First Report of a Toxic Nodularia spumigena (Nostocales/ Cyanobacteria) Bloom in Sub-Tropical Australia. I. Phycological and Public Health Investigations

Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of N. spumigena

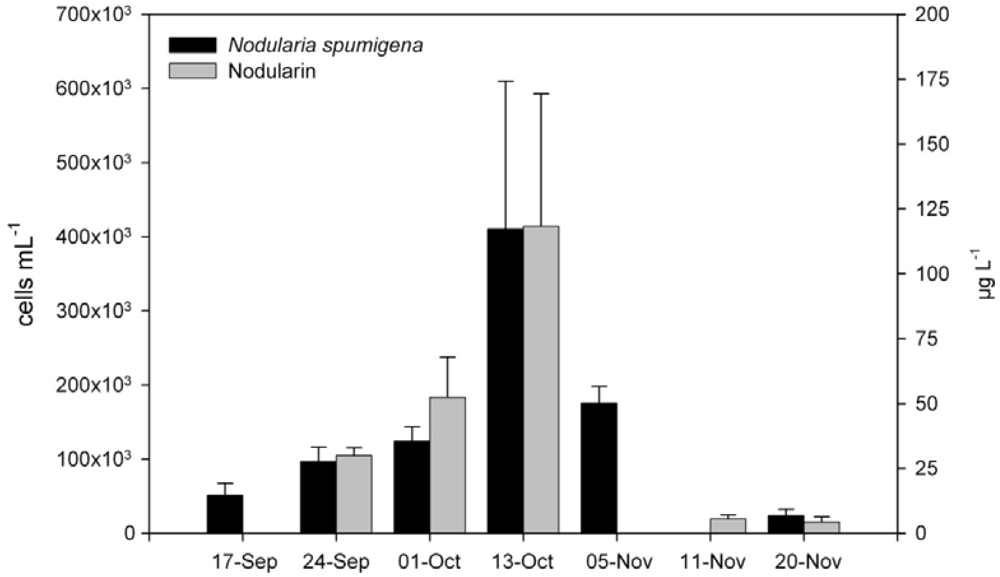
2.2. Phytoplankton

2.3. Toxin in Water
2.4. Toxin in Isolated Strains of N. spumigena
2.5. Molecular Confirmation of Toxin Origin and Identification of N. spumigena
2.6. Water Quality

{kind=link}
{kind=link}
| Groundwater bore | Lake water | ||||
|---|---|---|---|---|---|
| Parameter | 17 September | 17 September | 24 September | 01 October | 14 October |
| Conductivity | 28.5 | nd * | 25 | nd * | nd * |
| (mS cm−1) | |||||
| Total nitrogen | nd * | 3,800 | 2,500 | 1,833 | nd * |
| (µg L−1 as N) | |||||
| Total dissolved nitrogen | nd * | 3,200 | 2,600 | 1,833 | nd * |
| (µg L−1 as N) | |||||
| Ammonia nitrogen | 4,000 | 1,400 | 653 | 10 | nd * |
| (µg L−1 as N) | |||||
| Nitrogen oxides | 3 | 20 | 68 | 7 | nd * |
| (µg L−1 as N) | |||||
| Total phosphorus | nd * | 180 | 51 | 34 | 830 |
| (µg L−1 as P) | |||||
| Total dissolved phosphorus (µg L−1 as P) | nd * | 40 | 50 | 33 | nd * |
| Filterable reactive phosphorus (µg L−1 as P) | 68 | 14 | 10 | 3 | 2 |
2.7. Public Health Response to the Toxic Nodularia Bloom
3. Experimental Section
3.1. Study Area
3.2. Phytoplankton Analysis
3.3. Cyanotoxin Analysis
3.4. Isolation of N. spumigena from Bloom Samples
3.5. Molecular Analysis
3.5.1. DNA Extraction
3.5.2. Amplification of the ndaF Gene Associated with Nodularin Biosynthesis
3.5.3. Amplification of the 16S rRNA Gene
3.5.4. Sequencing of ndaF and 16S rDNA Amplicons
3.6. Chemical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Blackburn, S.I.; McCausland, M.A.; Bolch, C.J.S.; Newman, S.J.; Jones, G.J. Effect of salinity on growth and toxin production in cultures of the bloom-forming cyanobacterium Nodularia spumigena from Australian waters. Phycologia 1996, 35, 511–522. [Google Scholar] [CrossRef]
- Huber, A.L. Factors affecting the germination of akinetes of Nodularia spumigena (Cyanobacteriaceae). Appl. Environ. Microbiol. 1985, 49, 73–78. [Google Scholar]
- John, J.; Kemp, A. Cyanobacterial blooms in the wetlands of the Perth region, taxonomy and distribution: An overview. J. R. Soc. West Aust. 2006, 89, 51–56. [Google Scholar]
- Baker, P.D.; Humpage, A.R. Toxicity associated with commonly occurring cyanobacteria in surface waters of the Murray-Darling basin, Australia. Aust. J. Mar. Freshw. Res. 1994, 45, 773–786. [Google Scholar] [CrossRef]
- Francis, G. Poisonous Australian lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef]
- Camargo, J.A.; Alonso, A. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment. Environ. Int. 2006, 32, 831–849. [Google Scholar] [CrossRef]
- Eriksson, J.E.; Meriluoto, J.A.O.; Kujari, H.P.; Osterlund, K.; Fagerlund, K.; Hallbom, L. Preliminary characterization of a toxin isolated from the cyanobacterium Nodularia spumigena. Toxicon 1988, 26, 161–166. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Matsushima, R.; Watanabe, M.F.; Harada, K.; Ichihara, A.; Carmichael, W.W.; Fujiki, H. Inhibition of protein phosphatases by microcystins and nodularin associated with hepatotoxicity. J. Cancer Res. Clin. Oncol. 1990, 116, 609–614. [Google Scholar] [CrossRef]
- Honkanan, R.E.; Dukelow, M.; Zwiller, J.; Moore, R.E.; Khatra, B.S.; Boynton, A.L. Cyanobacterial nodularin is a potent inhibitor of type 1 and type 2A protein phosphatases. Mol. Pharmacol. 1991, 40, 577–583. [Google Scholar]
- Ohta, T.; Sueoka, E.; Iida, N.; Komori, A.; Suganuma, M.; Nishiwaki, R.; Tatematsu, M.; Kim, S.; Carmichael, W.W.; Fujiki, H. Nodularin, a potent inhibitor of protein phosphatases 1 and 2A, is a new environmental carcinogen in male F344 rat liver. Cancer Res. 1994, 154, 6402–6406. [Google Scholar]
- Nehring, S. Mortality of dogs associated with a mass development of Nodularia spumigena (Cyanophyceae) in a brackish lake at the German North Sea coast. J. Plankton Res. 1993, 15, 867–872. [Google Scholar] [CrossRef]
- Harding, W.R.; Rowe, N.; Wessels, J.C.; Beattie, K.A.; Codd, G.A. Death of a dog attributed to the cyanobacterial (blue-green algal) hepatotoxin nodularin in South Africa. J. S Afr. Vet. Assoc. 1995, 66, 256–259. [Google Scholar]
- Van Halderen, A.; Harding, W.R.; Wessels, J.C.; Schneider, D.J.; Heine, E.W.; van der Merwe, J.; Fourie, J.M. Cyanobacterial (blue-green algae) poisoning of livestock in the western Cape Province of South Africa. J. S. Afr. Vet. Assoc. 1995, 66, 260–264. [Google Scholar]
- Main, D.C.; Berry, P.H.; Peet, R.L.; Robertson, J.P. Sheep mortalities associated with the blue green alga: Nodularia spumigena. Aust. Vet. J. 2008, 53, 578–581. [Google Scholar]
- Elmgren, R.; Larsson, U. Nitrogen and the Baltic sea: Managing nitrogen in relation to phosphorus. Sci. World J. 2001, 1(Suppl 2), 371–377. [Google Scholar] [CrossRef]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar]
- Dodds, W.K.; Bouska, W.W.; Eitzmann, J.L.; Pilger, T.J.; Pitts, K.L.; Riley, A.J. Eutrophication of U.S. freshwaters: Analysis of potential economic damages. Environ. Sci. Technol. 2009, 43, 12–19. [Google Scholar]
- Petersen, S.A. Lake restoration by sediment removal. J. Am. Water Resour. Assoc. 1982, 18, 423–436. [Google Scholar] [CrossRef]
- Weidner, C.; Rücker, J.; Brüggemann, R.; Nixdorf, B. Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions. Oecologia 2007, 152, 473–484. [Google Scholar] [CrossRef]
- Vilhena, L.C.; Hillmer, I.; Imberger, J. The role of climate change in the occurrence of algal blooms: Lake Burragorang, Australia. Limnol. Oceanogr. 2010, 55, 1188–1200. [Google Scholar] [CrossRef]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar]
- Wagner, C.; Adrian, R. Cyanobacteria dominance: Quantifying the effects of climate change. Limnol. Oceanogr. 2009, 54, 2460–2468. [Google Scholar] [CrossRef]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar]
- Murphy, R.; Ribbe, J. Variability of southeast Queensland rainfall and its predictors. Int. J. Climatol. 2004, 24, 703–721. [Google Scholar] [CrossRef]
- Kling, H.J. Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria): A brief historic overview and recent discovery in the Assiniboine River (Canada). Fottea 2009, 9, 45–47. [Google Scholar]
- Hamilton, P.B.; Ley, L.M.; Dean, S.; Pick, F.R. The occurrence of the cyanobacterium Cylindrospermopsis raciborskii in Constance Lake: An exotic cyanoprokaryote new to Canada. Phycologia 2005, 44, 17–25. [Google Scholar] [CrossRef]
- Briand, J.F.; Leboulanger, C.; Humbert, J.F.; Bernard, C.; Dufour, P. Cylindrospermopsis raciborskii (Cyanobacteria) invasion at mid-latitudes: Selection, wide physiological tolerance, or global warming? J. Phycol. 2004, 40, 231–238. [Google Scholar] [CrossRef]
- Padisák, J. Cylindrospermopsis raciborskii (Woloszyńska) Seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: Worldwide distribution and review of its ecology. Arch. Hydrobiol. 1997, 107, 563–593. [Google Scholar]
- Komárek, J.; Hübel, M.; Hübel, H.; Smarda, J. The Nodularia studies. 2. Taxonomy. Arch. Hydrobiol. 1993, 96, 1–25. [Google Scholar]
- Bolch, C.J.; Orr, P.T.; Jones, G.J.; Blackburn, S.I. Genetic, morphological, and toxicological variation among globally distributed strains of Nodularia (Cyanobacteria). J. Phycol. 1999, 35, 339–355. [Google Scholar] [CrossRef]
- Moffit, C.; Neilan, B.A. Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Environ. Microbiol. 2004, 70, 6353–6362. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Zeglińska, J.; Pliński, M. The effect of salinity on the growth, toxin production, and morphology of Nodularia spumigena isolated from the Gulf of Gdańsk, southern Baltic Sea. J. Appl. Phycol. 2005, 17, 171–179. [Google Scholar] [CrossRef]
- Heresztyn, T.; Nicholson, B.C. Nodularin concentrations in Lakes Alexandrina and Albert, South Australia, during a bloom of the cyanobacterium (blue-green alga) Nodularia spumigena and degradation of the toxin. Environ. Toxicol. Water Qual. 1997, 12, 273–282. [Google Scholar] [CrossRef]
- Elliott, A. The seasonal sensitivity of cyanobacteria and other phytoplankton to changes in flushing rate and water temperature. Glob. Chang. Biol. 2010, 16, 864–876. [Google Scholar] [CrossRef]
- Carvalho, L.; Miller, C.A.; Scott, E.M.; Codd, G.A.; Davies, P.S.; Tyler, A.N. Cyanobacterial blooms: Statistical models describing risk factors for national-scale lake assessment and lake management. Sci. Total Environ. 2011, 409, 5353–5358. [Google Scholar] [Green Version]
- Reynolds, C.S. Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- National Health and Medical Research Council (NHMRC), Guidelines for Managing Risks in Recreational Water; Australian Government: Canberra, Australia, 2008.
- Stewart, I.; Carmichael, W.W.; Sadler, R.; McGregor, G.B.; Reardon, K.; Eaglesham, G.K.; Wickramasinghe, W.A.; Seawright, A.A.; Shaw, G.R. Occupational and environmental hazard assessments for the isolation, purification and toxicity testing of cyanobacterial toxins. Environ. Health 2009, 8. [Google Scholar] [CrossRef]
- Stewart, I.; Webb, P.M.; Schluter, P.J.; Shaw, G.R. Recreational and occupational field exposure to freshwater cyanobacteria—A review of anecdotal and case reports, epidemiological studies and the challenges for epidemiologic assessment. Environ. Health 2006, 5. [Google Scholar] [CrossRef]
- Giannuzzi, L.; Sedan, D.; Echenique, R.; Andrinolo, D. An acute case of intoxication with cyanobacteria and cyanotoxins in recreational water in Salto Grande Dam, Argentina. Mar. Drugs 2011, 9, 2164–2175. [Google Scholar] [CrossRef]
- Probert, C.S.; Robinson, R.J.; Jayanthi, V.; Mayberry, J.F. Microcystin hepatitis. Arq. Gastroenterol. 1995, 32, 199. [Google Scholar]
- Ibelings, B.W.; Chorus, I. Accumulation of cyanobacterial toxins in freshwater “seafood” and its consequences for public health: A review. Environ. Pollut. 2007, 150, 177–192. [Google Scholar] [CrossRef]
- Sipiä, V.O.; Karlsson, K.M.; Meriluoto, J.A.; Kankaanpää, H.T. Eiders (Somateria mollissima) obtain nodularin, a cyanobacterial hepatotoxin, in Baltic Sea food web. Environ. Toxicol. Chem. 2004, 23, 1256–1260. [Google Scholar] [CrossRef]
- Van Buynder, P.G.; Oughtred, T.; Kirkby, B.; Phillips, S.; Eaglesham, G.; Thomas, K.; Burch, M. Nodularin uptake by seafood during a cyanobacterial bloom. Environ. Toxicol. 2001, 16, 468–471. [Google Scholar] [CrossRef]
- Stark, C.H.; Richards, K.G. The continuing challenge of nitrogen loss to the environment: Environmental consequences and mitigation strategies. Dyn. Soil Dyn. Plant 2008, 2, 41–55. [Google Scholar]
- Steffensen, D.A. Economic cost of cyanobacterial blooms. Adv. Exp. Med. Biol. 2008, 619, 855–865. [Google Scholar] [CrossRef]
- Smith, J.L.; Boyer, G.L.; Zimba, P.V. A review of cyanobacterial odorous and bioactive metabolites: Impacts and management alternatives in aquaculture. Aquaculture 2008, 280, 5–20. [Google Scholar] [CrossRef]
- Tucker, C.S. Off-flavor problems in aquaculture. Rev. Fish. Sci. 2000, 8, 45–88. [Google Scholar] [CrossRef]
- Healy, M.; Cawley, A.M. Nutrient processing capacity of a constructed wetland in western Ireland. J. Environ. Qual. 2002, 31, 1739–1747. [Google Scholar] [CrossRef]
- Vymazal, J. Plants used in constructed wetlands with horizontal subsurface flow: A review. Hydrobiologia 2011, 674, 133–156. [Google Scholar] [CrossRef]
- Jones, G.J.; Orr, P.T. Release and degradation of microcystin following algicide treatment of a Microcystis aeruginosa bloom in a recreational lake, as determined by HPLC and protein phosphatase inhibition assay. Water Res. 1994, 28, 871–876. [Google Scholar] [CrossRef]
- Hötzel, G.; Croome, R. A Phytoplankton Methods Manual for Australian Freshwaters; Occasional Paper 22/29; Land and Water Research and Development Corporation: Canberra, Australia, 1999. [Google Scholar]
- Lund, J.W.G.; Kipling, C.; Le Cren, E.D. The inverted microscope method of estimating algal numbers and the statistical basis for counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.; Kirschtel, D.; Pollinger, U.; Zohary, H. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Lawton, L.A.; Edwards, C.; Codd, G.A. Extraction and high-performance liquid chromatographic method for the determination of microcystins in raw and treated waters. Analyst 1994, 119, 1525–1530. [Google Scholar] [CrossRef]
- Bolch, C.J.; Blackburn, S. Isolation and purification of Australian isolates of the toxic cyanobacterium Microcystis aeruginosa Kütz. J. Appl. Phycol. 1996, 8, 5–13. [Google Scholar] [CrossRef]
- Koskenniemi, K.; Lyra, C.; Rajaniemi-Wacklin, P.; Jokela, J.; Sivonen, K. Quantitative real-time PCR detection of toxic Nodularia cyanobacteria in the Baltic sea. Appl. Environ. Microbiol. 2007, 73, 2173–2179. [Google Scholar]
- Neilan, B.A.; Jacobs, D.; Del Dot, T.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman, A.E. rRNA sequences and evolutionary relationships among toxic and non-toxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef]
- Iteman, I.; Rippka, R.; Tandeau de Marsac, N.; Herdman, M. rDNA analyses of planktonic heterocystous cyanobacteria, including members of the genera Anabaenopsis and Cyanospira. Microbiology 2002, 148, 481–496. [Google Scholar]
- Shaw, G.R.; Sukenik, A.; Livne, A.; Chiswell, R.K.; Smith, M.J.; Seawright, A.A.; Norris, R.L.; Eaglesham, G.K.; Moore, M.R. Blooms of the cylindrospermopsin containing cyanobacterium, Aphanizomenon ovalisporum (Forti), in newly constructed lakes, Queensland, Australia. Environ. Toxicol. 1999, 14, 167–177. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- American Public Health Association (APHA), Standard Methods for the Examination of Water & Wastewater, 21; Eaton, A.D.; Clesceri, L.S.; Rice, E.W.; Greenberg, A.E.; Franson, M.A.H. (Eds.) APHA: Washington, DC, USA, 2005.
- Hosomi, M.; Sudo, R. Simultaneous determination of total nitrogen and total phosphorus in freshwater samples using persulfate digestion. Int. J. Environ. Stud. 1986, 27, 267–275. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
McGregor, G.B.; Stewart, I.; Sendall, B.C.; Sadler, R.; Reardon, K.; Carter, S.; Wruck, D.; Wickramasinghe, W. First Report of a Toxic Nodularia spumigena (Nostocales/ Cyanobacteria) Bloom in Sub-Tropical Australia. I. Phycological and Public Health Investigations. Int. J. Environ. Res. Public Health 2012, 9, 2396-2411. https://doi.org/10.3390/ijerph9072396
McGregor GB, Stewart I, Sendall BC, Sadler R, Reardon K, Carter S, Wruck D, Wickramasinghe W. First Report of a Toxic Nodularia spumigena (Nostocales/ Cyanobacteria) Bloom in Sub-Tropical Australia. I. Phycological and Public Health Investigations. International Journal of Environmental Research and Public Health. 2012; 9(7):2396-2411. https://doi.org/10.3390/ijerph9072396
Chicago/Turabian StyleMcGregor, Glenn B., Ian Stewart, Barbara C. Sendall, Ross Sadler, Karen Reardon, Steven Carter, Dan Wruck, and Wasa Wickramasinghe. 2012. "First Report of a Toxic Nodularia spumigena (Nostocales/ Cyanobacteria) Bloom in Sub-Tropical Australia. I. Phycological and Public Health Investigations" International Journal of Environmental Research and Public Health 9, no. 7: 2396-2411. https://doi.org/10.3390/ijerph9072396