
MiRNAs and miRNA Polymorphisms Modify Drug Response

Abstract
:1. Introduction
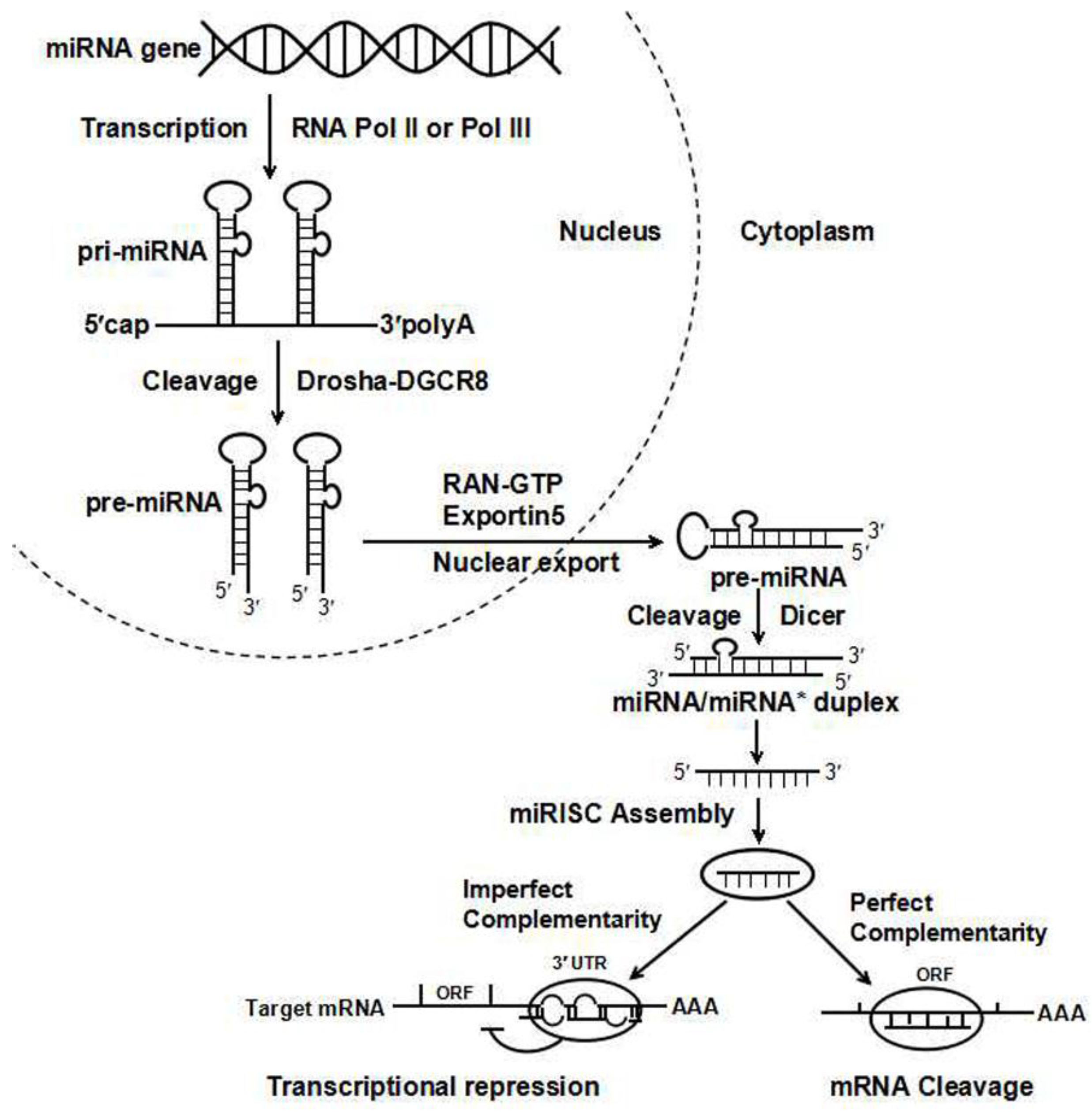
2. MiRNAs: A New Player in Gene Function and Drug Response
3. MiRNAs-Mediated Regulation of DMEs and Drug Transporters
3.1. Direct Repression of DME by miRNAs
3.2. Direct Repression of Drug Transporters by miRNAs
3.3. Indirect Regulation of DMEs and Drug Transporters by miRNAs
3.4. MiRNAs Mediate Drug-Drug Interactions in Pharmacokinetics
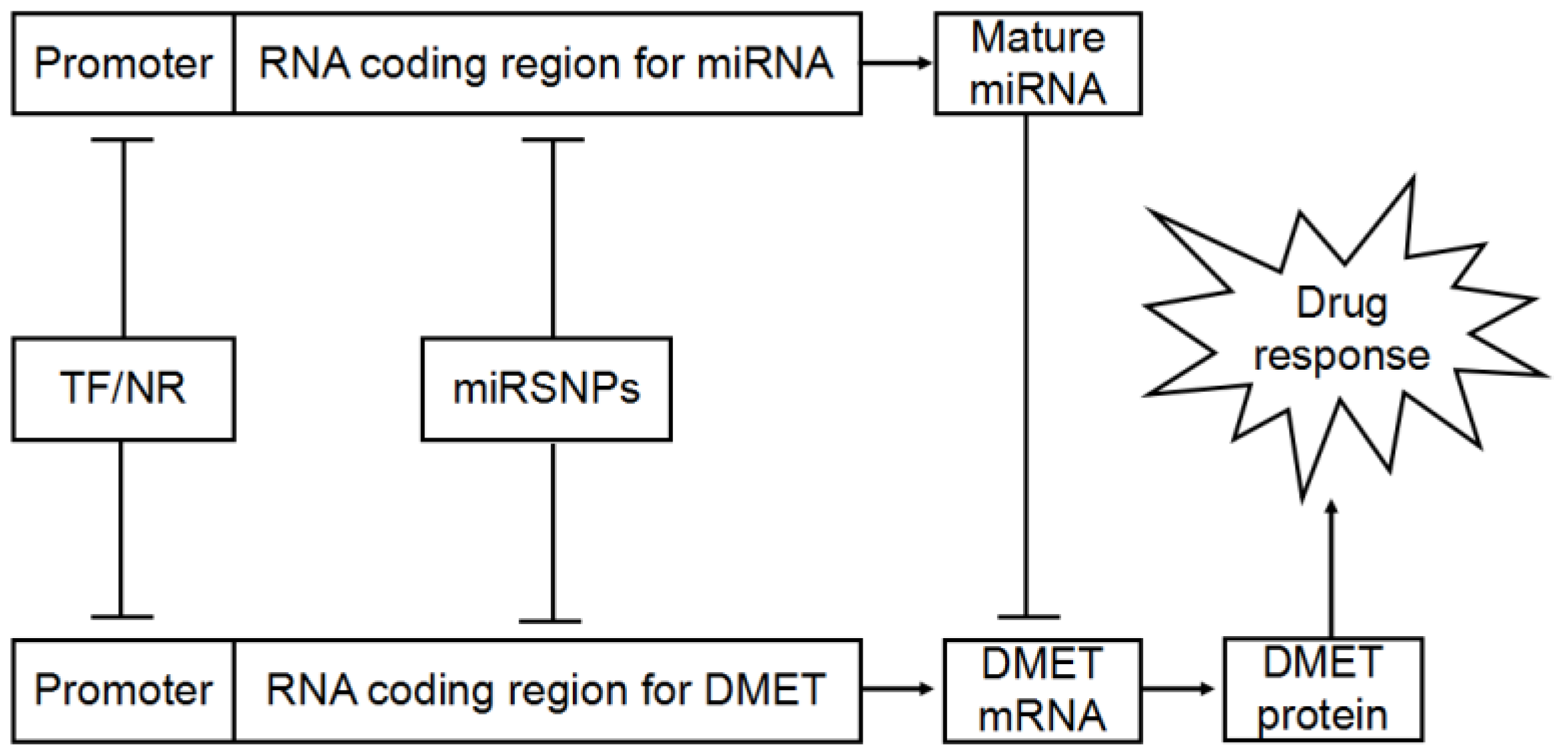
4. MiRSNPs Modify Cancer Chemotherapy Response and Survival
4.1. MiRNA Target Site Polymorphisms
4.2. Polymorphisms in miRNA Biogenesis Gene
4.3. CNV-miRNA: Possible Causative Variations Affecting Drug Response
5. MiRNAs and Ethnic Difference in Drug Response
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Baer-Dubowska, W.; Majchrzak-Celinska, A.; Cichocki, M. Pharmocoepigenetics: A new approach to predicting individual drug responses and targeting new drugs. Pharmacol. Rep. 2011, 63, 293–304. [Google Scholar] [CrossRef]
- Nakajima, M.; Yokoi, T. MicroRNAs from biology to future pharmacotherapy: Regulation of cytochrome P450s and nuclear receptors. Pharmacol. Ther. 2011, 131, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Cascorbi, I. Overlapping effects of genetic variation and epigenetics on drug response: Challenges of pharmacoepigenomics. Pharmacogenomics 2013, 14, 1807–1809. [Google Scholar] [CrossRef] [PubMed]
- Rukov, J.L.; Shomron, N. MicroRNA pharmacogenomics: Post-transcriptional regulation of drug response. Trends Mol. Med. 2011, 17, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Fromm, B.; Billipp, T.; Peck, L.E.; Johansen, M.; Tarver, J.E.; King, B.L.; Newcomb, J.M.; Sempere, L.F.; Flatmark, K.; Hovig, E.; et al. A Uniform System for the Annotation of Vertebrate microRNA Genes and the Evolution of the Human microRNAome. Annu. Rev. Genet. 2015, 49, 213–242. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-coding RNAs in development and disease: Background, mechanisms, and therapeutic approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chevillet, J.R.; Liu, G.; Kim, T.K.; Wang, K. The effects of microRNA on the absorption, distribution, metabolism and excretion of drugs. Br. J. Pharmacol. 2015, 172, 2733–2747. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Oberg, A.L.; Asmann, Y.W.; Sicotte, H.; McDonnell, S.K.; Riska, S.M.; Liu, W.; Steer, C.J.; Subramanian, S.; Cunningham, J.M.; et al. Genome-wide transcriptional profiling reveals microRNA-correlated genes and biological processes in human lymphoblastoid cell lines. PLoS ONE 2009, 4, e5878. [Google Scholar] [CrossRef] [PubMed]
- Rieger, J.K.; Klein, K.; Winter, S.; Zanger, U.M. Expression variability of absorption, distribution, metabolism, excretion-related microRNAs in human liver: Influence of nongenetic factors and association with gene expression. Drug Metab. Dispos. 2013, 41, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.M.; An, S.; Lee, E.M.; Kim, K.; Choi, S.J.; Kim, J.S.; Jang, H.H.; An, I.S.; Bae, S. CYP1A1 is a target of miR-892a-mediated post-transcriptional repression. Int. J. Oncol. 2012, 41, 331–336. [Google Scholar] [PubMed]
- Wang, H.; Tan, W.; Hao, B.; Miao, X.; Zhou, G.; He, F.; Lin, D. Substantial reduction in risk of lung adenocarcinoma associated with genetic polymorphism in CYP2A13, the most active cytochrome P450 for the metabolic activation of tobacco-specific carcinogen NNK. Cancer Res. 2003, 63, 8057–8061. [Google Scholar] [PubMed]
- Kalscheuer, S.; Zhang, X.; Zeng, Y.; Upadhyaya, P. Differential expression of microRNAs in early-stage neoplastic transformation in the lungs of F344 rats chronically treated with the tobacco carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone. Carcinogenesis 2008, 29, 2394–2399. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, Y.; Nakajima, M.; Takagi, S.; Taniya, T.; Yokoi, T. MicroRNA regulates the expression of human cytochrome P450 1B1. Cancer Res. 2006, 66, 9090–9098. [Google Scholar] [CrossRef] [PubMed]
- Chang, I.; Mitsui, Y.; Fukuhara, S.; Gill, A.; Wong, D.K.; Yamamura, S.; Shahryari, V.; Tabatabai, Z.L.; Dahiya, R.; Shin, D.M.; et al. Loss of miR-200c up-regulates CYP1B1 and confers docetaxel resistance in renal cell carcinoma. Oncotarget 2015, 6, 7774–7787. [Google Scholar] [CrossRef] [PubMed]
- Devlin, A.H.; Thompson, P.; Robson, T.; McKeown, S.R. Cytochrome P450 1B1 mRNA untranslated regions interact to inhibit protein translation. Mol. Carcinogenes. 2010, 49, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Surapureddi, S.; Coulter, S.; Ferguson, S.S.; Goldstein, J.A. Human CYP2C8 is post-transcriptionally regulated by microRNAs 103 and 107 in human liver. Mol. Pharmacol. 2012, 82, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Green, B.; Marrone, A.; Guo, Y.; Kadlubar, S.; Lin, D.; Fuscoe, J.; Pogribny, I.; Ning, B. Suppression of CYP2C9 by microRNA hsa-miR-128-3p in human liver cells and association with hepatocellular carcinoma. Sci. Rep. 2015, 5, 8534. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Green, B.; Tolleson, W.H.; Jin, Y.; Mei, N.; Guo, Y.; Deng, H.; Pogribny, I.; Ning, B. MicroRNA hsa-miR-29a-3p modulates CYP2C19 in human liver cells. Biochem. Pharmacol. 2015, 98, 215–123. [Google Scholar] [CrossRef] [PubMed]
- Mohri, T.; Nakajima, M.; Fukami, T.; Takamiya, M.; Aoki, Y.; Yokoi, T. Human CYP2E1 is regulated by miR-378. Biochem. Pharmacol. 2010, 79, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Shukla, U.; Tumma, N.; Gratsch, T.; Dombkowski, A.; Novak, R.F. Insights into insulin-mediated regulation of CYP2E1: miR-132/-212 targeting of CYP2E1 and role of phosphatidylinositol 3-kinase, Akt (protein kinase B), mammalian target of rapamycin signaling in regulating miR-132/-212 and miR-122/-181a expression in primary cultured rat hepatocytes. Drug Metab. Dispos. 2013, 41, 1769–1777. [Google Scholar] [PubMed]
- Chen, F.; Chen, C.; Yang, S.; Gong, W.; Wang, Y.; Cianflone, K.; Tang, J.; Wang, D.W. Let-7b inhibits human cancer phenotype by targeting cytochrome P450 epoxygenase 2J2. PLoS ONE 2012, 7, e39197. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.Z.; Gao, W.; Yu, A.M. MicroRNAs regulate CYP3A4 expression via direct and indirect targeting. Drug Metab. Dispos. 2009, 37, 2112–2117. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Jiang, S.; Zhang, Y.; Wang, X.; Peng, X.; Meng, C.; Liu, Y.; Wang, H.; Guo, L.; Qin, S.; et al. The effect of microRNAs in the regulation of human CYP3A4: A systematic study using a mathematical model. Sci. Rep. 2014, 4, 4283. [Google Scholar] [CrossRef] [PubMed]
- Vuppalanchi, R.; Liang, T.; Goswami, C.P.; Nalamasu, R.; Li, L.; Jones, D.; Wei, R.; Liu, W.; Sarasani, V.; Janga, S.C.; et al. Relationship between differential hepatic microRNA expression and decreased hepatic cytochrome P450 3A activity in cirrhosis. PLoS ONE 2013, 8, e74471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Liu, Y.; Wei, Z.; Zhang, Y.; Zhang, L.; Jiang, S.; Xiong, Y.; Shen, L.; He, L.; Xing, Q.; et al. Hsa-miR-27a is involved in the regulation of CYP3A4 expression in human livers from Chinese Han population. Pharmacogenomics 2015, 16, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Song, K.H.; Li, T.; Owsley, E.; Chiang, J.Y. A putative role of micro RNA in regulation of cholesterol 7alpha-hydroxylase expression in human hepatocytes. J. Lipid Res. 2010, 51, 2223–2233. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Kim, S.H.; King, A.N.; Zhao, L.; Simpson, R.U.; Christensen, P.J.; Wang, Z.; Thomas, D.G.; Giordano, T.J.; Lin, L.; et al. CYP24A1 is an independent prognostic marker of survival in patients with lung adenocarcinoma. Clin. Cancer Res. 2011, 17, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Komagata, S.; Nakajima, M.; Takagi, S.; Mohri, T.; Taniya, T.; Yokoi, T. Human CYP24 catalyzing the inactivation of calcitriol is post-transcriptionally regulated by miR-125b. Mol. Pharmacol. 2009, 76, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Mohri, T.; Nakajima, M.; Takagi, S.; Komagata, S.; Yokoi, T. MicroRNA regulates human vitamin D receptor. Int. J. Cancer 2009, 125, 1328–1333. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Yu, A.M. ABC transporters in multidrug resistance and pharmacokinetics, and strategies for drug development. Curr. Pharm. Des. 2014, 20, 793–807. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.T.; Li, Z.L.; He, Z.X.; Qiu, J.X.; Zhou, S.F. Molecular mechanisms for tumour resistance to chemotherapy. Clin. Exp. Pharmacol. Physiol. 2016, 43, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Boyerinas, B.; Park, S.M.; Murmann, A.E.; Gwin, K.; Montag, A.G.; Zillhardt, M.; Hua, Y.J.; Lengyel, E.; Peter, M.E. Let-7 modulates acquired resistance of ovarian cancer to Taxanes via IMP-1-mediated stabilization of multidrug resistance 1. Int. J. Cancer 2012, 130, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wu, H.; Liu, X.; Evans, B.R.; Medina, D.J.; Liu, C.G.; Yang, J.M. Role of MicroRNA miR-27a and miR-451 in the regulation of MDR1/P-glycoprotein expression in human cancer cells. Biochem. Pharmacol. 2008, 76, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hu, S.; Wang, J.; Cai, J.; Xiao, L.; Yu, L.; Wang, Z. MiR-27a modulates MDR1/P-glycoprotein expression by targeting HIPK2 in human ovarian cancer cells. Gynecol. Oncol. 2010, 119, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Bitarte, N.; Bandres, E.; Boni, V.; Zarate, R.; Rodriguez, J.; Gonzalez-Huarriz, M.; Lopez, I.; Javier Sola, J.; Alonso, M.M.; Fortes, P.; et al. MicroRNA-451 is involved in the self-renewal, tumorigenicity, and chemoresistance of colorectal cancer stem cells. Stem Cell 2011, 29, 1661–1671. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Hazari, S.; Mehra, S.; Kaushal, D.; Moroz, K.; Dash, S. Increased expression of P-glycoprotein and doxorubicin chemoresistance of metastatic breast cancer is regulated by miR-298. Am. J. Pathol. 2012, 180, 2490–2503. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zheng, Z.M.; Li, X.N.; Li, Z.F.; Wang, Y.; Geng, Y.F.; Bai, L.; Zhang, X.B. MiR-223 modulates multidrug resistance via downregulation of ABCB1 in hepatocellular carcinoma cells. Exp. Biol. Med. 2013, 238, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Zhang, Z.; Liu, Z.; Feng, B.; Ren, G.; Li, K.; Zhou, L.; Sun, Y.; Li, M.; Zhou, J.; et al. MiR-508-5p regulates multidrug resistance of gastric cancer by targeting ABCB1 and ZNRD1. Oncogene 2014, 33, 3267–3276. [Google Scholar] [CrossRef] [PubMed]
- Ikemura, K.; Yamamoto, M.; Miyazaki, S.; Mizutani, H.; Iwamoto, T.; Okuda, M. MicroRNA-145 post-transcriptionally regulates the expression and function of P-glycoprotein in intestinal epithelial cells. Mol. Pharmacol. 2013, 83, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Zhong, Z.; Yang, L.; Wang, S.; Gong, Z. MicroRNA-31 inhibits cisplatin-induced apoptosis in non-small cell lung cancer cells by regulating the drug transporter ABCB9. Cancer Lett. 2014, 343, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.K.; Wang, Y.J.; Gupta, P.; Chen, Z.S. Multidrug Resistance Proteins (MRPs) and Cancer Therapy. AAPS J. 2015, 17, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wu, H.; Xia, J.; Li, Y.; Zhang, Y.; Huang, K.; Wagar, N.; Yoon, Y.; Cho, H.T.; Scala, S.; et al. Involvement of miR-326 in chemotherapy resistance of breast cancer through modulating expression of multidrug resistance-associated protein 1. Biochem. Pharmacol. 2010, 79, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Liu, Y.; Bai, Y.; Sun, Y.; Xiao, F.; Guo, Y. Gene expression profiling of drug-resistant small cell lung cancer cells by combining microRNA and cDNA expression analysis. Eur. J. Cancer 2010, 46, 1692–1702. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xia, F.; Ma, L.; Shan, J.; Shen, J.; Yang, Z.; Liu, J.; Cui, Y.; Bian, X.; Bie, P.; et al. MicroRNA-122 sensitizes HCC cancer cells to adriamycin and vincristine through modulating expression of MDR and inducing cell cycle arrest. Cancer Lett. 2011, 310, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.Z.; Zhou, A.; Hu, Z.; Yu, A.M. Small nucleolar RNA-derived microRNA hsa-miR-1291 modulates cellular drug disposition through direct targeting of ABC transporter ABCC1. Drug Metab. Dispos. 2013, 41, 1744–1751. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, X.; Huang, J.; Peng, J.; Guo, L. MiR-7 modulates chemoresistance of small cell lung cancer by repressing MRP1/ABCC1. Int. J. Exp. Pathol. 2015, 96, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Haenisch, S.; Laechelt, S.; Bruckmueller, H.; Werk, A.; Noack, A.; Bruhn, O.; Remmler, C.; Cascorbi, I. Down-regulation of ATP-binding cassette C2 protein expression in HepG2 cells after rifampicin treatment is mediated by microRNA-379. Mol. Pharmacol. 2011, 80, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Werk, A.N.; Bruckmueller, H.; Haenisch, S.; Cascorbi, I. Genetic variants may play an important role in mRNA-miRNA interaction: Evidence for haplotype-dependent downregulation of ABCC2 (MRP2) by miRNA-379. Pharmacogenet. Genom. 2014, 24, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Liang, X.; Shen, K.; Cui, D.; Zheng, Y.; Xu, J.; Fan, Z.; Qiu, Y.; Li, Q.; Ni, L.; et al. MiR-297 modulates multidrug resistance in human colorectal carcinoma by down-regulating MRP-2. Biochem. J. 2012, 446, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Zhan, M.; Qu, Q.; Wang, G.; Zhou, H. Let-7c sensitizes acquired cisplatin-resistant A549 cells by targeting ABCC2 and Bcl-XL. Die Pharm. 2013, 68, 955–961. [Google Scholar]
- Borel, F.; Han, R.; Visser, A.; Petry, H.; van Deventer, S.J.; Jansen, P.L.; Konstantinova, P.; Réseau Centre de Ressources Biologiques Foie (French Liver Biobanks Network), France. Adenosine triphosphate-binding cassette transporter genes up-regulation in untreated hepatocellular carcinoma is mediated by cellular microRNAs. Hepatology 2012, 55, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Markova, S.M.; Kroetz, D.L. ABCC4 is regulated by microRNA-124a and microRNA-506. Biochem. Pharmacol. 2014, 87, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yu, F.; Jiao, Y.; Feng, J.; Tang, W.; Yao, H.; Gong, C.; Chen, J.; Su, F.; Zhang, Y.; et al. Reduced miR-128 in breast tumor-initiating cells induces chemotherapeutic resistance via Bmi-1 and ABCC5. Clin. Cancer Res. 2011, 17, 7105–7115. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.W.; Polgar, O.; Deeken, J.; To, K.W.; Bates, S.E. ABCG2: Determining its relevance in clinical drug resistance. Cancer Metast. Rev. 2007, 26, 39–57. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Ongkeko, W.M. ABCG2: The key to chemoresistance in cancer stem cells? Expert Opin. Drug Metab. Toxicol. 2009, 5, 1529–1542. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.Z.; Morris, M.E.; Yu, A.M. MicroRNA-328 negatively regulates the expression of breast cancer resistance protein (BCRP/ABCG2) in human cancer cells. Mol. Pharmacol. 2009, 75, 1374–1379. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Li, Y.M.; Tao, B.B.; Lu, Y.C.; Hu, G.H.; Liu, H.M.; He, J.; Xu, Y.; Yu, H.Y. Downregulation of ABCG2 expression in glioblastoma cancer stem cells with miRNA-328 may decrease their chemoresistance. Med. Sci. Monit. 2010, 16, HY27–HY30. [Google Scholar] [PubMed]
- Xu, X.T.; Xu, Q.; Tong, J.L.; Zhu, M.M.; Nie, F.; Chen, X.; Xiao, S.D.; Ran, Z.H. MicroRNA expression profiling identifies miR-328 regulates cancer stem cell-like SP cells in colorectal cancer. Br. J. Cancer 2012, 106, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pan, Y.Z.; Seigel, G.M.; Hu, Z.H.; Huang, M.; Yu, A.M. Breast cancer resistance protein BCRP/ABCG2 regulatory microRNAs (hsa-miR-328, -519c and -520h) and their differential expression in stem-like ABCG2+ cancer cells. Biochem. Pharmacol. 2011, 81, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Turrini, E.; Haenisch, S.; Laechelt, S.; Diewock, T.; Bruhn, O.; Cascorbi, I. MicroRNA profiling in K-562 cells under imatinib treatment: Influence of miR-212 and miR-328 on ABCG2 expression. Pharmacogenet. Genom. 2012, 22, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Liao, R.; Sun, J.; Zhang, L.; Lou, G.; Chen, M.; Zhou, D.; Chen, Z.; Zhang, S. MicroRNAs play a role in the development of human hematopoietic stem cells. J. Cell. Biochem. 2008, 104, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xue, X.; Wei, J.; An, Y.; Yao, J.; Cai, H.; Wu, J.; Dai, C.; Qian, Z.; Xu, Z.; et al. Hsa-miR-520h downregulates ABCG2 in pancreatic cancer cells to inhibit migration, invasion, and side populations. Br. J. Cancer 2010, 103, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Zhao, L.; Ma, M.; Bai, X.; He, M.; Yan, Y.; Wang, Y.; Chen, Q.; Zhao, X.; Zhou, M.; et al. MiR-181a enhances drug sensitivity in mitoxantone-resistant breast cancer cells by targeting breast cancer resistance protein (BCRP/ABCG2). Breast Cancer Res. Treat. 2013, 139, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.T.; He, M.; Wang, Y.; Jiao, X.Y.; Zhao, L.; Bai, X.F.; Yu, Z.J.; Wu, H.Z.; Sun, M.L.; Song, Z.G.; et al. MiR-487a resensitizes mitoxantrone (MX)-resistant breast cancer cells (MCF-7/MX) to MX by targeting breast cancer resistance protein (BCRP/ABCG2). Cancer Lett. 2013, 339, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Mouillet-Richard, S.; Schneider, B.; Launay, J.M.; Kellermann, O. MiR-16 targets the serotonin transporter: A new facet for adaptive responses to antidepressants. Science 2010, 329, 1537–1541. [Google Scholar] [CrossRef] [PubMed]
- Tamarapu Parthasarathy, P.; Galam, L.; Huynh, B.; Yunus, A.; Abuelenen, T.; Castillo, A.; Kollongod Ramanathan, G.; Cox, R., Jr.; Kolliputi, N. MicroRNA 16 modulates epithelial sodium channel in human alveolar epithelial cells. Biochem. Biophys. Res. Commun. 2012, 426, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Moya, P.R.; Wendland, J.R.; Salemme, J.; Fried, R.L.; Murphy, D.L. MiR-15a and miR-16 regulate serotonin transporter expression in human placental and rat brain raphe cells. Int. J. Neuropsychopharmacol. 2013, 16, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Miko, E.; Margitai, Z.; Czimmerer, Z.; Varkonyi, I.; Dezso, B.; Lanyi, A.; Bacso, Z.; Scholtz, B. MiR-126 inhibits proliferation of small cell lung cancer cells by targeting SLC7A5. FEBS Lett. 2011, 585, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Drayton, R.M.; Dudziec, E.; Peter, S.; Bertz, S.; Hartmann, A.; Bryant, H.E.; Catto, J.W. Reduced expression of miRNA-27a modulates cisplatin resistance in bladder cancer by targeting the cystine/glutamate exchanger SLC7A11. Clin. Cancer Res. 2014, 20, 1990–2000. [Google Scholar] [CrossRef] [PubMed]
- Gillen, A.E.; Gosalia, N.; Leir, S.H.; Harris, A. MicroRNA regulation of expression of the cystic fibrosis transmembrane conductance regulator gene. Biochem. J. 2011, 438, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Dalmasso, G.; Nguyen, H.T.; Yan, Y.; Laroui, H.; Charania, M.A.; Obertone, T.S.; Sitaraman, S.V.; Merlin, D. MicroRNA-92b regulates expression of the oligopeptide transporter PepT1 in intestinal epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G52–G59. [Google Scholar] [CrossRef] [PubMed]
- Li, K.K.; Pang, J.C.; Ching, A.K.; Wong, C.K.; Kong, X.; Wang, Y.; Zhou, L.; Chen, Z.; Ng, H.K. MiR-124 is frequently down-regulated in medulloblastoma and is a negative regulator of SLC16A1. Hum. Pathol. 2009, 40, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Pullen, T.J.; da Silva Xavier, G.; Kelsey, G.; Rutter, G.A. MiR-29a and miR-29b contribute to pancreatic beta-cell-specific silencing of monocarboxylate transporter 1 (Mct1). Mol. Cell. Biol. 2011, 31, 3182–3194. [Google Scholar] [CrossRef] [PubMed]
- Kamiyama, Y.; Matsubara, T.; Yoshinari, K.; Nagata, K.; Kamimura, H.; Yamazoe, Y. Role of human hepatocyte nuclear factor 4alpha in the expression of drug-metabolizing enzymes and transporters in human hepatocytes assessed by use of small interfering RNA. Drug Metab. Pharm. 2007, 22, 287–298. [Google Scholar] [CrossRef]
- Takagi, S.; Nakajima, M.; Kida, K.; Yamaura, Y.; Fukami, T.; Yokoi, T. MicroRNAs regulate human hepatocyte nuclear factor 4alpha, modulating the expression of metabolic enzymes and cell cycle. J. Biol. Chem. 2010, 285, 4415–4422. [Google Scholar] [CrossRef] [PubMed]
- Hatziapostolou, M.; Polytarchou, C.; Aggelidou, E.; Drakaki, A.; Poultsides, G.A.; Jaeger, S.A.; Ogata, H.; Karin, M.; Struhl, K.; Hadzopoulou-Cladaras, M.; et al. An HNF4alpha-miRNA inflammatory feedback circuit regulates hepatocellular oncogenesis. Cell 2011, 147, 1233–1247. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, A.; Li, L.; Gaedigk, A.; Bradford, L.D.; Benson, E.A.; Flockhart, D.A.; Skaar, T.C. In silico and in vitro identification of microRNAs that regulate hepatic nuclear factor 4alpha expression. Drug Metab. Dispos. 2012, 40, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Burke, P.A. The role of microRNAs in hepatocyte nuclear factor-4alpha expression and transactivation. Biochim. Biophys. Acta 2013, 1829, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Takagi, S.; Nakajima, M.; Mohri, T.; Yokoi, T. Post-transcriptional regulation of human pregnane X receptor by micro-RNA affects the expression of cytochrome P450 3A4. J. Biol. Chem. 2008, 283, 9674–9680. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Chen, M.; Zhang, Y.; Wang, X.; Jiang, S.; Wang, Y.; Wu, X.; Qin, S.; He, L.; Zhang, L.; et al. No correlation of hsa-miR-148a with expression of PXR or CYP3A4 in human livers from Chinese Han population. PLoS ONE 2013, 8, e59141. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Schuetz, E.G.; Xu, Y.; Thummel, K.E. Interplay between vitamin D and the drug metabolizing enzyme CYP3A4. J. Steroid Biochem. Mol. Biol. 2013, 136, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Keenan, C.R.; Lew, M.J.; Stewart, A.G. Biased signalling from the glucocorticoid receptor: Renewed opportunity for tailoring glucocorticoid activity. Biochem. Pharmacol. 2016, 112, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Pavek, P.; Cerveny, L.; Svecova, L.; Brysch, M.; Libra, A.; Vrzal, R.; Nachtigal, P.; Staud, F.; Ulrichova, J.; Fendrich, Z.; et al. Examination of Glucocorticoid receptor alpha-mediated transcriptional regulation of P-glycoprotein, CYP3A4, and CYP2C9 genes in placental trophoblast cell lines. Placenta 2007, 28, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Vreugdenhil, E.; Verissimo, C.S.; Mariman, R.; Kamphorst, J.T.; Barbosa, J.S.; Zweers, T.; Champagne, D.L.; Schouten, T.; Meijer, O.C.; de Kloet, E.R.; et al. MicroRNA 18 and 124a down-regulate the glucocorticoid receptor: Implications for glucocorticoid responsiveness in the brain. Endocrinology 2009, 150, 2220–2228. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.D.; Furneaux, H.; White, B.A. The micro-ribonucleic acid (miRNA) miR-206 targets the human estrogen receptor-alpha (ERalpha) and represses ERalpha messenger RNA and protein expression in breast cancer cell lines. Mol. Endocrinol. 2007, 21, 1132–1147. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Lin, J.; Yang, H.; Kong, W.; He, L.; Ma, X.; Coppola, D.; Cheng, J.Q. MicroRNA-221/222 negatively regulates estrogen receptor alpha and is associated with tamoxifen resistance in breast cancer. J. Biol. Chem. 2008, 283, 31079–31086. [Google Scholar] [CrossRef] [PubMed]
- Pandey, D.P.; Picard, D. MiR-22 inhibits estrogen signaling by directly targeting the estrogen receptor alpha mRNA. Mol. Cell. Biol. 2009, 29, 3783–3790. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Yu, D.; Wei, N.; Fu, H.; Cai, T.; Huang, Y.; Wu, C.; Zheng, X.; Du, Q.; Lin, D.; et al. An estrogen receptor alpha suppressor, microRNA-22, is downregulated in estrogen receptor alpha-positive human breast cancer cell lines and clinical samples. FEBS J. 2010, 277, 1684–1694. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Pu, Y.; Wong, D.K.; Liu, T.; Tang, H.; Xiang, T.; Yuen, M.F.; Ren, G. The hepatitis B virus-associated estrogen receptor alpha (ERalpha) was regulated by microRNA-130a in HepG2.2.15 human hepatocellular carcinoma cells. Acta Biochim. Biophys. Sin. 2011, 43, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Lv, G.C.; Sheng, J.; Yang, Y.D. Effect of miRNA-10b in regulating cellular steatosis level by targeting PPAR-alpha expression, a novel mechanism for the pathogenesis of NAFLD. J. Gastroenterol. Hepatol. 2010, 25, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Kida, K.; Nakajima, M.; Mohri, T.; Oda, Y.; Takagi, S.; Fukami, T.; Yokoi, T. PPARalpha is regulated by miR-21 and miR-27b in human liver. Pharm. Res. 2011, 28, 2467–2476. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.L.; Zhang, C.P.; Nie, F.; Xu, X.T.; Zhu, M.M.; Xiao, S.D.; Ran, Z.H. MicroRNA 506 regulates expression of PPAR alpha in hydroxycamptothecin-resistant human colon cancer cells. FEBS Lett. 2011, 585, 3560–3568. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z.; Wada, T.; Gramignoli, R.; Li, S.; Strom, S.C.; Huang, M.; Xie, W. MicroRNA hsa-miR-613 targets the human LXRalpha gene and mediates a feedback loop of LXRalpha autoregulation. Mol. Endocrinol. 2011, 25, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Zhang, Y.; Zeng, Y.J.; Gao, M.; Wu, G.Z.; Hu, C.J.; Huang, G.; He, F.T. MicroRNA-613 represses lipogenesis in HepG2 cells by downregulating LXRalpha. Lipids Health Dis. 2013, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Vinod, M.; Chennamsetty, I.; Colin, S.; Belloy, L.; De Paoli, F.; Schaider, H.; Graier, W.F.; Frank, S.; Kratky, D.; Staels, B.; et al. MiR-206 controls LXRalpha expression and promotes LXR-mediated cholesterol efflux in macrophages. Biochim. Biophys. Acta 2014, 1841, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.M. Role of microRNAs in the regulation of drug metabolism and disposition. Expert Opin. Drug Metab. Toxicol. 2009, 5, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.C.; Li, X.; Radecki, L.; Pan, Y.Z.; Winter, J.C.; Huang, M.; Yu, A.M. MicroRNA expression is differentially altered by xenobiotic drugs in different human cell lines. Biopharm. Drug Dispos. 2011, 32, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Pogribny, I.P.; Filkowski, J.N.; Tryndyak, V.P.; Golubov, A.; Shpyleva, S.I.; Kovalchuk, O. Alterations of microRNAs and their targets are associated with acquired resistance of MCF-7 breast cancer cells to cisplatin. Int. J. Cancer 2010, 127, 1785–1794. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tatsumi, N.; Fukami, T.; Yokoi, T.; Nakajima, M. Integrated analysis of rifampicin-induced microRNA and gene expression changes in human hepatocytes. Drug Metab. Pharm. 2014, 29, 333–340. [Google Scholar] [CrossRef]
- Benson, E.A.; Eadon, M.T.; Desta, Z.; Liu, Y.; Lin, H.; Burgess, K.S.; Segar, M.W.; Gaedigk, A.; Skaar, T.C. Rifampin Regulation of Drug Transporters Gene Expression and the Association of MicroRNAs in Human Hepatocytes. Front. Pharmacol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, F.; Li, T.; Lu, M.; Wang, L.; Yue, W.; Zhang, D. MirSNP, a database of polymorphisms altering miRNA target sites, identifies miRNA-related SNPs in GWAS SNPs and eQTLs. BMC Genom. 2012, 13, 661. [Google Scholar] [CrossRef] [PubMed]
- Dzikiewicz-Krawczyk, A. MicroRNA polymorphisms as markers of risk, prognosis and treatment response in hematological malignancies. Crit. Rev. Oncol. Hematol. 2015, 93, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2012. CA Cancer J. Clin. 2012, 62, 10–29. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Li, R.; Peng, J.; Li, S.; Guo, Z. A polymorphism at the miR-502 binding site in the 3’ untranslated region of the SET8 gene is associated with the outcome of small-cell lung cancer. Exp. Ther. Med. 2012, 3, 689–692. [Google Scholar] [PubMed]
- Xu, J.; Yin, Z.; Gao, W.; Liu, L.; Yin, Y.; Liu, P.; Shu, Y. Genetic variation in a microRNA-502 minding site in SET8 gene confers clinical outcome of non-small cell lung cancer in a Chinese population. PLoS ONE 2013, 8, e77024. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Yang, L.; Yang, R.; Yang, X.; Deng, J.; Yu, B.; Huang, D.; Zhang, S.; Wang, H.; Qiu, F.; et al. A microRNA-135a/b binding polymorphism in CD133 confers decreased risk and favorable prognosis of lung cancer in Chinese by reducing CD133 expression. Carcinogenesis 2013, 34, 2292–2299. [Google Scholar] [CrossRef] [PubMed]
- Campayo, M.; Navarro, A.; Vinolas, N.; Tejero, R.; Munoz, C.; Diaz, T.; Marrades, R.; Cabanas, M.L.; Gimferrer, J.M.; Gascon, P.; et al. A dual role for KRT81: A miR-SNP associated with recurrence in non-small-cell lung cancer and a novel marker of squamous cell lung carcinoma. PLoS ONE 2011, 6, e22509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, X.; Roth, J.A.; Hildebrandt, M.A.; Ye, Y.; Wei, H.; Minna, J.D.; Lippman, S.M.; Wu, X. MicroRNA-related genetic variants associated with clinical outcomes in early-stage non-small cell lung cancer patients. Cancer Res. 2013, 73, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zandi, R.; Gu, J.; Ye, Y.Q.; Pertsemlidis, A.; Wu, X.F.; Roth, J.A.; Ji, L. A SNP in the 3’-untranslated region of FZD4 linked to lung cancer survival modulates a miRNA-mediated FZD4 transcript binding, cleavage, expression, and Wnt-signaling in NSCLC cells. Cancer Res. 2015, 75, 196. [Google Scholar] [CrossRef]
- Graziano, F.; Canestrari, E.; Loupakis, F.; Ruzzo, A.; Galluccio, N.; Santini, D.; Rocchi, M.; Vincenzi, B.; Salvatore, L.; Cremolini, C.; et al. Genetic modulation of the Let-7 microRNA binding to KRAS 3’-untranslated region and survival of metastatic colorectal cancer patients treated with salvage cetuximab-irinotecan. Pharmacogenom. J. 2010, 10, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Sebio, A.; Pare, L.; Paez, D.; Salazar, J.; Gonzalez, A.; Sala, N.; del Rio, E.; Martin-Richard, M.; Tobena, M.; Barnadas, A.; et al. The LCS6 polymorphism in the binding site of let-7 microRNA to the KRAS 3’-untranslated region: Its role in the efficacy of anti-EGFR-based therapy in metastatic colorectal cancer patients. Pharmacogenet. Genom. 2013, 23, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Winder, T.; Ning, Y.; Pohl, A.; Yang, D.; Kahn, M.; Lurje, G.; Labonte, M.J.; Wilson, P.M.; Gordon, M.A.; et al. A let-7 microRNA-binding site polymorphism in 3’-untranslated region of KRAS gene predicts response in wild-type KRAS patients with metastatic colorectal cancer treated with cetuximab monotherapy. Ann. Oncol. 2011, 22, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Kjersem, J.B.; Ikdahl, T.; Guren, T.; Skovlund, E.; Sorbye, H.; Hamfjord, J.; Pfeiffer, P.; Glimelius, B.; Kersten, C.; Solvang, H.; et al. Let-7 miRNA-binding site polymorphism in the KRAS 3'UTR; colorectal cancer screening population prevalence and influence on clinical outcome in patients with metastatic colorectal cancer treated with 5-fluorouracil and oxaliplatin +/− cetuximab. BMC Cancer 2012, 12, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynendaele, J.; Bohnke, A.; Leucci, E.; Nielsen, S.J.; Lambertz, I.; Hammer, S.; Sbrzesny, N.; Kubitza, D.; Wolf, A.; Gradhand, E.; et al. An illegitimate microRNA target site within the 3’ UTR of MDM4 affects ovarian cancer progression and chemosensitivity. Cancer Res. 2010, 70, 9641–9649. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Meyer, L.; Chang, D.W.; Lin, J.; Pu, X.; Ye, Y.; Gu, J.; Wu, X.; Lu, K. Genetic variants in MicroRNA biosynthesis pathways and binding sites modify ovarian cancer risk, survival, and treatment response. Cancer Res. 2010, 70, 9765–9776. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wu, C.; Wang, X.; Wang, C.; Zhang, R.; Shan, B. A polymorphism at the miR-502 binding site in the 3’-untranslated region of the histone methyltransferase SET8 is associated with hepatocellular carcinoma outcome. Int. J. Cancer 2012, 131, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Munoz, C.; Gaya, A.; Diaz-Beya, M.; Gel, B.; Tejero, R.; Diaz, T.; Martinez, A.; Monzo, M. MiR-SNPs as markers of toxicity and clinical outcome in Hodgkin lymphoma patients. PLoS ONE 2013, 8, e64716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Larrea, C.F.; Navarro, A.; Tejero, R.; Tovar, N.; Diaz, T.; Cibeira, M.T.; Rosinol, L.; Ferrer, G.; Rovira, M.; Rozman, M.; et al. Impact of MiRSNPs on survival and progression in patients with multiple myeloma undergoing autologous stem cell transplantation. Clin. Cancer Res. 2012, 18, 3697–3704. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xiao, Y.; Ding, X.; Zhuo, Y.; Ren, P.; Zhou, C.; Zhou, J. A miR-200b/200c/429-binding site polymorphism in the 3’ untranslated region of the AP-2alpha gene is associated with cisplatin resistance. PLoS ONE 2011, 6, e29043. [Google Scholar]
- Bao, B.Y.; Pao, J.B.; Huang, C.N.; Pu, Y.S.; Chang, T.Y.; Lan, Y.H.; Lu, T.L.; Lee, H.Z.; Juang, S.H.; Chen, L.M.; et al. Polymorphisms inside microRNAs and microRNA target sites predict clinical outcomes in prostate cancer patients receiving androgen-deprivation therapy. Clin. Cancer Res. 2011, 17, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.P.; Levesque, E.; Guillemette, C.; Yu, C.C.; Huang, C.Y.; Lin, V.C.; Chung, I.C.; Chen, L.C.; Laverdiere, I.; Lacombe, L.; et al. Genetic variants in microRNAs and microRNA target sites predict biochemical recurrence after radical prostatectomy in localized prostate cancer. Int. J. Cancer 2014, 135, 2661–2667. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Wu, W.; Han, B.; Gao, G.; Qiao, R.; Lv, J.; Zhang, S.; Zhang, W.; Fan, W.; Chen, H.; et al. Hsa-miR-196a2 functional SNP is associated with severe toxicity after platinum-based chemotherapy of advanced nonsmall cell lung cancer patients in a Chinese population. J. Clin. Lab. Anal. 2012, 26, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.J.; Choi, Y.Y.; Jang, J.A.; Jung, H.J.; Lee, S.Y.; Lee, W.K.; Yoo, S.S.; Lee, J.; Cha, S.I.; Kim, C.H.; et al. Association between genetic variants in pre-microRNAs and survival of early-stage NSCLC. J. Thorac. Oncol. 2013, 8, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yin, Z.; Shen, H.; Gao, W.; Qian, Y.; Pei, D.; Liu, L.; Shu, Y. A genetic polymorphism in pre-miR-27a confers clinical outcome of non-small cell lung cancer in a Chinese population. PLoS ONE 2013, 8, e79135. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wei, Q.; Hu, L.; Chen, F.; Hu, Z.; Heist, R.S.; Su, L.; Amos, C.I.; Shen, H.; Christiani, D.C. Polymorphisms in MicroRNAs are associated with survival in non-small cell lung cancer. Cancer Epidemiol. Biomarkers Prev. 2014, 23, 2503–2511. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, H.; Li, Y.; Li, B.; Li, C.; Ding, C. A microRNA-related single nucleotide polymorphism of the XPO5 gene is associated with survival of small cell lung cancer patients. Biomed. Rep. 2013, 1, 545–548. [Google Scholar] [PubMed]
- Boni, V.; Zarate, R.; Villa, J.C.; Bandres, E.; Gomez, M.A.; Maiello, E.; Garcia-Foncillas, J.; Aranda, E. Role of primary miRNA polymorphic variants in metastatic colon cancer patients treated with 5-fluorouracil and irinotecan. Pharmacogenom. J. 2011, 11, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Gu, J.; Eng, C.; Ellis, L.M.; Hildebrandt, M.A.; Lin, J.; Huang, M.; Calin, G.A.; Wang, D.; Dubois, R.N.; et al. Genetic polymorphisms in MicroRNA-related genes as predictors of clinical outcomes in colorectal adenocarcinoma patients. Clin. Cancer Res. 2012, 18, 3982–3991. [Google Scholar] [CrossRef] [PubMed]
- Pardini, B.; Rosa, F.; Naccarati, A.; Vymetalkova, V.; Ye, Y.; Wu, X.; di Gaetano, C.; Buchler, T.; Novotny, J.; Matullo, G.; et al. Polymorphisms in microRNA genes as predictors of clinical outcomes in colorectal cancer patients. Carcinogenesis 2015, 36, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.L.; Chen, M.; Ye, Y.; Hildebrandt, M.A.; Wu, W.J.; Wei, H.; Huang, M.; Chang, D.W.; Dinney, C.P.; Wu, X. Genetic variations in micro-RNA biogenesis genes and clinical outcomes in non-muscle-invasive bladder cancer. Carcinogenesis 2013, 34, 1006–1011. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; An, J.; Lin, J.; Liu, Y.; Bao, L.; Zhang, W.; Zhao, J.J. Single nucleotide polymorphisms of microRNA processing machinery genes and outcome of hepatocellular carcinoma. PLoS ONE 2014, 9, e92791. [Google Scholar] [CrossRef] [PubMed]
- Henrichsen, C.N.; Chaignat, E.; Reymond, A. Copy number variants, diseases and gene expression. Hum. Mol. Genet. 2009, 18, R1–R8. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Mi, S.; Zhang, W.; Dolan, M.E. Comprehensive analysis of the impact of SNPs and CNVs on human microRNAs and their regulatory genes. RNA Biol. 2009, 6, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Zhang, X.; Tian, H.; Liang, N.; Wang, Y.; Liang, C.; Li, X.; Sun, F. Altered microRNA expression in patients with non-obstructive azoospermia. Reprod. Biol. Endocrinol. 2009, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Leidinger, P.; Lange, J.; Borries, A.; Schroers, H.; Scheffler, M.; Lenhof, H.P.; Ruprecht, K.; Meese, E. Multiple sclerosis: microRNA expression profiles accurately differentiate patients with relapsing-remitting disease from healthy controls. PLoS ONE 2009, 4, e7440. [Google Scholar] [CrossRef] [PubMed]
- Marrale, M.; Albanese, N.N.; Cali, F.; Romano, V. Assessing the impact of copy number variants on miRNA genes in autism by Monte Carlo simulation. PLoS ONE 2014, 9, e90947. [Google Scholar] [CrossRef] [PubMed]
- Marcinkowska, M.; Szymanski, M.; Krzyzosiak, W.J.; Kozlowski, P. Copy number variation of microRNA genes in the human genome. BMC Genom. 2011, 12, 183. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Zheng, Y. Comprehensive analysis of single nucleotide polymorphisms in human microRNAs. PLoS ONE 2013, 8, e78028. [Google Scholar] [CrossRef] [PubMed]
- Rawlings-Goss, R.A.; Campbell, M.C.; Tishkoff, S.A. Global population-specific variation in miRNA associated with cancer risk and clinical biomarkers. BMC Med. Genom. 2014, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Clark, A.G. Impact of microRNA regulation on variation in human gene expression. Genome Res. 2012, 22, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.S.; Gamazon, E.R.; Ziliak, D.; Wen, Y.; Im, H.K.; Zhang, W.; Wing, C.; Duan, S.; Bleibel, W.K.; Cox, N.J.; et al. Population differences in microRNA expression and biological implications. RNA Boil. 2011, 8, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Bovell, L.C.; Shanmugam, C.; Putcha, B.D.; Katkoori, V.R.; Zhang, B.; Bae, S.; Singh, K.P.; Grizzle, W.E.; Manne, U. The prognostic value of microRNAs varies with patient race/ethnicity and stage of colorectal cancer. Clin. Cancer Res. 2013, 19, 3955–3965. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Li, S.; Li, J.; Yin, L.; Zhou, T.; Zhang, C.; Chen, X.; Sun, K. Ethnic differences in microRNA-375 expression level and DNA methylation status in type 2 diabetes of Han and Kazak populations. J. Diabetes Res. 2014, 2014, 761938. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sundquist, J.; Zoller, B.; Memon, A.A.; Palmer, K.; Sundquist, K.; Bennet, L. Determination of 14 circulating microRNAs in Swedes and Iraqis with and without diabetes mellitus type 2. PLoS ONE 2014, 9, e86792. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Ji, P.; Ouyang, N.; Zhang, Y.; Wang, X.Y.; Rubin, D.C.; Davidson, N.O.; Bergamaschi, R.; Shroyer, K.R.; Burke, S.; et al. Differential expression of miRNAs in colon cancer between African and Caucasian Americans: Implications for cancer racial health disparities. Int. J. Oncol. 2014, 45, 587–594. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Function | Gene | Involved miRNA | Identification Methods | References |
|---|---|---|---|---|
| DMEs | CYP1A1 | miR-18b, -20b | Correlation, mRNA expression | [9] |
| miR-892a | Reporter assay, mRNA/protein expression, functional assay | [11] | ||
| CYP2A3 | miR-126* a | mRNA/protein expression | [13] | |
| CYP1B1 | miR-27b | Correlation, Reporter assays, protein expression, functional assay | [14] | |
| CYP2C8 | miR-103, -107 | Reporter assay, mRNA/protein expression, correlation | [17] | |
| CYP2C9 | miR-128-3p | EMSA, reporter assay, mRNA/protein expression, correlation | [18] | |
| CYP2C19 | miR-103, -107 | Protein expression | [17] | |
| miR-29a-3p | EMSA, mRNA/protein expression, correlation | [19] | ||
| CYP2E1 | miR-378 | Reporter assay, mRNA/protein expression, correlation, functional assay | [20] | |
| miR-132, -212 | Reporter assay, mRNA expression | [21] | ||
| CYP2J2 | let-7b | Reporter assay, protein expression, functional assay | [22] | |
| CYP3A4 | miR-27b | Reporter assay, mRNA/protein expression, functional assay | [23] | |
| miR-1, -532-3p, -577, -627 | Reporter assay, protein expression, correlation | [24] | ||
| miR-27a | Reporter assay, mRNA/protein expression, correlation | [26] | ||
| CYP7A1 | miR-122a, -422a a | Reporter assay, mRNA expression | [27] | |
| CYP24A1 | miR-125b | Reporter assay, mRNA/protein expression, functional assay | [29] | |
| Transporters | ABCB1/MDR1 | let-7g | mRNA/protein expression, correlation, functional assay | [33] |
| miR-27a | mRNA/protein expression, functional assay | [34,35] | ||
| miR-451 a | mRNA/protein expression, functional assay | [34,36] | ||
| miR-298 a | Reporter assay, protein expression, functional assay | [37] | ||
| miR-223 | Reporter assay, mRNA/protein expression, functional assay | [38] | ||
| miR-508-5p | Reporter assay, mRNA/protein expression, functional assay | [39] | ||
| miR-145 | Reporter assay, protein expression, functional assay | [40] | ||
| ABCB9 | miR-31 | Reporter assay, mRNA/protein expression, functional assay | [41] | |
| ABCC1/MRP1 | miR-326 | Reporter assay, mRNA/protein expression, functional assays | [43] | |
| miR-134 | mRNA/protein expression | [44] | ||
| miR-122 | mRNA/protein expression, functional assay | [45] | ||
| miR-1291 a | Reporter assay, mRNA/protein expression, and functional assays | [46] | ||
| miR-7 | Reporter assay, mRNA/protein expression, correlation | [47] | ||
| ABCC2/MRP2 | miR-379 | Reporter assay, mRNA/protein expression, functional assays | [48,49] | |
| miR-297 a | Reporter assay, mRNA/protein expression, functional assays | [50] | ||
| let-7c | Reporter assay, mRNA/protein expression, functional assays | [51] | ||
| ABCC4/MRP4 | miR-125a/b | Reporter assay, mRNA expression, correlation | [52] | |
| miR-124a, -506 a | Reporter assay, mRNA/protein expression, correlation, functional assay | [53] | ||
| ABCC5/MRP5 | miR-128 | Reporter assay, protein expression, functional assay | [54] | |
| ABCG2/BCRP | miR-328 | Reporter assay, mRNA/protein expression, functional assays | [57,58,59] | |
| miR-519c | Reporter assay, mRNA/protein expression, functional assays | [60] | ||
| miR-212 | Reporter assay, protein expression | [61] | ||
| miR-520h | Reporter assay, mRNA/protein expression | [62,63] | ||
| miR-181a | Reporter assay, mRNA/protein expression, functional assays | [64] | ||
| miR-487a | Reporter assay, mRNA/protein expression, functional assays | [65] | ||
| SLC6A4/SERT | miR-16 | Reporter assay, mRNA/protein expression, functional assays | [66,67,68] | |
| miR-15a | Reporter assay, protein expression | [68] | ||
| SLC7A5/LAT1 | miR-126 | Reporter assay, mRNA/protein expression, correlation | [69] | |
| SLC7A11 | miR-27a | Reporter assay, mRNA/protein expression, functional assays | [70] | |
| SLC12A2 | miR-384 a, -494, -1246 a | Reporter assay, mRNA expression | [71] | |
| SLC15A1/PEPT1 | miR-92b | Reporter assay, mRNA/protein expression, correlation, functional assays | [72] | |
| SLC16A1/MCT1 | miR-124 | Reporter assay, mRNA/protein expression, functional assays | [73,74] | |
| Nuclear Receptor | HNF4α | miR-24, -34a | Reporter assay, mRNA/protein expression, functional assays | [76,77] |
| miR-34a, -34c-5p, -449a | Reporter assay, mRNA/protein expression, functional assays | [78,79] | ||
| PXR | miR-148a | Reporter assay, protein expression | [80] | |
| VDR | miR-27b | Reporter assay, mRNA/protein expression | [23] | |
| miR-125b | Reporter assay, EMSA, protein expression | [30] | ||
| GR | miR-18, -124a | Reporter assay, protein expression, functional assays | [85] | |
| ESR1 | miR-206 | Reporter assay, mRNA/protein expression | [86] | |
| miR-221, -222 | Reporter assay, protein expression, functional assays | [87] | ||
| miR-22 | Reporter assay, mRNA/protein expression | [88,89] | ||
| miR-130a | Reporter assay, mRNA/protein expression | [90] | ||
| PPARα | miR-10b, -21, -27b | Reporter assay, mRNA/protein expression, correlation | [91,92] | |
| miR-506 | Reporter assay, mRNA/protein expression, functional assays | [93] | ||
| LXRα | miR-613 a | EMSA, reporter assay, mRNA/protein expression, functional assays | [94,95] | |
| miR-206 | Reporter assay, mRNA/protein expression, functional assays | [96] |
| Gene | MirSNPs | Involved miRNAs | Cancer Types | Association with Variant Allele | References |
|---|---|---|---|---|---|
| SET8 | rs16917496 T > C | miR-502 | SCLC | Increased OS | [105] |
| NSCLC | Increased OS and reduced risk of death | [106] | |||
| CD133 | rs2240688 A > C | miR-135a/b | LC | Increased OS | [107] |
| KRT81 | rs3660 C > G | miR-17 | NSCLC | Increased time to recurrence | [108] |
| FAS | rs2234978 A > G | miR-651 | NSCLC | Reduced OS | [109] |
| FZD4 | rs713065 A > G | miR-204 | NSCLC | Increased OS | [109,110] |
| KRAS | rs61764370 T > G | let-7 | CRC | Poor response to cetuximab-irinotecan therapy and reduced OS/PFS | [111] |
| CRC | Resistance to anti-EGFR-based therapy | [112] | |||
| CRC | Improved OS/PFS | [113,114] | |||
| MDM4 | rs4245739 C > A | miR-191 | OC | Delayed progression and tumor related death | [115] |
| rs10900596 G > A | miR-409-3p a | OC | Better treatment response | [116] | |
| PDGFC | rs1425486 G > A | miR-425 | OC | Poor treatment response and reduced OS | [116] |
| KRAS | rs10771184 T > A | miR-544 | OC | Better treatment response and increased OS | [116] |
| SET8 | rs16917496 T > C | miR-502 | HCC | Increased OS | [117] |
| KRT81 | rs3660 C > G | miR-17 | HL | Increased risk of neurological toxicity | [118] |
| KRT81 | rs3660 C > G | miR-17 | MM | Increased OS | [119] |
| AP-2α | rs1045385 A > C | miR-200b, -200c, -429 | BC | Increased cisplatin sensitivity | [120] |
| CDON | rs3737336 T > C | miR-181c, -5007 a | PC | Recreased PFS | [121,122] |
| miR-196a-2 | rs11614913 C > T | NSCLC | Decreased risk of overall toxicity and increased OS/DFS | [123,124] | |
| miR-149 | rs2292832 T > C | NSCLC | Increased OS/DFS | [124] | |
| pre-miR-27a | rs895819 A > G | NSCLC | Poor treatment response and reduced OS | [125] | |
| miR-5197 a | rs2042253 T > C | NSCLC | Increased OS | [126] | |
| DROSHA | rs6886834 G > A | NSCLC | Reduced RFS | [109] | |
| XPO5 | rs11077 A > C | NSCLC | Increased time to recurrence | [108] | |
| SCLC | Reduced OS | [127] | |||
| pri-miR-26a-1 | rs7372209 C > T | CRC | Reduced time to progression | [128] | |
| pri-miR-100 | rs1834306 T > C | CRC | Increased time to progression | [128] | |
| miR-219-1 | rs213210 T > C | CRC | Increased RFS | [129] | |
| CRC | Increased OS/RFS | [130] | |||
| miR-608 | rs4919510 C > G | CRC | Decreased RFS | [129] | |
| CRC | Increased EFS | [130] | |||
| KIF3C | rs6728684 T > G | PC | Reduced PFS | [124] | |
| IFI30 | rs1045747 T > C | PC | Reduced PFS | [121] | |
| DDX20 | rs197412 C > T | BC | Decreased risk of recurrence | [131] | |
| DGCR8 | rs2073778 G > T | BC | Increased risk of progression | [131] | |
| XPO5 | rs11077 A > C | MM | Increased OS | [119] | |
| HL | Increased OS/DFS for heterozygotes | [118] | |||
| HCC | Increased OS | [132] | |||
| RNASEN | rs17408716 A > G | OC | Better treatment response and increased OS | [116] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.-P.; Hu, Y.-D.; Hu, X.-L.; Zhang, Y.-J.; Yang, Y.-L.; Jiang, C.; Tang, J.; Chen, X.-P. MiRNAs and miRNA Polymorphisms Modify Drug Response. Int. J. Environ. Res. Public Health 2016, 13, 1096. https://doi.org/10.3390/ijerph13111096
Li M-P, Hu Y-D, Hu X-L, Zhang Y-J, Yang Y-L, Jiang C, Tang J, Chen X-P. MiRNAs and miRNA Polymorphisms Modify Drug Response. International Journal of Environmental Research and Public Health. 2016; 13(11):1096. https://doi.org/10.3390/ijerph13111096
Chicago/Turabian StyleLi, Mu-Peng, Yao-Dong Hu, Xiao-Lei Hu, Yan-Jiao Zhang, Yong-Long Yang, Chun Jiang, Jie Tang, and Xiao-Ping Chen. 2016. "MiRNAs and miRNA Polymorphisms Modify Drug Response" International Journal of Environmental Research and Public Health 13, no. 11: 1096. https://doi.org/10.3390/ijerph13111096