C.M. Fixing Broken Windows: Restoring Order and Reducing Crime in Our Communities; Manhattan Institute for Policy Research Inc.: New York, NY, USA, 1996. [Google Scholar]
- Kraut, D.T. Hanging out the no vacancy sign: Eliminating the blight of vacant buildings from urban areas. N. Y. Univ. Law Rev. 1999, 74, 1139–1177. [Google Scholar]
- Skogan, W.G. The national crime survey redesign. Public Opin. Q. 1990, 54, 256–272. [Google Scholar] [CrossRef]
- Reisen, W.K.; Takahashi, R.M.; Carroll, B.D.; Quiring, R. Delinquent mortgages, neglected swimming pools, and West Nile virus, California. Emerg. Infect. Dis. 2008, 14, 1747–1749. [Google Scholar] [CrossRef]
- Frazier, A.E.; Bagchi-Sen, S.; Knight, J. The spatio-temporal impacts of demolition land use policy and crime in a shrinking city. Appl. Geogr. 2013, 41, 55–64. [Google Scholar] [CrossRef]
- LaDeau, S.L.; Leisnham, P.T.; Biehler, D.; Bodner, D. Higher mosquito production in low-income neighborhoods of Baltimore and Washington, DC: Understanding ecological drivers and mosquito-borne disease risk in temperate cities. Int. J. Environ. Res. Public Health 2013, 10, 1505–1526. [Google Scholar] [CrossRef]
- Leisnham, P.T.; Juliano, S.A. Impacts of climate, land use, and biological invasion on the ecology of immature Aedes mosquitoes: Implications for La Crosse emergence. Ecohealth 2012, 9, 217–228. [Google Scholar] [CrossRef]
- Leisnham, P.T.; Slaney, D.P. Urbanization and the Increasing Risk from Mosquito-Borne Diseases: Linking Human Well-Being with Ecosystem Health. In Focus on Urbanization Trends; De Smet, L.M., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2009; pp. 47–82. [Google Scholar]
- LaDeau, S.L.; Calder, C.A.; Doran, P.J.; Marra, P.P. West Nile virus impacts in American crow populations are associated with human land use and climate. Ecol. Res. 2011, 26, 909–916. [Google Scholar] [CrossRef]
- Alemu, A.; Tsegaye, W.; Golassa, L.; Abebe, G. Urban malaria and associated risk factors in Jimma town, south-west Ethiopia. Malar. J. 2011, 10. [Google Scholar] [CrossRef]
- Delatte, H.; Bagny, L.; Brengue, C.; Bouetard, A.; Paupy, C.; Fontenille, D. The invaders: Phylogeography of dengue and chikungunya viruses Aedes vectors, on the South West islands of the Indian Ocean. Infec. Genet. Evol. 2011, 11, 1769–1781. [Google Scholar] [CrossRef]
- Patz, J.A.; Graczyk, T.K.; Geller, N.; Vittor, A.Y. Effects of environmental change on emerging parasitic diseases. Int. J. Parasitol. 2000, 30, 1395–1405. [Google Scholar] [CrossRef]
- Keesing, F.; Brunner, J.; Duerr, S.; Killilea, M.; LoGiudice, K.; Schmidt, K.; Vuong, H.; Ostfeld, S.R. Hosts as ecological traps for the vector of Lyme disease. P. Roy. Soc. BBio. Sci. 2009, 276, 3911–3919. [Google Scholar] [CrossRef]
- Allan, B.F.; Langerhans, R.B.; Ryberg, W.A.; Landesman, W.J.; Griffin, N.W.; Katz, R.S.; Oberle, B.J.; Schutzenhofer, M.R.; Smyth, K.N.; de St Maurice, A.; et al. Ecological correlates of risk and incidence of West Nile virus in the United States. Oecologia 2009, 158, 699–708. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Daszak, P.; Jones, M.J.; Marra, P.P.; Kramer, L.D. Host heterogeneity dominates West Nile virus transmission. P. Roy. Soc. B Bio. Sci. 2006, 273, 2327–2333. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Kramer, L.D.; Jones, M.J.; Marra, P.P.; Daszak, P. West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior. PloS Biol. 2006, 4, 606–610. [Google Scholar]
- Juliano, S.A.; Lounibos, L.P. Ecology of invasive mosquitoes: Effects on resident species and on human health. Ecol. Lett. 2005, 8, 558–574. [Google Scholar] [CrossRef]
- Gratz, N.G. Emerging and resurging vector-borne diseases. Annu. Rev. Entomol. 1999, 44, 51–75. [Google Scholar] [CrossRef]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef]
- Dowling, Z.; Ladeau, S.L.; Armbruster, P.; Biehler, D.; Leisnham, P.T. Socioeconomic status affects mosquito (Diptera: Culicidae) larval habitat type availability and infestation level. J. Med. Entomol. 2013, 50, 764–772. [Google Scholar] [CrossRef]
- Rochlin, I.; Ninivaggi, D.V.; Hutchinson, M.L.; Farajollahi, A. Climate change and range expansion of the Asian tiger mosquito (Aedes albopictus) in Northeastern USA: Implications for public health practitioners. PloS One 2013, 8. [Google Scholar] [CrossRef]
- Moore, C.G.; Mitchell, C.J. Aedes albopictus in the United States: Ten-year presence and public health implications. Emerg. Infect. Dis. 1997, 3, 329–334. [Google Scholar] [CrossRef]
- Bartlett-Healy, K.; Unlu, I.; Obenauer, P.; Hughes, T.; Healy, S.; Crepeau, T.; Farajollahi, A.; Kesavaraju, B.; Fonseca, D.; Schoeler, G.; et al. Larval mosquito habitat utilization and community dynamics of Aedes albopictus and Aedes japonicus (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 813–824. [Google Scholar] [CrossRef]
- Leisnham, P.T.; Juliano, S.A. Spatial and temporal patterns of coexistence between competing Aedes mosquitoes in urban Florida. Oecologia 2009, 160, 343–352. [Google Scholar] [CrossRef]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; Oguinn, M.L.; Andreadis, T.G.; Blow, J.A. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef]
- Gerhardt, R.R.; Gottfried, K.L.; Apperson, C.S.; Davis, B.S.; Erwin, P.C.; Smith, A.B.; Panella, N.A.; Powell, E.E.; Nasci, R.S. First isolation of La Crosse virus from naturally infected Aedes albopictus. Emerg. Infect. Dis. 2001, 7, 807–811. [Google Scholar] [CrossRef]
- Ibanez-Bernal, S.; Briseno, B.; Mutebi, J.P.; Argot, E.; Rodriguez, G.; Martinez-Campos, C.; Paz, R.; de la Fuente-San Roman, P.; Tapia-Conyer, R.; Flisser, A. First record in America of Aedes albopictus naturally infected with dengue virus during the 1995 outbreak at Reynosa, Mexico. Med. Vet. Entomol. 1997, 11, 305–309. [Google Scholar] [CrossRef]
- Darsie, R.; Ward, R. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico; University of Florida Press: Gainesville, FL, USA, 2005. [Google Scholar]
- Sardelis, M.R.; Turell, M.J.; Dohm, D.J.; O’Guinn, M.L. Vector competence of selected North American Culex and Coquillettidia mosquitoes for West Nile virus. Emerg. Infect. Dis. 2001, 7, 1018–1022. [Google Scholar] [CrossRef]
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Marm Kilpatrick, A. “Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef]
- Fonseca, D.M.; Keyghobadi, N.; Malcolm, C.A.; Mehmet, C.; Schaffner, F.; Mogi, M.; Fleischer, R.C.; Wilkerson, R.C. Emerging vectors in the Culex pipiens complex. Science 2004, 303, 1535–1538. [Google Scholar] [CrossRef]
- Diuk-Wasser, M.A.; Brown, H.E.; Andreadis, T.G.; Fish, D. Modeling the spatial distribution of mosquito vectors for West Nile virus in Connecticut, USA. Vector Borne Zoonotic Dis. 2006, 6, 283–295. [Google Scholar]
- Brown, H.; Diuk-Wasser, M.; Andreadis, T.; Fish, D. Remotely-sensed vegetation indices identify mosquito clusters of West Nile virus vectors in an urban landscape in the northeastern United States. Vector Borne Zoonotic Dis. 2008, 8, 197–206. [Google Scholar] [CrossRef]
- Juliano, S.A. Population dynamics. J. Am. Mosq. Control Assoc. 2007, 23, 265–275. [Google Scholar] [CrossRef]
- Juliano, S.A. Species Interactions Among Larval Mosquitoes: Context Dependence Across Habitat Gradients. Annu. Rev. Entomol. 2009, 54, 37–56. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Nishimura, N.; Escher, R.L. Fitness of a treehole mosquito—Influences of food type and predation. Oikos 1993, 66, 114–118. [Google Scholar] [CrossRef]
- Su, T.Y.; Mulla, M.S. Effects of temperature on development, mortality, mating and blood feeding behavior of Culiseta incidens (Diptera: Culicidae). J. Vector Ecol. 2001, 26, 83–92. [Google Scholar]
- Blaustein, L.; Kiflawi, M.; Eitam, A.; Mangel, M.; Cohen, J.E. Oviposition habitat selection in response to risk of predation in temporary pools: Mode of detection and consistency across experimental venue. Oecologia 2004, 138, 300–305. [Google Scholar] [CrossRef]
- Stav, G.; Blaustein, L.; Margalit, Y. Individual and interactive effects of a predator and controphic species on mosquito populations. Ecol. Appl. 2005, 15, 587–598. [Google Scholar] [CrossRef]
- Yee, D.A.; Juliano, S.A. Consequences of detritus type in an aquatic microsystem: Effects on water quality, micro-organisms and performance of the dominant consumers. Freshw. Biol. 2006, 51, 448–459. [Google Scholar] [CrossRef]
- Blaustein, L.; Chase, J.M. Interactions between mosquito larvae and species that share the same trophic level. Annu. Rev. Entomol. 2007, 52, 489–507. [Google Scholar] [CrossRef]
- Delatte, H.; Gimonneau, G.; Triboire, A.; Fontenille, D. Influence of temperature on immature development, survival, longevity, fecundity, and gonotrophic cycles of Aedes albopictus, vector of chikungunya and dengue in the Indian Ocean. J. Med. Entomol. 2009, 46, 33–41. [Google Scholar]
- Chase, J.M.; Shulman, R.S. Wetland isolation facilitates larval mosquito density through the reduction of predators. Ecol. Entomol. 2009, 34, 741–747. [Google Scholar] [CrossRef]
- Costanzo, K.S.; Kesavaraju, B.; Juliano, S.A. Condition-specific competition in container mosquitoes: The role of noncompeting life-history stages. Ecology 2005, 86, 3289–3295. [Google Scholar] [CrossRef]
- Buyantuyev, A.; Wu, J.G. Urban heat islands and landscape heterogeneity: Linking spatiotemporal variations in surface temperatures to land-cover and socioeconomic patterns. Landsc. Ecol. 2010, 25, 17–33. [Google Scholar] [CrossRef]
- Landau, K.I.; van Leeuwen, W.J.D. Fine scale spatial urban land cover factors associated with adult mosquito abundance and risk in Tucson, Arizona. J. Vector Ecol. 2012, 37, 407–418. [Google Scholar] [CrossRef]
- Richards, S.L.; Ghosh, S.K.; Zeichner, B.C.; Apperson, C.S. Impact of source reduction on the spatial distribution of larvae and pupae of Aedes albopictus (Diptera: Culicidae) in suburban neighborhoods of a Piedmont community in North Carolina. J. Med. Entomol. 2008, 45, 617–628. [Google Scholar] [CrossRef]
- Yee, D.A.; Allgood, D.; Kneitel, J.M.; Kuehn, K.A. Constitutive differences between natural and artificial container mosquito habitats: Vector communities, resources, microorganisms, and habitat parameters. J. Med. Entomol. 2012, 49, 482–491. [Google Scholar]
- Yee, D.A. Tires as habitats for mosquitoes: A review of studies within the eastern United States. J. Med. Entomol. 2008, 45, 581–593. [Google Scholar]
- Reisen, W.K.; Meyer, R.P.; Tempelis, C.H.; Spoehel, J.J. Mosquito abundance and bionomics in residential communities in Orange and Los Angeles counties, California. J. Med. Entomol. 1990, 27, 356–367. [Google Scholar]
- Reisen, W.K.; Carroll, B.D.; Takahashi, R.; Fang, Y.; Garcia, S.; Martinez, V.M.; Quiring, R. Repeated West Nile virus epidemic transmission in Kern County, California, 2004–2007. J. Med. Entomol. 2009, 46, 139–157. [Google Scholar] [CrossRef]
- Harrigan, R.J.; Thomassen, H.A.; Buermann, W.; Cummings, R.F.; Kahn, M.E.; Smith, T.B. Economic conditions predict prevalence of West Nile virus. PloS One 2010, 5. [Google Scholar] [CrossRef]
- Joshi, V.; Sharma, R.C.; Sharma, Y.; Adha, S.; Sharma, K.; Singh, H.; Purohit, A.; Singhi, M. Importance of socioeconomic status and tree holes in distribution of Aedes mosquitoes (Diptera: Culicidae) in Jodhpur, Rajasthan, India. J. Med. Entomol. 2006, 43, 330–336. [Google Scholar] [CrossRef]
- Unlu, I.; Farajollahi, A.; Healy, S.P.; Crepeau, T.; Bartlett-Healy, K.; Williges, E.; Strickman, D.; Clark, G.G.; Gaugler, R.; Fonseca, D.M. Area-wide management of Aedes albopictus: Choice of study sites based on geospatial characteristics, socioeconomic factors and mosquito populations. Pest Manag. Sci. 2011, 67, 965–974. [Google Scholar] [CrossRef]
- Bradman, A.; Chevrier, J.; Tager, I.; Lipsett, M.; Sedgwick, J.; Macher, J.; Vargas, A.B.; Cabrera, E.B.; Camacho, J.M.; Weldon, R.; et al. Association of housing disrepair indicators with cockroach and rodent infestations in a cohort of pregnant Latina women and their children. Environ. Health Perspect. 2005, 113, 1795–1801. [Google Scholar] [CrossRef]
- Rauh, V.A.; Chew, G.L.; Garfinkel, R.S. Deteriorated housing contributes to high cockroach allergen levels in inner-city households. Environ. Health Perspect. 2002, 110, 323–327. [Google Scholar] [CrossRef]
- Baltimore City: Vacant Building Index. Available online: http://cityview.baltimorecity.gov/vacantbuildings/ (accessed on 7 February 2012).
- Baltimore City: Open Data Catalog. Available online: http://cityview.baltimorecity.gov/ (accessed on 7 February 2012).
- Honorio, N.A.; Silva, W.D.; Leite, P.J.; Goncalves, J.M.; Lounibos, L.P.; Lourenco-de-Oliveira, R. Dispersal of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in an urban endemic dengue area in the State of Rio de Janeiro, Brazil. Mem. Inst. Oswaldo Cruz. 2003, 98, 191–198. [Google Scholar]
- Cianci, D.; van den Broek, J.; Caputo, B.; Marini, F.; Della Torre, A.; Heesterbeek, H.; Hartemink, N. Estimating mosquito population size from mark-release-recapture data. J. Med. Entomol. 2013, 50, 533–542. [Google Scholar] [CrossRef]
- Marini, F.; Caputo, B.; Pombi, M.; tarsitani, G.; della Torre, A. Study of Aedes albopictus dispersal in Rome, Italy, using sticky traps in mark-release-recapture experiments. Med. Vet. Entomol. 2010, 24, 361–368. [Google Scholar] [CrossRef]
- Crepeau, T.N.; Healy, S.P.; Bartlett-Healy, K.; Unlu, I.; Farajollahi, A.; Fonseca, D.M. Effects of biogents sentinel trap field placement on capture rates of adult Asian tiger mosquitoes, Aedes albopictus. PloS One 2013, 8. [Google Scholar] [CrossRef]
- Leisnham, P.T.; Lounibos, L.P.; O’Meara, G.F.; Juliano, S.A. Interpopulation divergence in competitive interactions of the mosquito Aedes albopictus. Ecology 2009, 90, 2405–2413. [Google Scholar]
- Johnson, B.J.; Sukhdeo, M.V. Successional mosquito dynamics in surrogate treehole and ground-container habitats in the northeastern United States: Where does Aedes albopictus fit in? J. Vector Ecol. 2013, 38, 168–174. [Google Scholar]
- Johnson, B.J.; Sukhdeo, M.V.K. Drought-Induced amplification of local and regional West Nile virus infection rates in New Jersey. J. Med. Entomol. 2013, 50, 195–204. [Google Scholar] [CrossRef]
- Maryland Climatological Data: Baltimore, Inner Harbor. Available online: http://w1.weather.gov/obhistory/KDMH.html (accessed on 7 February 2012).
- Silver, J.B. Mosquito Ecology: Field Sampling Methods; Springer: New York, NY, USA, 2008. [Google Scholar]
- Bartlett-Healy, K.; Healy, S.P.; Hamilton, G.C. A model to predict evaporation rates in habitats used by container-dwelling mosquitoes. J. Med. Entomol. 2011, 48, 712–716. [Google Scholar] [CrossRef]
- Renshaw, M.; Service, M.W.; Birley, M.H. Size variation and reproductive success in the mosquito Aedes Cantans. Med. Vet Entomol. 1994, 8, 179–186. [Google Scholar] [CrossRef]
- Alto, B.W.; Muturi, E.J.; Lampman, R.L. Effects of nutrition and density in Culex pipiens. Med. Vet. Entomol. 2012, 26, 396–406. [Google Scholar]
- Nasci, R.S.; Mitchell, C.J. Larval diet, adult size, and susceptibility of Aedes aegypti (Diptera: Culicidae) to infection with Ross River virus. J. Med. Entomol. 1994, 31, 123–126. [Google Scholar]
- Westbrook, C.J.; Reiskind, M.H.; Pesko, K.N.; Greene, K.E.; Lounibos, L.P. Larval environmental temperature and the susceptibility of Aedes albopictus Skuse (Diptera: Culicidae) to Chikungunya virus. Vector Borne Zoonotic Dis. 2010, 10, 241–247. [Google Scholar] [CrossRef]
- Alto, B.W.; Lounibos, L.P.; Mores, C.N.; Reiskind, M.H. Larval competition alters susceptibility of adult Aedes mosquitoes to dengue infection. P. Roy. Soc. B Biol. Sci. 2008, 275, 463–471. [Google Scholar] [CrossRef]
- Alto, B.W.; Lounibos, L.P.; Higgs, S.; Juliano, S.A. Larval competition differentially affects arbovirus infection in Aedes mosquitoes. Ecology 2005, 86, 3279–3288. [Google Scholar] [CrossRef]
- Juliano, S.A.; Stoffregen, T.L. Effects of habitat drying on size at and time to metamorphosis in the tree hole mosquito Aedes triseriatus. Oecologia 1994, 97, 369–376. [Google Scholar]
- Teng, H.J.; Apperson, C.S. Development and survival of immature Aedes albopictus and Aedes triseriatus (Diptera: Culicidae) in the laboratory: Effects of density, food, and competition on response to temperature. J. Med. Entomol. 2000, 37, 40–52. [Google Scholar] [CrossRef]
- Leisnham, P.T.; LaDeau, S.L.; Juliano, S.A. Spatial and temporal habitat segregation of mosquitoes in urban Florida. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Juliano, S.A.; O’Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef]
- Dowling, Z.; Armbruster, P.; LaDeau, S.L.; DeCotiis, M.; Mottley, J.; Leisnham, P.T. Linking mosquito infestation to resident socioeconomic status, knowledge, and source reduction practices in suburban Washington, DC. Ecohealth 2013, 10, 36–47. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Becker, B.; Leisnham, P.T.; LaDeau, S.L. A Tale of Two City Blocks: Differences in Immature and Adult Mosquito Abundances between Socioeconomically Different Urban Blocks in Baltimore (Maryland, USA). Int. J. Environ. Res. Public Health 2014, 11, 3256-3270. https://doi.org/10.3390/ijerph110303256
Becker B, Leisnham PT, LaDeau SL. A Tale of Two City Blocks: Differences in Immature and Adult Mosquito Abundances between Socioeconomically Different Urban Blocks in Baltimore (Maryland, USA). International Journal of Environmental Research and Public Health. 2014; 11(3):3256-3270. https://doi.org/10.3390/ijerph110303256
Chicago/Turabian StyleBecker, Brian, Paul T. Leisnham, and Shannon L. LaDeau. 2014. "A Tale of Two City Blocks: Differences in Immature and Adult Mosquito Abundances between Socioeconomically Different Urban Blocks in Baltimore (Maryland, USA)" International Journal of Environmental Research and Public Health 11, no. 3: 3256-3270. https://doi.org/10.3390/ijerph110303256