Histological Lesions, Cell Cycle Arrest, Apoptosis and T Cell Subsets Changes of Spleen in Chicken Fed Aflatoxin-contaminated Corn

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diets and Study Design

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet 1 | Control | 25% | 50% | 75% | 100% | Control Corn | Contaminated Corn |
|---|---|---|---|---|---|---|---|
| 1~21d | |||||||
| AFB1 | ND 2 | 16.3 | 36.9 | 45.6 | 82.4 | ND | 149.6 |
| 22~42d | |||||||
| AFB1 | ND | 34.3 | 69.3 | 95.2 | 134 | ND | 229 |
2.2. Relative Weight of Spleen
2.3. Pathological Observation
2.4. Cell Cycle Phase Detection
2.5. Apoptosis Detection
2.6. Cell T Cell Subsets Detection by Flow Cytometry
3. Results
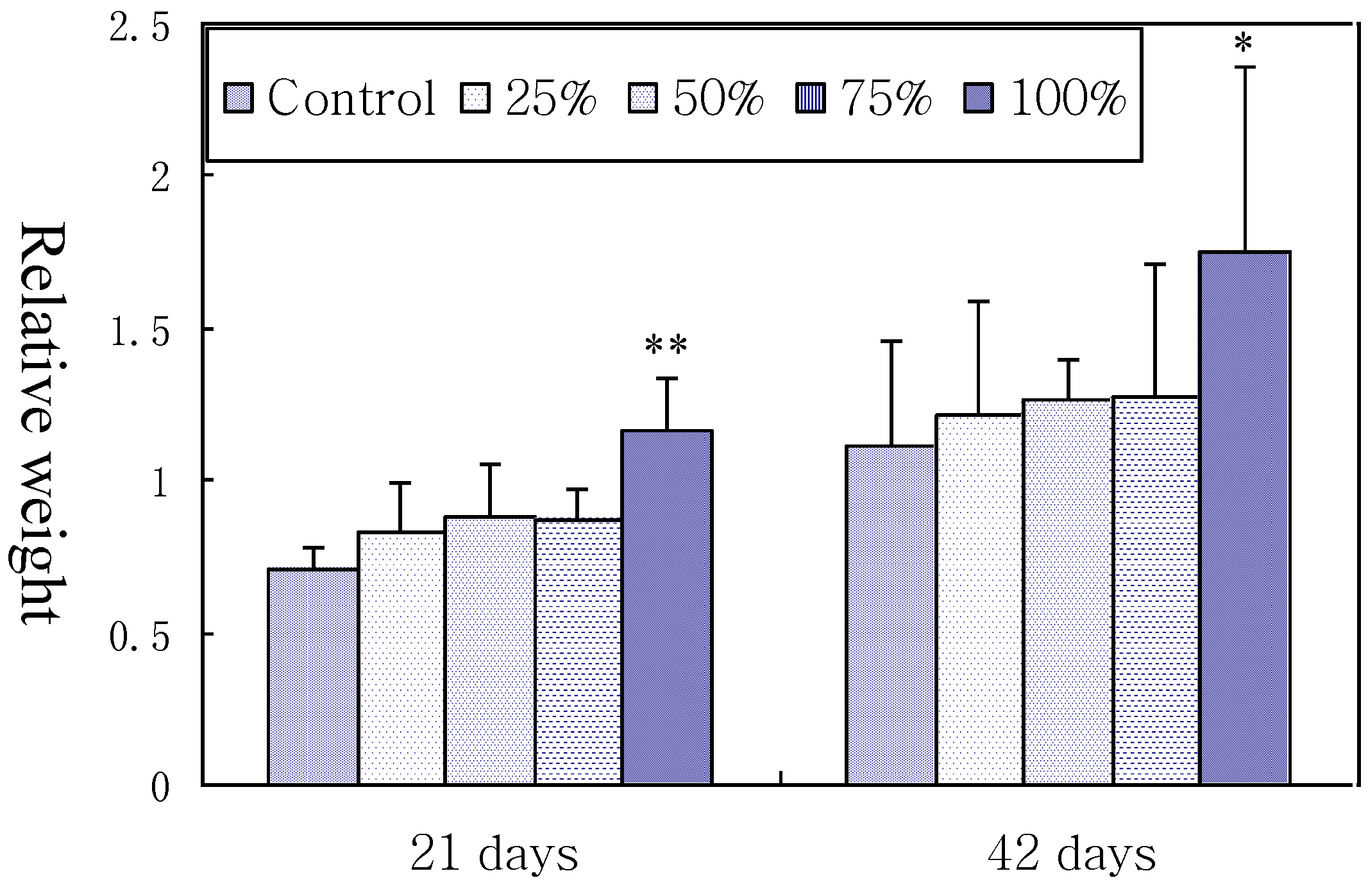
3.1. Relative Weight of Spleen

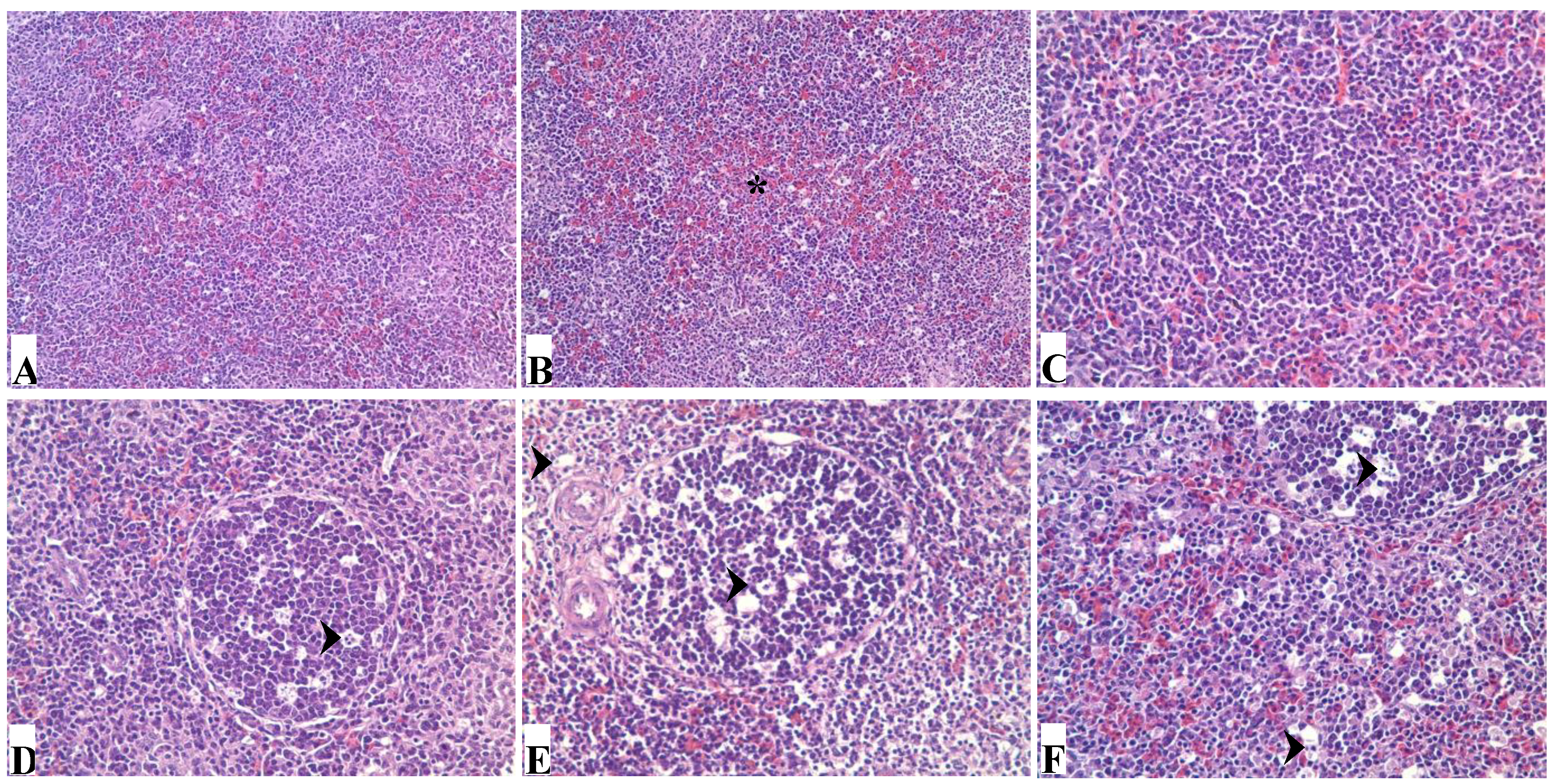
3.2. Pathological Lesions


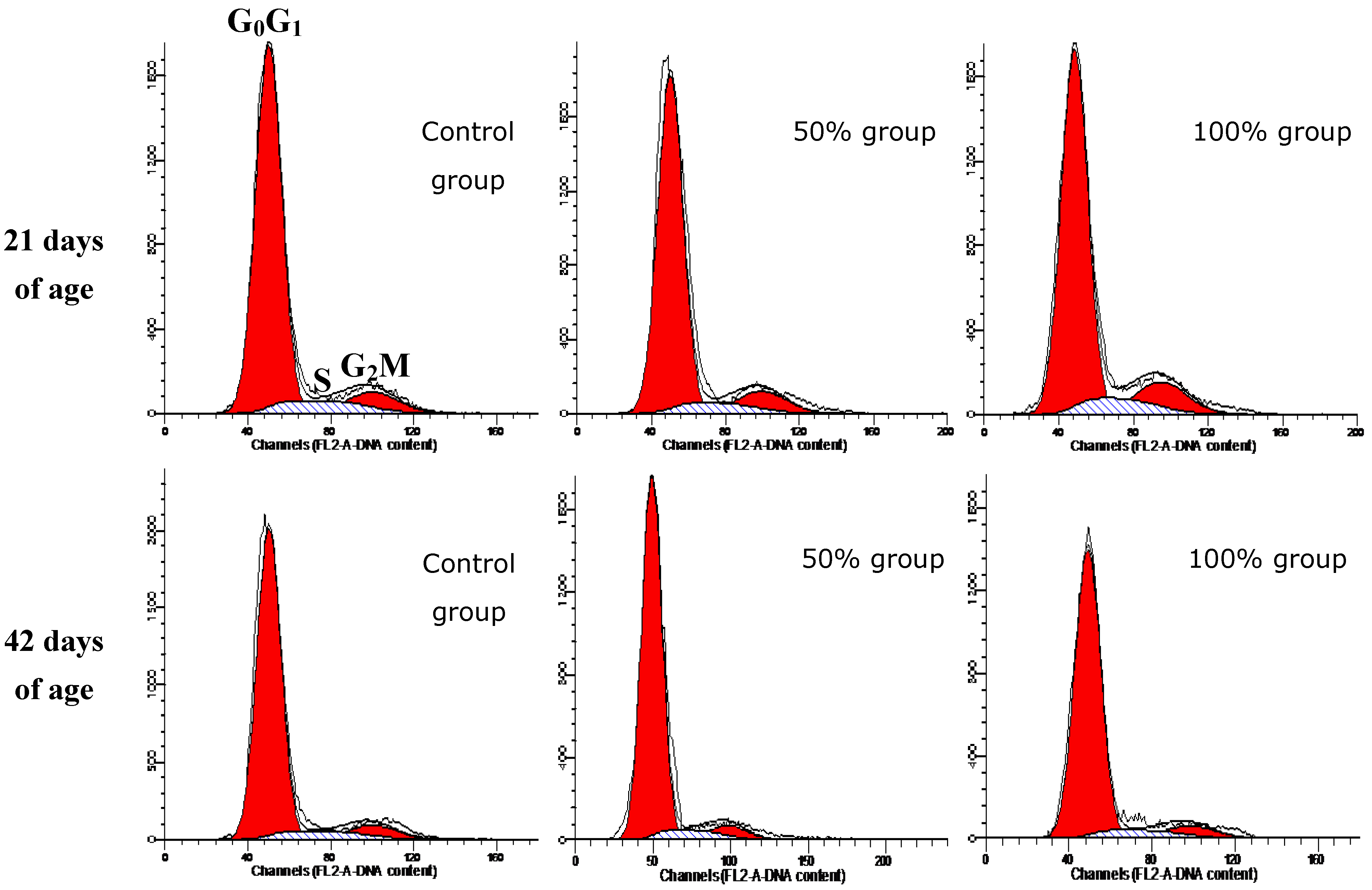
3.3. Cell Cycle Phase-Distribution of Splenocytes
| Time | Group | G0G1 Phase (%) | G2M Phase (%) | S Phase (%) | PI |
|---|---|---|---|---|---|
| 21 days | Control | 82.04 ± 2.87 | 9.05 ± 1.51 | 8.91 ± 1.40 | 17.96 ± 2.87 |
| 25% | 80.95 ± 3.27 | 9.49 ± 1.56 | 9.55 ± 1.72 | 19.05 ± 3.27 | |
| 50% | 79.60 ± 1.17 | 10.71 ± 0.74 * | 9.69 ± 0.82 | 20.40 ± 1.17 | |
| 75% | 79.32 ± 1.86 | 11.68 ± 1.28 ** | 9.00 ± 1.02 | 20.68 ± 1.86 | |
| 100% | 78.37 ± 0.80 * | 12.88 ± 0.50 ** | 8.74 ± 0.38 | 21.63 ± 0.80 * | |
| 42 days | Control | 84.56 ± 0.67 | 7.69 ± 0.73 | 7.75 ± 0.41 | 15.44 ± 0.67 |
| 25% | 84.47 ± 2.12 | 7.50 ± 0.99 | 8.02 ± 1.14d | 15.53 ± 2.12 | |
| 50% | 85.52 ± 1.65 | 7.25 ± 0.91 | 7.23 ± 1.00 | 14.45 ± 1.65 | |
| 75% | 86.80 ± 1.32 * | 6.81 ± 0.64 | 6.39 ± 0.84 * | 13.20 ± 1.32 * | |
| 100% | 87.42 ± 0.70 ** | 6.25 ± 0.22 ** | 6.33 ± 0.84 * | 12.58 ± 0.70 ** |

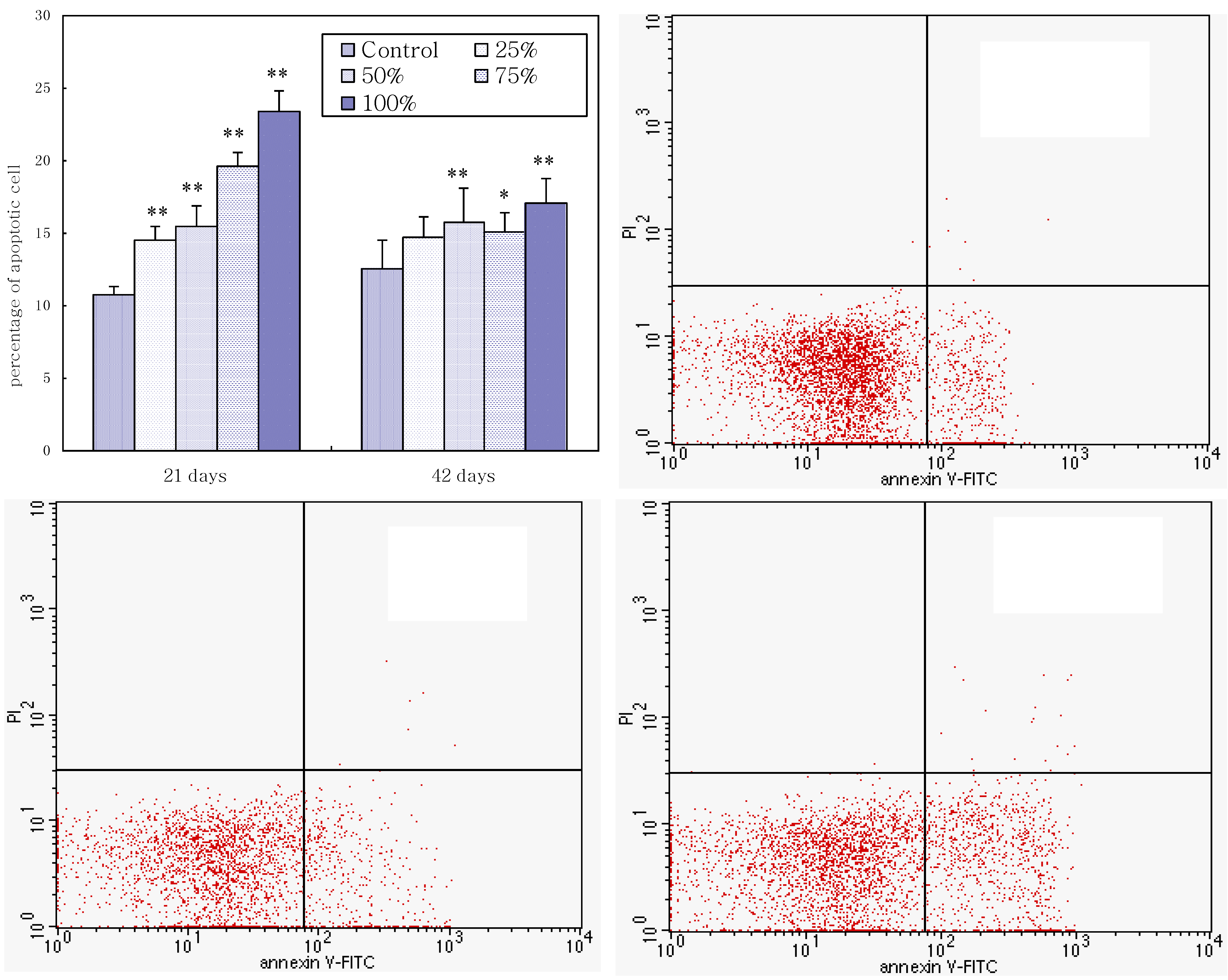
3.4. Annexin V-FITC Staining Assay by Flow Cytometry

3.5. Splenic T Cell Subsets
| Time | Items | CD3+ | CD3+CD4+ | CD3+CD8+ | CD3+CD4+/CD3+CD8+ |
|---|---|---|---|---|---|
| 21 days | Control | 64.73 ± 3.05 | 34.67 ± 2.08 | 27.47 ± 2.19 | 1.27 ± 0.08 |
| 25% | 65.38 ± 4.85 | 33.02 ± 6.06 | 32.34 ± 7.39 | 1.06 ± 0.30 | |
| 50% | 64.13 ± 5.85 | 33.98 ± 4.84 | 32.17 ± 5.91 | 1.09 ± 0.30 | |
| 75% | 62.26 ± 5.11 | 35.39 ± 5.68 | 36.04 ± 5.49 * | 0.98 ± 0.07 * | |
| 100% | 59.57 ± 4.77 | 30.19 ± 1.96 | 34.34 ± 2.09 * | 0.88 ± 0.08 ** | |
| 42 days | Control | 77.00 ± 5.26 | 31.83 ± 5.32 | 34.94 ± 3.48 | 0.91 ± 0.07 |
| 25% | 74.95 ± 4.83 | 30.11 ± 5.47 | 34.94 ± 6.59 | 0.89 ± 0.25 | |
| 50% | 77.34 ± 2.94 | 29.34 ± 6.67 | 37.20 ± 2.92 | 0.79 ± 0.19 | |
| 75% | 75.20 ± 3.81 | 31.37 ± 3.92 | 36.59 ± 2.44 | 0.86 ± 0.14 | |
| 100% | 79.06 ± 5.13 | 26.65 ± 4.08 | 43.72 ± 5.91 ** | 0.62 ± 0.12 * |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bryden, W.L. Mycotoxins in the food chain: Human health implications. Asia Pac. J. Clin. Nutr. 2007, 16, 95–101. [Google Scholar] [PubMed]
- Wogan, G.N. Aflatoxins as risk factors for hepatocellular carcinoma in humans. Cancer Res. 1992, 52, 2114s–2118s. [Google Scholar] [PubMed]
- International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; World Health Organization: Lyon, France, 2002; pp. 171–175. [Google Scholar]
- Wu, H.C.; Wang, Q.; Yang, H.I.; Ahsan, H.; Tsai, W.Y.; Wang, L.Y.; Chen, S.Y.; Chen, C.J.; Santella, R.M. Aflatoxin B1 exposure, hepatitis B virus infection, and hepatocellular carcinoma in Taiwan. Cancer Epidem. Biomar. 2009, 18, 846. [Google Scholar]
- Wild, C.P.; Turner, P.C. The toxicology of aflatoxins as a basis for public health decisions. Mutagenesis 2002, 17, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Rawal, S.; Kim, J.E.; Coulombe, R., Jr. Aflatoxin B1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Magnoli, A.P.; Monge, M.P.; Miazzo, R.D.; Cavaglieri, L.R.; Magnoli, C.E.; Merkis, C.I.; Cristofolini, A.L.; Dalcero, A.M.; Chiacchiera, S.M. Effect of low levels of aflatoxin B1 on performance, biochemical parameters, and aflatoxin B1 in broiler liver tissues in the presence of monensin and sodium bentonite. Poult. Sci. 2011, 90, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.A.; Khan, M.Z.; Khan, A.; Hussain, I. Pathological effects of aflatoxin and their amelioration by vitamin E in White Leghorn layers. Pak. Vet. J. 2010, 30, 155–162. [Google Scholar]
- Altamura, M.; Caradonna, L.; Amati, L.; Pellegrino, N.M.; Urgesi, G.; Miniello, S. Splenectomy and sepsis: The role of the spleen in the immune-mediated bacterial clearance. Immunopharmacol. Immunotoxicol. 2001, 23, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Hinton, D.M.; Myers, M.J.; Raybourne, R.A.; Francke, C.S.; Sotomayor, R.E.; Shaddock, J.; Warbritton, A.; Chou, M.W. Immunotoxicity of aflatoxin B1 in rats: Effects on lymphocytes and the inflammatory response in a chronic intermittent dosing study. Toxicol. Sci. 2003, 73, 362–377. [Google Scholar] [PubMed]
- Mary, V.S.; Theumer, M.G.; Arias, S.L.; Rubinstein, H.R. Reactive oxygen species sources and biomolecular oxidative damage induced by aflatoxin B1 and fumonisin B1 in rat spleen mononuclear cells. Toxicology 2012, 302, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Theumer, M.G.; Lopez, A.G.; Masih, D.T.; Chulze, S.N.; Rubinstein, H.R. Immuno-biological effects of AFB1 and AFB1–FB1 mixture in experimental subchronic mycotoxicoses in rats. Toxicology 2003, 186, 159–170. [Google Scholar] [PubMed]
- Morris, S.M.; Aidoo, A.; Chen, J.J.; Chou, M.W.; Casciano, D.A. Aflatoxin B1-induced Hprt mutations in splenic lymphocytes of fischer 344 rats: Results of an intermittent feeding trial. Mutat. Res. 1999, 423, 33–38. [Google Scholar] [PubMed]
- Sabourin, P.J.; Price, J.A.; Casbohm, S.L.; Perry, M.R.; Tuttle, R.S.; Rogers, J.V.; Rowell, K.S.; Estep, J.E.; Sabourin, C.L. Evaluation of acute immunotoxicity of aerosolized aflatoxin B1 in female C57BL/6N mice. J. Immunotoxicol. 2006, 3, 11–20. [Google Scholar] [PubMed]
- Méndez-Albores, A.; Del Río-García, J.C.; Moreno-Martínez, E. Decontamination of aflatoxin duckling feed with aqueous citric acid treatment. Anim. Feed Sci. Tech. 2007, 135, 249–262. [Google Scholar]
- Ortatatli, M.; Oğuz, H.; Hatipoğlu, F.; Karaman, M. Evaluation of pathological changes in broilers during chronic aflatoxin (50 and 100 ppb) and clinoptilolite exposure. Res. Vet. Sci. 2005, 78, 61–68. [Google Scholar] [PubMed]
- Yang, J.; Bai, F.; Zhang, K.; Lv, X.; Bai, S.; Zhao, L.; Peng, X.; Ding, X.; Li, Y.; Zhang, J. Effects of feeding corn naturally contaminated with AFB1 and AFB2 on performance and aflatoxin residues in broilers. Czech. J. Anim. Sci. 2012, 57, 506–515. [Google Scholar]
- Yang, J.; Bai, F.; Zhang, K.; Bai, S.; Peng, X.; Ding, X.; Li, Y.; Zhang, J.; Zhao, L. Effects of feeding corn naturally contaminated with aflatoxin B1 and B2 on hepatic functions of broilers. Poult. Sci. 2012, 91, 2792–2801. [Google Scholar] [PubMed]
- Administration of Quality and Technology Supervision of Sichuan Province. Official Standard of Administration of Quality and Technology Supervision of Sichuan Province of PRC, Determination of Aflatoxin B1, B2, G1, G2, T-2 Toxin, Deoxynicalenol, Zearalenone, Ochratoxin, Fumonisin B1 with HPLC; People’s Government of Sichuan Province: Chengdu, China, 2010. (In Chinese)
- Edrington, T.S.; Kubena, L.F.; Harvey, R.B.; Rottinghaus, G.E. Influence of a superactivated charcoal on the toxic effects of aflatoxin or T-2 toxin in growing broilers. Poult. Sci. 1997, 76, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Rosa, C.A.; Miazzo, R.C.; Magnoli, M.; Salvano, S.; Chiacchiera, M.; Frrero, S.; Saenz, M.; Carvalho, E.C.; Dalcero, A. Evaluation of the efficacy of bentonite from the south of Argentina to ameliorate the toxic effects of aflatoxin in broilers. Poult. Sci. 2001, 80, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Solcan, C.; Gogu, M.; Floristean, V.; Oprisan, B.; Solcan, G. The hepatoprotective effect of sea buckthorn (Hippophae rhamnoides) berries on induced aflatoxin B1 poisoning in chickens. Poult. Sci. 2013, 92, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Riddell, C. Lymphoid System; Allen Press Inc.: Kansas, KC, USA, 1987. [Google Scholar]
- Scott, T.R.; Rowland, S.M.; Rodgers, R.S.; Bodine, A.B. Genetic selection for aflatoxin B1 resistance influences chicken T-cell and thymocyte proliferation. Dev. Comp. Immunol. 1991, 15, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Bianco, G.; Russo, R.; Marzocco, S.; Velotto, S.; Autore, G.; Severino, L. Modulation of macrophage activity by aflatoxins B1 and B2 and their metabolites aflatoxins M1 and M2. Toxicon 2012, 59, 644–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Zhang, Z.; Wang, X.; Wang, Y.; Zhang, X.; Lu, H.; Wang, S. Cytochrome P450 2A13 enhances the sensitivity of human bronchial epithelial cells to aflatoxin B1-induced DNA damage. Toxicol. Appl. Pharm. 2013, 270, 114–121. [Google Scholar] [CrossRef]
- Barreto Filho, J.B.; Marques Júnior, A.P. Histological aspects of Zebu Cow’s placenta. Arq. Bras. Med. Vet. Zootec. 1993, 45, 385–393. [Google Scholar]
- Hetts, S.W. To die or not to die: An overview of apoptosis and its role in disease. J. Am. Med. Assoc. 1998, 279, 300–307. [Google Scholar] [CrossRef]
- Sun, E.W.; Shi, Y.F. Apoptosis: The quiet death silences the immune system. Pharmacol. Therapeut. 2001, 92, 135–145. [Google Scholar] [CrossRef]
- Meki, A.R.; Abdel-Ghaffar, S.K.; El-Gibaly, I. Aflatoxin B1 induces apoptosis in rat liver: Protective effect of melatonin. Neuro. Endocrinol. Lett. 2001, 22, 417–426. [Google Scholar] [PubMed]
- Hanumantharao, G.R.; Ekta, K.; Vishwajeet, R.; Dwarakanat, B.S.; Virinder, S.P.; Smita, M.; Adhikari, J.S.; Yogesh, K.T.; Sanjay, G.; Mridula, B.; et al. Acetoxy-4-Methylcoumarins confer differential protection from aflatoxin B1-induced micronucei and apoptosis in lung and bone marrow cells. Mutat. Res. 2001, 494, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Lu, H.Y.; Li, Z.Y.; Bian, Q.; Qiu, L.L.; Li, Z.; Liu, Q.; Li, J.; Wang, X.J.; Wang, S. Cytochrome P450 2A13 mediates Aflatoxin B1 induced cytotoxicity and apoptosis in human bronchial epithelial cells. Toxicology 2012, 300, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Y.; Shu, G.; Peng, X.; Fang, J.; Chen, K.J.; Cui, H.M.; Chen, Z.L.; Zuo, Z.C.; Deng, J.L.; Geng, Y.; et al. Protective effects of sodium selenite against aflatoxin B1-Induced oxidative stress and apoptosis in broiler spleen. Inter. J. Env. Res. Pub. Heal. 2013, 7, 2834–2844. [Google Scholar] [CrossRef]
- Duan, X.X.; Ou, J.S.; Li, Y.; Su, J.J.; Ou, C.; Yang, C.H.; Yue, F.; Ban, K.C. Dynami expression of apoptosis-related genes during development of laboratory hepatocellular carcinoma and its relation to apoptosis. World J. Gastroenterol. 2005, 11, 4740–4744. [Google Scholar] [PubMed]
- Van Vleet, T.R.; Watterson, T.L.; Klein, P.J.; Coulombe, R.A., Jr. Aflatoxin B1 alters the expression of p53 in cytochrome P450-expressing human lung cells. Toxicol. Sci. 2006, 89, 399–407. [Google Scholar]
- Chen, K.J.; Shu, G.; Peng, X.; Fang, J.; Cui, H.M.; Chen, J.; Wang, F.Y.; Chen, Z.L.; Zuo, Z.C.; Deng, J.L.; et al. Protective role of sodium selenite on histopathological lesions, decreased T-cell subsets and increased apoptosis of thymus in broilers intoxicated with aflatoxin B1. Food Chem. Toxicol. 2013, 59, 308–316. [Google Scholar]
- Pavelka, M.; Roth, J. Functional Ultrastructure-Atlas of Tissue Biology and Pathology; Springer: New York, NY, USA, 2005. [Google Scholar]
- Tomková, I.; Ševčiková, Z.; Levkut, M.; Revajová, V.; Čonková, E.; Laciaková, A.; Lenhardt, L. Effect of aflatoxin B1 on CD3 T cells and alkaline phosphatase in the intestine of mice. Mycopathologia 2002, 154, 15–19. [Google Scholar]
- Sabourin, P.J.; Price, J.A.; Casbohm, S.L. Perry, M.R.; Tuttle, R.S.; Rogers, J.V.; Rowell, K.S.; Estep, J.E.; Sabourin, C.L. Evaluation of acute immunotoxicity of aerosolized aflatoxin B1 in female C57BL/6N mice. J. Immunotoxicol. 2006, 3, 11–20. [Google Scholar]
- Xie, Y.; Wang, L.; Freywald, A.; Qureshi, M.; Chen, Y.; Xiang, J. A novel T cell-based vaccine capable of stimulating long-term functional CTL memory against B16 melanoma via CD40L signaling. Cell. Mol. Immunol. 2012, 10, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; Bevan, M.J. Effector and memory CTL differentiation. Annu. Rev. Immunol. 2007, 25, 171–192. [Google Scholar] [CrossRef] [PubMed]
- Prado-Garcia, H.; Romero-Garcia, S.; Aguilar-Cazares, D.; Meneses-Flores, M.; Lopez-Gonzalez, J.S. Tumor-induced CD8+ T-cell dysfunction in lung cancer patients. Clin. Dev. Immunol. 2012. [Google Scholar] [CrossRef]
- Benford, D.; Leblanc, J.; Setzer, R.W. Application of the margin of exposure (MoE) approach to substances in food that are genotoxic and carcinogenic: Example: Aflatoxin B1 (AFB1). Food Chem. Toxicol. 2010, 48, S34–S41. [Google Scholar] [CrossRef] [PubMed]
- Bailey, G.S.; Williams, D.E.; Wilcox, J.S.; Loveland, P.M.; Coulombe, R.A.; Hendricks, J.D. Aflatoxin B1 carcinogenesis and its relation to DNA adduct formation and adduct persistence in sensitive and resistant salmonid fish. Carcinogenesis 1988, 9, 1919–1926. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Peng, X.; Zhang, K.; Bai, S.; Ding, X.; Zeng, Q.; Yang, J.; Fang, J.; Chen, K. Histological Lesions, Cell Cycle Arrest, Apoptosis and T Cell Subsets Changes of Spleen in Chicken Fed Aflatoxin-contaminated Corn. Int. J. Environ. Res. Public Health 2014, 11, 8567-8580. https://doi.org/10.3390/ijerph110808567
Peng X, Zhang K, Bai S, Ding X, Zeng Q, Yang J, Fang J, Chen K. Histological Lesions, Cell Cycle Arrest, Apoptosis and T Cell Subsets Changes of Spleen in Chicken Fed Aflatoxin-contaminated Corn. International Journal of Environmental Research and Public Health. 2014; 11(8):8567-8580. https://doi.org/10.3390/ijerph110808567
Chicago/Turabian StylePeng, Xi, Keying Zhang, Shiping Bai, Xuemei Ding, Qiufeng Zeng, Jun Yang, Jing Fang, and Kejie Chen. 2014. "Histological Lesions, Cell Cycle Arrest, Apoptosis and T Cell Subsets Changes of Spleen in Chicken Fed Aflatoxin-contaminated Corn" International Journal of Environmental Research and Public Health 11, no. 8: 8567-8580. https://doi.org/10.3390/ijerph110808567