LRBA Gene Polymorphisms and Risk of Coal Workers’ Pneumoconiosis: A Case–Control Study from China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. SNP Selection
2.3. Genotyping
2.4. Bioinformatical Analysis and Gene Expression Levels
2.5. Statistical Analysis
3. Results
3.1. Characteristics of the Study Subjects
3.2. Associations between the LRBA Polymorphisms and Risk of CWP
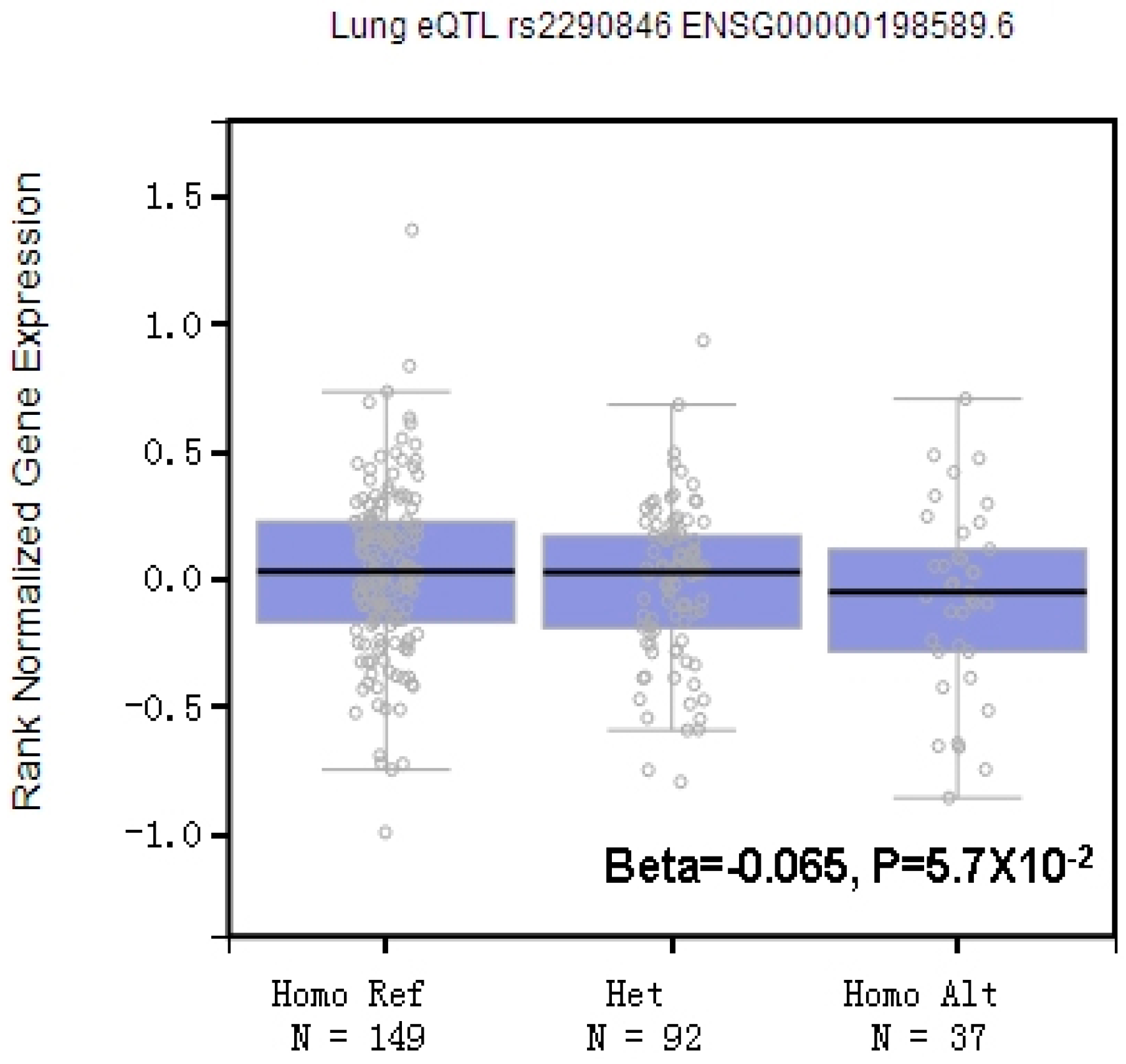
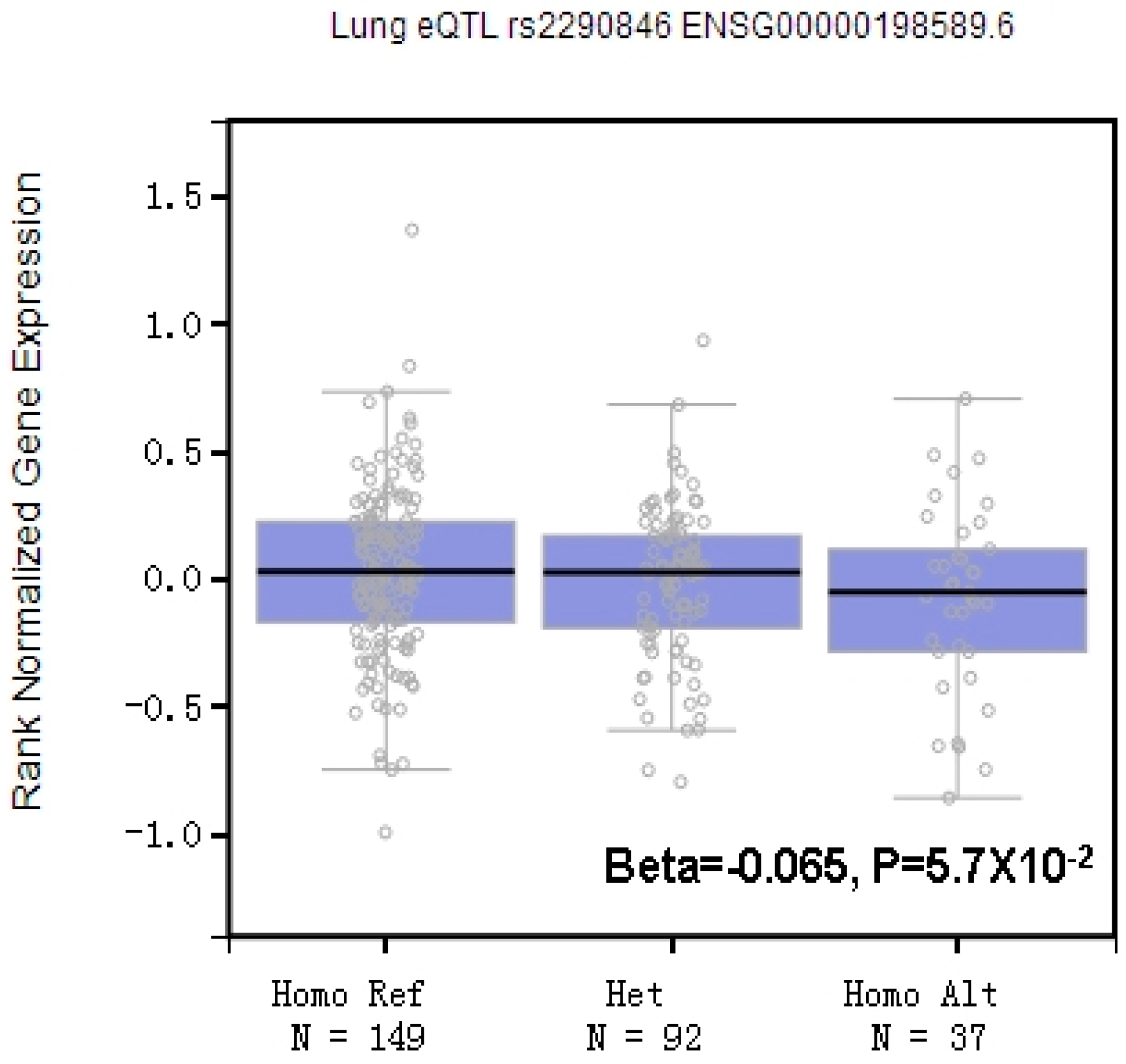
3.3. Potential Biological Roles of LRBA rs2290846 in CWP
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chuang, C.S.; Ho, S.C.; Lin, C.L.; Lin, M.C.; Kao, C.H. Risk of cerebrovascular events in pneumoconiosis patients: A population-based study, 1996–2011. Medicine 2016, 95, e2944. [Google Scholar] [CrossRef] [PubMed]
- National Report of Occupational Diseases in 2014. Available online: http://www.nhfpc.gov.cn/jkj/s5899t/201512/c5a99f823c5d4dd48324c6be69b7b2f9.shtml (accessed on 3 December 2015).
- Oyunbileg, S.; Wang, J.D.; Sumberzul, N.; Chang, Y.Y.; Erdenchimeg, E. Health impact of pneumoconiosis in Mongolia: Estimation of losses in life expectancy and quality adjusted life expectancy. Am. J. Ind. Med. 2011, 54, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Castranova, V.; Vallyathan, V. Silicosis and coal workers’ pneumoconiosis. Environ. Health Perspect. 2000, 108 (Suppl. 4), 675–684. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, A.T.; Jenkins, G. Molecular Endotyping of Pulmonary Fibrosis. Chest 2016, 149, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Pollard, K.M. Silica, Silicosis, and Autoimmunity. Front. Immunol. 2016, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Hahon, N.; Morgan, W.K.; Petersen, M. Serum immunoglobulin levels in coal workers’ pneumoconiosis. Ann. Occup. Hyg. 1980, 23, 165–174. [Google Scholar] [PubMed]
- Kotsianidis, I.; Nakou, E.; Bouchliou, I.; Tzouvelekis, A.; Spanoudakis, E.; Steiropoulos, P.; Sotiriou, I.; Aidinis, V.; Margaritis, D.; Tsatalas, C.; et al. Global impairment of CD4+CD25+FOXP3+ regulatory T cells in idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2009, 179, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Barbarin, V.; Arras, M.; Misson, P.; Delos, M.; McGarry, B.; Phan, S.H.; Lison, D.; Huaux, F. Characterization of the effect of interleukin-10 on silica-induced lung fibrosis in mice. Am. J. Respir. Cell Mol. Biol. 2004, 31, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Huaux, F.; Louahed, J.; Hudspith, B.; Meredith, C.; Delos, M.; Renauld, J.C.; Lison, D. Role of interleukin-10 in the lung response to silica in mice. Am. J. Respir. Cell Mol. Biol. 1998, 18, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.P.; Hemsley, S.; Canfield, P.J. Association of uterine and salpingeal fibrosis with chlamydial hsp60 and hsp10 antigen-specific antibodies in Chlamydia-infected koalas. Clin. Diagn. Lab. Immunol. 2005, 12, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Klionsky, D.J. Development by self-digestion: Molecular mechanisms and biological functions of autophagy. Dev. Cell 2004, 6, 463–477. [Google Scholar] [CrossRef]
- Ravikumar, B.; Sarkar, S.; Davies, J.E.; Futter, M.; Garcia-Arencibia, M.; Green-Thompson, Z.W.; Jimenez-Sanchez, M.; Korolchuk, V.I.; Lichtenberg, M.; Luo, S.; et al. Regulation of mammalian autophagy in physiology and pathophysiology. Physiol. Rev. 2010, 90, 1383–1435. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.; Wang, F.; Gao, R.; Wu, J.; Ou, Y.; Chen, X.; Wang, T.; Zhou, X.; Zhu, W.; Li, P.; et al. Autophagy inhibition of hsa-miR-19a-3p/19b-3p by targeting TGF-beta R II during TGF-beta1-induced fibrogenesis in human cardiac fibroblasts. Sci. Rep. 2016, 6, 24747. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Ogura, Y.; Suzuki, T.; Sen, S.; Lee, S.M.; Kanasaki, K.; Kume, S.; Koya, D. A very-low-protein diet ameliorates advanced diabetic nephropathy through autophagy induction by suppression of the mTORC1 pathway in Wistar fatty rats, an animal model of type 2 diabetes and obesity. Diabetologia 2016, 59, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Schmitz, U.; Raatz, Y.; Schonherr, M.; Kottek, T.; Schauer, M.; Franz, S.; Saalbach, A.; Anderegg, U.; Wolkenhauer, O.; et al. miR-638 promotes melanoma metastasis and protects melanoma cells from apoptosis and autophagy. Oncotarget 2015, 6, 2966–2980. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yuan, J.; Yao, S.; Jin, Y.; Chen, G.; Tian, W.; Xi, J.; Xu, Z.; Weng, D.; Chen, J. Lipopolysaccharides may aggravate apoptosis through accumulation of autophagosomes in alveolar macrophages of human silicosis. Autophagy 2015, 11, 2346–2357. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Ji, X.; Rong, R.; Li, Y.; Yao, W.; Yuan, J.; Wu, Q.; Yang, J.; Yan, W.; Han, L.; et al. MiR-449a regulates autophagy to inhibit silica-induced pulmonary fibrosis through targeting Bcl2. J. Mol. Med. 2016, 94, 1267–1279. [Google Scholar] [CrossRef] [PubMed]
- Lo, B.; Zhang, K.; Lu, W.; Zheng, L.; Zhang, Q.; Kanellopoulou, C.; Zhang, Y.; Liu, Z.; Fritz, J.M.; Marsh, R.; et al. AUTOIMMUNE DISEASE. Patients with LRBA deficiency show CTLA4 loss and immune dysregulation responsive to abatacept therapy. Science 2015, 349, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Charbonnier, L.M.; Janssen, E.; Chou, J.; Ohsumi, T.K.; Keles, S.; Hsu, J.T.; Massaad, M.J.; Garcia-Lloret, M.; Hanna-Wakim, R.; Dbaibo, G.; et al. Regulatory T-cell deficiency and immune dysregulation, polyendocrinopathy, enteropathy, X-linked-like disorder caused by loss-of-function mutations in LRBA. J. Allergy Clin. Immunol. 2015, 135, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Revel-Vilk, S.; Fischer, U.; Keller, B.; Nabhani, S.; Gamez-Diaz, L.; Rensing-Ehl, A.; Gombert, M.; Honscheid, A.; Saleh, H.; Shaag, A.; et al. Autoimmune lymphoproliferative syndrome-like disease in patients with LRBA mutation. Clin. Immunol. 2015, 159, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Alkhairy, O.K.; Abolhassani, H.; Rezaei, N.; Fang, M.; Andersen, K.K.; Chavoshzadeh, Z.; Mohammadzadeh, I.; El-Rajab, M.A.; Massaad, M.; Chou, J.; et al. Spectrum of Phenotypes Associated with Mutations in LRBA. J. Clin. Immunol. 2016, 36, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ye, Y.; Qian, H.; Song, Z.; Jia, X.; Zhang, Z.; Zhou, J.; Ni, C. Common genetic variants in pre-microRNAs are associated with risk of coal workers’ pneumoconiosis. J. Hum. Genet. 2010, 55, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.D.; Pan, Z.Q.; Liu, S.L.; Chen, L.; Ma, J.Z.; Yang, M.L.; Wang, N.P. The estimation of the number of underground coal miners and the annual dose to coal miners in China. Health Phys. 2007, 93, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Sosulski, M.L.; Gongora, R.; Danchuk, S.; Dong, C.; Luo, F.; Sanchez, C.G. Deregulation of selective autophagy during aging and pulmonary fibrosis: The role of TGFbeta1. Aging Cell 2015, 14, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Herrera, G.; Tampella, G.; Pan-Hammarstrom, Q.; Herholz, P.; Trujillo-Vargas, C.M.; Phadwal, K.; Simon, A.K.; Moutschen, M.; Etzioni, A.; Mory, A.; et al. Deleterious mutations in LRBA are associated with a syndrome of immune deficiency and autoimmunity. Am. J. Hum. Genet. 2012, 90, 986–1001. [Google Scholar] [CrossRef] [PubMed]
- Ballabio, A. Disease pathogenesis explained by basic science: Lysosomal storage diseases as autophagocytic disorders. Int. J. Clin. Pharmacol. Ther. 2009, 47 (Suppl. 1), S34–S38. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Ji, X.; Han, R.; Han, L.; Wang, T.; Yang, J.; Zhu, B.; Ni, C. GITR promoter polymorphism contributes to risk of coal workers’ pneumoconiosis: A case-control study from China. Immunol. Lett. 2014, 162, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Hessel, P.A.; Gamble, J.F.; Nicolich, M. Relationship between silicosis and smoking. Scand. J. Work Environ. Health 2003, 29, 329–336. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Variables | CWP (n = 703) | Controls (n = 705) | p | ||
|---|---|---|---|---|---|
| n | % | n | % | ||
| Age, years (mean ± SD) | 67.3 ± 10.8 | 66.3 ± 9.7 | 0.086 | ||
| Exposure years (mean ± SD) | 24.9 ± 8.5 | 25.5 ± 7.1 | 0.170 | ||
| Smoking | 0.088 | ||||
| No | 374 | 53.2 | 343 | 48.7 | |
| Yes | 329 | 46.8 | 362 | 51.3 | |
| Pack-years smoked | <0.001 | ||||
| 0 | 374 | 53.2 | 343 | 48.7 | |
| 0–20 | 201 | 28.6 | 165 | 23.4 | |
| >20 | 128 | 18.2 | 197 | 27.9 | |
| Job type | 0.703 | ||||
| Tunnel and coal mining | 590 | 83.9 | 599 | 85.0 | |
| Transport | 40 | 5.7 | 42 | 6.0 | |
| Others | 73 | 10.4 | 64 | 9.1 | |
| Stage | |||||
| I | 431 | 61.3 | |||
| II | 209 | 29.7 | |||
| III | 63 | 9.0 | |||
| Cluster ID | Region | dbSNP Allele | Function | Protein Residue | MAF | HWE a | |
|---|---|---|---|---|---|---|---|
| Case | Control | ||||||
| rs2290846 | Exon-56/57 | G > A | missense | Ser/Leu | 0.256 | 0.279 | 0.060 |
| rs3749574 | Exon-54/57 | C > T | missense | Ala/Thr | 0.251 | 0.275 | 0.504 |
| rs1782360 | Exon-54/57 | G > C | missense | Ala/Gly | 0.125 | 0.123 | 0.091 |
| Variables | CWP Cases | Controls | OR (95% CI) a | p a | OR (95% CI) b | p b | ||
|---|---|---|---|---|---|---|---|---|
| n | % | n | % | |||||
| rs2290846 | n = 695 | n = 684 | ||||||
| GG | 384 | 55.3 | 371 | 54.2 | 1.00 | - | 1.00 | - |
| GA | 266 | 38.3 | 245 | 35.8 | 1.05 (0.84–1.31) | 0.677 | 1.05 (0.84–1.32) | 0.680 |
| AA | 45 | 6.5 | 68 | 9.9 | 0.64 (0.43–0.96) | 0.030 | 0.61 (0.41–0.92) | 0.018 |
| Dominant | 0.96 (0.78–1.19) | 0.706 | 0.95 (0.77–1.18) | 0.658 | ||||
| Recessive | 0.63 (0.42–0.93) | 0.020 | 0.60 (0.40–0.89) | 0.011 | ||||
| Additive | 0.90 (0.76–1.06) | 0.196 | 0.89 (0.75–1.05) | 0.152 | ||||
| rs3749574 | n = 698 | n = 692 | ||||||
| CC | 386 | 55.3 | 360 | 52.0 | 1.00 | - | 1.00 | - |
| CT | 274 | 39.3 | 284 | 41.0 | 0.90 (0.72–1.12) | 0.346 | 0.90 (0.72–1.13) | 0.364 |
| TT | 38 | 5.4 | 48 | 6.9 | 0.74 (0.47–1.16) | 0.186 | 0.70 (0.45–1.10) | 0.124 |
| Dominant | 0.88 (0.71–1.08) | 0.221 | 0.88 (0.71–1.09) | 0.232 | ||||
| Recessive | 0.77 (0.50–1.20) | 0.249 | 0.73 (0.47–1.14) | 0.162 | ||||
| Additive | 0.88 (0.74–1.05) | 0.146 | 0.87 (0.73–1.04) | 0.127 | ||||
| rs1782360 | n = 695 | n = 686 | ||||||
| GG | 539 | 77.6 | 534 | 77.8 | 1.00 | - | 1.00 | - |
| CG | 139 | 20.0 | 135 | 19.7 | 1.02 (0.78–1.33) | 0.883 | 1.01 (0.77–1.32) | 0.943 |
| CC | 17 | 2.4 | 17 | 2.5 | 0.99 (0.50–1.96) | 0.979 | 1.01 (0.51–2.02) | 0.971 |
| Dominant | 1.02 (0.79–1.31) | 0.898 | 1.01 (0.78–1.30) | 0.964 | ||||
| Recessive | 0.99 (0.50–1.95) | 0.969 | 1.01 (0.51–2.00) | 0.985 | ||||
| Additive | 1.01 (0.81–1.26) | 0.922 | 1.01 (0.81–1.25) | 0.964 | ||||
| Variables | rs2290846 | |||
|---|---|---|---|---|
| Cases a | Controls a | OR (95% CI) b | p b | |
| Exposure years | ||||
| <27 | 341/27 | 355/37 | 0.73 (0.43–1.23) | 0.237 |
| ≥27 | 309/18 | 261/31 | 0.51 (0.28–0.94) | 0.032 |
| Pack-years smoked | ||||
| 0 | 343/24 | 302/36 | 0.58 (0.34–1.00) | 0.049 |
| >0–20 | 184/16 | 138/19 | 0.65 (0.32–1.33) | 0.241 |
| >20 | 123/5 | 176/13 | 0.57 (0.20–1.69) | 0.313 |
| Variables | Cases/Controls | Genotypes (Cases/Controls) | p a | OR (95% CI) a | |||
|---|---|---|---|---|---|---|---|
| AA | GG/GA | ||||||
| N | % | N | % | ||||
| Total | 703/705 | 45/68 | 6.4/9.6 | 658/637 | 93.6/90.4 | 0.011 | 0.60 (0.40–0.89) |
| Stage | |||||||
| Ι | 431/705 | 26/68 | 6.0/9.6 | 405/637 | 94.0/90.4 | 0.035 | 0.60 (0.37–0.97) |
| II | 209/705 | 14/68 | 6.7/9.6 | 195/637 | 93.3/90.4 | 0.137 | 0.62 (0.34–1.16) |
| III | 63/705 | 5/68 | 7.9/9.6 | 58/637 | 92.1/90.4 | 0.381 | 0.65 (0.24–1.72) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Yang, J.; Wu, Q.; Han, R.; Yan, W.; Yuan, J.; Ji, X.; Li, Y.; Yao, W.; Ni, C. LRBA Gene Polymorphisms and Risk of Coal Workers’ Pneumoconiosis: A Case–Control Study from China. Int. J. Environ. Res. Public Health 2017, 14, 1138. https://doi.org/10.3390/ijerph14101138
Liu Y, Yang J, Wu Q, Han R, Yan W, Yuan J, Ji X, Li Y, Yao W, Ni C. LRBA Gene Polymorphisms and Risk of Coal Workers’ Pneumoconiosis: A Case–Control Study from China. International Journal of Environmental Research and Public Health. 2017; 14(10):1138. https://doi.org/10.3390/ijerph14101138
Chicago/Turabian StyleLiu, Yi, Jingjin Yang, Qiuyun Wu, Ruhui Han, Weiwen Yan, Jiali Yuan, Xiaoming Ji, Yan Li, Wenxi Yao, and Chunhui Ni. 2017. "LRBA Gene Polymorphisms and Risk of Coal Workers’ Pneumoconiosis: A Case–Control Study from China" International Journal of Environmental Research and Public Health 14, no. 10: 1138. https://doi.org/10.3390/ijerph14101138