Association of Bisphenol A Exposure with LINE-1 Hydroxymethylation in Human Semen

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Data Collection
2.2. BPA Measurement
2.3. Semen Collection
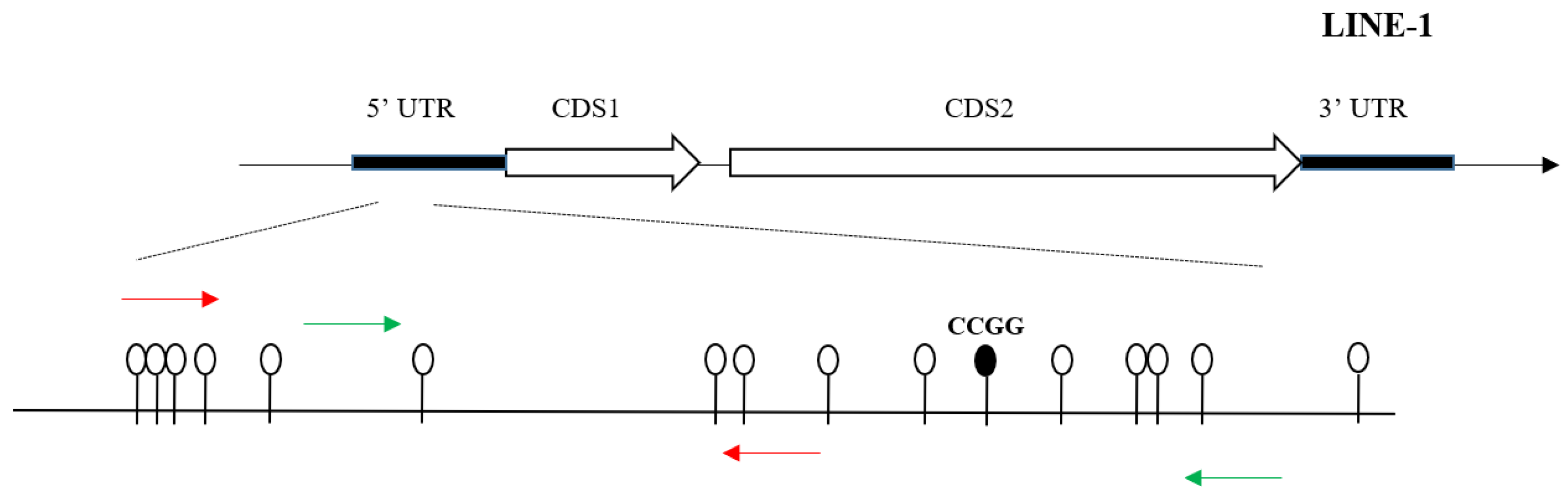
2.4. DNA Extraction and Hydroxymethylation Measurement of LINE-1
2.5. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgements
Conflict of interest
References
- Calafat, A.M.; Ye, X.; Wong, L.Y.; Reidy, J.A.; Needham, L.L. Exposure of the U.S. Population to Bisphenol A and 4-Tertiary-Octylphenol: 2003–2004. Environ. Health Perspect. 2008, 116, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Kuklenyik, Z.; Reidy, J.A.; Caudill, S.P.; Ekong, J.; Needham, L.L. Urinary Concentrations of Bisphenol A and 4-Nonylphenol in a Human Reference Population. Environ. Health Perspect. 2005, 113, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, J.C.; Leonard, L.S.; Maness, S.C.; Wagner, B.L.; Conner, K.; Zacharewski, T.; Safe, S.; Mcdonnell, D.P.; Gaido, K.W. Bisphenol A Interacts with the Estrogen Receptor α in a Distinct Manner from Estradiol. Mol. Cell. Endocrinol. 1998, 142, 203–214. [Google Scholar] [CrossRef]
- Bonefeld-Jørgensen, E.C.; Long, M.; Hofmeister, M.V.; Vinggaard, A.M. Endocrine-Disrupting Potential of Bisphenol A, Bisphenol A Dimethacrylate, 4-n-Nonylphenol, and 4-n-Octylphenol in Vitro: New Data and a Brief Review. Environ. Health Perspect. 2007, 115, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.L.; Wang, X.; Zhang, X.H.; Zhang, Z.; Gu, J.; Liu, L.; Wang, Y.; Wang, X.; Wang, S.L. Decreased Androgen Receptor Expression May Contribute to Spermatogenesis Failure in Rats Exposed to Low Concentration of Bisphenol A. Toxicol. Lett. 2013, 219, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Tainaka, H.; Takahashi, H.; Umezawa, M.; Tanaka, H.; Nishimune, Y.; Oshio, S.; Takeda, K. Evaluation of the Testicular Toxicity of Prenatal Exposure to Bisphenol A Based on Microarray Analysis Combined with MeSH Annotation. J. Toxicol. Sci. 2012, 37, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Lassen, T.H.; Frederiksen, H.; Jensen, T.K.; Petersen, J.H.; Joensen, U.N.; Main, K.M.; Skakkebaek, N.E.; Juul, A.; Jørgensen, N.; Andersson, A.-M. Urinary Bisphenol A Levels in Young Men: Association with Reproductive Hormones and Semen Quality. Environ. Health Perspect. 2014, 122, 478–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstone, A.E.; Chen, Z.; Perry, M.J.; Kannan, K.; Louis, G.M.B. Urinary Bisphenol a and Semen Quality: The Life Study. Reprod. Toxicol. 2015, 51, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Meeker, J.D.; Ehrlich, S.; Toth, T.L.; Wright, D.L.; Calafat, A.M.; Trisini, A.T.; Ye, X.; Hauser, R. Semen quality and sperm DNA damage in relation to urinary bisphenol A among men from an infertility clinic. Reprod. Toxicol. 2010, 30, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-K.; Zhou, Z.; Miao, M.; He, Y.; Qing, D.; Wu, T.; Wang, J.; Weng, X.; Ferber, J.; Herrinton, L.J.; et al. Relationship Between Urine Bisphenol-A Level and Declining Male Sexual Function. J. Androl. 2010, 31, 500–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.K.; Zhou, Z.; Miao, M.; He, Y.; Wang, J.; Ferber, J.; Herrinton, L.J.; Gao, E.; Yuan, W. Urine bisphenol-A (BPA) level in relation to semen quality. Fertil. Steril. 2011, 95, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Zhang, X.; Wang, D.; Baccarelli, A. Environmental chemical exposures and human epigenetics. Int. J. Epidemiol. 2012, 41, 79–105. [Google Scholar] [CrossRef] [PubMed]
- Fleisch, A.F.; Wright, R.O.; Baccarelli, A.A. Environmental epigenetics: A role in endocrine disease? J. Mol. Endocrinol. 2012, 49, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Zhou, X.; Li, Y.; Zhang, O.; Zhou, Z.; Li, T.; Yuan, W.; Li, R.; Li, D.-K. LINE-1 Hypomethylation in Spermatozoa Is Associated with Bisphenol A Exposure. Andrology-US 2014, 2, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Skinner, M.K. Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLoS ONE 2013, 8, e55387. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Dai, Y.; Jiang, X.; Liu, Y.; Chen, H.; Han, F.; Cao, J.; Liu, J. Role of DNA Methylation in Bisphenol A Exposed Mouse Spermatocyte. Environ Toxicol. Pharmacol. 2016, 48, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Kriaucionis, S.; Heintz, N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science 2009, 324, 929–930. [Google Scholar] [CrossRef] [PubMed]
- Tahiliani, M.; Koh, K.P.; Shen, Y.; Pastor, W.A.; Bandukwala, H.; Brudno, Y.; Agarwal, S.; Iyer, L.M.; Liu, D.R.; Aravind, L.; et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 2009, 324, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; D’ Alessio, A.C.; Taranova, O.V.; Hong, K.; Sowers, L.C.; Zhang, Y. Role of Tet proteins in 5mC to 5hmC conversion, ES cell self-renewal, and ICM specification. Nature 2010, 466, 1129. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Hon, G.C.; Szulwach, K.E.; Song, C.X.; Zhang, L.; Kim, A.; Li, X.; Dai, Q.; Shen, Y.; Park, B.; et al. Base-resolution analysis of 5-hydroxymethylcytosine in the mammalian genome. Cell 2012, 149, 1368–1380. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.; Christensen, J.; Pedersen, M.T.; Johansen, J.V.; Cloos, P.A.; Rappsilber, J.; Helin, K. TET1 and hydroxymethylcytosine in transcription and DNA methylation fidelity. Nature 2011, 473, 343–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellen, M.; Ayata, P.; Dewell, S.; Kriaucionis, S.; Heintz, N. MeCP2 binds to 5hmC enriched within active genes and accessible chromatin in the nervous system. Cell 2012, 151, 1417–1430. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, O.; Li, R.; Hung, J.H.; Chen, P.B.; Dong, X.; Ee, L.S.; Weng, Z.; Rando, O.J; Fazzio, T.G. Mbd3/NURD complex regulates expression of 5-hydroxymethylcytosine marked genes in embryonic stem cells. Cell 2011, 147, 1498–1510. [Google Scholar] [CrossRef] [PubMed]
- Bachman, M.; Uribe-Lewis, S.; Yang, X.; Williams, M.; Murrell, A.; Balasubramanian, S. 5-Hydroxymethylcytosine is a predominantly stable DNA modification. Nat. Chem. 2014, 6, 1049–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, H.; Wen, L.; Liao, S.; Lin, X.; Ma, T.; Liu, J.; Song, C.-X.; Wang, M.; He, C.; Han, C.; et al. Dynamics of 5-Hydroxymethylcytosine during Mouse Spermatogenesis. Nat. Commun. 2013, 4, 1995. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Maksoud, F.M.; Leasor, K.R.; Butzen, K.; Braden, T.D.; Akingbemi, B.T. Prenatal Exposures of Male Rats to the Environmental Chemicals Bisphenol A and Di(2-Ethylhexyl) Phthalate Impact the Sexual Differentiation Process. Endocrinology 2015, 156, 4672–4683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Zhang, Y.; Liu, Y.; Wang, S.; Wang, Z. DNA demethylation mediated by down-regulated TETs in the testes of rare minnow Gobiocypris rarus under bisphenol A exposure. Chemosphere 2017, 171, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [PubMed]
- Khare, T.; Pai, S.; Koncevicius, K.; Pal, M.; Kriukiene, E.; Liutkeviciute, Z.; Irimia, M.; Jia, P.; Ptak, C.; Xia, M.; et al. 5-HmC in the Brain Is Abundant in Synaptic Genes and Shows Differences at the Exon-Intron Boundary. Nat. Struct. Mol. Biol. 2012, 19, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Booth, M.J.; Branco, M.R.; Ficz, G.; Oxley, D.; Krueger, F.; Reik, W.; Balasubramanian, S. Quantitative sequencing of 5-methylcytosine and 5-hydroxymethylcytosine at single-base resolution. Science 2012, 336, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zhou, X.; Li, D.-K.; Yang, F.; Pan, H.; Li, T.; Miao, M.; Li, R.; Yuan, W. Genome-Wide Alteration in DNA Hydroxymethylation in the Sperm from Bisphenol A-Exposed Men. PLoS ONE 2017, 12, e178535. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Miao, M.; Herrinton, L.J.; Wu, C.; Yuan, W.; Zhou, Z.; Li, D.-K. Bisphenol A Levels in Blood and Urine in a Chinese Population and the Personal Factors Affecting the Levels. Environ. Res. 2009, 109, 629–633. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Laboratory Manual for the Examination of Human Semen and Sperm-Cervical Mucus Interaction, 4th ed.; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Hur, K.; Cejas, P.; Feliu, J.; Moreno-Rubio, J.; Burgos, E.; Boland, C.R.; Goel, A. Hypomethylation of Long Interspersed Nuclear Element-1 (LINE-1) Leads to Activation of Proto-Oncogenes in Human Colorectal Cancer Metastasis. Gut 2013, 63, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Tammen, S.A.; Dolnikowski, G.G.; Ausman, L.M.; Liu, Z.; Sauer, J.; Friso, S.; Choi, S.-W. Aging and Alcohol Interact to Alter Hepatic DNA Hydroxymethylation. Alcohol. Clin. Exp. Res. 2014, 38, 2178–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tellez-Plaza, M.; Tang, W.-Y.; Shang, Y.; Umans, J.G.; Francesconi, K.A.; Goessler, W.; Ledesma, M.; Leon, M.; Laclaustra, M.; Pollak, J.; et al. Association of Global DNA Methylation and Global DNA Hydroxymethylation with Metals and Other Exposures in Human Blood DNA Samples. Environ. Health Perspect. 2014, 122, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.A.; Da, S.I.; Mariz, B.A.; Da, S.M.; Freitas-Ribeiro, G.M.; de Oliveira, N.F. Influence of smoking on methylation and hydroxymethylation levels in global DNA and specific sites of KRT14, KRT19, MIR-9-3 and MIR-137 genes of oral mucosa. Arch. Oral Biol. 2016, 72, 56–65. [Google Scholar] [CrossRef] [PubMed]
- James, S.J.; Shpyleva, S.; Melnyk, S.; Pavliv, O.; Pogribny, I.P. Elevated 5-hydroxymethylcytosine in the Engrailed-2 (EN-2) promoter is associated with increased gene expression and decreased MeCP2 binding in autism cerebellum. Transl. Psychiatry 2014, 4, e460. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.R.; Burns, K.H.; Boeke, J.D. Active transposition in genomes. Annu. Rev. Genet. 2012, 46, 651–675. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.; Jiang, L.; Liu, X.; Geng, C.; Wang, W.; Zhong, L.; Yang, G.; Chen, M. Bisphenol A Induces Oxidative Stress-Associated DNA Damage in INS-1 Cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 769, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, S.; Mousavi, S.N.; Aghapour, F.; Rezaee, B.; Sadeghi, F.; Moghadamnia, A.A. Induction Effect of Bisphenol A on Gene Expression Involving Hepatic Oxidative Stress in Rat. Oxid. Med. Cell. Longev. 2016, 2016, 6298515. [Google Scholar] [CrossRef] [PubMed]
- Chia, N.; Wang, L.; Lu, X.; Senut, M.C.; Brenner, C.; Ruden, D.M. Hypothesis: Environmental regulation of 5-hydroxymethylcytosine by oxidative stress. Epigenetics-US 2011, 6, 853–856. [Google Scholar] [CrossRef] [Green Version]
- Niedzwiecki, M.M.; Liu, X.; Hall, M.N.; Thomas, T.; Slavkovich, V.; Ilievski, V.; Levy, D.; Alam, S.; Siddique, A.B.; Parvez, F.; et al. Sex-Specific Associations of Arsenic Exposure with Global DNA Methylation and Hydroxymethylation in Leukocytes: Results from Two Studies in Bangladesh. Cancer Epidemiol. Biomarkers Prev. 2015, 24, 1748–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Guerra, M.; Zheng, Y.; Osorio-Yanez, C.; Zhong, J.; Chervona, Y.; Wang, S.; Chang, D.; McCracken, J.P.; Díaz, A.; Bertazzi, P.A.; et al. Effects of particulate matter exposure on blood 5-hydroxymethylation: Results from the Beijing truck driver air pollution study. Epigenetics 2015, 10, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Coulter, J.B.; O'Driscoll, C.M.; Bressler, J.P. Hydroquinone increases 5-hydroxymethylcytosine formation through ten eleven translocation 1 (TET1) 5-methylcytosine dioxygenase. J. Biol. Chem. 2013, 288, 28792–28800. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Kwon, W.S.; Lee, J.S.; Yoon, S.J.; Ryu, B.Y.; Pang, M.G. Bisphenol-A affects male fertility via fertility-related proteins in spermatozoa. Sci. Rep. 2015, 5, 9169. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lin, H.; Xu, H.; Zhang, L.; Cheng, L.; Wen, B.; Shou, J.; Guan, K.; Xiong, Y.; Ye, D. TET-Catalyzed 5-Methylcytosine Hydroxylation Is Dynamically Regulated by Metabolites. Cell Res. 2014, 24, 1017–1020. [Google Scholar] [CrossRef] [PubMed]
- Gassman, N.R.; Coskun, E.; Stefanick, D.F.; Horton, J.K.; Jaruga, P.; Dizdaroglu, M.; Wilson, S.H. Bisphenol A Promotes Cell Survival Following Oxidative DNA Damage in Mouse Fibroblasts. PLoS ONE 2015, 10, e118819. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.U.; Su, Y.; Zhong, C.; Ming, G.L.; Song, H. Hydroxylation of 5-methylcytosine by TET1 promotes active DNA demethylation in the adult brain. Cell 2011, 145, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Branco, M.R.; Ficz, G.; Reik, W. Uncovering the role of 5-hydroxymethylcytosine in the epigenome. Nat. Rev. Genet. 2011, 13, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Colquitt, B.M.; Allen, W.E.; Barnea, G.; Lomvardas, S. Alteration of genic 5-hydroxymethylcytosine patterning in olfactory neurons correlates with changes in gene expression and cell identity. Proc. Natl. Acad. Sci. USA 2013, 110, 14682–14687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, R.J.; Hamra, F.K.; Richardson, J.A.; Qi, X.; Bassel-Duby, R.; Olson, E.N. MOV10L1 is necessary for protection of spermatocytes against retrotransposons by Piwi-interacting RNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 11847–11852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tunc, O.; Tremellen, K. Oxidative DNA damage impairs global sperm DNA methylation in infertile men. J. Assist. Reprod. Genet. 2009, 26, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanassy, L.; Carrell, D.T. Analysis of the methylation pattern of six gene promoters in sperm of men with abnormal protamination. Asian J. Androl. 2011, 13, 342–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Characteristics | Exposed (n = 72) | Unexposed (n = 86) | p |
|---|---|---|---|
| Age (years) | |||
| <29 | 23 (31.94) | 32 (37.21) | 0.48 |
| 30–35 | 21 (29.17) | 18 (21.18) | |
| >36 | 28 (38.89) | 36 (41.86) | |
| Education | |||
| ≤Middle school | 21 (29.17) | 20 (23.26) | 0.57 |
| High school | 39 (54.17) | 47 (54.65) | |
| ≥College | 12 (16.67) | 19 (22.09) | |
| Smoking | |||
| Yes | 47 (65.28) | 60 (69.77) | 0.55 |
| No | 25 (34.72) | 26 (30.23) | |
| Alcohol consumption | |||
| Yes | 16 (22.22) | 22 (25.58) | 0.62 |
| No | 56 (77.78) | 64 (74.42) | |
| History of disease | |||
| Yes | 13 (18.06) | 16 (18.60) | 0.93 |
| No | 59 (81.94) | 70 (81.40) | |
| Groups | N | GM (std) | Median (Q1, Q3) |
|---|---|---|---|
| By occupational exposure status | |||
| BPA-exposed | 72 | 158.41 (17.92) | 238.78 (24.14, 2043.22) |
| Unexposed | 86 | 0.84 (6.53) | LOD (LOD, 6.07) |
| By urine BPA level | |||
| BPA undetectable (below LOD) | 53 | LOD | LOD |
| Low tertile (0–13.28) | 35 | 3.77 (3.07) | 4.78 (1.55, 8.19) |
| Middle tertile (13.28–274.82) | 35 | 33.94 (2.13) | 30.88 (22.67, 53.72) |
| Top tertile (274.82–) | 35 | 1698.88 (6.74) | 2158.44 (679.92, 9771.63) |
| Groups | N | Mean% (std) | 5th% | 25th% | 50th% | 75th% | 95th% | β | p |
|---|---|---|---|---|---|---|---|---|---|
| By occupational exposure status | |||||||||
| Unexposed | 86 | 9.68 (4.97) | 2.13 | 6.48 | 8.99 | 11.62 | 20.95 | Ref | - |
| Exposed | 72 | 12.97 (5.07) | 5.62 | 9.51 | 12.45 | 15.71 | 23.45 | 0.034 | <0.0001 |
| By urine BPA level | |||||||||
| BPA undetected | 53 | 8.79 (4.22) | 1.91 | 6.32 | 8.36 | 10.24 | 16.09 | Ref | - |
| Low tertile | 35 | 12.16 (5.75) | 5.25 | 6.92 | 11.85 | 17.13 | 22.55 | 0.031 | 0.006 |
| Middle tertile | 35 | 11.53 (4.58) | 5.99 | 8.54 | 10.26 | 15.22 | 21.51 | 0.023 | 0.047 |
| Top tertile | 35 | 13.45 (5.58) | 4.71 | 10.23 | 12.71 | 18.46 | 24.15 | 0.048 | <0.0001 |
| Stratified Analysis | N | Crude | Adjusted | ||
|---|---|---|---|---|---|
| β | p | β | p | ||
| Smoking | |||||
| NO | 51 | 0.042 | 0.002 | 0.047 | 0.001 |
| YES | 107 | 0.029 | 0.005 | 0.028 | 0.007 |
| Alcohol consumption | |||||
| NO | 120 | 0.034 | <0.001 | 0.035 | <0.001 |
| YES | 38 | 0.031 | 0.073 | 0.027 | 0.148 |
| Age group (years) | |||||
| <30 | 55 | 0.030 | 0.047 | 0.031 | 0.046 |
| 30–36 | 39 | 0.017 | 0.334 | 0.013 | 0.473 |
| ≥36 | 64 | 0.040 | <0.001 | 0.045 | <0.001 |
| Disease | |||||
| NO | 129 | 0.035 | <0.001 | 0.036 | <0.001 |
| YES | 29 | 0.023 | 0.167 | 0.025 | 0.167 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Zhou, X.; Miao, M.; Li, D.-k.; Wang, Z.; Li, R.; Liang, H.; Yuan, W. Association of Bisphenol A Exposure with LINE-1 Hydroxymethylation in Human Semen. Int. J. Environ. Res. Public Health 2018, 15, 1770. https://doi.org/10.3390/ijerph15081770
Tian Y, Zhou X, Miao M, Li D-k, Wang Z, Li R, Liang H, Yuan W. Association of Bisphenol A Exposure with LINE-1 Hydroxymethylation in Human Semen. International Journal of Environmental Research and Public Health. 2018; 15(8):1770. https://doi.org/10.3390/ijerph15081770
Chicago/Turabian StyleTian, Youping, Xiaoyu Zhou, Maohua Miao, De-kun Li, Ziliang Wang, Runsheng Li, Hong Liang, and Wei Yuan. 2018. "Association of Bisphenol A Exposure with LINE-1 Hydroxymethylation in Human Semen" International Journal of Environmental Research and Public Health 15, no. 8: 1770. https://doi.org/10.3390/ijerph15081770