Leg Length, Body Proportion, and Health: A Review with a Note on Beauty

Abstract
:
1. Introduction
2. Leg Length Defined
3. Practical Methods and Techniques
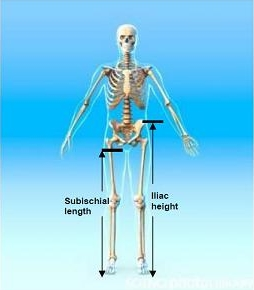
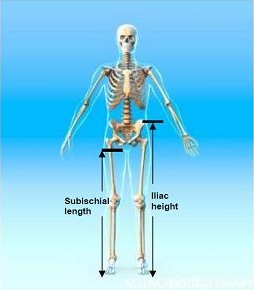
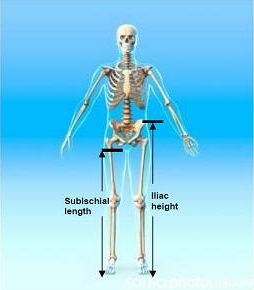
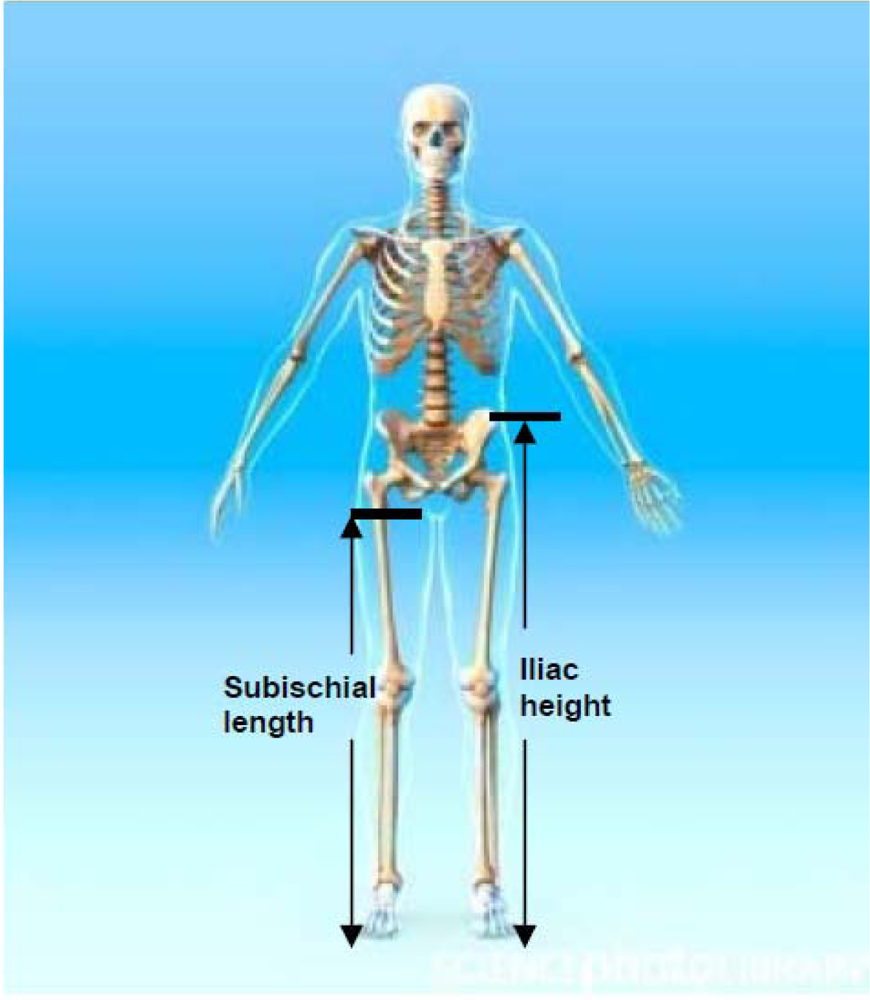
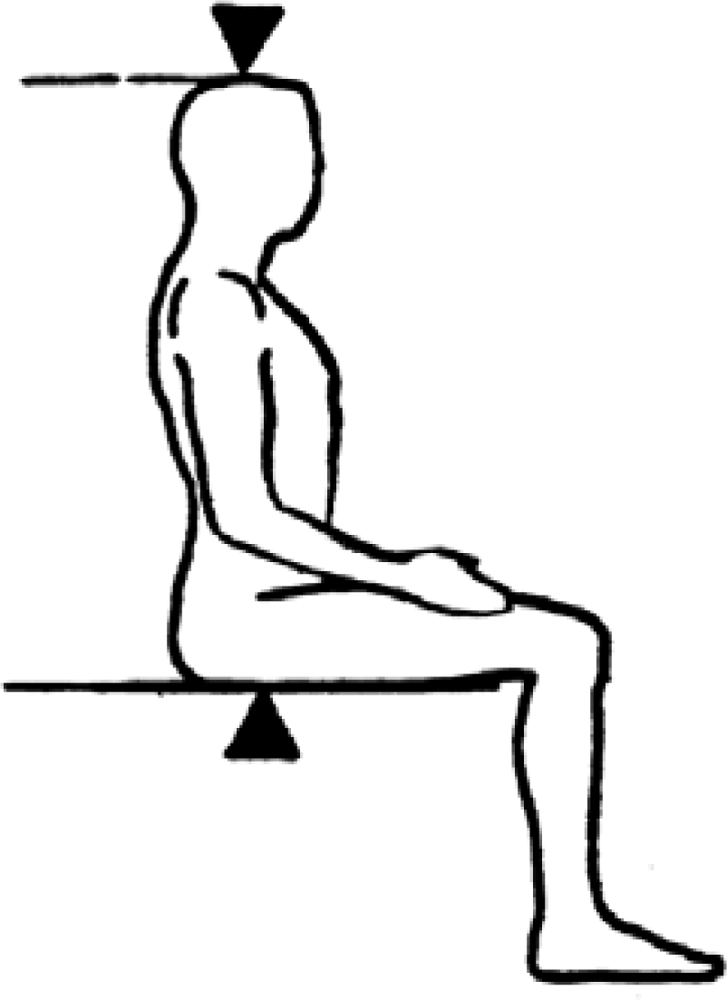
3.1. Iliac Height (IH)
3.2. Subischial Leg Length (SLL)
3.3. Thigh Length (TL)
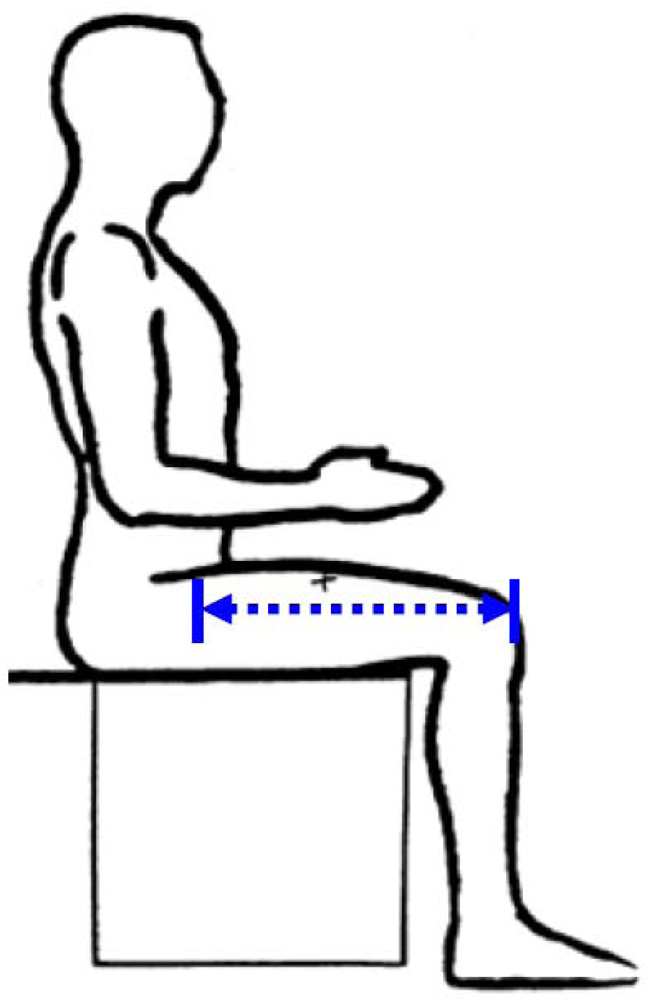
3.4. Knee Height (KH)
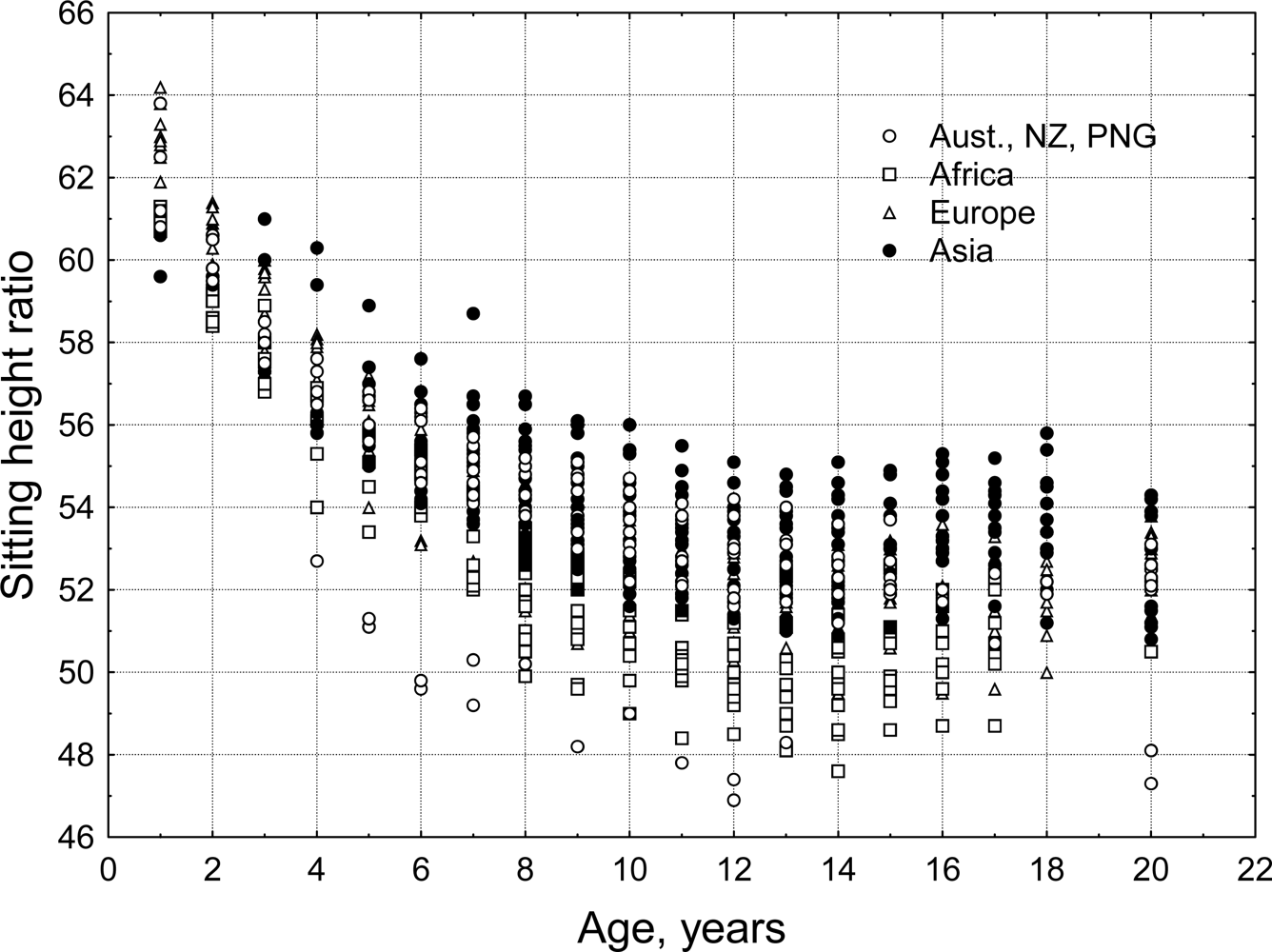
3.5. Sitting Height Ratio (SHR)
3.6. Relative Subischial Leg Length (RSLL)
3.7. Knee Height Ratio (KHR)
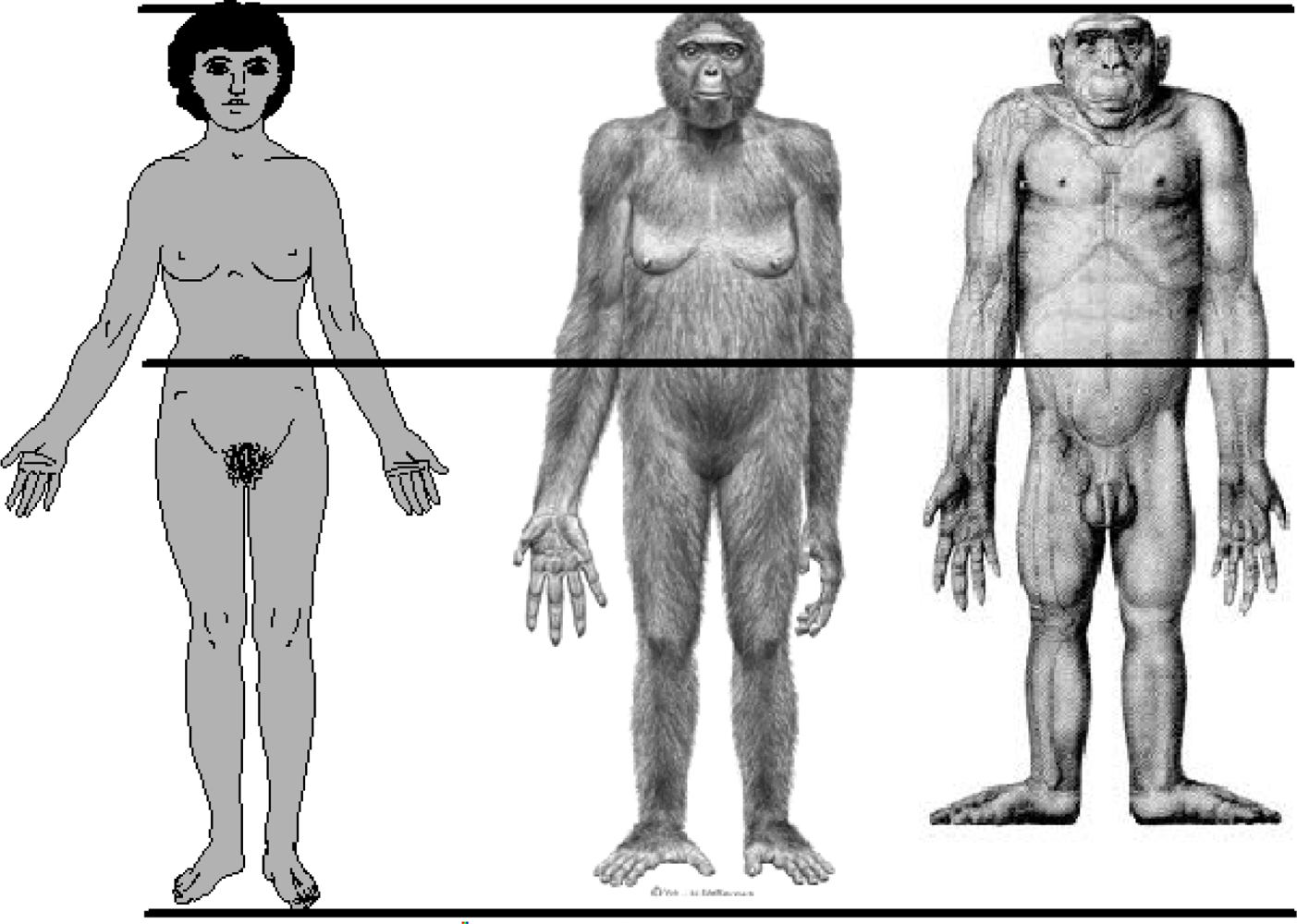
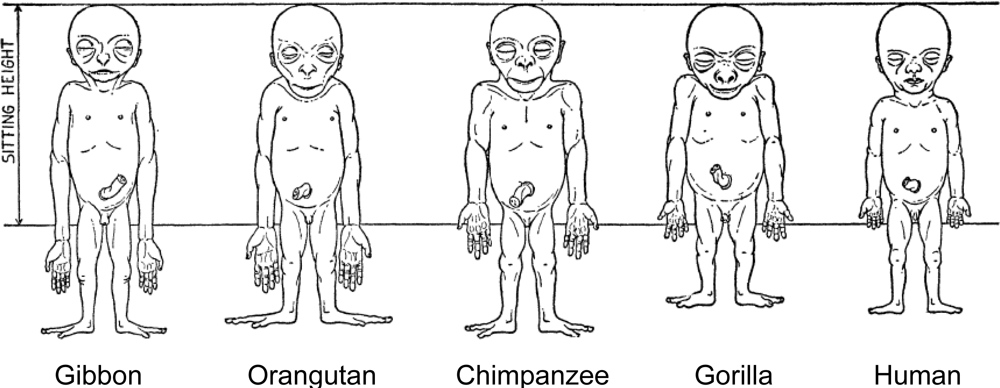
4. Evolutionary Background of Human Body Shape
5. Size and Shape in Living Humans
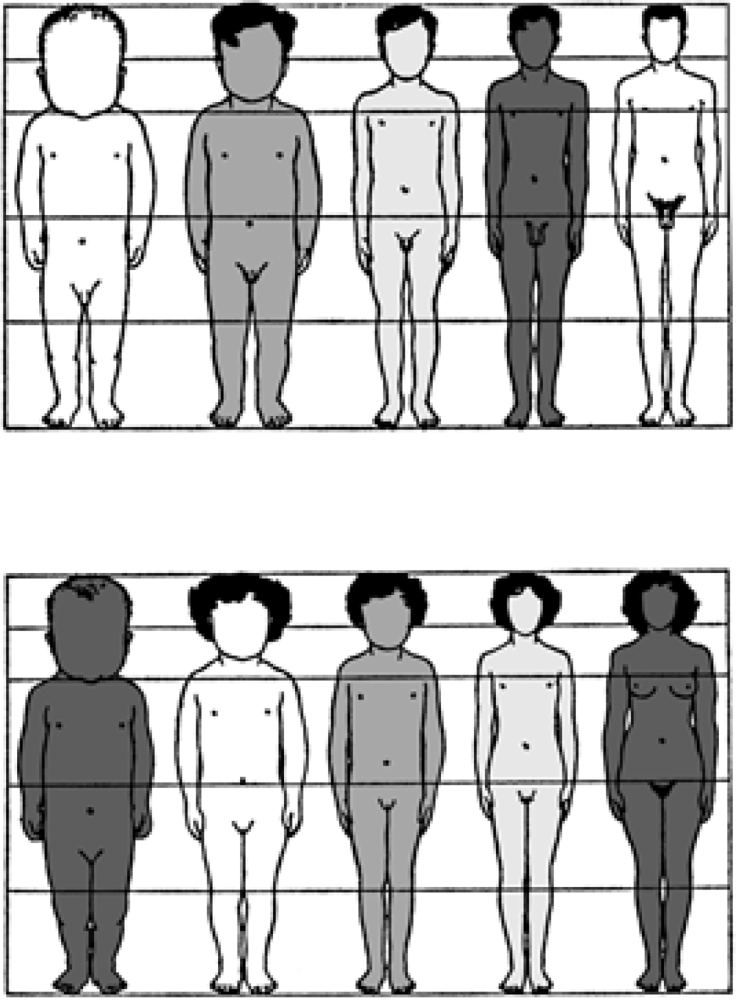
6. Developmental Plasticity
7. The Use of Leg Length in Human Biology and Environmental Epidemiology
7.1. Leg Length and Human Environmental Health
7.2. Leg Length and Risk for Morbidity and Mortality
8. Leg Length and Beauty
9. Conclusion
References
- Gould, SJ. The Mismeasure of Man; Norton: New York, NY, USA, 1981. [Google Scholar]
- Marks, J. Human Biodiversity: Genes, Race, and History; Aldine De Gruyter: New York, NY, USA, 1995. [Google Scholar]
- Coon, C. The Origin of Races; Knopf: New York, NY, USA, 1962. [Google Scholar]
- Tishkoff, SA; Kidd, KK. Implications of biogeography of human populations for ‘race’ and medicine. Nat. Genet 2004, 36, S21–S27. [Google Scholar]
- Ramachandran, S; Deshpande, O; Roseman, CC; Rosenberg, NA; Feldman, MW; Cavalli-Sforza, LL. Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa. Proc. Natl. Acad. Sci. U.S.A 2005, 102, 15942–15947. [Google Scholar]
- Tattersall, I. Out of Africa: modern human origins special feature: human origins: out of Africa. Proc. Natl. Acad. Sci. U.S.A 2009, 106, 16018–16021. [Google Scholar]
- Shea, BT; Bailey, RC. Allometry and adaptation of body proportions and stature in African pygmies. Amer. J. Phy. Anthropol 1996, 100, 311–340. [Google Scholar]
- Lohman, TG; Roche, AF; Martorell, R. Anthropometric Standardization Reference Manual; Human Kinetics Publishers: Champaign, IL, USA, 1988. [Google Scholar]
- Bogin, B; Varela-Silva, MI. Fatness biases the use of estimated leg length as an epidemiological marker for adults in the NHANES III sample. Int. J. Epidemiol 2008, 8, 201–209. [Google Scholar]
- Frisancho, AR. Anthropometric Standards An Interactive Nutritional Reference of Body Size and Body Composition for Children and Adults; The University of Michigan Press: Ann Arbor, MI, USA, 2008. [Google Scholar]
- Bogin, B. Patterns of Human Growth, 2nd ed; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Darwin, C. The Descent of Man, and Selection in Relation to Sex; John Murray: London, UK, 1981. [Google Scholar]
- Underwood, CR; Ward, EJ. The solar radiation area of man. Ergonomics 1966, 9, 155–168. [Google Scholar]
- Newman, RW. Why man is such a sweaty and thirsty naked animal: a speculative review. Hum. Biol 1970, 42, 12–27. [Google Scholar]
- Ruff, C. Variation in human body size and shape. Ann. Rev. Anthropol 2002, 31, 211–232. [Google Scholar]
- Frisancho, AR. Human Adaptation and Accommodation; University of Michigan Press: Ann Arbor, MI, USA, 1993. [Google Scholar]
- Zihlman, A. Woman the gatherer: the role of women in early hominid evolution. In Gender and Anthropology: Critical Reviews for Teaching and Research; Sandra, M, Ed.; American Anthropological Association: Washington, DC, USA, 1989; pp. 23–43. [Google Scholar]
- Bramble, DM; Lieberman, DE. Endurance running and the evolution of Homo. Nature 2004, 18, 345–352. [Google Scholar]
- Corballis, MC. From Hand to Mouth: The Origins of Language; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Aiello, L; Dean, MC. Human Evolutionary Anatomy; Academic Press: London, UK, 1990. [Google Scholar]
- Scammon, RE; Calkins, LA. The Development and Growth of the External Dimensions of the Human Body in the Fetal Period; University of Minnesota Press: Minneapolis, MN, USA, 1929. [Google Scholar]
- Scammon, RE. The measurement of the body in childhood. In The Measurement of Man; Harris, JA, Jackson, CM, Paterson, DG, Scammon, RE, Eds.; University of Minnesota Press: Minneapolis, MN, USA, 1930; pp. 173–215. [Google Scholar]
- Schultz, AH. Fetal growth of man and other primates. Quart. Rev. Biol 1926, 1, 465–521. [Google Scholar]
- Martin, RD. Human Brain Evolution in an Ecological Context (Fifty-second James Arthur Lecture); American Museum of Natural History: New York, NY, USA, 1983. [Google Scholar]
- Leigh, SR. Brain growth, life history, and cognition in primate and human evolution. Amer. J. Primatol 2004, 62, 139–164. [Google Scholar]
- Leonard, WR; Robertson, ML. Evolutionary perspectives on human nutrition: the influence of brain and body size on diet and metabolism. Amer. J. Hum. Biol 1994, 6, 77–88. [Google Scholar]
- Klingenberg, CP; Nijhout, HF. Competition among growing organs and developmental control of morphological asymmetry. Proc. R. Soc. Lond 1998, 265, 1135–1139. [Google Scholar]
- Charnov, EL. Life History Invariants; Oxford University Press: Oxford, UK, 1993. [Google Scholar]
- Stearns, SC. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Bailey, SM; Xu, J; Feng, JH; Hu, X; Zhang, C; Qui, S. Tradeoffs between oxygen and energy in tibial growth at high altitude. Amer. J. Hum. Biol 2007, 19, 662–668. [Google Scholar]
- Bogin, B; Varela Silva, MI; Rios, L. Life history trade-offs in human growth: adaptation or pathology? Amer. J. Hum. Biol 2007, 19, 631–642. [Google Scholar]
- Rauch, F. Bone growth in length and width: the Yin and Yang of bone stability. J. Musculoskelet. Neuronal Interact 2005, 5, 194–201. [Google Scholar]
- Tanner, JM. A historical perspective on human auxology. Humanbiol. Budapest 1994, 25, 9–22. [Google Scholar]
- Norgard, EA; Jarvis, JP; Roseman, CC; Maxwell, TJ; Kenney-Hunt, JP; Samocha, KE; Pletscher, LS; Wang, B; Fawcett, GL; Leatherwood, CJ; Wolf, JB; Cheverud, JM. Replication of long bone length QTL in the F9−F10 LG,SM advanced intercross. Mamm Genome 2009, 20, 224–235. [Google Scholar]
- Reno, PL; McCollum, MA; Cohn, MJ; Meindl, RS; Hamrick, M; Lovejoy, CO. Patterns of correlation and covariation of anthropoid distal forelimb segments correspond to Hoxd expression territories. J. Exp. Zool. (Mol. Dev. Evol.) 2008, 310B, 240–258. [Google Scholar]
- Kajantie, E. Insulin-like growth factor (IGF)-I, IGF binding protein (IGFBP)-3, phosphoisoforms of IGFBP-1 and postnatal growth in very-low-birth-weight infants. Horm Res 2003, 60, 124–130. [Google Scholar]
- Serrat, MA; Lovejoy, CO; King, D. Age- and site-specific decline in insulin-like growth factor-1 receptor expression is correlated with differential growth plate activity in the mouse hindlimb. Anatom. Record 2007, 290, 375–381. [Google Scholar]
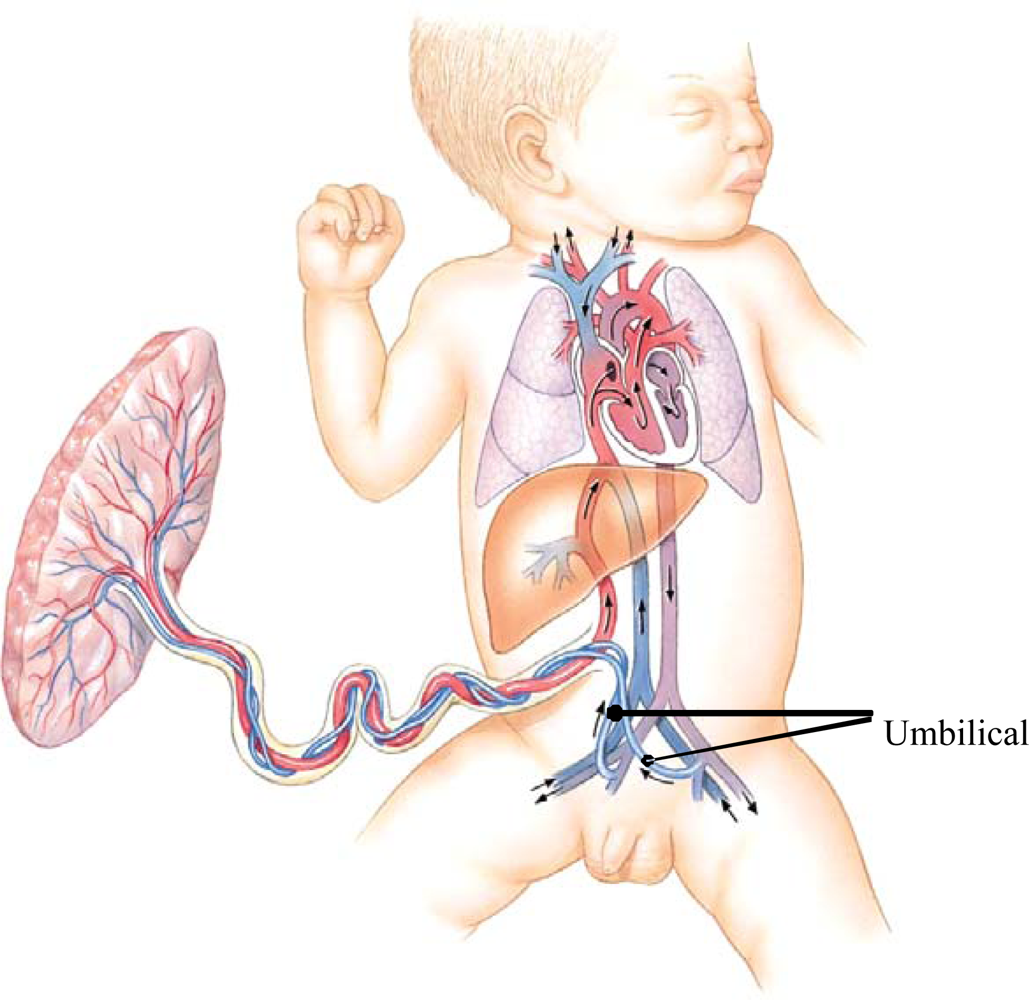
- Boros, SJ; Nystrom, J; Thompson, T; Reynolds, J; Williams, H. Leg growth following umbilical artery catheter-associated thrombus formation: a 4-year follow-up. J. Pediatrics 1975, 87, 973–976. [Google Scholar]
- Martini, FH; Bartholomew, EF. Essentials of Anatomy & Physiology; Pearson Education: San Francisco, CA, USA, 2007. [Google Scholar]
- Boyd, E. Origins of the Study of Human Growth; Savara, BS, Schilke, JF, Eds.; University of Oregon Press: Eugene, OR, USA, 1980. [Google Scholar]
- Tanner, JM. A History of the Study of Human Growth; University of Cambridge Press: Cambridge, UK, 1981. [Google Scholar]
- Tate, C; Bendersky, G. Olmec sculptures of the human fetus. Perspect. Biol. Med 1999, 42, 303–332. [Google Scholar]
- Dietz, WH; Marino, B; Peacock, NR; Bailey, RC. Nutritional status of Efe pygmies and Lese horticulturalists. Am. J. Phys. Anthropol 1989, 78, 509–518. [Google Scholar]
- Fredriks, AM; van Buuren, S; Burgmeijer, RJ; Meulmeester, JF; Beuker, RJ; Brugman, E; Roede, MJ; Verloove-Vanhorick, SP; Wit, JM. Continuing positive secular growth change in The Netherlands 1955–1997. Pediatr. Res 2000, 47, 316–323. [Google Scholar]
- Eveleth, PB; Tanner, JM. Worldwide Variation in Human Growth; Cambridge University Press: Cambridge, UK, 1976. [Google Scholar]
- Eveleth, PB; Tanner, JM. Worldwide Variation in Human Growth, 2nd ed; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Bergmann, K. Über die Verhältnisse der Wärmeökonomie der Thiere zu ihrer Grösse. Göttinger Studien 1847, 3, 95–108. [Google Scholar]
- Allen, JA. The influence of physical conditions in the genesis of species. Radical Review 1877, 1, 108–140. [Google Scholar]
- Serrat, MA; King, D; Lovejoy, CO. Temperature regulates limb length in homeotherms by directly modulating cartilage growth. PNAS 2008, 105, 19348–19353. [Google Scholar]
- Roberts, DF. Bodyweight, race, and climate. Am. J. Phys. Anthropol 1953, 11, 533–558. [Google Scholar]
- Roberts, DF. Climate and Human Variability, 2nd ed; Cummings: Menlo Park, CA, USA, 1978. [Google Scholar]
- Katzmarzyk, PT; Leonard, WR. Climatic influences on human body size and proportions: ecological adaptations and secular trends. Am. J. Phy. Anthrop 1998, 106, 483–503. [Google Scholar]
- Bogin, B; Keep, R. Eight thousand years of economic and political history in Latin America revealed by anthropometry. Ann. Hum. Biol 1999, 26, 333–351. [Google Scholar]
- Andersen, H. The influence of hormones on human development. In Human Development; Falkner, F, Ed.; W.B. Saunders: Philadelphia, PA, USA, 1966; pp. 184–221. [Google Scholar]
- Kramer, K. Variation in juvenile dependence: helping behavior among Maya children. Hum. Nat 2002, 13, 299–325. [Google Scholar]
- Bogin, B; Smith, PK; Orden, AB; Varela Silva, MI; Loucky, J. Rapid change in height and body proportions of Maya American children. Am. J. Hum. Biol 2002, 14, 753–761. [Google Scholar]
- Fulwood, R; Abraham, S; Johnson, C. Height and weight of adults ages 18−74 years by socioeconomic and geographic variables. In Vital and Health Statistics; Series 11, No. 224, DHEW Pub. No. (PHS) 81-1674. U.S. Government Printing Office: Washington, DC, USA, 1981. [Google Scholar]
- Krogman, WM. Growth of the head, face, trunk, and limbs in Philadelphia white and Negro children of elementary and high school age. Monog. Soc. Res. Child Develop 1970, 20, 1–91. [Google Scholar]
- Hamill, PVV; Johnston, FE; Lemshow, S. Body weight, stature, and sitting height: white and Negro youths 12–17 years, United States. DHEW Publication No. (HRA) 74-1608. U.S. Government Printing Office: Washington, DC, USA, 1973. [Google Scholar]
- Livshits, G; Roset, A; Yakovenko, K; Trofimov, S; Kobyliansky, E. Genetics of human body size and shape: body proportions and indices. Ann. Hum. Biol 2002, 29, 271–289. [Google Scholar]
- Bogin, B; Kapell, M; Varela Silva, MI; Orden, AB; Smith, PK; Loucky, J. How genetic are human body proportions? In Perspectives in Human Growth, Development and Maturation; Dasgupta, P, Hauspie, R, Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 205–221. [Google Scholar]
- Aulchenko, YS; Struchalin, MV; Belonogova, NM; Axenovich, TI; Weedon, MN; Hoffman, A; Uitterlinden, AG; Kayser, M; Oostra, BA; van Duijn, CM; Janssens, AC; Borodin, PM. Predicting human height by Victorian and genomic methods. Eur. J. Hum. Genet 2009, 17, 1070–1075. [Google Scholar]
- Holliday, TW; Falsetti, AB. A new method for discriminating African–American from European–American skeletons using postcranial osteometrics reflective of body shape. J. Forensic Sci 1999, 44, 926–930. [Google Scholar]
- Martorell, R; Malina, RM; Castillo, RO; Mendoza, FS. Body proportions in three ethnic groups: children and youths 2−17 years in NHANES and HHANES. Hum. Biol 1988, 60, 205–222. [Google Scholar]
- Feldesman, MR; Fountain, RL. “Race” specificity and the femur/stature ratio. Amer. J. Phys. Anthropol 1996, 100, 207–224. [Google Scholar]
- Mark, M; Rijli, FM; Chambon, P. Homeobox genes in embryogenesis and pathogenesis. Pediatric Res 1997, 42, 421–429. [Google Scholar]
- Blum, WF; Crowe, BJ; Quigley, CA; Jung, H; Cao, D; Ross, JL; Braun, L; Rappold, G; SHOX Study Group. Growth hormone is effective in treatment of short stature associated with short stature homeobox-containing gene deficiency: Two-year results of a randomized, controlled, multicenter trial. J. Clin. Endocrinol. Metab 2007, 92, 219–228. [Google Scholar]
- Neufeld, ND; Lippe, BM; Kaplan, SA. Disproportionate growth of the lower extremities. A major determinant of short stature in Turner’s syndrome. Am. J. Dis. Child 1978, 132, 296–298. [Google Scholar]
- Ogata, T; Inokuchi, M; Ogawa, M. Growth pattern and body proportion in a female with short stature homeobox-containing gene overdosage and gonadal estrogen deficiency. Europ. J. Endocrinol 2002, 147, 249–254. [Google Scholar]
- Hughes, PC; Ribeiro, J; Hughes, IA. Body proportions in Turner’s syndrome. Arch. Dis. Child 1986, 61, 506–507. [Google Scholar]
- Anderssen, L; Haley, CS; Ellegren, H; Knott, SA; Johansson, M; Andersson, K; Andersson-Eklund, L; Edfors-Lilja, I; Fredholm, M; Hansson, I; Hakansson, J; Lundstrom, K. Genetic mapping of quantitative trait loci for growth and fatness in pigs. Science 1994, 262, 1771–1774. [Google Scholar]
- Quignon, P; Schoenebeck, JJ; Chase, K; Parker, HG; Mosher, DS; Johnson, GS; Lark, KG; Ostrander, EA. Fine mapping a locus controlling leg morphology in the domestic dog. Quant Biol 2009. [Google Scholar]
- Tost, J. DNA methylation: an introduction to the biology and the disease-associated changes of a promising biomarker. Methods Mol. Biol 2009, 507, 3–20. [Google Scholar]
- Lasker, GW. Human biological adaptability. Science 166, 1480–1486.
- Hales, CN; Barker, DJ. Type 2 (non-insulin-dependent) diabetes mellitus: the thrifty phenotype hypothesis. Diabetologia 1992, 35, 595–601. [Google Scholar]
- Wells, JCK. The thrifty phenotype as an adaptive maternal effect. Biol. Rev 2007, 82, 143–172. [Google Scholar]
- Emanuel, I. Maternal health during childhood and later reproductive performance. Ann. NY Acad. Sci 1986, 477, 27–39. [Google Scholar]
- Varela-Silva, MI; Frisancho, AR; Bogin, B; Chatkoff, D; Smith, P; Dickinson, F; Winham, D. Behavioral, environmental, metabolic and intergenerational components of early life undernutrition leading to later obesity in developing nations and in minority groups in the USA. Coll. Antropol 2007, 31, 315–319. [Google Scholar]
- Barker, DJP; Eriksson, JG; Forsén, T; Osmond, C. Fetal origins of adult disease: strength of effects and biological basis. Int. J. Epidemiol 2002, 31, 1235–1239. [Google Scholar]
- Gluckman, PD; Hanson, MA. The Fetal Matrix; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Gluckman, PD; Hanson, MA; Beedle, AS. Early life events and their consequences for later disease: a life history and evolutionary perspective. Am. J. Hum. Biol 2007, 19, 1–19. [Google Scholar]
- Leitch, I. Growth and health. Brit. J. Nutr 1951, 5, 142–151. [Google Scholar]
- Crews, DE. Human Senescence: Evolutionary and Biocultural Perspectives; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Larke, A; Crews, DE. Parental investment, late reproduction, and increased reserve capacity are associated with longevity in humans. J. Phy. Anthropol 2006, 25, 119–131. [Google Scholar]
- Bogin, B. Childhood, adolescence, and longevity: a multilevel model of the evolution of reserve capacity in human life history. Am. J. Hum. Biol 2009, 21, 567–577. [Google Scholar]
- Thomson, AM; Duncan, DL. The diagnosis of malnutrition in man. Nutr. Abstr. Rev 1954, 24, 1–18. [Google Scholar]
- Wolanski, N. Parent-offspring similarity in body size and proportions. Stud. Hum. Ecol 1979, 3, 7–26. [Google Scholar]
- Ramos Rodríguez, RM. El significado del miembro superior una hipótesis a considerar. Bol Med Hosp Infant Mex 1981, 38, 373–377. [Google Scholar]
- Ramos Rodríguez, RM. Algunos aspectos de proporcionalidad lineal de una población del estado de Oaxaca. Anales de Antropología 1990, 27, 85–96. [Google Scholar]
- Tanner, JM; Hayashi, T; Preece, MA; Cameron, N. Increase in length of leg relative to trunk in Japanese children and adults from 1957 to 1977: comparison with British and with Japanese Americans. Ann. Hum. Biol 1982, 9, 411–423. [Google Scholar]
- Buschang, PH; Malina, RM; Little, BB. Linear growth in Zapotec schoolchildren: growth status and early velocity for leg length and sitting height. Ann. Hum. Biol 1986, 13, 225–234. [Google Scholar]
- Dickinson, F; Cervera, M; Murguía, R; Uc, L. Growth, nutritional status and environmental change in Yucatan, Mexico. Stud. Hum. Ecol 1990, 9, 135–149. [Google Scholar]
- Gurri, FD; Dickinson, F. Effects of socioeconomic, ecological, and demographic conditions on the development of the extremities and the trunk: a case study with adult females from Chiapas. J. Hum. Ecol 1990, 1, 125–138. [Google Scholar]
- Murguía, R; Dickinson, F; Cervera, M; Uc, L. Socio-economic activities, ecology and somatic differences in Yucatan, Mexico. Stud. Hum. Ecol 1990, 9, 111–134. [Google Scholar]
- Bolzán, AG; Guimarey, LM; Pucciarelli, HM. Crecimiento y dimorfismo sexual de escolares según la ocupación laboral paterna. Archivos Latinoamericanos de Nutrición 1993, 43, 132–38. [Google Scholar]
- Wolanski, N; Dickinson, F; Siniarska, A. Biological traits and living conditions of Maya Indian and non-Maya girls from Merida, Mexico. Int. J. Anthropol 1993, 8, 233–246. [Google Scholar]
- Siniarska, A. Family environment and body build in adults of Yucatan Mexico. Am. J. Phys. Anthropol 1995, 20, 196. [Google Scholar]
- Wolanski, N. Household and family as environment for child growth. Cross cultural studies in Poland, Japan, South Korea and Mexico. In Human Ecology: Progress through Integrative Perspectives; Wright, SD, Meeker, DE, Griffore, R, Eds.; The Society for Human Ecology: Bar Harbor, ME, USA, 1995; pp. 140–152. [Google Scholar]
- Jantz, LM; Jantz, RL. Secular change in long bone length and proportion in the United States, 1800−1970. Am. J. Phys. Anthropol 1999, 110, 57–67. [Google Scholar]
- Whitley, E; Gunnell, G; Davey-Smith, G; Holly, JMP; Martin, RM. Childhood circumstances and anthropometry: The Boyd Orr cohort. Ann. Hum. Biol 2008, 35, 518–534. [Google Scholar]
- Kim, J-M; Stewart, R; Shin, I-S; Kim, SW; Yang, S-J; Yoon, J-S. Associations between head circumference, leg length and dementia in a Korean population. Internat. J. Geriatric Psych 2008, 23, 41–48. [Google Scholar]
- Gunnell, D; Whitley, E; Upton, MN; McConnachie, A; Davey-Smith, G; Watt, GC. Associations of height, leg length, and lung function with cardiovascular risk factors in the Midspan Family Study. J. Epidemiol. Comm. Health 2003, 57, 141–146. [Google Scholar]
- Ferrie, JE; Langenberg, C; Shipley, MJ; Marmot, MG. Birth weight, components of height and coronary heart disease: evidence from the Whitehall II study. Int. J. Epidemiol 2006, 35, 1532–1542. [Google Scholar]
- Wadsworth, ME; Hardy, RJ; Paul, AA; Marshall, SF; Cole, TJ. Leg and trunk length at 43 years in relation to childhood health, diet and family circumstances: evidence from the 1946 national birth cohort. Int. J. Epidemiol 2002, 31, 383–390. [Google Scholar]
- Li, L; Dangour, AL; Power, C. Early life influences on adult leg and trunk length in the 1958 British Birth Cohort. Amer. J. Hum. Biol 2007, 19, 836–843. [Google Scholar]
- Dixon, B; Darlow, B; Prickett, T. How useful is measuring neonatal growth? J. Paediat. Child Health 2008, 44, 444–448. [Google Scholar]
- Frisancho, AR; Guilding, N; Tanner, S. Growth of leg length is reflected in socio-economic differences. Acta Med. Auxol 2001, 33, 47–50. [Google Scholar]
- Malina, RM; Pena Reyes, ME; Tan, SK; Buschang, PH; Little, BB; Koziel, S. Secular change in height, sitting height and leg length in rural Oaxaca, southern Mexico: 1968–2000. Ann. Hum. Biol 2004, 31, 615–633. [Google Scholar]
- Dasgupta, P; Saha, R; Nubé, M. Changes in body size, shape and nutritional status of middle class Bengali boys of Kolkata, India, 1982–2002. Econ. Hum. Biol 2008, 6, 75–94. [Google Scholar]
- Floyd, B. Intergenerational gains in relative knee height as compared to gains in relative leg length within Taiwanese families. Amer. J. Hum. Biol 2008, 20, 462–464. [Google Scholar]
- Gunnell, DJ; Smith, GD; Frankel, SJ; Kemp, M; Peters, TJ. Socio-economic and dietary influences on leg length and trunk length in childhood: A reanalysis of the Carnegie (Boyd Orr) survey of diet and health in pre-war Britain (1937–39). Paediatr. Perinat. Epidemiol 1998, 12, 96–113. [Google Scholar]
- Lawlor, DA; Davey-Smith, G; Ebrahim, S. Association between leg length and offspring birthweight: partial explanation for the trans-generational association between birthweight and cardiovascular disease: findings from the British Women’s Heart and Health Study. Paediatr. Perinat. Epidemiol 2003, 17, 148–155. [Google Scholar]
- Martin, RM; Davey-Smith, G; Frankel, S; Gunnell, D. Parents’ growth in childhood and the birth weight of their offspring. Epidemiol 2004, 15, 308–316. [Google Scholar]
- Leary, S; Davey-Smith, G; Ness, A; ALSPAC Study Team. Smoking during pregnancy and components of stature in offspring 2006. Am. J. Hum. Biol 2006, 18, 502–512. [Google Scholar]
- Dangour, AD. Growth of upper- and lower-body segments in Patamona and Wapishana Amerindian children (cross-sectional data). Ann. Hum. Biol 2001, 28, 649–663. [Google Scholar]
- Bogin, B; Rios, L. Rapid morphological change in living humans: implications for modern human origins. Comp. Biochem. Physiol. A, Mol. Integr. Physiol 2003, 136, 71–84. [Google Scholar]
- Bogin, B; Varela-Silva, MI. Anthropometric variation and health: a biocultural model of human growth. J. Children’s Health 2003, 1, 149–172. [Google Scholar]
- Smith, PK; Bogin, B; Varela-Silva, MI; Orden, AB; Loucky, J. Does immigration help or harm children’s health? The Mayan case. Soc. Sci. Quart 2002, 83, 994–1002. [Google Scholar]
- Smith, PK; Bogin, B; Varela-Silva, MI. Economic and anthropological assessments of the health of children in Maya families in the United States. Econ Hum Biol 2003, 1–2, 145–160. [Google Scholar]
- Han, TS; Hooper, JP; Morrison, CE; Lean, ME. Skeletal proportions and metabolic disorders in adults. Eur. J. Clin. Nutr 1997, 51, 804–809. [Google Scholar]
- Gunnell, DJ; Davey-Smith, G; Frankel, S; Nanchahal, K; Braddon, FE; Pemberton, J; Peters, TJ. Childhood leg length and adult mortality: follow up of the Carnegie (Boyd Orr) Survey of Diet and Health in Pre-war Britain. J. Epidemiol. Comm. Health 1998, 52, 142–152. [Google Scholar]
- Jarvelin, MR. Fetal and infant markers of adult heart diseases. Heart 2000, 84, 219–226. [Google Scholar]
- Smith, GD; Greenwood, R; Gunnell, D; Sweetnam, P; Yarnell, J; Elwood, P. Leg length, insulin resistance, and coronary heart disease risk: the Caerphilly Study. J. Epidemiol. Comm. Health 2001, 55, 867–872. [Google Scholar]
- Langenberg, C; Hardy, R; Kuh, D; Wadsworth, ME. Influence of height, leg and trunk length on pulse pressure, systolic and diastolic blood pressure. J. Hypertens 2003, 21, 537–543. [Google Scholar]
- Lawlor, DA; Taylor, M; Davey-Smith, G; Gunnell, D; Ebrahim, S. Associations of components of adult height with coronary heart disease in postmenopausal women: the British women’s heart and health study. Heart 2004, 90, 745–749. [Google Scholar]
- Fraser, A; Ebrahim, S; Smith, GD; Lawlor, DA. The associations between height components (leg and trunk length) and adult levels of liver enzymes. J. Epidemiol. Community. Health 2008, 62, 48–53. [Google Scholar]
- Gunnell, D; Okasha, M; Smith, GD; Oliver, SE; Sandhu, J; Holly, JM. Height, leg length, and cancer risk: a systematic review. Epidemiol. Rev 2001, 23, 313–342. [Google Scholar]
- Ogilvy-Stuart, AL; Gleeson, H. Cancer risk following growth hormone use in childhood: implications for current practice. Drug Saf 2004, 27, 369–382. [Google Scholar]
- Lima, GA; Corrêa, LL; Gabrich, R; Miranda, LC; Gadelha, MR. IGF-I, insulin and prostate cancer. Arq. Bras. Endocrinol. Metabol 2009, 53, 969–975. [Google Scholar]
- Weng, CJ; Hsieh, YH; Tsai, CM; Chu, YH; Ueng, KC; Liu, YF; Yeh, YH; Su, SC; Chen, YC; Chen, MK; Yang, SF. Relationship of Insulin-Like Growth Factors system gene polymorphisms with the susceptibility and pathological development of hepatocellular carcinoma. Ann Surg Oncol 2010. [Google Scholar]
- Schooling, CM; Jiang, CQ; Heys, M; Zhang, WS; Adab, P; Cheng, KK; Lam, TH; Leung, GM. Are height and leg length universal markers of childhood conditions? The Guangzhou Biobank cohort study. J. Epidemiol. Comm. Health 2008, 62, 607–614. [Google Scholar]
- Schooling, CM; Jiang, CQ; Heys, M; Zhang, WS; Lao, XQ; Adab, P; Cowling, BJ; Thomas, GN; Cheng, KK; Lam, TH; Leung, GM. Is leg length a biomarker of childhood conditions in older Chinese women? The Guangzhou Biobank Cohort Study. J. Epidemiol. Comm. Health 2008, 62, 160–166. [Google Scholar] [Green Version]
- Padez, C; Varela-Silva, MI; Bogin, B. Height and relative leg length as indicators of the quality of the environment among Mozambican jveniles and adolescents. Amer. J. Hum. Biol 2009, 21, 200–209. [Google Scholar]
- Cuenca-Guerra, R; Daza-Flores, JL; Saade-Saade, AJ. Calf implants. Aesthetic Plast. Surg 2009, 33, 505–513. [Google Scholar]
- Fan, J; Liu, F; Wu, J; Dai, W. Visual perception of female physical attractiveness. Proc. Biol. Sci 2004, 271, 347–352. [Google Scholar]
- Weeden, J; Sabini, J. Physical attractiveness and health in Western societies: a review. Psychol. Bull 2005, 131, 635–653. [Google Scholar]
- Gründl, M; Eisenmann-Klein, M; Prantl, L. Quantifying female bodily attractiveness by a statistical analysis of body measurements. Plast. Reconstr. Surg 2009, 123, 1064–1071. [Google Scholar]
- Martins, PA; Hoffman, DJ; Fernandes, MT; Nascimento, CR; Roberts, SB; Sesso, R; Sawaya, AL. Stunted children gain less lean body mass and more fat mass than their non-stunted counterparts: a prospective study. Br. J. Nutr 2004, 92, 819–825. [Google Scholar]
- Velásquez-Meléndez, G; Silveira, EA; Allencastro-Souza, P; Kac, G. Relationship between sitting-height-to-stature ratio and adiposity in Brazilian women. Am. J. Hum. Biol 2005, 17, 646–653. [Google Scholar]
- Hoffman, DJ; Sawaya, AL; Verreschi, I; Tucker, KL; Roberts, SB. Why are nutritionally stunted children at increased risk of obesity? Studies of metabolic rate and fat oxidation in shantytown children from São Paulo, Brazil. Amer. J. Clin. Nutr 2000, 72, 702–707. [Google Scholar]
- Sawaya, AL; Martins, PA; Baccin Martins, VJ; Florêncio, TT; Hoffman, D; Franco, MdCP; das Neves, J. Malnutrition, long-term health and the effect of nutritional recovery. In Emerging Societies—Coexistence of Childhood Malnutrition and Obesity; Kalhan, SC, Prentice, AM, Yajnik, CS, Eds.; Nestlé Nutrition Institute Workshop Series, Pediatric Program, Nestec Ltd: Vevey; S. Karger AG: Basel, Switzerland, 2009; Volume 63, pp. 95–108. [Google Scholar]
- Harvey, P; Martin, RD; Cluton-Brock, TH. Life histories in comparative perspective. In Primate Societies; Smuts, B, Cheney, DL, Seyfarth, RM, Wrangham, RW, Struhsaker, TT, Eds.; University of Chicago Press: Chicago, IL, USA, 1983; pp. 181–196. [Google Scholar]
- Bénazet, JD; Zeller, R. Vertebrate limb development: moving from classical morphogen gradients to an integrated 4-dimensional patterning system. Cold Spring Harbor Perspect. Biol 2009, 1, a001339. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Intermembral Index | Humerofemoral Index |
|---|---|---|
| Human (male) | 69.7 | 71.4 |
| Human (female) | 68.5 | 69.8 |
| Chimpanzee (male) | 108.0 | 101.1 |
| Chimpanzee (female) | 109.4 | 102 |
| Bonobo (male & female) | 102.2 | 98.0 |
| Neonatal mass (grams) | Adult mass (grams) | |||||
|---|---|---|---|---|---|---|
| Species | Brain | Body | Br/Bo ratio | Brain | Body | Br/Bo ratio |
| Pongo (orangutan) | 170.3 | 1,728.0 | 0.10 | 413.3 | 53,000.0 | 0.008 |
| Pan (chimpanzee) | 128.0 | 1,756.0 | 0.07 | 410.3 | 36,350.0 | 0.011 |
| Gorilla | 227.0 | 2,110.0 | 0.11 | 505.9 | 126,500.0 | 0.004 |
| Homo sapiens | 384.0 | 3,300.0 | 0.12 | 1,250.0 | 44,000.0 | 0.284 |
| Measure of “leg length” | Sample sizes | Sample | Results | Source |
|---|---|---|---|---|
| IH | Total: 2,209 M: 1,062 F: 1,147 | 2–14 years Extracted from The Boyd Orr Survey. Children from 1343 working class families in England and Scotland, measured between 1937 and 1939 | M&F: positive association with length of breastfeeding, decreasing numbers of children in the household and increasing household income. Overall, the individual components of stature mostly associated with childhood environment was leg length (measured as IH) and foot length (not in the scope of this entry). | [100] |
| Total: 916 M: 376 F: 540 | 65+ years inhabitants of Kwangju, South Korea, assessed in 2003. | Shorter limb length is associated with markers of lower early-life socioeconomic status and is associated with dementia later in life, especially in women. | [101] | |
| SLL | Total: 2,338 M: 1,040 F: 1,298 | 30–59 years (United Kingdom) | M&F: inverse association with systolic BP, diastolic BP, total cholesterol and fibrinogen. Direct association with FEV, FVC, BW, and BMI | [102] |
| Total: 10,308 M: 6,895 F: 3,413 | 35–55 years (London) | M&F: Strong inverse association with pulse pressure and systolic BP. Strong positive association with lower total/HDL cholesterol ratio, triglycerides, and 2hr glucose M: Strong inverse association with total cholesterol. F: Strong inverse association with diastolic BP. | [103] | |
| Total: 3,262 | Longitudinal study, births from 3–9 March 1946. 21 assessment occasions between birth and 53 years). MRC National Survey of Health & Development (United Kingdom) | M&F: Positive association with mother’s & father’s height, BW. SLL greater among individuals from non-manual social class and among individuals who were breastfed | [104] | |
| Total: 5,900 | The 1958 British Birth Cohort. Participants assessed at birth and at ages 7, 11, 16, 23, 32, 42, and 45 | Adult SLL associated with parental height, birth weight. Taller prepubertal stature is associated with higher SLL. Maternal smoking during pregnancy resulted in lower adult SLL. Overall, adult SLL is related to a greater extent than trunk length to early life factors and prepubertal height | [105] | |
| KH | Total: 50 M: 27 F: 23 | Infants grouped by gestation time at birth: <28 weeks, 28–31 weeks, 32–36 weeks, >36 weeks. Births occurred in 2004–2005, in the neonatal intensive care, Christchurch, New Zealand. | Changes in KH (using a kneemometer) correlate very well with changes in weight. If gain in weight is achieved, normal linear growth may be assumed. Because of this, kneemometry is not a useful addition to routine measurements of growth in the neonatal unit | [106] |
| SHR | Total: 2,985 M: 1,465 F: 1,520 | 2–17 years Mexican-Americans (NHANES III, USA) | M&F: Individuals with relatively shorter legs in proportion to total stature are poorer than longer “legged” individuals (poverty assessed by Poverty Income Ratio) | [107] |
| Total: 1,472 M: 747 F: 707 | 6–13 years, Oaxaca, Southern Mexico Urban in 1972: Total:409, M:218, F:173 Rural in 1978: Total:363, M:179, F:184 Urban in 2000: Total:339, M:173, F:166 Rural in 2000: Total:361, M:177, F:184 | Positive time trend in leg length from 1972 to 2000 both in rural and urban settings | [108] | |
| Total: 2003 M: 2003 F: 0 | 7–16 years. Two cross-sectional surveys among school aged boys from Kolkata, India. 1982–1983 (n = 816) 1999–2002 (n = 1187) | Positive time trend in relative leg length. Boys measured in 1999–2002 had relatively longer legs in proportion to total stature than their counterparts in 1983–1983. | [109] | |
| Total: 1995 M:977 F: 1018 | 5–12 years. Maya migrants to the USA in 1992 (n = 211), Maya migrants to the USA in 2000 (n = 431) and Maya in Guatemala in 1998 (n = 1353) | Leg length is a sensitive indicator of the quality of the environment. Maya children in the USA show relatively longer legs in proportion to stature than their counterparts in Guatemala. By 2000, Maya migrants to the USA were 11.54 cm taller and 6.83 cm longer-legged than Maya children in Guatemala. | [56] | |
| RSLL | Total: 273 | Intergenerational sample Parents’ generation: Total:165, M:80, F:85 Offspring generation: Total:108, M:49, F:59 From Auckland and Taipei | Is an effective marker of intergenerational changes | [110] |
| KHR | Total: 273 | Intergenerational sample Parents’ generation: Total:165, M:80, F:85 Offspring generation: Total:108, M:49, F:59 From Auckland and Taipei | Is an effective marker of intergenerational changes. Lower leg growth, as represented by KHR is similar to changes in overall leg length in sensitivity to environmental change. | [110] |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bogin, B.; Varela-Silva, M.I. Leg Length, Body Proportion, and Health: A Review with a Note on Beauty. Int. J. Environ. Res. Public Health 2010, 7, 1047-1075. https://doi.org/10.3390/ijerph7031047
Bogin B, Varela-Silva MI. Leg Length, Body Proportion, and Health: A Review with a Note on Beauty. International Journal of Environmental Research and Public Health. 2010; 7(3):1047-1075. https://doi.org/10.3390/ijerph7031047
Chicago/Turabian StyleBogin, Barry, and Maria Inês Varela-Silva. 2010. "Leg Length, Body Proportion, and Health: A Review with a Note on Beauty" International Journal of Environmental Research and Public Health 7, no. 3: 1047-1075. https://doi.org/10.3390/ijerph7031047