A Synthetic Thiourea-Based Tripodal Receptor that Impairs the Function of Human First Trimester Cytotrophoblast Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Experimental Section
2.1. General Information
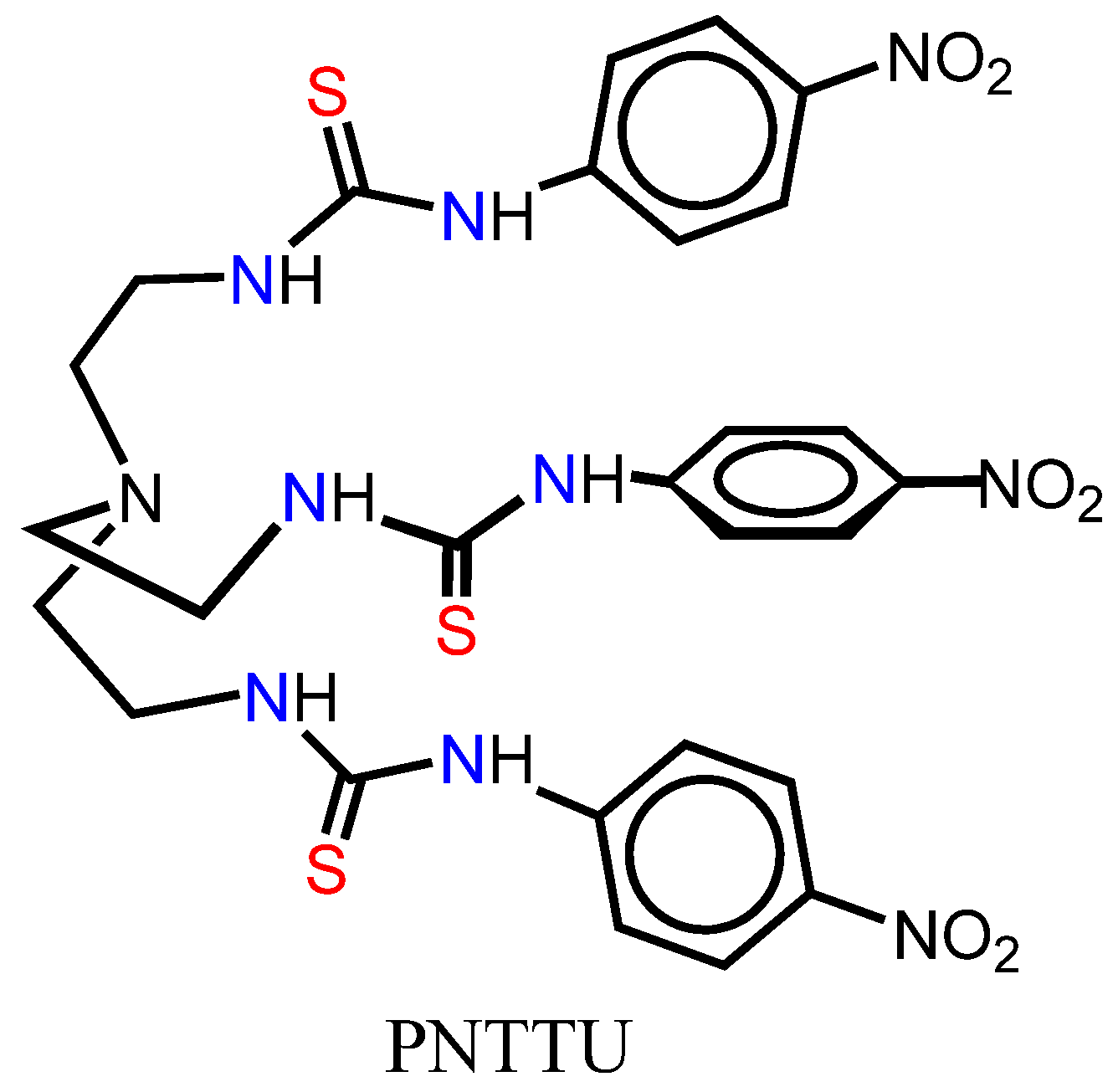
2.2. Synthesis of PNTTU
2.3. Cell Culture
2.4. Effect of PNTTU on CTB Cells Function
2.5. ELISA for Angiogenic and Anti-angiogenic Factors
2.6. Western Blots for VEGFR-1, AT1, and AT2 Receptors
2.7. Statistical Methods
3. Results and Discussion
3.1. Results
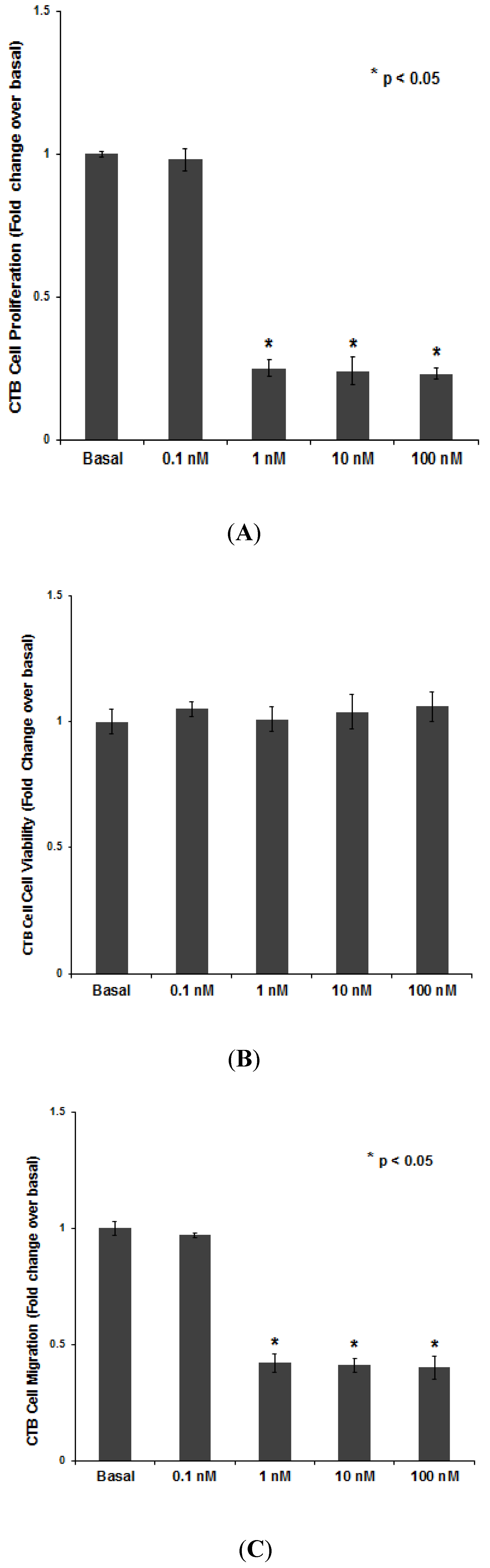
3.1.1. PNTTU Inhibited the CTB Cells Function

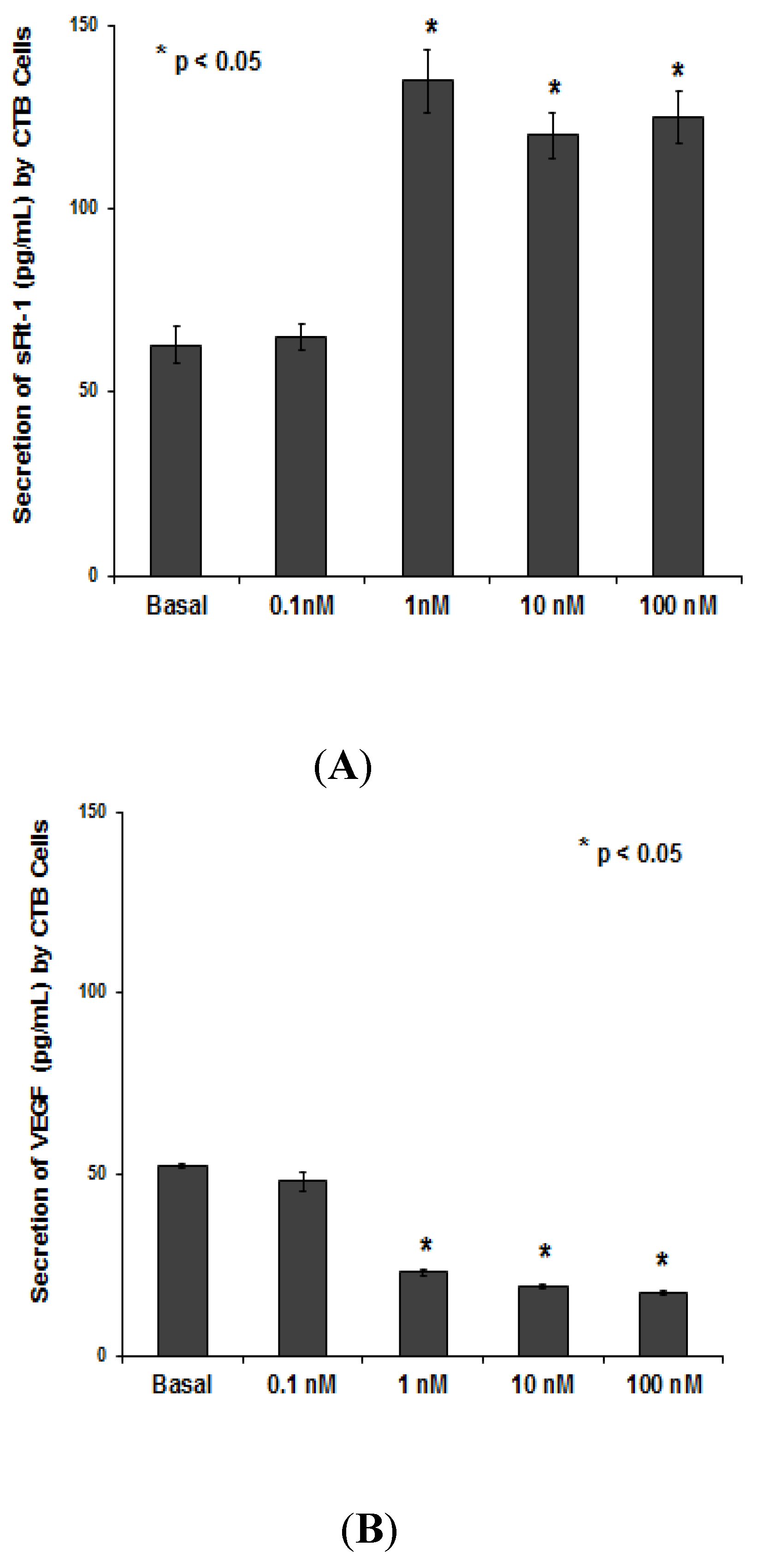
3.1.2. PNTTU Upregulated Anti-angiogenic Factor (sFlt-1) and Downregulated Angiogenic Factor (VEGF)

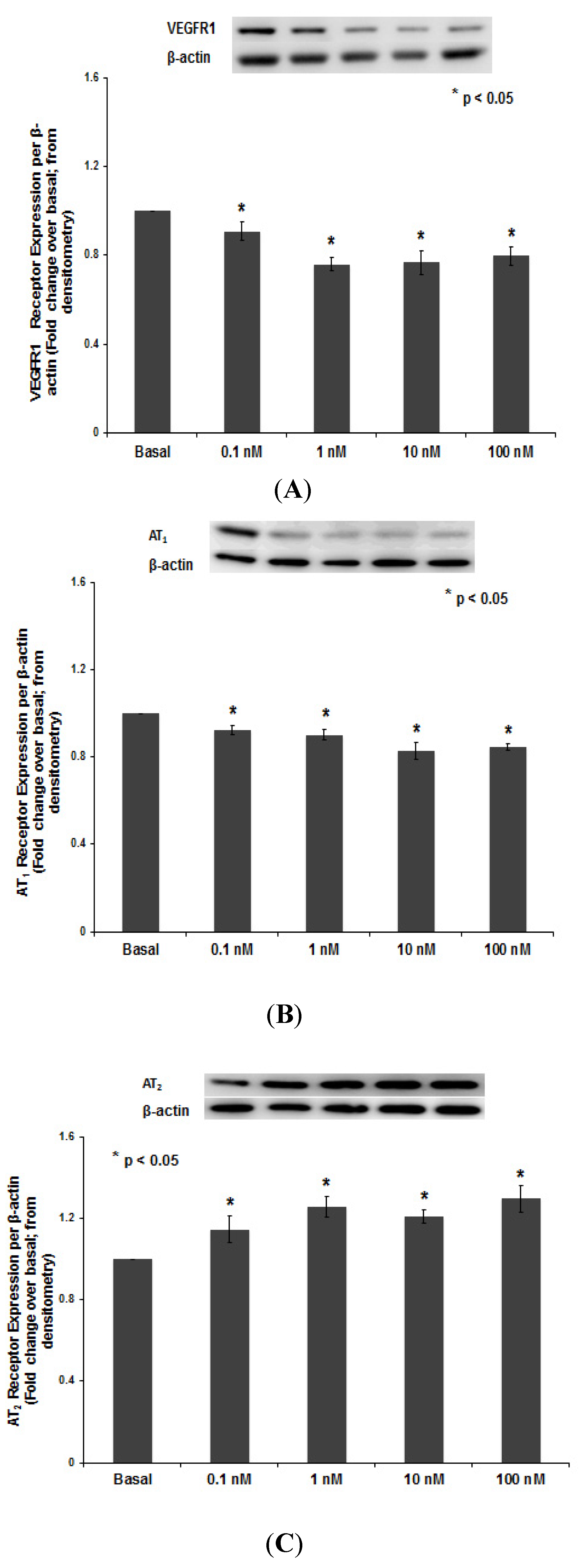
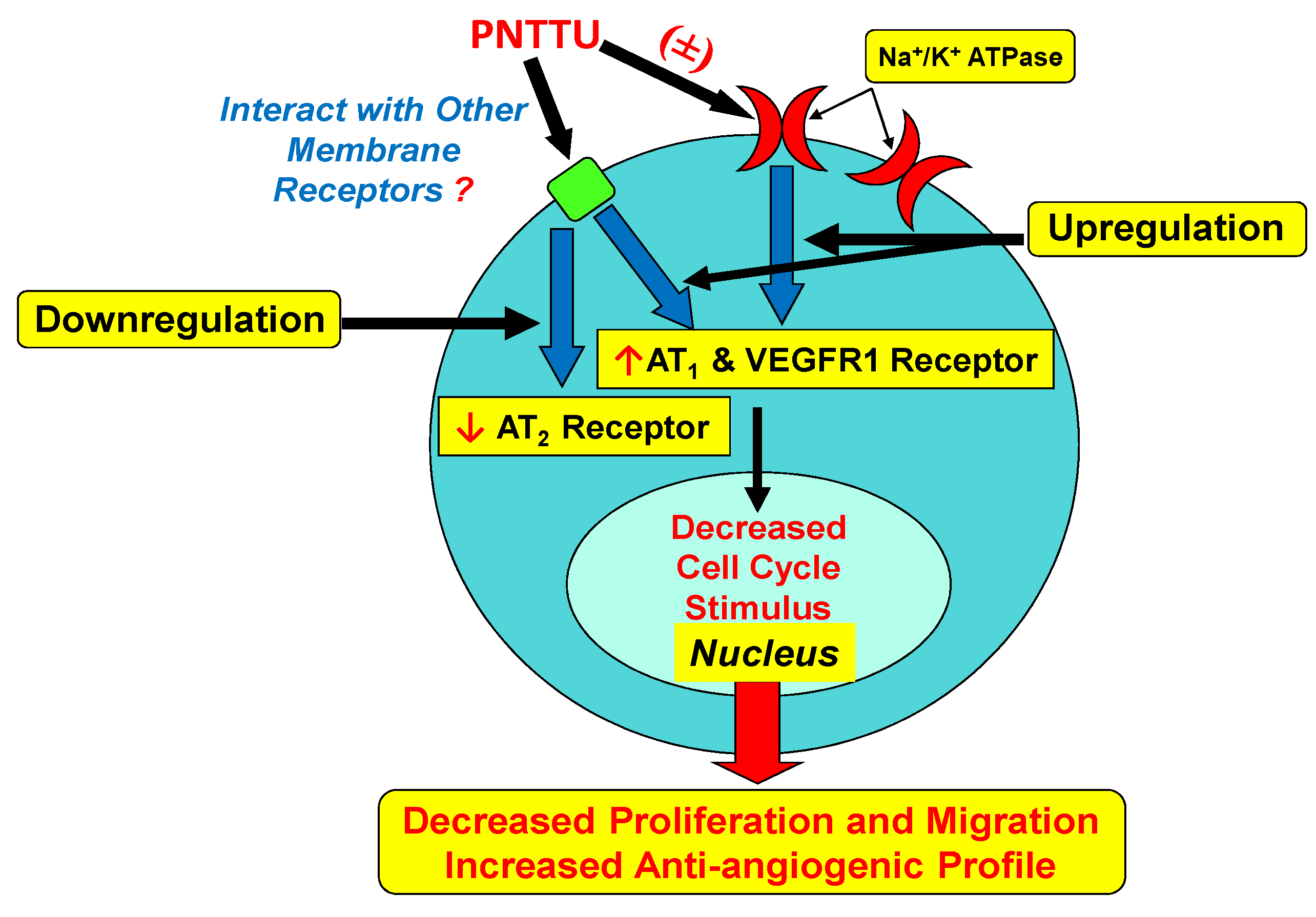
3.1.3. PNTTU Downregulated VEGFR-1 and AT1 Receptor Expression and Upregulated AT2 Receptor Expression

3.2. Discussion

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Berg, C.J.; Atrash, H.K.; Koonin, L.M.; Tucker, M. Pregnancy-related mortality in the United States, 1987. Obstet. Gynecol. 1996, 88, 161–167. [Google Scholar] [CrossRef]
- Pridjian, G.; Puschett, J.B. Preeclampsia. Part 1: Clinical and pathophysiologic considerations. Obstet. Gynecol. Surv. 2002, 57, 598–618. [Google Scholar] [CrossRef]
- Vu, H.; Ianosi-Irimie, M.R.; Pridjian, C.; Whitbred, J.M.; Durst, J.M.; Bagrov, A.Y.; Fedorova, O.V.; Pridjian, G.; Puschett, J.B. The involvement of marinobufagenin in a rat model of human preeclampsia. Amer. J. Nephrol. 2005, 25, 520–528. [Google Scholar] [CrossRef]
- Hamlyn, J.M.; Ringel, R.; Schaeffer, J.; Levinson, P.D.; Hamilton, B.P.; Kowarski, A.A.; Blaustein, M.P. A circulating inhibitor of (Na+ + K+) ATPase associated with essential hypertension. Nature 1982, 300, 650–652. [Google Scholar]
- Mohammadi, K.; Liu, L.; Tian, J.; Kometiani, P.; Xie, Z.; Askari, A. Positive inotropic effect of ouabain on isolated heart is accompanied by activation of signal pathways that link Na+/K+-ATPase to ERK1/2. J. Cardiovasc. Pharmacol. 2003, 41, 609–614. [Google Scholar] [CrossRef]
- Liu, J.; Tian, J.; Haas, M.; Shapiro, J.I.; Askari, A.; Xie, Z. Ouabain interaction with cardiac Na+/K+-ATPase initiates signal cascades independent of changes in intracellular Na+ and Ca2+ concentrations. J. Biol. Chem. 2000, 275, 27838–27844. [Google Scholar]
- Iwamoto, T. Vascular Na+/Ca2+ exchanger: Implications for the pathogenesis and therapy of salt-dependent hypertension. Amer. J. Physiol.-Regul. Integr. C. 2006, 290, 536–545. [Google Scholar] [CrossRef]
- Uddin, M.N.; Horvat, D.; Glaser, S.S.; Danchuk, S.; Mitchell, B.M.; Sullivan, D.E.; Morris, C.A.; Puschett, J.B. Marinobufagenin inhibits proliferation and migration of cytotrophoblast and CHO cells. Placenta 2008, 29, 266–273. [Google Scholar] [CrossRef]
- Uddin, M.N.; Horvat, D.; Glaser, S.S.; Mitchell, B.M.; Puschett, J.B. Examination of the cellular mechanisms by which marinobufagenin inhibits cytotrophoblast function. J. Biol. Chem. 2008, 283, 17946–17953. [Google Scholar]
- Uddin, M.N.; Horvat, D.; Childs, E.W.; Puschett, J.B. Marinobufagenin causes endothelial cell monolayer hyperpermeability by altering apoptotic signaling. Amer. J. Physiol.-Regul. Integr. C. 2009, 296, 1726–1734. [Google Scholar] [CrossRef]
- Uddin, M.N.; Agunanne, E.; Horvat, D.; Puschett, J.B. Marinobufagenin causes enhanced permeability in human brain microvascular endothelial cells via apoptotic signaling. J. Am. Soc. Nephrol. 2009, 20, 534–535. [Google Scholar]
- Uddin, M.N.; Allen, S.; Jones, R.; Zawieja, D.C.; Kuehl, T.J. Pathogenesis of preeclampsia: Marinobufagenin and angiogenic imbalance as biomarkers of the syndrome. Transl. Res. 2012, 160, 99–113. [Google Scholar] [CrossRef]
- Ismet, B.; Emami Khansari, M.; Pramanik, A.; Wong, B.M.; Hossain, M.A. An exclusive fluoride receptor: Fluoride-induced proton transfer to a quinoline-based thiourea. Tetrahedron Lett. 2014, 55, 1467–1470. [Google Scholar]
- Khansari, M.E.; Wallace, K.D.; Hossain, M.A. Synthesis and anion recognition studies of a dipodal thiourea-based sensor for anions. Tetrahedron Lett. 2014, 55, 438–440. [Google Scholar] [CrossRef]
- Russ, T.H.; Pramanik, A.; Khansari, M.E.; Wong, B.M.; Hossain, M.A. A quinoline based bis-urea receptor for anions: A selective receptor for hydrogen sulfate. Nat. Prod. Commun. 2012, 7, 301–304. [Google Scholar]
- Pramanik, A.; Thompson, B.; Hayes, T.; Tucker, K.; Powell, D.R.; Bonnesen, P.V.; Ellis, E.D.; Lee, K.S.; Yu, H.; Hossain, M.A. Seven-coordinate anion complex with a tren-based urea: Binding discrepancy of hydrogen sulfate in solid and solution states. Org. Biomol. Chem. 2011, 9, 4444–4447. [Google Scholar] [CrossRef]
- Busschaert, N.; Wenzel, M.; Light, M.E.; Iglesias-Hernandez, P.; Perez-Tomas, R.; Gale, P.A. Structure-activity relationships in tripodal transmembrane anion transporters: The effect of fluorination. J. Am. Chem. Soc. 2011, 133, 14136–14148. [Google Scholar]
- Werner, F.; Schneider, H.-J. Complexation of anions including nucleotide anions by open-chain host compounds with amide, urea, and aryl functions. Helv. Chim. Acta 2000, 83, 465–478. [Google Scholar] [CrossRef]
- Dey, S.K.; Das, G. Encapsulation of trivalent phosphate anion within a rigidified p-stacked dimeric capsular assembly of tripodal receptor. Dalton Trans. 2011, 40, 12048–12051. [Google Scholar]
- Dey, S.K.; Das, G. Selective inclusion of PO43− within persistent dimeric capsules of a tris(thiourea) receptor and evidence of cation/solvent sealed unimolecular capsules. Dalton Trans. 2012, 41, 8960–8972. [Google Scholar]
- Pramanik, A.; Powell, D.R.; Wong, B.M.; Hossain, M.A. Spectroscopic, structural, and theoretical studies of halide complexes with a urea-based tripodal receptor. Inorg. Chem. 2012, 51, 4274–4284. [Google Scholar]
- Bagrov, A.Y.; Shapiro, J.I.; Fedorova, O.V. Endogenous cardiotonic steroids: Physiology, pharmacology, and novel therapeutic targets. Pharmacol. Rev. 2009, 61, 9–38. [Google Scholar] [CrossRef]
- Blaustein, M.P. Sodium ions, calcium ions, blood pressure regulation and hypertension: A reasessment and a hypothesis. Amer. J. Physiol. 1977, 232, 167–173. [Google Scholar]
- Dmitrieva, R.I.; Doris, P.A. Ouabain is a potent promoter of growth and activator of ERK1/2 in ouabain-resistant rat renal epithelial cells. J. Biol. Chem. 2003, 278, 28160–28166. [Google Scholar]
- Saunders, R.; Scheiner-Bobis, G. Ouabain stimulates endothelin release and expression in human endothelial cells without inhibiting the sodium pump. Eur. J. Biochem. 2004, 271, 1054–1062. [Google Scholar] [CrossRef]
- Chueh, S.C.; Guh, J.H.; Chen, J.; Lai, M.K.; Teng, C.M. Dual effects of ouabain on the regulation of proliferation and apoptosis in human prostatic smooth muscle cells. J. Urol. 2001, 166, 347–353. [Google Scholar] [CrossRef]
- Huang, Y.T.; Chueh, S.C.; Teng, C.M.; Guh, J.H. Investigation of ouabain-induced anticancer effect in human androgen-independent prostate cancer PC-3 cells. Biochem. Pharmacol. 2004, 67, 727–733. [Google Scholar] [CrossRef]
- Schoner, W.; Scheiner-Bobis, G. Endogenous and exogenous cardiac glycosides: Their roles in hypertension, salt metabolism, and cell growth. Amer. J. Physiol. Cell Physiol. 2007, 293, 509–536. [Google Scholar] [CrossRef]
- Uddin, M.N.; Horvat, D.; DeMorrow, S.; Agunanne, E.; Puschett, J.B. Marinobufagenin is an upstream modulator of Gadd45a stress signaling in preeclampsia. Biochim. Biophys. Acta 2011, 1812, 49–58. [Google Scholar]
- Cartwright, J.E.; Holden, D.P.; Whitley, G.S. Hepatocyte growth factor regulates human trophoblast motility and invasion: A role for nitric oxide. Brit. J. Pharmacol. 1999, 128, 181–189. [Google Scholar] [CrossRef]
- Cartwright, J.E.; Kenny, L.C.; Dash, P.R.; Crocker, I.P.; Aplin, J.D.; Baker, P.N.; Whitley, G.S. Trophoblast invasion of spiral arteries: A novel in vitro model. Placenta 2002, 23, 232–235. [Google Scholar] [CrossRef]
- Choy, M.Y.; Manyonda, M.Y. The phagocytic activity of human first trimester extravillous trophoblast. Hum. Reprod. 1998, 13, 2941–2949. [Google Scholar]
- Choy, M.Y.; Whitley, G.S.; Manyonda, I.T. Efficient, rapid and reliable establishment of human trophoblast cell lines using poly-L-ornithine. Early Preg. 2000, 4, 124–143. [Google Scholar]
- Ehrig, J.; Horvat, D.; Fothergill, R.E.; Allen, S.R.; Jones, R.O.; Zawieja, D.C.; Kuehl, T.J.; Uddin, M.N. Cardiotonic steroids induce an anti-angiogenic profile in first trimester cytotrophoblast cells. Amer. J. Obstet Gynecol. 2013, 208. [Google Scholar] [CrossRef]
- Fisher, S.J. The placental problem: Linking abnormal cytotrophoblast differentiation to the maternal symptoms of preeclampsia. Reprod. Biol. Endocrinol. 2004, 2. [Google Scholar] [CrossRef] [Green Version]
- Kharfi, A.; Giguère, Y.; Sapin, V.; Massé, J.; Dastugue, B.; Forest, J.C. Trophoblastic remodeling in normal and preeclamptic pregnancies: Implication of cytokines. Clin. Biochem. 2003, 36, 323–331. [Google Scholar] [CrossRef]
- Cunningham, F.G.; Leveno, K.J.; Bloom, S.L.; Hauth, J.C.; Rouse, D.J.; Spong, C.Y. Implantation, Embryogenesis and Placentation. In Williams Obstetrics, 23rd ed.; McGraw-Hill: New York, NY, USA, 2010. [Google Scholar]
- Gonick, H.C.; Ding, Y.; Vaziri, N.D.; Bagrov, A.Y.; Fedorova, O.V. Simultaneous measurement of marinobufagenin, ouabain, and hypertension-associated protein in various disease states. Clin. Exp. Hypertension 1998, 20, 617–627. [Google Scholar] [CrossRef]
- Puschett, J.B.; Agunanne, E.; Uddin, M.N. Emerging role of the bufodienolides in cardiovascular and renal disease. Amer. J. Kidney Dis. 2010, 56, 359–370. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Horvat, D.; Khansari, M.E.; Pramanik, A.; Beeram, M.R.; Kuehl, T.J.; Hossain, M.A.; Uddin, M.N. A Synthetic Thiourea-Based Tripodal Receptor that Impairs the Function of Human First Trimester Cytotrophoblast Cells. Int. J. Environ. Res. Public Health 2014, 11, 7456-7469. https://doi.org/10.3390/ijerph110707456
Horvat D, Khansari ME, Pramanik A, Beeram MR, Kuehl TJ, Hossain MA, Uddin MN. A Synthetic Thiourea-Based Tripodal Receptor that Impairs the Function of Human First Trimester Cytotrophoblast Cells. International Journal of Environmental Research and Public Health. 2014; 11(7):7456-7469. https://doi.org/10.3390/ijerph110707456
Chicago/Turabian StyleHorvat, Darijana, Maryam Emami Khansari, Avijit Pramanik, Madhava R. Beeram, Thomas J. Kuehl, Md. Alamgir Hossain, and Mohammad Nasir Uddin. 2014. "A Synthetic Thiourea-Based Tripodal Receptor that Impairs the Function of Human First Trimester Cytotrophoblast Cells" International Journal of Environmental Research and Public Health 11, no. 7: 7456-7469. https://doi.org/10.3390/ijerph110707456