Thermostable Bacterial Bioflocculant Produced by Cobetia Spp. Isolated from Algoa Bay (South Africa)


 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. The Test Bacteria
2.2. Identification of the Bioflocculant-Producing Bacteria
2.2.1. DNA Extraction
2.2.2. Polymerase Chain Reaction (PCR) Amplification of 16S rRNA Gene
2.3. Media and Cultivation Conditions
2.4. Measurement of Flocculating Activity
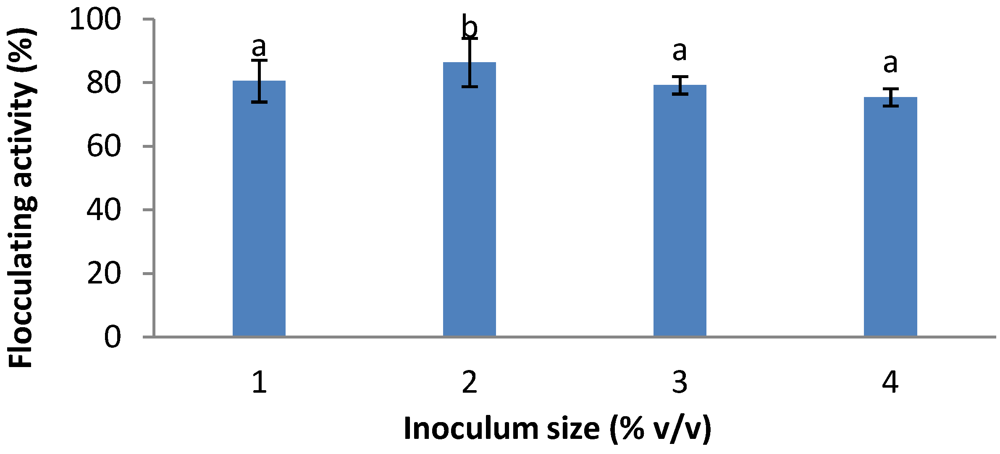
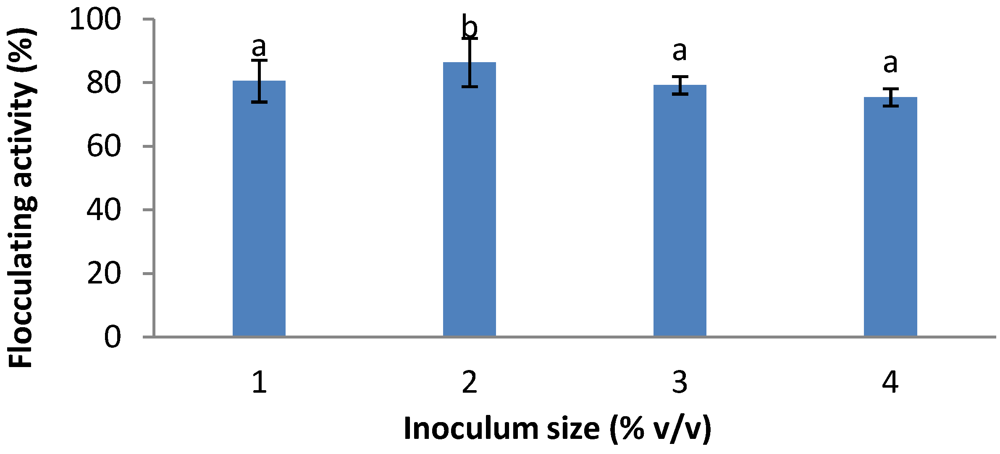
2.5. Effects of Inoculum Size
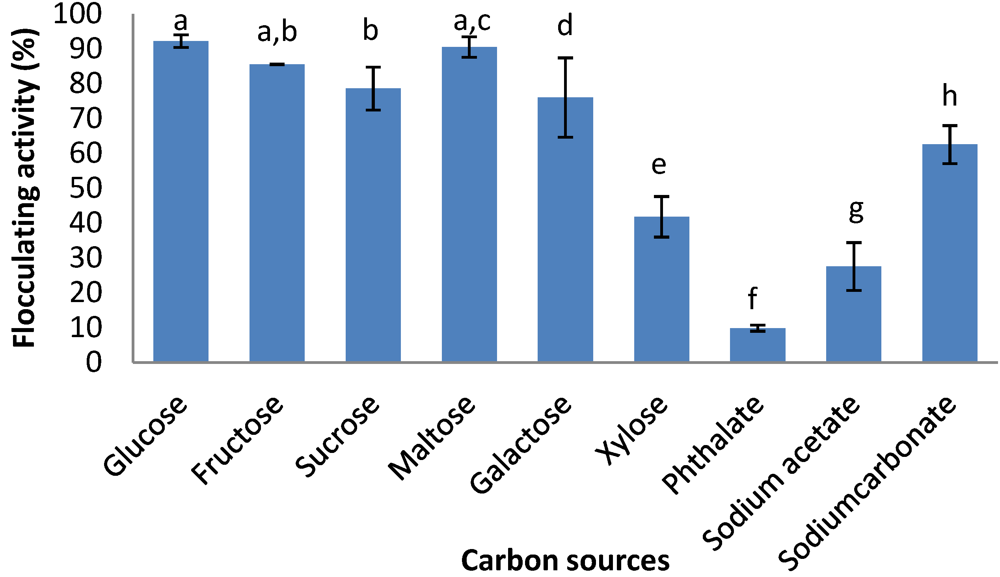
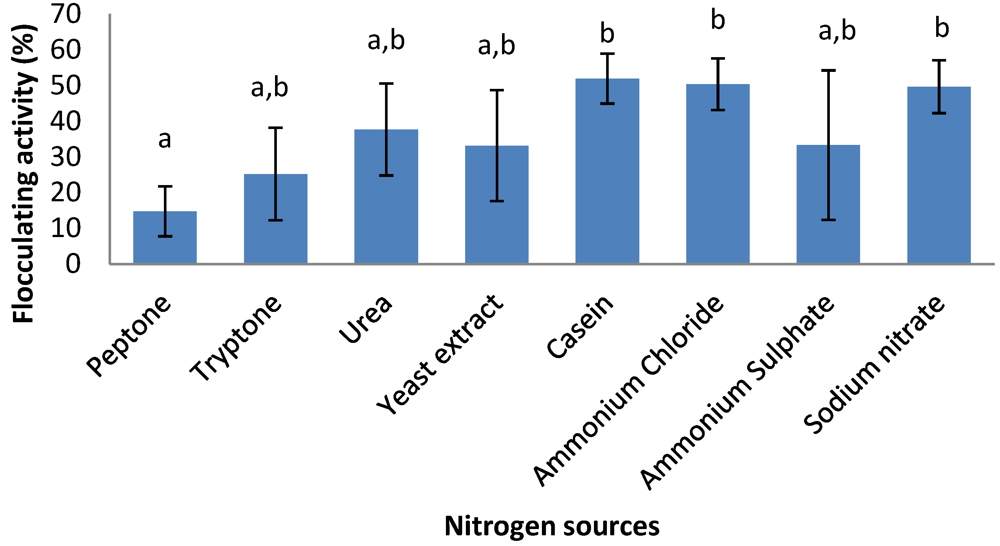
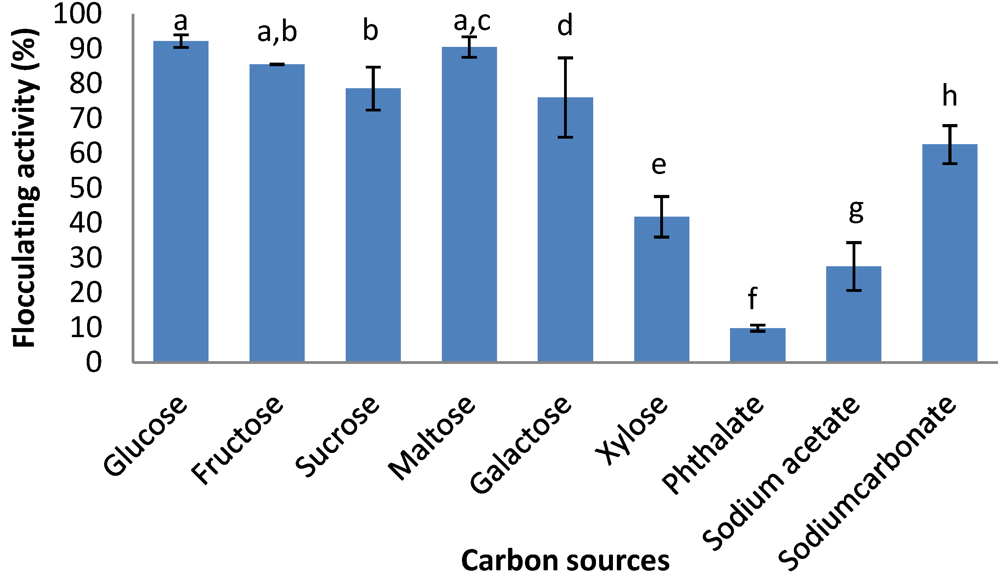
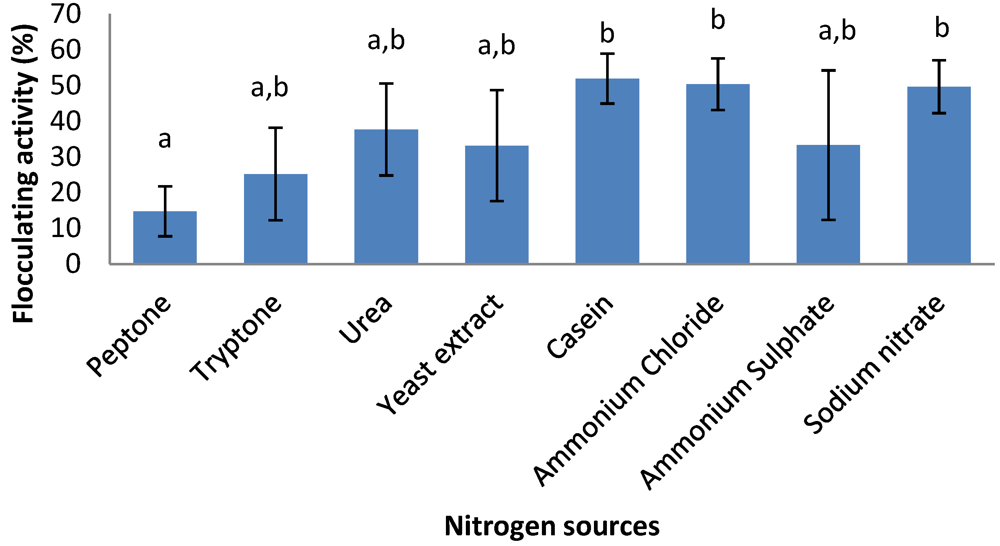
2.6. Effects of Carbon and Nitrogen Sources
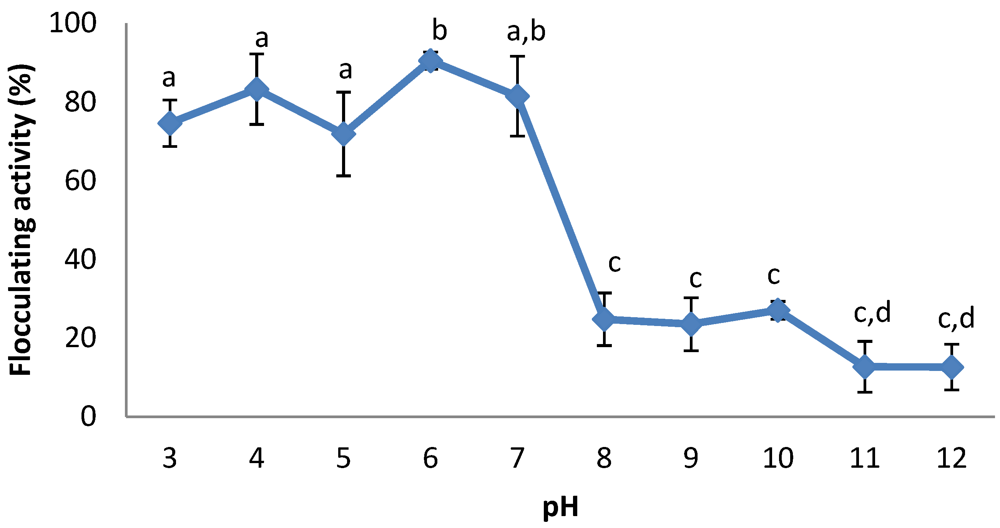
2.7. Effects of Initial pH and Cations
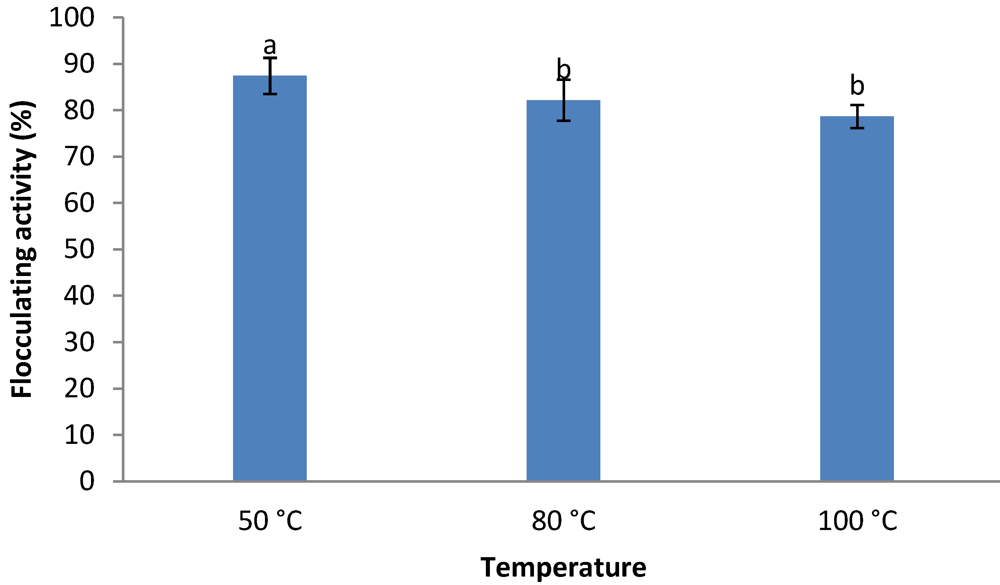
2.8. Thermal Stability of the Crude Bioflocculant
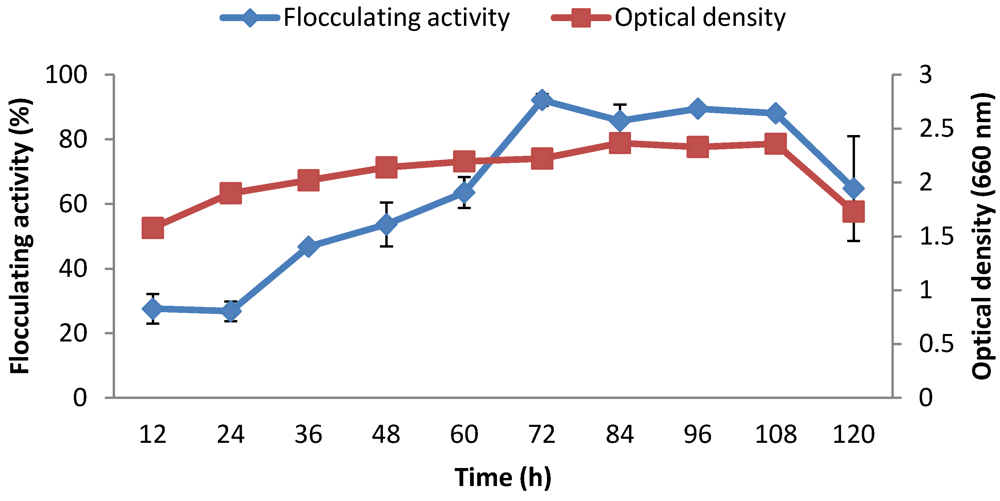
2.9. Time Course Assay
2.10. Extraction and Purification of Bioflocculant
2.11. Analysis of Purified Bioflocculant
2.12. Statistical Analysis
3. Results and Discussion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| METAL IONS | FLOCCULATING ACTIVITY (%) |
|---|---|
| Na+ | 93.61 ± 1.27 a |
| K+ | 94.36 ± 0.51 a |
| Li+ | 91.35 ± 1.69 a |
| Ca2+ | 92.20 ± 1.83 a |
| Mn2+ | 95.02 ± 1.17 a |
| Mg2+ | 90.90 + 0.78 a |
| Al3+ | 77.41 ± 1.94 a |
| Fe3+ | 0.00 b |


| Component | % (w/w) |
|---|---|
| Protein | 5 |
| Neutral sugar | 1.8 |
| Uronic acid | 93 |
4. Conclusions
Acknowledgements
Conflict of Interest
References
- Hubbard, A.T. Encyclopedia of Surface and Colloid Science; CRC Press: Boca Raton, FL, USA, 2004; p. 4230. [Google Scholar]
- Mabinya, L.V.; Cosa, S.; Mkwetshana, N.; Okoh, A.I. Halomonas sp. OKOH—A marine bacterium isolated from the bottom sediment of Algoa Bay—Produces a polysaccharide bioflocculant: Partial characterization and biochemical analysis of its properties. Molecules 2011, 16, 4358–4370. [Google Scholar]
- Nakata, K.; Kurane, R. Production of an extracellular polysaccharide bioflocculant by Klebsiella pneumonia. Biosci. Biotechnol. Biochem. 1999, 63, 2064–2068. [Google Scholar] [CrossRef]
- Salehizadeh, H.; Shojaosadati, S.A. Extracellular biopolymeric flocculants: Recent trends and biotechnology importance. Biotechnol. Adv. 2001, 19, 371–385. [Google Scholar] [CrossRef]
- Master, C.L.; Multhaup, G.; Simms, G. Neuronal origin of a cerebral amyloid: Neurofibrallary tangles of Alzheimer’s disease contain the same protein as the amyloid of plaque cores and blood vessels. EMBO J. 1985, 4, 2757–2763. [Google Scholar]
- Kowall, N.W.; Pendleury, W.W.; Kessler, J.B. Aluminium-induced neurofibrillary degeneration affects a subset of neurons in rabbit cerebral cortex, basal forebrain and upper brain stem. Neuroscience 1989, 29, 329–337. [Google Scholar] [CrossRef]
- Dearfield, K.L.; Abermathy, C.O. Acrylamide: Its metabolism developmental and reproductive effects, genotoxicity and carcinogenicity. Mutat. Res. 1988, 195, 45–77. [Google Scholar]
- Li, Z.; Zhong, S.; Lei, H.; Chen, R.; Yu, Q.; Li, H.L. Production of a novel bioflocculant by Bacillus licheniformis X14 and its application to low temperature drinking water treatment. Bioresour. Technol. 2009, 100, 3650–3656. [Google Scholar] [CrossRef]
- Liu, W.; Wang, K.; Li, B.; Yuan, H.; Yang, J. Production and characterization of an intracellular biofloculant by Chryseobacterium daeguense W6 cultured in low nutrition medium. Bioresour. Technol. 2010, 101, 1044–1048. [Google Scholar] [CrossRef]
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Fenical, W.; Baden, D.; Burg, M.; de Goyet, C.V.; Grimes, J.D.; Katz, M.; Marcus, N.H.; Pomponi, S.; Rhines, P.; Tester, P.; Vena, J. Marine-derived Pharmaceuticals and Related Bioactive Compounds. In From Monsoons to Microbes: Understanding the Ocean’s Role in Human Health; Fenical, W., Ed.; National Academics Press: Washington, DC, USA, 1999; pp. 71–86. [Google Scholar]
- Cook, A.E.; Meyers, P.R. Rapid identification of filamentous actinomycetes to the genus level using genus-specific 16S rRNA gene restriction fragment patterns. Int. J. Syst. Evol. Microbiol. 2003, 53, 1907–1915. [Google Scholar] [CrossRef]
- Zhang, Z.; Lin, B.; Xia, S.; Wang, X.; Yang, A. Production and application of a novel bioflocculant by multiple-microorganism consortia using brewery wastewater as carbon source. J. Environ. Sci. 2007, 19, 667–673. [Google Scholar] [CrossRef]
- Cosa, S.; Mabinya, L.V.; Olaniran, A.O.; Okoh, O.O.; Bernard, K.; Deyzel, S.; Okoh, A.I. Bioflocculant production by Virgibacillus sp. rob isolated from the bottom sediment of Algoa Bay in the Eastern Cape, South Africa. Molecules 2011, 16, 2431–2442. [Google Scholar] [CrossRef]
- Piyo, N.; Cosa, S.; Mabinya, V.L.; Okoh, I.A. Assessment of bioflocculant production by Bacillus sp. gilbert, a marine bacterium isolated from the bottom sediment of Algoa Bay. Mar. Drugs 2011, 9, 1232–1242. [Google Scholar] [CrossRef]
- Kurane, R.; Takeda, K.; Suzuki, T. Screening for characteristics of microbial flocculants. Agric. Biol. Chem. 1986, 50, 2301–2307. [Google Scholar] [CrossRef]
- Wang, S.; Gong, W.; Liu, X.; Lin, T.; Yue, Y.; Gao, B. Production of a novel bioflocculant by culture of Klebsiella mobilis using dairy wastewater. Biochem. Eng. J. 2007, 36, 81–86. [Google Scholar] [CrossRef]
- Gong, W.; Wang, S.; Sun, X.; Liu, X.; Yue, Q.; Gao, B. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour. Technol. 2008, 99, 4668–4674. [Google Scholar] [CrossRef]
- Gao, J.; Bao, H.; Xin, M.; Liu, Y.; Li, Q.; Zhang, Y. Characterization of a bioflocculant from a newly isolated Vagococcus sp. W31. J. Zhejiang Univ. Sci. B 2006, 7, 186–192. [Google Scholar] [CrossRef]
- Chang, W.C.; Soon, A.Y.; In, H.O.; Sang, H.P. Characterization of an extracellular flocculating substance produced by a planktonic cyanobacterium, Anabaena sp. Biotechnol. Lett. 1998, 20, 643–646. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, J.F.; Jiang, P.J.; Yang, S.L.; Liu, Z.L. Composition and characterization of microbiological flocculant SC06. Environ. Chem. 2002, 21, 360–364. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with Folin Phenol reagent. J. Biol. Chem. 1951, 196, 256–275. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Jayaraman, J. Laboratory Manual in Biochemistry; Wiley Eastern: Delhi, India, 1981; pp. 75–76. [Google Scholar]
- Zhong, J.J.; Yoshida, T. High-density cultivation of Perilla frutescens cell suspensions for anthocyanin production: Effects of sucrose concentration and inoculum size. Enzym. Microb. Technol. 1995, 17, 1073–1079. [Google Scholar] [CrossRef]
- Kallos, M.S.; Behie, L.A. Inoculation and growth conditions for high-cell-density expansion of mammalian neural stem cells in suspension bioreactors. Biotechnol. Bioeng. 1999, 63, 473–483. [Google Scholar] [CrossRef]
- Jang, J.H.; Ike, M.; Kim, S.M.; Fujita, M. Production of a novel bioflocculant by fed-batch culture of Citrobacter sp. Biotechnol. Lett. 2001, 23, 593–597. [Google Scholar] [CrossRef]
- Lachhwani, P. Studies on Polymeric Bioflocculant Producing Microorganisms. M.S. Thesis, Thapar Institute of Engineering and Technology, Patiala, India, 2005. [Google Scholar]
- Xia, S.; Zhang, Z.; Wang, X.; Yang, A.; Chen, L.; Zhao, J.; Leonard, D.; Jaffrezic-Renault, N. Production and characterization of bioflocculant by Proteus mirabilis TJ-1. Bioresour. Technol. 2008, 99, 6520–6527. [Google Scholar]
- Liu, L.; Cheng, W. Characteristics and culture conditions of a bioflocculant produced by Penicillium sp. Biomed. Environ. Sci. 2010, 23, 213–218. [Google Scholar] [CrossRef]
- Yokoi, H.; Natsuda, O.; Hirose, J.; Hayashi, S.; Takasaki, Y. Characteristics of a biopolymer flocculant produced by Bacillus sp. PY-90. J. Ferment. Bioeng. 1995, 79, 378–380. [Google Scholar] [CrossRef]
- Shimofuruya, H.; Koide, A.; Shirota, K.; Tsuji, T.; Nakamura, M.; Suzuki, J. The production of flocculating substance(s) by Streptomyces griseus. Biosci. Biotechnol. Biochem. 1995, 60, 498–500. [Google Scholar]
- Takadi, H.; Kodowaki, K. Flocculant production by Paecilomyces sp. Taxanomic studies and culture conditions for production. Agric. Biol. Chem. 1985, 49, 3151–3157. [Google Scholar] [CrossRef]
- Feng, D.L.; Xu, S.H. Characterization of bioflocculant MBF3-3 produced by an isolated Bacillus sp. World J. Microbiol. Biotechnol. 2008, 24, 1627–1632. [Google Scholar] [CrossRef]
- Kwon, G.S.; Moon, S.H.; Hong, S.D.; Lee, H.M.; Kim, H.S.; Oh, H.M.; Yoon, B.D. A novel flocculant biopolymer produced by Pestalotiopsis sp. KCTC 8637p. Biotechnol. Lett. 1996, 18, 1459–1464. [Google Scholar] [CrossRef]
- Wang, L.; Ma, F.; Qu, Y.; Sun, D.; Li, A.; Guo, J.; Yu, B. Characterization of a compound bioflocculant produced by mixed culture of Rhizobium radiobacter F2 and Bacillus sphaeicus F6. World J. Microbiol. Biotechnol. 2011, 27, 2559–2565. [Google Scholar] [CrossRef]
- He, N.; Li, Y.; Chen, J. Production of a novel polygalacturonic acid bioflocculant REA-11 by Corynebacterium glutamicum. Bioresour. Technol. 2004, 94, 99–105. [Google Scholar] [CrossRef]
- Salehizadeh, H.; Vossoughi, M.; Alemzadeh, I. Some investigations on bioflocculant producing bacteria. Biochem. Eng. J. 2000, 5, 39–44. [Google Scholar] [CrossRef]
- Lu, W.Y.; Zhang, T.; Zhang, D.Y.; Li, C.H.; Wen, J.P.; Du, L.X. A novel bioflocculant produced by Enterobacter aerogenes and its use in defecating the trona suspension. Biochem. Eng. J. 2005, 27, 1–7. [Google Scholar] [CrossRef]
- Tago, Y.; Aida, K.O. Extracellular mucopolysaccharide closely related to bacterial floc formation. Appl. Environ. Microbiol. 1977, 34, 308–314. [Google Scholar]
- Salehizadeh, H.; Shojaosadati, S.A. Isolation and characterisation of a bioflocculant produced by Bacillus firmus. Biotechnol. Lett. 2002, 24, 35–40. [Google Scholar] [CrossRef]
- Li, Q.; Liu, H.L.; Qi, Q.S.; Wang, F.S.; Zhang, Y.Z. Isolation and characterization of temperature and alkaline stable bioflocculant from Agrobacterium sp. M-503. New Biotechnol. 2010, 27, 789–794. [Google Scholar] [CrossRef]
- Fattom, A.; Shilo, M. Phormidium J-1 bioflocculant: Production and activity. Arch. Microbiol. 1984, 139, 421–426. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ugbenyen, A.; Cosa, S.; Mabinya, L.; Babalola, O.O.; Aghdasi, F.; Okoh, A. Thermostable Bacterial Bioflocculant Produced by Cobetia Spp. Isolated from Algoa Bay (South Africa). Int. J. Environ. Res. Public Health 2012, 9, 2108-2120. https://doi.org/10.3390/ijerph9062108
Ugbenyen A, Cosa S, Mabinya L, Babalola OO, Aghdasi F, Okoh A. Thermostable Bacterial Bioflocculant Produced by Cobetia Spp. Isolated from Algoa Bay (South Africa). International Journal of Environmental Research and Public Health. 2012; 9(6):2108-2120. https://doi.org/10.3390/ijerph9062108
Chicago/Turabian StyleUgbenyen, Anthony, Sekelwa Cosa, Leonard Mabinya, Olubukola O. Babalola, Farhad Aghdasi, and Anthony Okoh. 2012. "Thermostable Bacterial Bioflocculant Produced by Cobetia Spp. Isolated from Algoa Bay (South Africa)" International Journal of Environmental Research and Public Health 9, no. 6: 2108-2120. https://doi.org/10.3390/ijerph9062108