Morphological and Physiological Responses of Morning Glory (Ipomoea lacunosa L.) Grown in a Lead- and Chelate-Amended Soil

Abstract
:Introduction
Materials and Methods
Experimental Design and Soil Preparation and Analysis
Chlorophyll Analysis and Photosynthetic Rates
Statistical Analysis
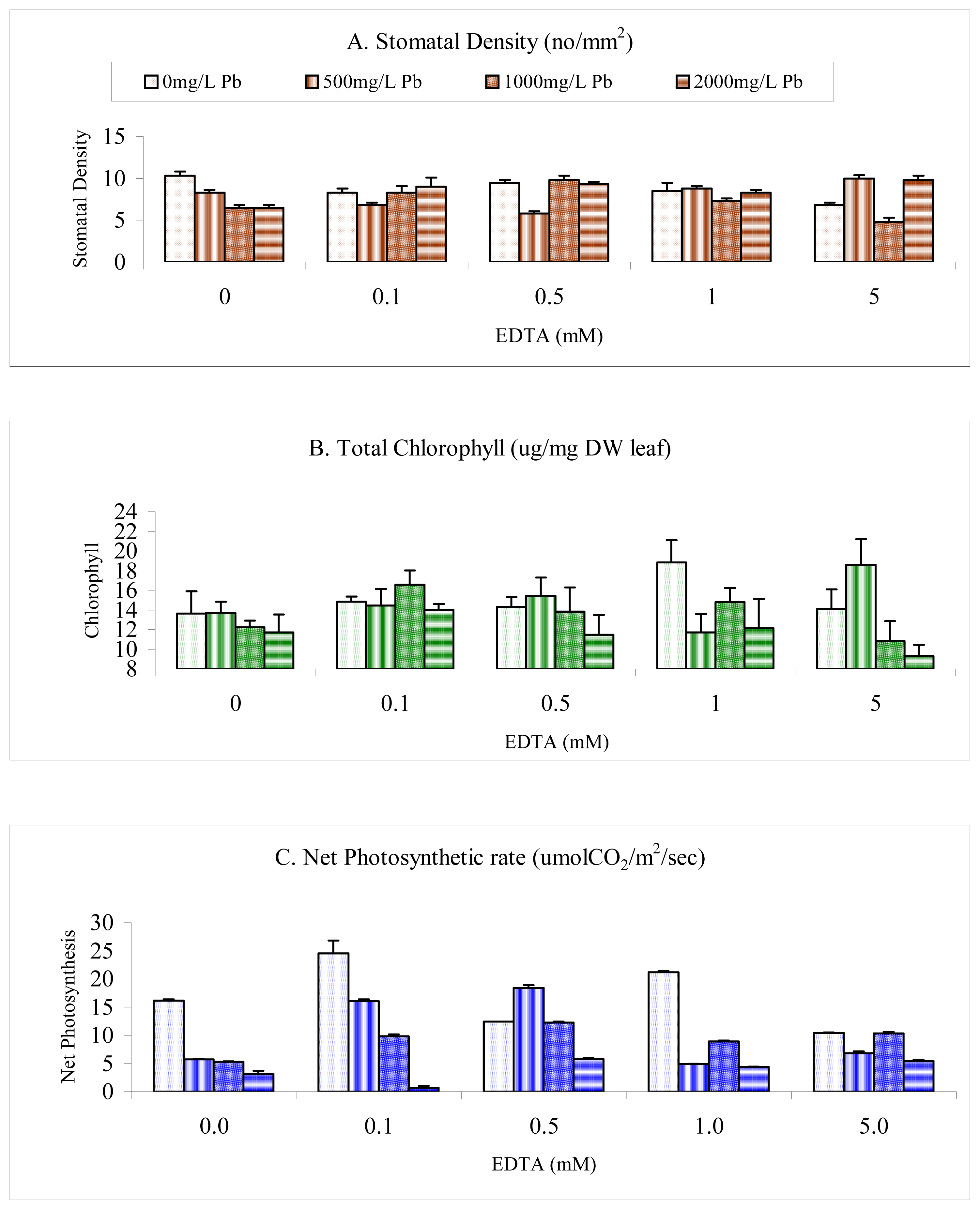
Results
Stomatal Density
Chlorophyll
Photosynthesis
Discussion
Conclusion


{kind=link}
| Source | DF | SS | MS | F-Value | P-Value |
|---|---|---|---|---|---|
| EDTA (mM) | 4 | 5.7 | 1.425 | 1.513 | 0.210 |
| Pb (mg/L) | 3 | 23.7 | 7.900 | 8.389* | 0.000 |
| EDTA vs. Pb | 12 | 149.3 | 12.442 | 13.212* | 0.000 |
| Between Groups | 19 | 178.7 | 9.405 | ||
| Error | 60 | 56.5 | 0.942 | ||
| Total | 79 | 235.200 | 2.977 | ||
| Source | DF | SS | MS | F-Value | P-value |
|---|---|---|---|---|---|
| EDTA (mM) | 4 | 47.58 | 11.89 | 0.893 | 0.474 |
| Pb (mg/L) | 3 | 140.57 | 46.85 | 3.518* | 0.020 |
| EDTA vs. Pb | 12 | 251.500 | 20.95 | 1.574 | 0.124 |
| Between Groups | 19 | 439.665 | 23.14 | ||
| Error | 60 | 799.110 | 13.31 | ||
| Total | 79 | 1238.775 | 15.681 | ||
| Source | DF | SS | MS | F-Value | P-value |
|---|---|---|---|---|---|
| EDTA (mM) | 4 | 257.122 | 64.28 | 64.163* | 0.000 |
| Pb (mg/L) | 3 | 1297.766 | 432.58 | 431.797* | 0.000 |
| EDTA vs. Pb | 12 | 776.295 | 64.69 | 64.573* | 0.000 |
| Between Groups | 19 | 2331.183 | 122.69 | ||
| Error | 40 | 40.073 | 1.00 | ||
| Total | 59 | 2371.256 | 40.191 | ||
Acknowledgments
References
- Singer, M. J.; Munn, D. N. Soils: An Introduction; Prentice Hall: New York, 1996. [Google Scholar]
- Krupa, Z.; Baszynski, T. Some aspects of heavy metals toxicity towards photosynthetic apparatus–direct and indirect effects on light and dark reactions. Acta. Physiol. Plant 1995, 17, 177–190. [Google Scholar]
- Sarvari, E.; Gasper, L.; Fodor, F.; Cseh, E.; Kropfl, K.; Varga, A.; Baron, M. Comparison of the effects of Pb treatment on thylakoid development in poplar and cucumber plants. Acta. Biolog. Szeged 2002, 46, 163–165. [Google Scholar]
- Zaman, M. S.; Zereen, F. Growth responses of radish plants to soil cadmium and lead contamination. Bull. Environ. Contam. Toxicol 1998, 61, 44–50. [Google Scholar]
- Rousos, P. A.; Harrison, H. C.; Steffen, K. L. Physiological responses of cabbage to incipient copper toxicity. J. Amer. Soc. Hort. Sci 1989, 114, 149–152. [Google Scholar]
- Pahlsson, A. M. B. Toxicity of heavy metals (Zn, Cu, Cd, Pb) to vascular plants. Water, Air, Soil Pollut 1989, 47, 287–319. [Google Scholar]
- Zayed, A.; Gowthaman, S.; Terry, N. Phytoaccumulation of trace elements by wetland plants: I. Duckweed. J. Environ. Qual 1998, 27, 715–721. [Google Scholar]
- Begonia, G. B. Comparative lead uptake and responses of some plants grown on lead contaminated soils. J. Mississippi Acad. Sci 1997, 42, 101–106. [Google Scholar]
- Einhellig, F. A.; Rasmussen, J. A. Effects of three phenolic acids on chlorophyll content and growth of soybean and grain sorghum seedlings. J. Chem. Ecol 1979, 5, 815–823. [Google Scholar]
- Baker, A. J. M. Metal tolerance. New Phytol 1987, 106 suppl, 93–111. [Google Scholar]
- Tanton, T. W.; Crowdy, S. H. The distribution of lead chelate in the transpiration stream of higher plants. Pesticide Sci 1971, 2, 211–213. [Google Scholar]
- Mac Robbie, E. A. C. Ionic relations of guard cells. In Stomatal Function; Zeigler, E., Farquhar, G. D., Cohen, I. R., Eds.; Stanford University Press: Stanford, CA, 1987; pp. 125–162. [Google Scholar]
- Outlaw, W. H., Jr. An introduction to carbon metabolism in guard cells. In Stomatal Function; Zeigler, E., Farquhar, G. D., Cohen, I. R., Eds.; Stanford University Press: Stanford, CA, 1987; pp. 115–123. [Google Scholar]
- Abdelbasset, R.; Issa, A. A.; Adam, M. S. Chlorophyllase activity-effects of heavy metals and calcium. Photosynthetica 1995, 31, 421–425. [Google Scholar]
- Johnson, W. R.; Proctor, J. A comparative study of metal levels in plants from two contrasting lead-mine sites. Plant Soil 1977, 46, 251–257. [Google Scholar]
- Johnson, M. S.; McNeilly, T.; Putwain, P. O. Revegetation of metalliferous mine spoil contaminated by lead and zinc. Environ. Pollut 1977, 12, 261–277. [Google Scholar]
- Salisbury, F. B.; Ross, C. W. Plant physiology; Wadsworth Publishing Company: Belmont, CA, 1992. [Google Scholar]
- Bazzaz, F. A.; Rolfe, G. L.; Windle, P. Differing sensitivity of corn and soybean photosynthesis and transpiration to lead concentration. J. Environ. Quality 1974, 3, 156–158. [Google Scholar]
© 2005 MDPI. All rights reserved.
Share and Cite
Kambhampati, M.S.; Begonia, G.B.; Begonia, M.F.T.; Bufford, Y. Morphological and Physiological Responses of Morning Glory (Ipomoea lacunosa L.) Grown in a Lead- and Chelate-Amended Soil. Int. J. Environ. Res. Public Health 2005, 2, 299-303. https://doi.org/10.3390/ijerph2005020014
Kambhampati MS, Begonia GB, Begonia MFT, Bufford Y. Morphological and Physiological Responses of Morning Glory (Ipomoea lacunosa L.) Grown in a Lead- and Chelate-Amended Soil. International Journal of Environmental Research and Public Health. 2005; 2(2):299-303. https://doi.org/10.3390/ijerph2005020014
Chicago/Turabian StyleKambhampati, Murty S., Gregorio B. Begonia, Maria F. T. Begonia, and Yolanda Bufford. 2005. "Morphological and Physiological Responses of Morning Glory (Ipomoea lacunosa L.) Grown in a Lead- and Chelate-Amended Soil" International Journal of Environmental Research and Public Health 2, no. 2: 299-303. https://doi.org/10.3390/ijerph2005020014