
Effects of Ambient Fine Particles PM2.5 on Human HaCaT Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- The mechanism linking skin damage and environmental factors may have the following aspects.
- Oxidative damage: PM may damage skin through oxidative stress production, which is one of the important causes of skin aging [10].
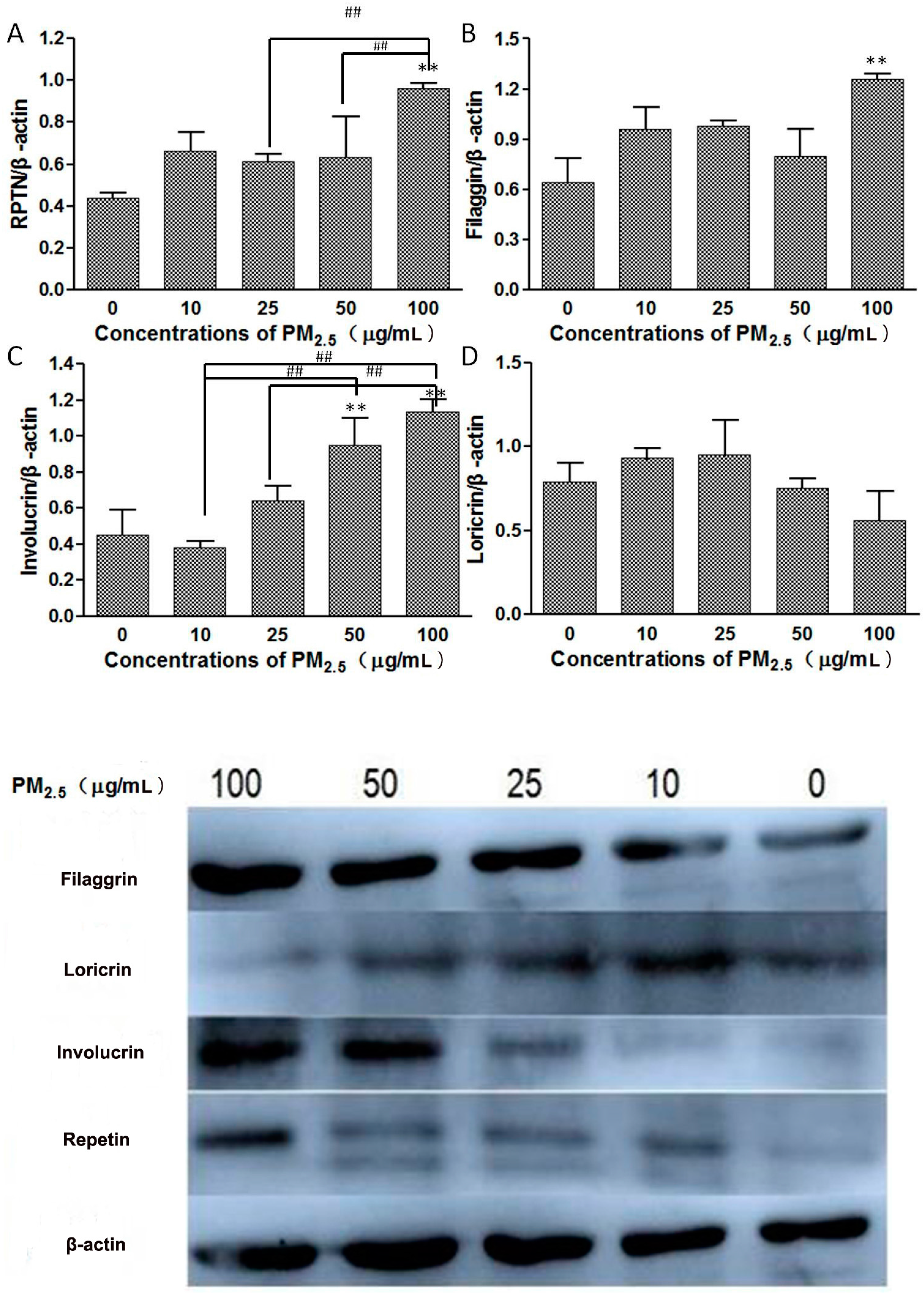
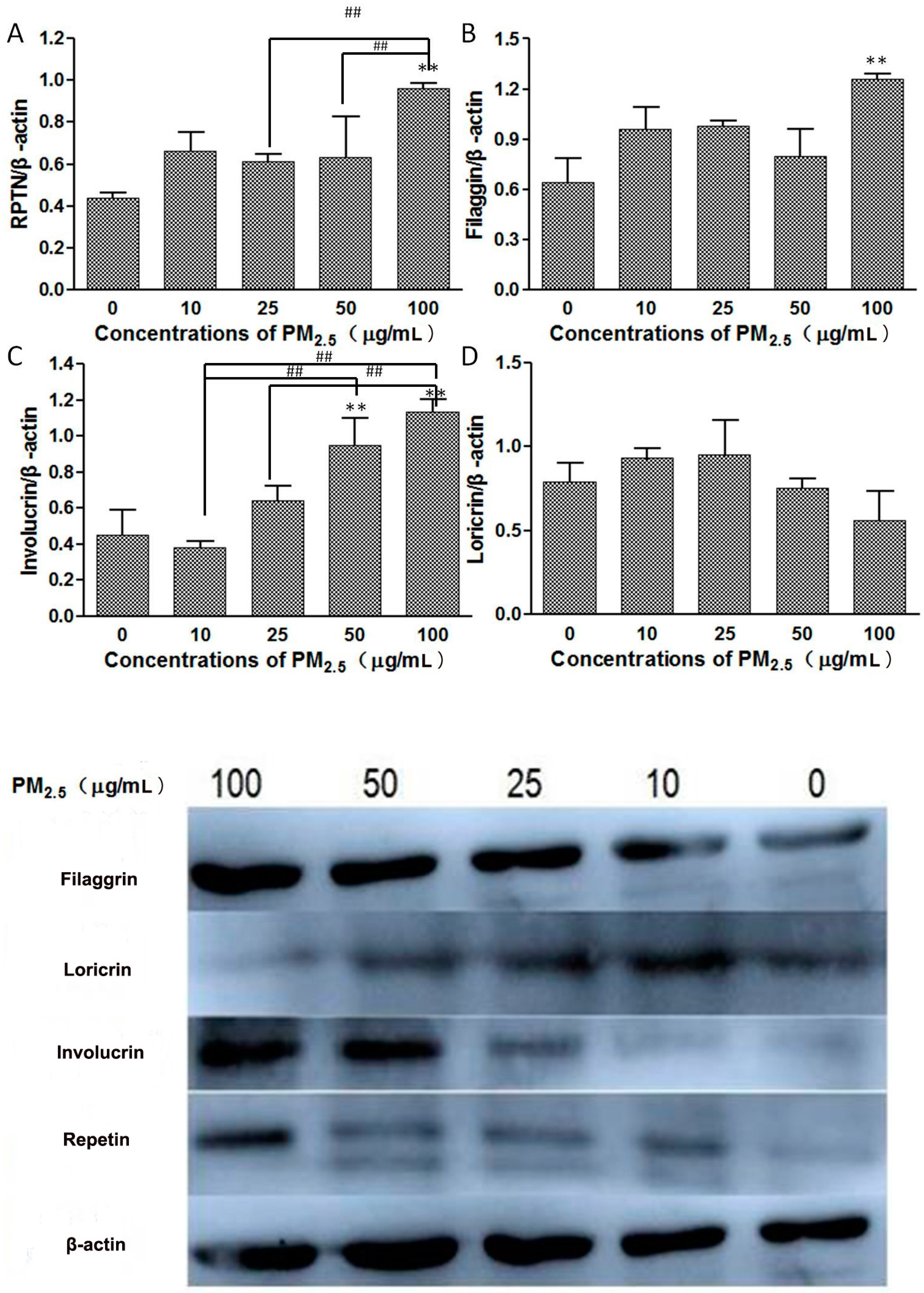
- Skin barrier function impairment: In the process of keeping skin barrier integrity, several important structural proteins play an important role in the process of skin cell differentiation, such as Loricrin (LOR), Involucrin (IVL), and Filaggrin (FLG) [11].
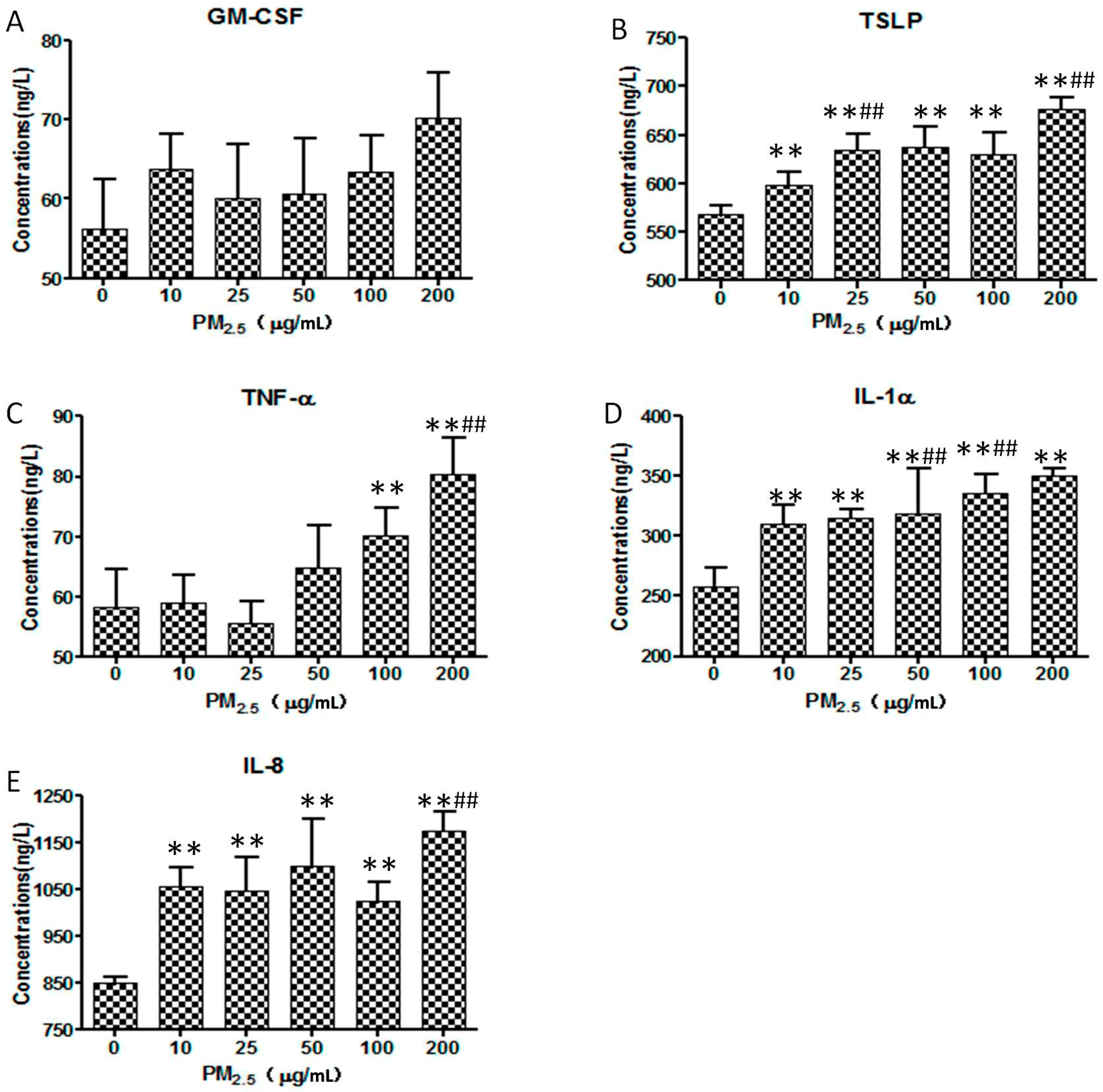
- Immune injury: Airborne particulate matter may increase keratinocyte inflammatory mediators [12] and cause cutaneous injury in cellular or humoral through immunity and innate immunity pathways. Increasing evidence has shown that the immune injury may play a role in the pathogenesis of various cutaneous inflammatory diseases by releasing cytokines or by expressing membrane-associated adhesion molecules [13,14].
2. Materials and Methods
2.1. PM2.5 Sampling
2.2. Cell Culture
2.3. PM2.5 Treatment
2.4. Cell Viability Determination
2.5. Western Blot
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Statistical Analyses
3. Results
3.1. The Morphology of HaCaT Cells
3.2. Cell Viability Determination
3.2.1. Relationship between PM2.5 Concentration and Cell Viability
3.2.2. Relationship between Exposure Time and Cell Viability
3.3. Western Blot
3.4. ELISA
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ministry of Environmental Protection of the P.R. China; General Administration of Quality Supervision, Inspection and Quarantine of the P.R. China. Ambient Air Quality Standards; GB 3095-2012; Standard Press of China: Beijing, China, 2012.
- Pope, C.A., III; Burnett, R.T.; Thun, M.J.; Calle, E.E.; Krewski, D.; Ito, K.; Thurston, G.D. Lung cancer, cardiopulmonary mortality, and long-term exposure to fine particulate air pollution. JAMA 2002, 287, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.J.; Kim, B.; Lee, K. Air pollution exposure and cardiovascular disease. Toxicol. Res. 2014, 30, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Xue, C.H.; Hwang, S.K.; Li, W.H.; Chen, Z.; Zhang, J.Z. Exposure to fine particulate matter associated with senile lentigo in Chinese women: A cross-sectional study. J. Eur. Acad. Dermatol. Venereol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liang, W.L.; Zhang, Y.; Sun, L.D.; Liang, W.Y. Effect of atmospheric fine particles on epidermal growth factor receptor mRNA expression in mouse skin tissue. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K. The role of air pollutants in atopic dermatitis. J. Allergy Clin. Immunol. 2014, 134, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J.; Liu, W.; Li, L.; Pan, X.; Crawford, M.; Sore, G.; Seite, S. Pollution and skin: From epidemiological and mechanistic studies to clinical implications. J. Dermatol. Sci. 2014, 76, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Lee, K.; Lee, Y.M.; Lee, J.H.; Lee, S.I.; Yu, S.D.; Paek, D. Acute health effects of urban fine and ultrafine particles on children with atopic dermatitis. Environ. Res. 2011, 111, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Vierkötter, A.; Schikowski, T.; Ranft, U.; Sugiri, D.; Matsui, M.; Krämer, U.; Krutmann, J. Airborne particle exposure and extrinsic skin aging. J. Investig. Dermatol. 2010, 130, 2719–2726. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.C.; Lee, C.W.; Tsai, M.H.; Ko, H.H.; Fang, J.Y.; Chiang, Y.C.; Liang, C.J.; Hsu, L.F.; Hu, S.C.; Yen, F.L. Eupafolin nanoparticles protect HaCaT keratinocytes from particulate matter-induced inflammation and oxidative stress. Int. J. Nanomed. 2016, 11, 3907–3926. [Google Scholar]
- Cork, M.J.; Danby, S.G.; Vasilopoulos, Y.; Hadgraft, J.; Lane, M.E.; Moustafa, M.; Guy, R.H.; MacGowan, A.L.; Tazi-Ahnini, R.; Ward, S.J. Epidermal barrier dysfunction in atopic dermatitis. J. Investig. Dermatol. 2009, 129, 1892–1908. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Shin, D.W.; Kim, W.; Doh, S.J.; Lee, S.H.; Noh, M. Asian dust storm particles induce a broad toxicological transcriptional program in human epidermal keratinocytes. Toxicol. Lett. 2011, 200, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.N.; Jones, M.L.; Mitra, R.S.; Crockett-Torabe, E.; Fantone, J.C.; Kunkel, S.L.; Warren, J.S.; Dixit, V.M.; Nickoloff, B.J. Modulation of keratinocyte-derived interleukin-8 which is chemotactic for neutrophils and T lymphocytes. Am. J. Pathol. 1991, 139, 869–876. [Google Scholar] [PubMed]
- Peguet-Navarro, J.; Dalbiez-Gauthier, C.; Moulon, C.; Berthier, O.; Réano, A.; Gaucherand, M.; Banchereau, J.; Rousset, F.; Schmitt, D. CD40 ligation of human keratinocytes inhibits their proliferation and induces their differentiation. J. Immunol. 1997, 158, 144–152. [Google Scholar] [PubMed]
- Jensen, J.M.; Fölster-Holst, R.; Baranowsky, A.; Schunck, M.; Winoto-Morbach, S.; Neumann, C.; Schütze, S.; Proksch, E. Impaired sphingomyelinase activity and epidermal differentiation in atopic dermatitis. J. Investig. Dermatol. 2004, 122, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Crish, J.F.; Gopalakrishnan, R.; Bone, F.; Gilliam, A.C.; Eckert, R.L. The distal and proximal regulatory regions of the involucrin gene promoter have distinct functions and are required for in vivo involucrin expression. J. Investig. Dermatol. 2006, 126, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Seguchi, T.; Cui, C.-Y.; Kusuda, S.; Takahashi, M.; Aisu, K.; Tezuka, T. Decreased expression of filaggrin in atopic skin. Arch. Dermatol. Res. 1996, 288, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, H.; Ebise, H.; Tazawa, T.; Tanaka, K.; Sugiura, Y.; Uehara, M.; Kikuchi, K.; Kimura, T. Large-scale DNA microarray analysis of atopic skin lesions shows overexpression of an epidermal differentiation gene cluster in the alternative pathway and lack of protective gene expression in the cornified envelope. Br. J. Dermatol. 2005, 152, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Hirao, T.; Terui, T.; Takeuchi, I.; Kobayashi, H.; Okada, M.; Takahashi, M.; Tagami, H. Ratio of immature cornified envelopes does not correlate with parakeratosis in inflammatory skin disorders. Exp. Dermatol. 2003, 12, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Jarzab, J.; Filipowska, B.; Zebracka, J.; Kowalska, M.; Bozek, A.; Rachowska, R.; Gubala, E.; Grzanka, A.; Hadas, E.; Jarzab, B. Locus 1q21 Gene expression changes in atopic dermatitis skin lesions: Deregulation of small proline-rich region 1A. Int. Arch. Allergy Immunol. 2010, 151, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Hazlewood, G.; Bernatsky, S.; Kaplan, G.G.; Eksteen, B.; Barnabe, C. Association between Air Pollution and the Development of Rheumatic Disease: A Systematic Review. Int. J. Rheumatol. 2016, 2016, 5356307. [Google Scholar] [CrossRef] [PubMed]
- Friend, S.L.; Hosier, S.; Nelson, A.; Foxworthe, D.; Williams, D.E.; Farr, A. A thymic stromal cell line supports in vitro development of surface IgM+ B cells and produces a novel growth factor affecting B and T lineage cells. Exp. Hematol. 1994, 22, 321–328. [Google Scholar] [PubMed]
- Li, M.; Messaddeq, N.; Teletin, M.; Pasquali, J.L.; Metzger, D.; Chambon, P. Retinoid X receptor ablation in adult mouse keratinocytes generates an atopic dermatitis triggered by thymic stromal lymphopoietin. Proc. Natl. Acad. Sci. USA 2005, 102, 14795–14800. [Google Scholar] [PubMed]
- Komine, M. Analysis of the mechanism for the development of allergic skin inflammation and the application for its treatment: Keratinocytes in atopic dermatitis—Their pathogenic involvement. J. Pharmacol. Sci. 2009, 110, 260–264. [Google Scholar] [PubMed]
- Lee, K.H.; Cho, K.A.; Kim, J.Y.; Kim, J.Y.; Baek, J.H.; Woo, S.Y.; Kim, J.W. Filaggrin knockdown and Toll-like receptor 3 (TLR3) stimulation enhanced the production of thymic stromal lymphopoietin (TSLP) from epidermal layers. Exp. Dermatol. 2011, 20, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Dumortier, A.; Durham, A.D.; Di Piazza, M.; Vauclair, S.; Koch, U.; Ferrand, G.; Ferrero, I.; Demehri, S.; Song, L.L.; Farr, A.G.; et al. Atopic dermatitis-like disease and associated lethal myeloproliferative disorder arise from loss of Notch signaling in the murine skin. PLoS ONE 2010, 5, e9258. [Google Scholar]
- Wang, Z.; Feng, A.; Cui, M.; Liu, Y.; Wang, L.; Wang, Q. First discovery and stucture-activity relationship study of phenanthroquinolizidines as novel antiviral agents against tobacco mosaic virus (TMV). PLoS ONE 2012, 7, e52933. [Google Scholar]
- Quay, J.L.; Reed, W.; Samet, J.; Devlin, R.B. Air pollution particles induce IL-6 gene expression in human airway epithelial cells via NF-κB activation. Am. J. Respir. Cell Mol. Biol. 1998, 19, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, H.; Ohtoshi, T.; Kawasaki, S.; Kohyama, T.; Desaki, M.; Kasama, T.; Kobayashi, K.; Nakahara, K.; Yamamoto, K.; Matsushima, K.; et al. Diesel exhaust particles induce NF-κB activation in human bronchial epithelial cells in vitro: Importance in cytokine transcription. J. Immunol. 1999, 162, 4705–4711. [Google Scholar] [PubMed]
- Martinez, V.; Corsini, E.; Mitjans, M.; Pinazo, A.; Vinardell, M.P. Evaluation of eye and skin irritation of arginine-derivative surfactants using different in vitro endpoints as alternatives to the in vivo assays. Toxicol. Lett. 2006, 164, 259–267. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Kang, Z.; Jiang, S.; Zhao, J.; Yan, S.; Xu, F.; Xu, J. Effects of Ambient Fine Particles PM2.5 on Human HaCaT Cells. Int. J. Environ. Res. Public Health 2017, 14, 72. https://doi.org/10.3390/ijerph14010072
Li Q, Kang Z, Jiang S, Zhao J, Yan S, Xu F, Xu J. Effects of Ambient Fine Particles PM2.5 on Human HaCaT Cells. International Journal of Environmental Research and Public Health. 2017; 14(1):72. https://doi.org/10.3390/ijerph14010072
Chicago/Turabian StyleLi, Qiao, Zhihua Kang, Shuo Jiang, Jinzhuo Zhao, Shuxian Yan, Feng Xu, and Jinhua Xu. 2017. "Effects of Ambient Fine Particles PM2.5 on Human HaCaT Cells" International Journal of Environmental Research and Public Health 14, no. 1: 72. https://doi.org/10.3390/ijerph14010072