Effects of Benzo(a)pyrene on Intra-testicular Function in F-344 Rats

Abstract
:Introduction
Materials and Methods
Animals and Exposure
Post-exposure Processing Of Tissue Samples
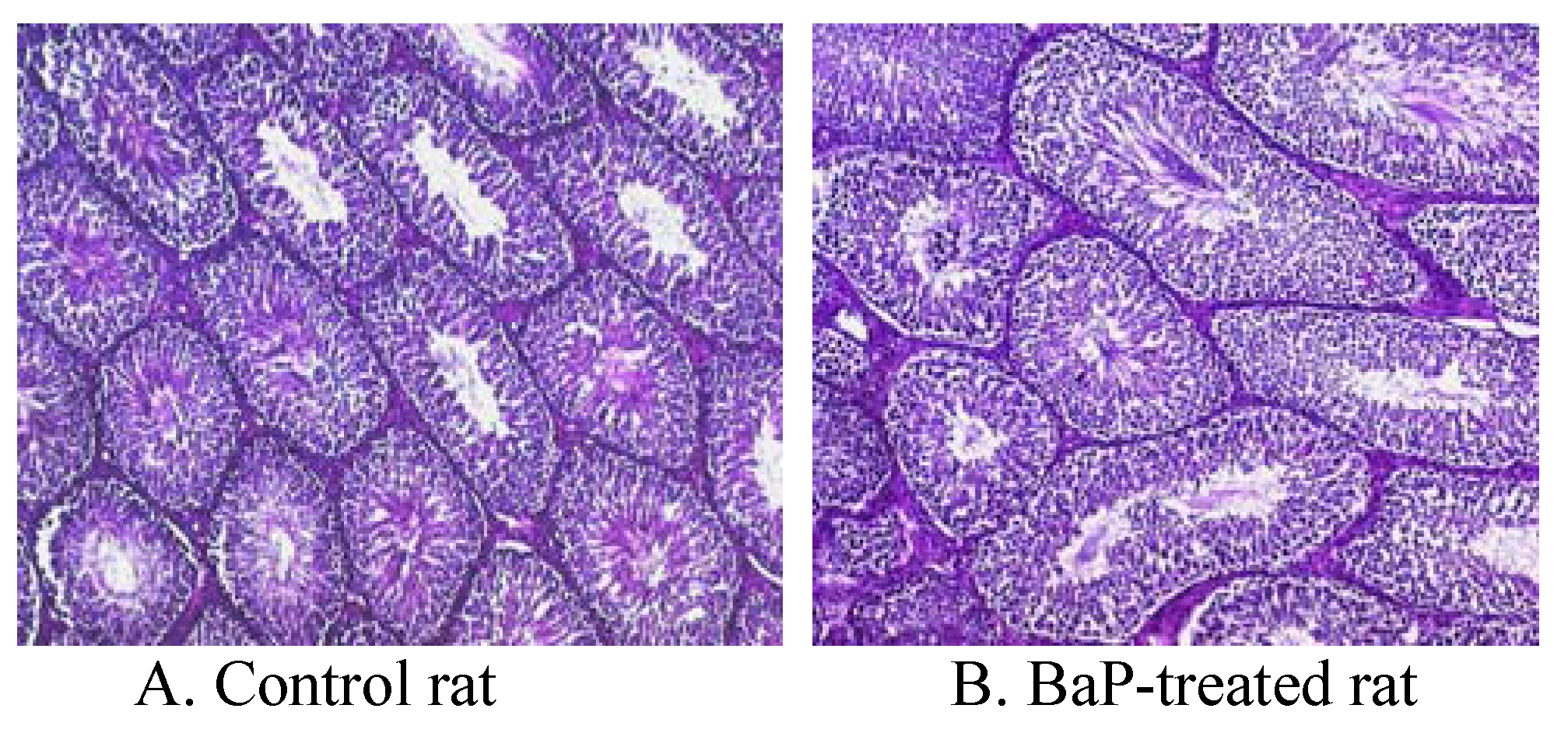
Morphometry & Histopathology
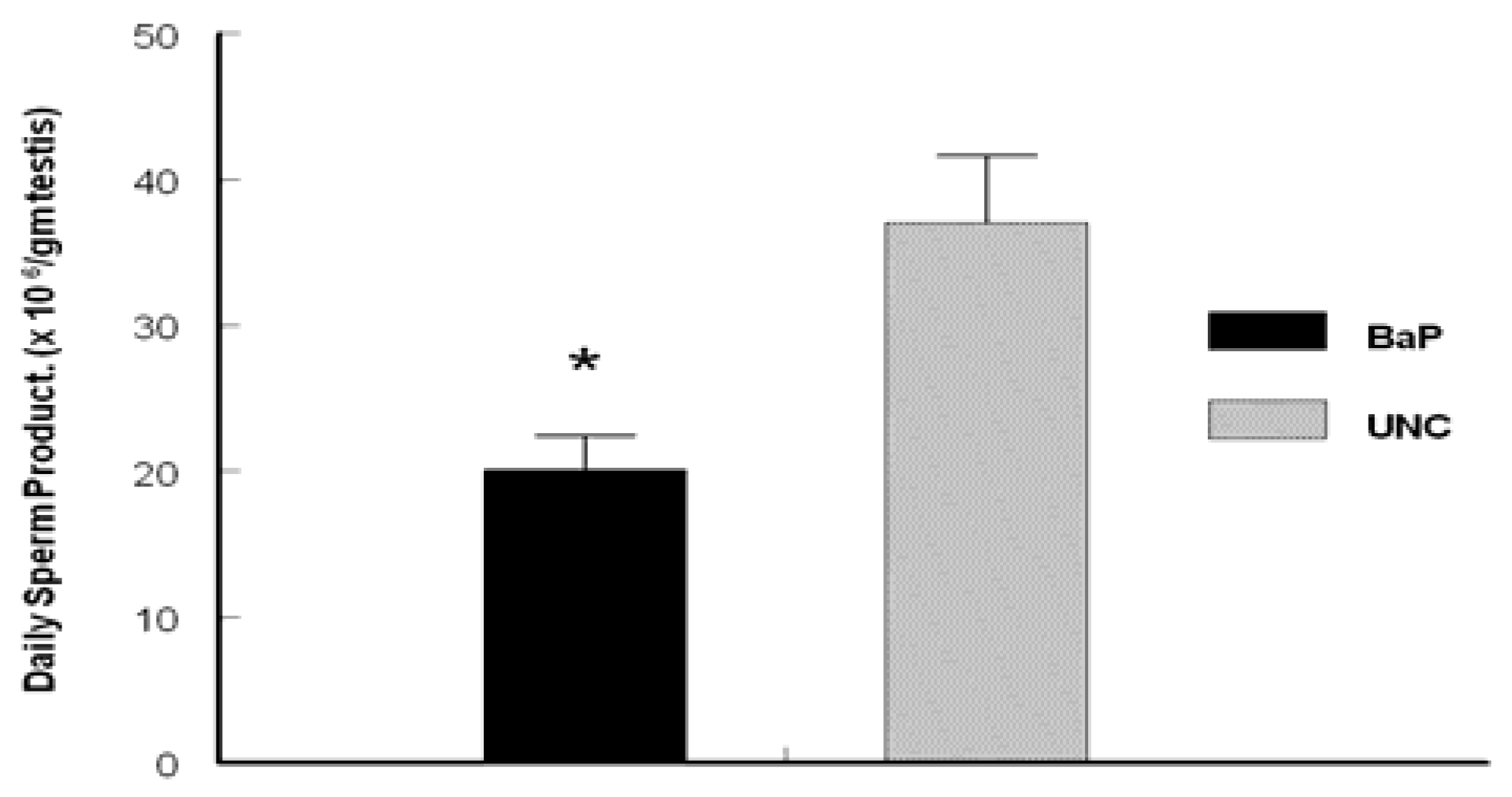
Determination of Daily Sperm Production (DSP)
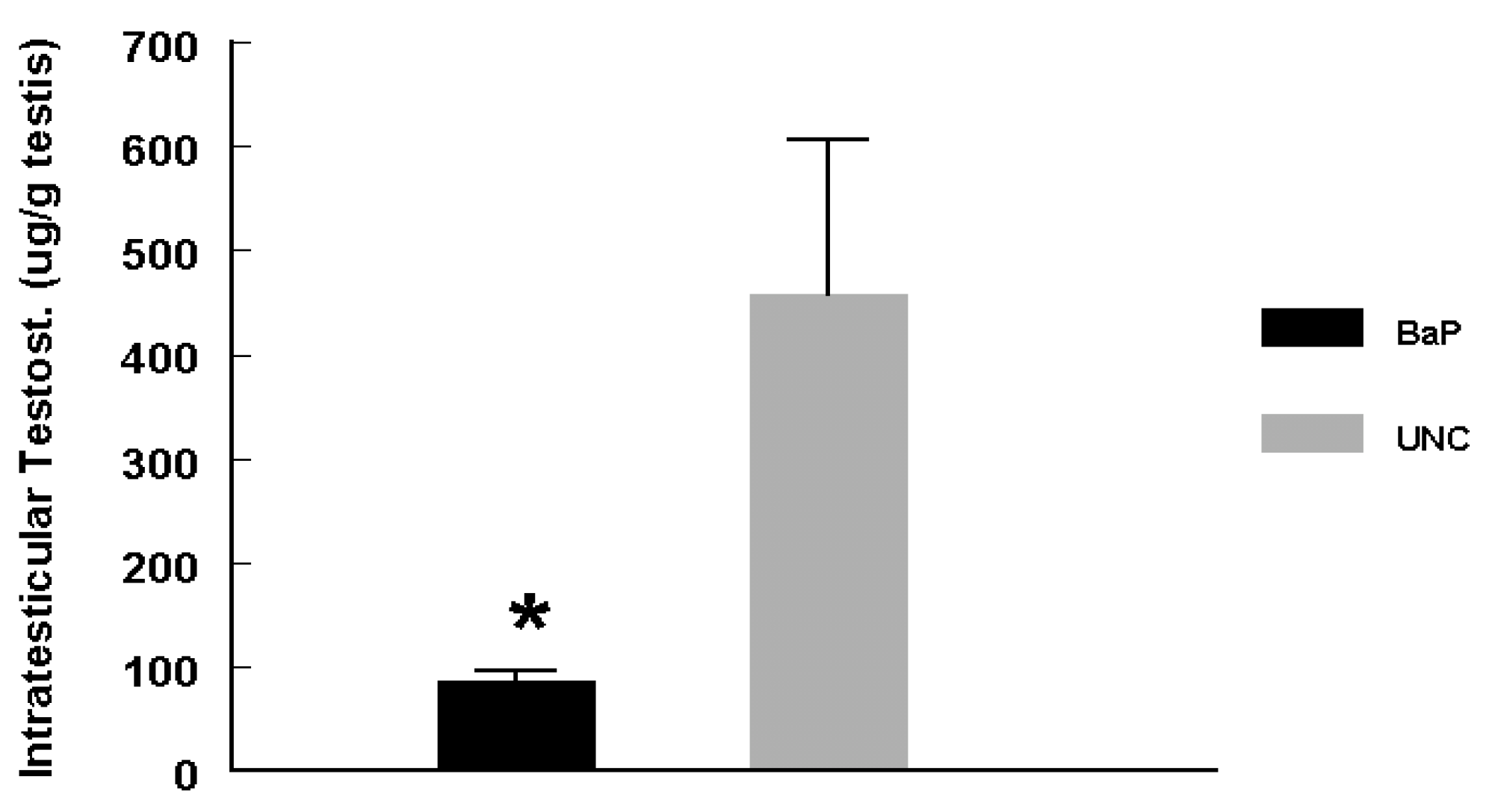
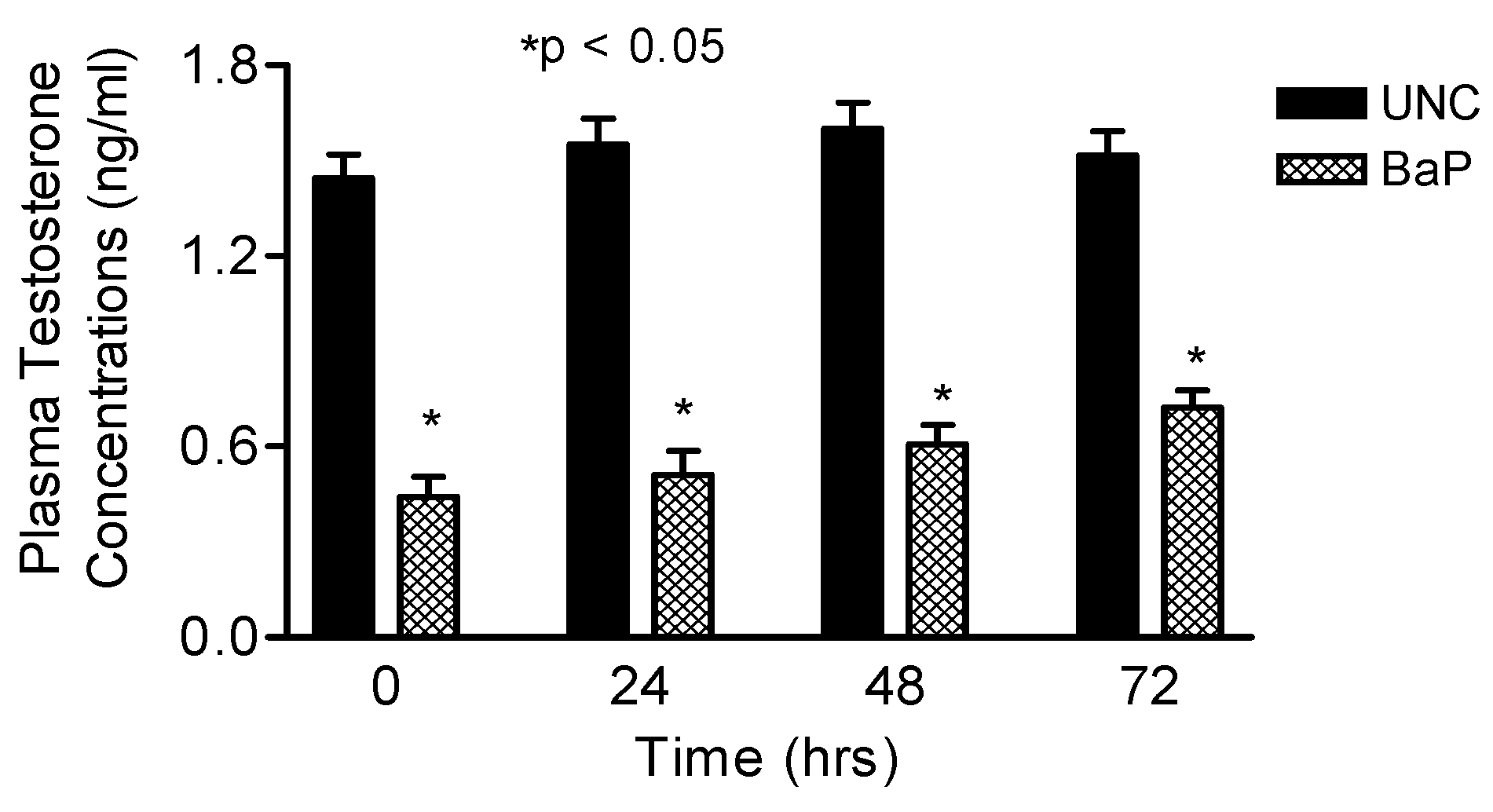
Plasma and Intratesticular Testosterone and Plasma LH Determination
Statistical Analyses
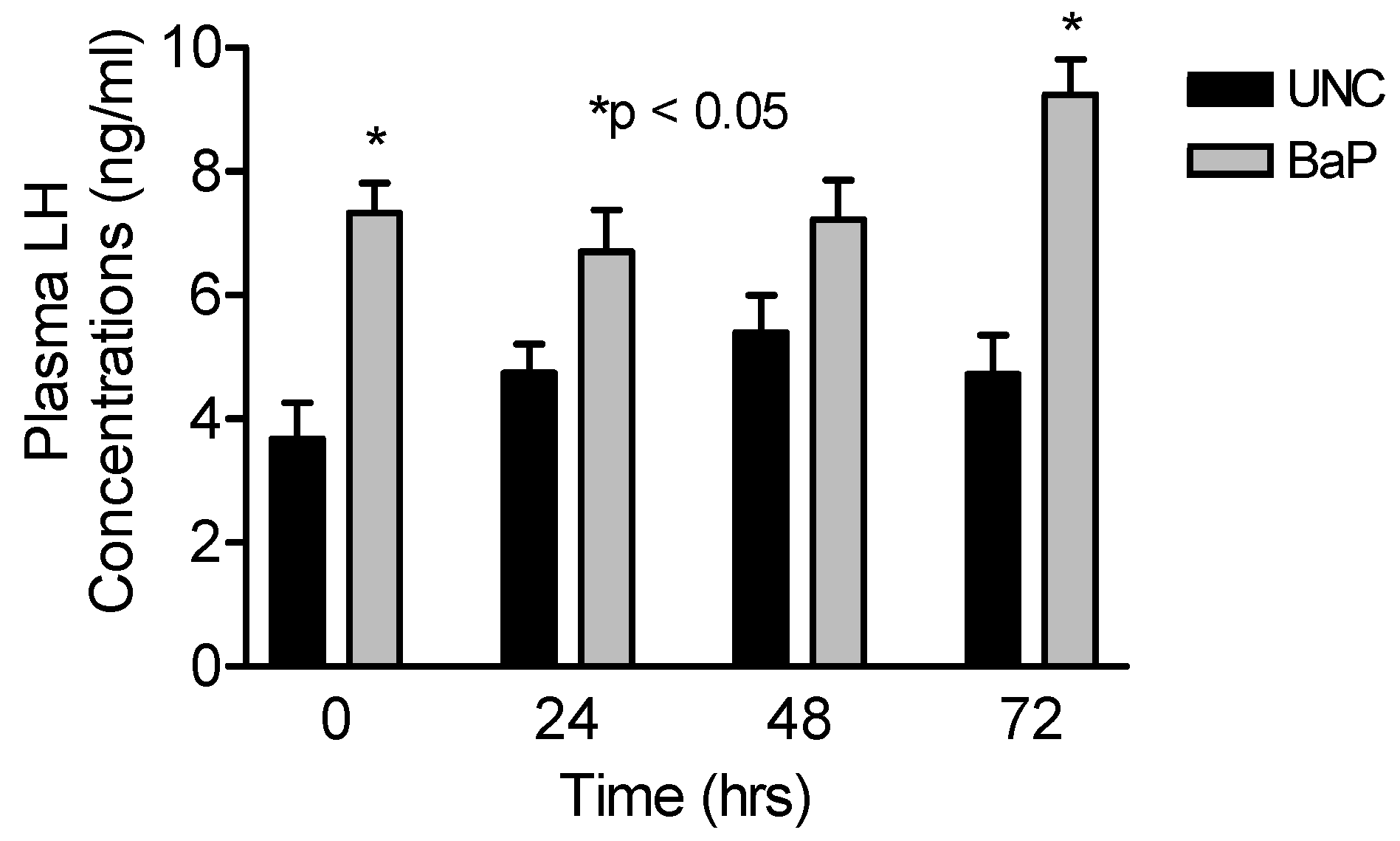
Results






Discussion
Acknowledgements
References
- Moline, J. M.; Golden, A. L.; Bar-Chama, N.; Smith, E.; Rauch, M. E.; Chapin, R. E.; Perreault, S. D.; Schrader, S. M.; Suk, W. A.; Landrigan, P.J. : Exposure to hazardous substances and male reproductive health: a research framework. Environ Health Perspect. 2000 108, 803–813. [CrossRef]
- Solomon, G.M.; Schettler, T. : Environment and health: 6. Endocrine disruption and potential human health implications. CMAJ 2000 163, 1471–1476.
- Arcand-hoy, L. D.; Nimrod, A. C.; Benson, W. H. : Endocrine-modulating substances in the environment:estrogenic effects of pharmaceutical products. Int. J. Toxicol. 1998, 17, 139–158. [Google Scholar] [CrossRef]
- Jarow, J. P.; Zirkin, B. : The androgen microenvironment of the human testis and hormonal control of spermatogenesis. Ann NY Sci 2005, 1061, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Inyang, F.; Ramesh, A.; Kopsombut, P.; Niaz, M.S.; Hood, D.B.; Nyanda, A.M.; et al. : Disruption of testicular steroidogenesis and epididymal function by inhaled benzo(a)pyrene. Reprod Toxicol 2003, 17, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Agency for Toxic Substances and Disease Registry (ATSDR). Toxicological profile for Polycyclic Aromatic Hydrocarbons (PAHs). US Department of Health & Human Services: Atlanta, Georgia; 1995, 458pp.
- WHO. Selected non-heterocyclic polycyclic aromatic hydrocarbons. Environmental Health Criteria 202. World Health Organization, Geneva, 1998, 883pp.
- Ramesh, A.; Walker, S. A.; Hood, D. B.; Guillen, M. D.; Schneider, H.; Weyand, E. H. : Bioavailability and risk assessment of orally ingested polycyclic aromatic hydrocarbons. Int. J. Toxicol. 2004, 23, 301–333. [Google Scholar] [CrossRef] [PubMed]
- Nebert, D. W; Roe, A. L.; Dieter, M. Z.; Solis, W. A; Yang, Y.; Dalton, T. P. : Role of the aromatic hydrocarbon receptor and [Ah] gene battery in the oxidative stress response, cell cycle control, and apoptosis. Biochem. Pharmacol. 2000, 59, 65–85. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Guengerich, F. P. : Inhibition of cytochrome P450 1A1-, 1A2-, and 1B1-mediated activation of procarcinogens to genotoxic metabolites by polycyclic aromatic hydrocarbons. Chem. Res. Toxicol. 2006, 19, 288–294. [Google Scholar] [CrossRef]
- Nebert, D. W; Gonzalez, F. J. : P450 genes: structure, evolution, and regulation. Annu Rev Biochem. 1987, 56, 945–993. [Google Scholar] [CrossRef]
- Flowers, L.; Ohnishi, S.; Penning, T. M. : DNA strand scission by polycyclic aromatic hydrocarbon oquinones: role of reactive oxygen species, Cu (II)/Cu (I) redox cycling, and o-semiquinone anion radicals. Biochemistry, 1997, 36, 8640–8648. [Google Scholar] [CrossRef]
- Aust, S. D.; Chignell, C. F.; Bray, T. M.; Kalyanaraman, B.; Mason, R.P. Free radicals in toxicology. Toxicol. Appl. Pharmacol. 1993, 120, 168–178. [Google Scholar] [CrossRef]
- Bolton, J. L.; Trush, M. A.; Penning, T. M; Dryhurst, G; Monks, T. J.: Role of quinones in toxicology. Chem. Res. Toxicol. 2000, 13, 135–160. [Google Scholar] [CrossRef]
- Burchiel, S. W.; Luster, M. I. : Signaling by environmental polycyclic aromatic hydrocarbons in human lymphocytes. Clin. Immunol. 2001, 98, 2–10. [Google Scholar] [CrossRef]
- McLachlan, R. I.; O’Donnell, L.; Stanton, P. G.; Balourdos, G.; Frydenberg, M.; de Kretser, D. M.; Robertson, D. M. : Effects of testosterone plus medroxyprogesterone acetate on semen quality, reproductive hormones, and germ cell populations in normal young men. J Clin Endocrinol Metab 2002, 87, 546–556. [Google Scholar]
- Matsumoto, A. M.; Bremner, W. J. ; Stimulation of sperm production by human chorionic gonadotropin after prolonged gonadotropin suppression in normal men. J Androl. 1985 6, 137–143. [CrossRef]
- Matsumoto, A. M. : Effects of chronic testosterone administration in normal men: safety and efficacy of high dosage testosterone and parallel dose-dependent suppression of luteinizing hormone, follicle- stimulating hormone, and sperm production. J Clin Endocrinol Metab. 1990 70, 282–287. [CrossRef]
- Morse, H. C.; Horike, N.; Rowley, M. J.; Heller, C. G. : Testosterone concentrations in testes of normal men: Effects of testosterone propionate administration. J Clin Endocrinol Metab. 1973, 37, 882–886. [Google Scholar] [CrossRef] [PubMed]
- Huhtaniemi, I.; Nikula, H.; Rannikko, S. : Pituitary- testicular function of prostatic cancer patients during treatment with a gonadotropin-releasing hormone agonist analog. I. Circulating hormone levels. J Androl. 1987, 8, 355–362. [Google Scholar] [CrossRef] [PubMed]
- WHO. Contraceptive efficacy of testosterone-induced azoospermia in normal men. Lancet. 1990, 336, 995–999. [Google Scholar]
- Hill, C. M.; Anway, M. D.; Zirkin, B. R.; Brown, T. R. : Intratesticular androgen levels, androgen receptor localization, and androgen receptor expression in adult rat Sertoli cells. Biol. Reprod. 2004, 71, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Hood, D. B.; Nayyar, T.; Ramesh, A.; Greenwood, M.; Inyang, F. : Modulation in the developmental expression profile of Sp1 subsequent to transplacental exposure of fetal rats to desorbed benzo(a)pyrene following maternal inhalation. Inhal Toxicol, 2000, 12, 511–535. [Google Scholar] [PubMed]
- Accardi-Dey, A.; Gschwend, P. M. : Reinterpreting literature sorption data considering both absorption into organic carbon and adsorption onto black carbon. Environ Sci Technol 2003, 37, 99–106. [Google Scholar]
- Anon. Carbon black users guide. Safety, health and environmental information, International Carbon Black Association, 2004.
- Archibong, A. E.; Inyang, F.; Ramesh, A.; Greenwood, M.; Nayyar, T.; Kopsombut, P.; et al. : Alteration of pregnancy related hormones and fetal survival in F-344 rats by inhaled benzo(a)pyrene. Reprod Toxicol, 2002, 16, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Weibel, E. R. ; Stereological techniques for electron microscopic morphometry. In: Hayat, M. A.: ed. Principles and Techniques of Electron Microscopy, Vol 3, Biological Applications. Van Nostrand Reinhold Co., New York; 1973: pp.237-296.
- Lunstra, D. D.; Ford, J. J.; Christenson, R. K.; Allrich, R. D. : Changes in Leydig cell ultrastructure and function during pubertal development in the boar. Biol Reprod 1986, 34, 145–158. [Google Scholar]
- Jiménez-Severiano, H.; Mussard, M. L.; Fitzpatrick, L. A.; D’Occhio, M. J.; Ford, J. J.; Lunstra D., D.; et al. : Testicular development of Zebu bulls after chronic treatment with a gonadotropin-releasing hormone agonist. J. Anim Sci, 2005, 83, 2111–2122. [Google Scholar] [CrossRef] [PubMed]
- Okwun, O. E.; Igboeli, G.; Ford, J. J.; Lunstra, D. D.; Johnson, L. : Sertoli cell number and function, spermatogonial number and yield and daily sperm production in three breeds of boars. J. Reprod Fertil 1996, 107, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Goyal, H. O.; Braden, T. D.; Mansour, M.; Williams, C. S.; Kamaleldin, A.; Srivastava, K. K. : Diethylstilbestrol-treated adult rats with altered epididymal sperm numbers and sperm motility parameters, but without alterations in sperm production and sperm morphology. Biol Reprod. 2001, 64, 927–934. [Google Scholar] [CrossRef]
- Robb, G. W.; Amann, R. P.; Killian, G. J. : Daily sperm production and epididymal sperm reserves of pubertal and adult rats. J Reprod Fertil. 1978, 54, 103–107. [Google Scholar] [CrossRef]
- SAS/STAT. SAS Institute SAS/STAT Statistical Analysis System User’s Guide (Version 6, 5th Edition). Cary, North Carolina: Statistical Analysis System Institute, Inc. 1990, pp.113-709.
- Chorazy, M.; Szeliga, J.; Strozyk, M.; Cimander, B. : Ambient air pollutants in Upper Silesia: partial chemical composition and biological activity. Environ Health Perspect. 1994, 102 (Suppl 4), 61–66. [Google Scholar]
- Viau, C.; Hakizimana, G.; Bouchard, M. : Indoor exposure to polycyclic aromatic hydrocarbons and carbon monoxide in traditional houses in Burundi. Int Arch Occup Environ Health, 2000, 73, 331–338. [Google Scholar]
- Occupational Safety and Health Administration (OSHA). Occupational Safety and Health Standards, Toxic and Hazardous Substances. Code of Federal Regulations. 29 CFR 1910.1000. 1998.
- Blazak, W. F.; Ernst, T. L.; Stewart, B. E. : Potential indicators of reproductive toxicity: testicular sperm production and epididymal sperm number, transit time, and motility in Fischer 344 rats. Fundam Appl Toxicol 1985, 5, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Tate, F. : Antispermatogenic effects of ethylmethanesulfonate and benzo(a)pyrene in PD4 Lakeview hamsters. J. Toxicol. Environ. Health, 1981; 8, 929–937. [Google Scholar]
- Lipshultz, L. I.; Corriere, J. N Jr.: Progressive testicular atrophy in the varicocele patient. J Urol 1977, 117, 175–176. [Google Scholar] [CrossRef]
- Lipshultz, L. I.; Witt, M. A. : Infertility in the male. In: Hammond, M. G, Talbert L M, editors. Infertility, a practical guide for the physician. Blackwell Scientific Publications, Boston; pp. 26-55, 1993.
- Raychoudhury, S. S.; Kubinski, D. : Polycyclic aromatic hydrocarbon induced cytotoxicity in cultured rat sertoli cells involves differential apoptotic response. Environ. Hlth. Perspect, 2003, 111, 33–38. [Google Scholar] [CrossRef]
- Ku, W. W.; Chapin, R. E. : Preparation and use of Sertoli cell-enriched cultures from 18-Day - old rat. In: Chapin, R. E.; Heindel, J. J.: ed. Male Reproductive Toxicology, Methods in Toxicology, Academic Press, San Diego: 1993, Vol. 3, pp. 210-229.
- Revel, A.; Roanani, H.; Younglai, E.; Xu, J.; Han, R.; Savouret, J-F et al. Resveratrol, a natural aryl hydrocarbon receptor antagonist, protects sperm from DNA damage and apoptosis caused by benzo(a)pyrene. Reprod. Toxicol. 2001, 15, 479–486. [Google Scholar] [CrossRef]
- Plant, T. M.; Marshall, G. R. : The functional significance of FSH in spermatogenesis and the control of its secretion in male primates. Endocrine Reviews, 2001, 22, 764–786. [Google Scholar] [CrossRef] [PubMed]
- Page, S. T.; Kalhorn, T. F.; Bremner, W. J.; Anawalt, B. D.; Matsumoto, A. M.; Amory, J. K. : Intratesticular androgens and spermatogenesis during severe gonadotropin suppression induced by male hormonal contraceptive treatment. J. Androl, 2007, 28, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Polyakov, L. M.; Chasovskikh, M. I.; Panin, L. E. : Binding and transport of benzo(a)pyrene by blood plasma lipoproteins: the possible role of apolipoprotein B in this process. Bioconjugate Chem 1996, 7, 396–400. [Google Scholar] [CrossRef]
- Mandal, P. K.; McDaniel, L. R.; Prough, R. A.; Clark, B. J. : 7, 12- Dimethlbenz(a)anthracene inhibition of steroid production in MA-10 mouse Leydig tumor cells is not directly linked to induction of CYP1B1. Toxicol Appl Pharmacol., 2001, 175, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, A.; Inyang, F.; Hood, D. B.; Knuckles, M. E. : Aryl hydrocarbon hydroxylase activity in F-344 rats subchronically exposed to benzo(a)pyrene and fluoranthene through diet. J Biochem Mol Toxicol., 2000, 14, 155–161. [Google Scholar] [CrossRef]
- Ramesh, A.; Hood, D. B.; Inyang, F.; Greenwood, M.; Archibong, A.E.; Knuckles, M. E et al.: Comparative metabolism, bioavailability and toxicokinetics of benzo(a)pyrene in rats after acute oral, inhalation, and intravenous administration. Polycyclic Aromatic Compounds, 2002, 22, 969–980. [Google Scholar]
- Lee, I. P.; Nagayama, J. : Metabolism of benzo(a)pyrene by the isolated perfused rat testis. Cancer Res., 1980, 40, 3297–3303. [Google Scholar] [PubMed]
- Williams, J. A.; Martin, F. C.; Muir, G. H.; Hewer, A.; Grover, P. L.; Phillips, D. H. : Metabolic activation of carcinogens and expression of various cytochromes P450 in human prostate tissue. Carcinogenesis, 2000, 21, 1683–1689. [Google Scholar] [CrossRef]
- Senft, A. P.; Dalton, T. P.; Nebert, D. W.; Genter, M. B.; Puga, A.; Hutchinson, R. J, et al.: Mitochondrial reactive oxygen production is dependent on the aromatic hydrocarbon receptor. Free Radic Biol Med, 2002, 33, 1268–1278. [Google Scholar] [CrossRef]
- Peltola, V.; Huhtaniemi, I.; Metsa-Ketela, T.; Ahotupa, M. : Induction of lipid peroxidation during steroidogenesis in the rat testis. Endocrinology 1996, 137, 105–112. [Google Scholar] [CrossRef]
- Diemer, T.; Allen, J. A.; Hales, K. H.; Hales, D. B. : Reactive oxygen disrupts mitochondriain MA-10 tumor Leydig cells and inhibits steroidogenic acute regulatory (StAR) protein and steroidogenesis. Endocrinology, 2003, 144, 2882–2891. [Google Scholar] [CrossRef]
- Yardimci, S.; Atan, A.; Delibasi, T.; Sunguroglu, K.; Guven, M. C. : Long-term effects of cigarette-smoke exposure on plasma testosterone, luteinizing hormone and follicle-stimulating hormone levels in male rats. Br J Urol., 1997, 79, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Jeyaraj, D. A.; Grossman, G.; Petrusz, P. : Altered bioavailability of testosterone in androgen-binding protein-transgenic mice. Steroids, 2005, 70, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Karabelyos, C. S.; Csaba, G. : Benzpyrene treatment decreases the sexual activity of adult rats, what is reversed in neonatally allylestrenol treated animals. Acta Physiol Hung, 1996, 84, 131–137. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | BaP-exposed |
| Tubule diameter (μm) | 250 ± 8.5 | 230 ± 8.2 |
| Tubules with elongated spermatids (%) | 99 ± 1.0 | 99 ± 1.0 |
| Tubular volumne (X 109 μm3) | 2.0 ± 0.7 | 1.6 ± 0.004*** |
| Total weight of tubules (gm) | 2.20 ± 0.7 | 1.6 ± 0.004*** |
| Total tubular length (μm) | 55 ± 1.0 | 33+1.0** |
| Total volume of interstitium per paired testis (μm3) | 0.43 ± 0.01 | 0.38 ± 0.01* |
| Total weight of interstitium per paired testis (gm) | 0.45 ± 0.01 | 0.37 ± 0.01* |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2008 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Archibong, A.E.; Ramesh, A.; Niaz, M.S.; Brooks, C.M.; Roberson, S.I.; Lunstra, D.D. Effects of Benzo(a)pyrene on Intra-testicular Function in F-344 Rats. Int. J. Environ. Res. Public Health 2008, 5, 32-40. https://doi.org/10.3390/ijerph5010032
Archibong AE, Ramesh A, Niaz MS, Brooks CM, Roberson SI, Lunstra DD. Effects of Benzo(a)pyrene on Intra-testicular Function in F-344 Rats. International Journal of Environmental Research and Public Health. 2008; 5(1):32-40. https://doi.org/10.3390/ijerph5010032
Chicago/Turabian StyleArchibong, Anthony E., Aramandla Ramesh, Mohammad S. Niaz, Cynthia M. Brooks, Shannon I. Roberson, and Donald D. Lunstra. 2008. "Effects of Benzo(a)pyrene on Intra-testicular Function in F-344 Rats" International Journal of Environmental Research and Public Health 5, no. 1: 32-40. https://doi.org/10.3390/ijerph5010032