
Changes in Carbon Electrode Morphology Affect Microbial Fuel Cell Performance with Shewanella oneidensis MR-1


Abstract
:1. Introduction
2. Experimental Section
2.1. Electrode Characterization
2.2. Cell Cultures
2.3. Micro-Electrolysis Cell (MEC) Operation
3. Results and Discussion
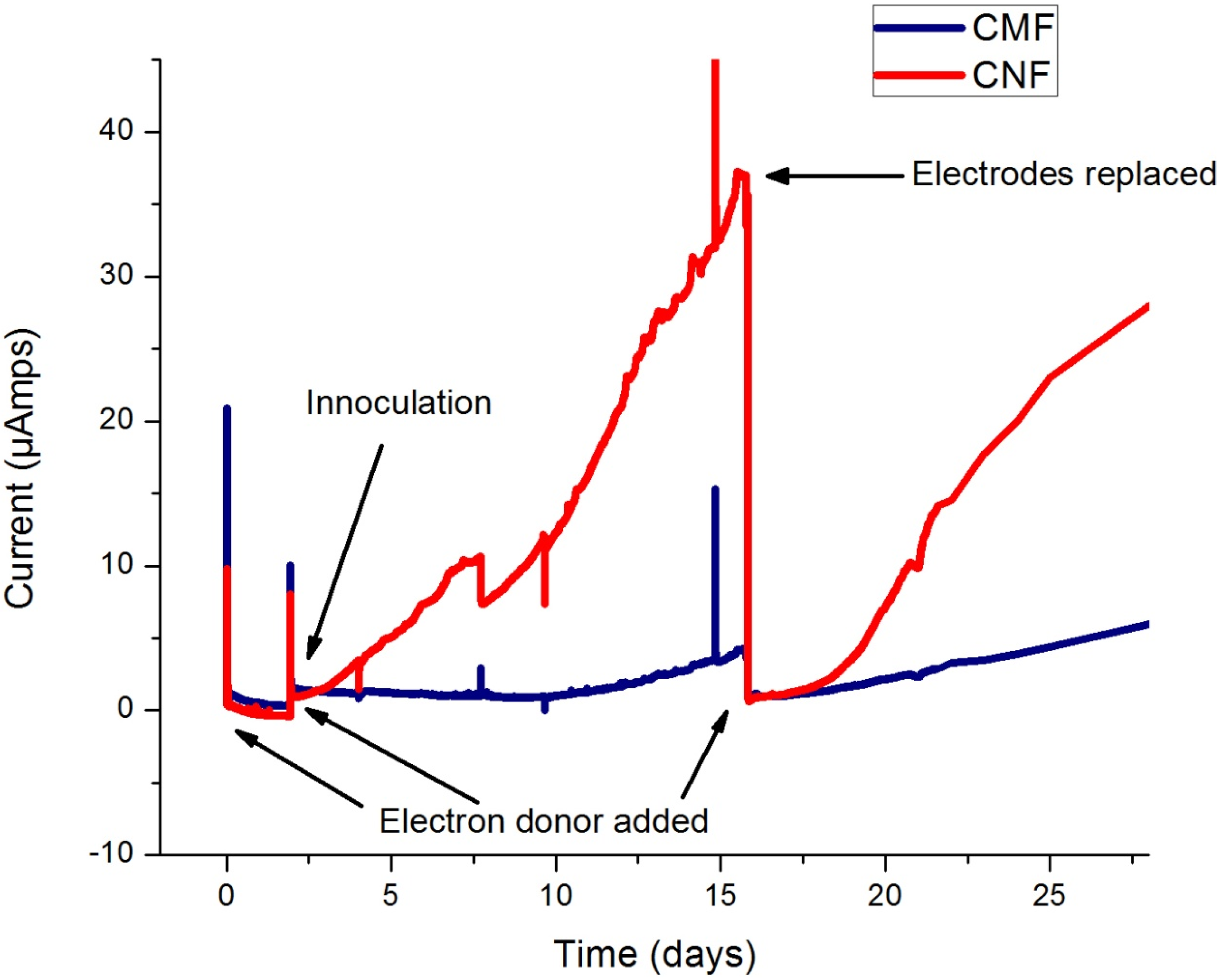
3.1. Current Production

3.2. Cyclic Voltammograms

3.2.1. Biofilm-Based Electron Transfer
3.2.2. Comparison of Electroactive Surface Area and Kinetics Using CVs
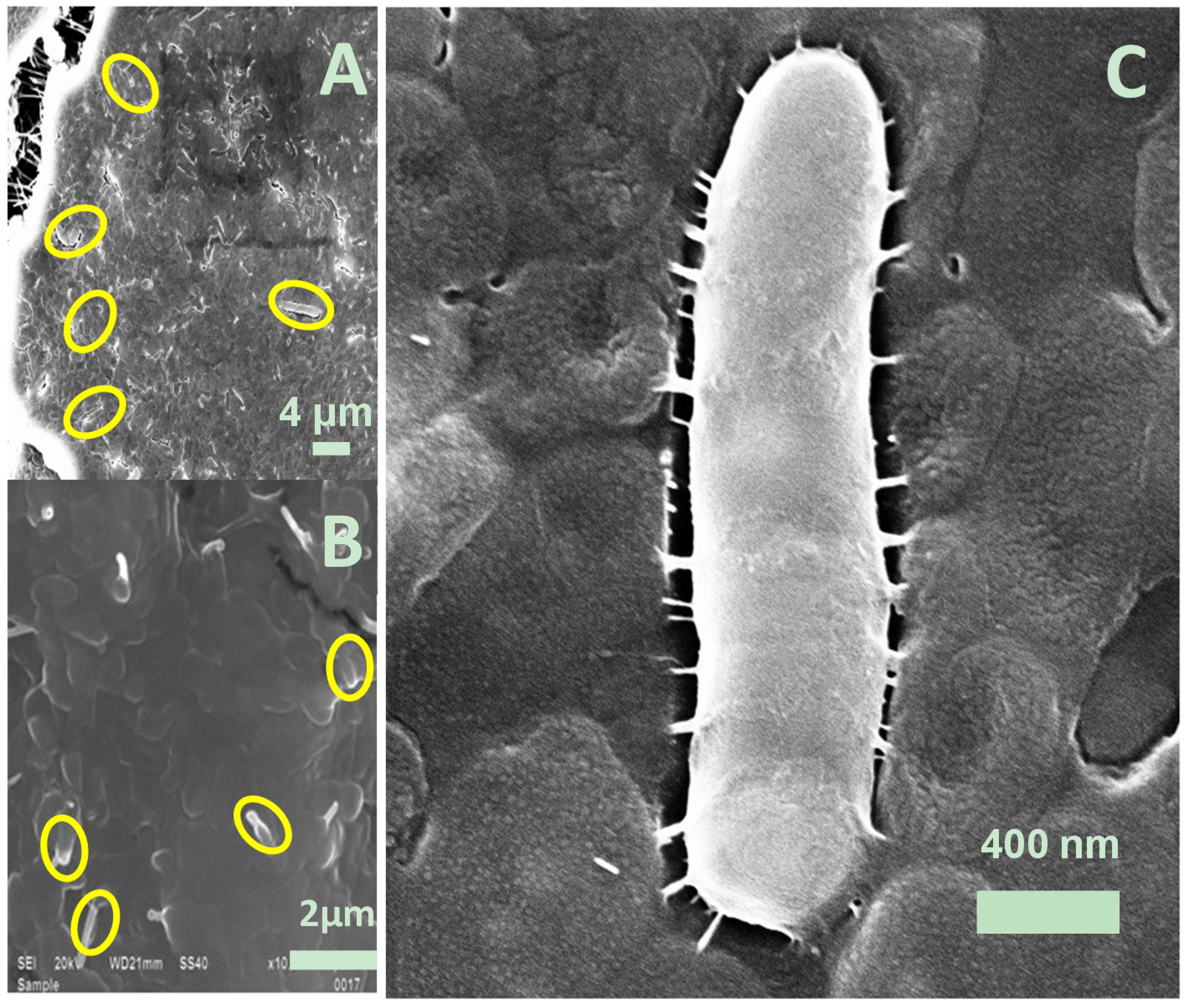
3.3. SEM Images for Biofilm Colonization

Biofilm Formation
3.4. Morphology of Sterile Electrodes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SEM images | Carbon microfiber paper | Carbon nanofiber mats |
|---|---|---|
| Areal Wt (g/m2) | 161 | 40 |
| Thickness (cm) | 0.038 | 0.015 |
| Bulk resistivity (Ohm-cm) | 0.006 | 0.075 |
| Measured sheet resistivity (Ω/sq) | 0.17 | 5 |
| Avg. diameter of constituent material | 10 μm | 0.2 μm |
| Bonding | sp2 | sp2 |
| Surface modifications | None | None |
Impact of Electrode Morphology.
3.5. Differences in Electrode Conductivity
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bond, D.R.; Lovley, D.R. Electricity production by Geobacter sulfurreducens attached to electrodes. Appl. Environ. Microbiol. 2003, 69, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Franks, A.E.; Malvankar, N.; Nevin, K.P. Bacterial biofilms: The powerhouse of a microbial fuel cell. Biofuels 2010, 1, 589–604. [Google Scholar] [CrossRef]
- Torres, C.I.; Marcus, A.K.; Lee, H.-S.; Parameswaran, P.; Krajmalnik-Brown, R.; Rittmann, B.E. A kinetic perspective on extracellular electron transfer by anode-respiring bacteria. FEMS Microbiol. Rev. 2010, 34, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Reguera, G.; Nevin, K.P.; Nicoll, J.S.; Covalla, S.F.; Woodard, T.L.; Lovley, D.R. Biofilm and nanowire production leads to increased current in Geobacter sulfurreducens fuel cells. Appl. Environ. Microbiol. 2006, 72, 7345–7348. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, S.K.; Lovley, D.R. Electricity generation by direct oxidation of glucose in mediatorless microbial fuel cells. Nat. Biotechnol. 2003, 21, 1229–1232. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-Y.; Wu, C.-C.; Lee, C.-Y.; Shih, E.P. Microbial fuel cell performance of multiwall carbon nanotubes on carbon cloth as electrodes. J. Power Sources 2009, 194, 199–205. [Google Scholar] [CrossRef]
- Liu, Y.; Harnisch, F.; Fricke, K.; Schröder, U.; Climent, V.; Feliu, J.M. The study of electrochemically active microbial biofilms on different carbon-based anode materials in microbial fuel cells. Biosens. Bioelectron. 2010, 25, 2167–2171. [Google Scholar] [CrossRef]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial fuel cells: Methodology and technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Aelterman, P.; Verstraete, W. Bioanode performance in bioelectrochemical systems: Recent improvements and prospects. Trends Biotechnol. 2009, 27, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Morozan, A.; Stamatin, I.; Stamatin, L.; Dumitru, A.; Scott, K. Carbon electrodes for microbial fuel cells. J. Optoelectron. Adv. Mater. 2007, 9, 221–224. [Google Scholar]
- Logan, B.; Cheng, S.; Watson, V.; Estadt, G. Graphite fiber brush anodes for increased power production in air-cathode microbial fuel cells. Environ. Sci. Technol. 2007, 41, 3341–3346. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Li, C.M.; Bao, S.-J.; Bao, Q.-L. Carbon nanotube/polyaniline composite as anode material for microbial fuel cells. J. Power Sources 2007, 170, 79–84. [Google Scholar] [CrossRef]
- Rosenbaum, M.; Zhao, F.; Quaas, M.; Wulff, H.; Schröder, U.; Scholz, F. Evaluation of catalytic properties of tungsten carbide for the anode of microbial fuel cells. Appl. Catal. B Environ. 2007, 74, 261–269. [Google Scholar] [CrossRef]
- Sanchez, D.V.P.; Huynh, P.; Kozlov, M.E.; Baughman, R.H.; Vidic, R.D.; Yun, M. Carbon nanotube/platinum (Pt) sheet as an improved cathode for microbial fuel cells. Energy Fuels 2010, 24, 5897–5902. [Google Scholar] [CrossRef]
- Dewan, A.; Beyenal, H.; Lewandowski, Z. Scaling up microbial fuel cells. Environ. Sci. Technol. 2008, 42, 7643–7648. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, S.; Fernández Morales, F.J.; Kleerebezem, R.; Heijnen, J.J.; van Loosdrecht, M.C.M. High-rate acidophilic ferrous iron oxidation in a biofilm airlift reactor and the role of the carrier material. Biotechnol. Bioeng. 2005, 90, 462–472. [Google Scholar] [CrossRef]
- Kappell, G.M.; Grover, J.P.; Chrzanowski, T.H. Micro-scale surface-patterning influences biofilm formation. Electron. J. Biotechnol. 2009, 12, 10–11. [Google Scholar] [CrossRef]
- Chen, S.; Hou, H.; Harnisch, F.; Patil, S.A.; Carmona-Martinez, A.A.; Agarwal, S.; Zhang, Y.; Sinha-Ray, S.; Yarin, A.L.; Greiner, A.; et al. Electrospun and solution blown three-dimensional carbon fiber nonwovens for application as electrodes in microbial fuel cells. Energy Environ. Sci. 2011, 4, 1417–1421. [Google Scholar] [CrossRef]
- He, G.; Gu, Y.; He, S.; Schröder, U.; Chen, S.; Hou, H. Effect of fiber diameter on the behavior of biofilm and anodic performance of fiber electrodes in microbial fuel cells. Bioresour. Technol. 2011, 102, 10763–10766. [Google Scholar] [CrossRef]
- Baron, D.; LaBelle, E.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Electrochemical measurement of electron transfer kinetics by Shewanella oneidensis MR-1. J. Biol. Chem. 2009, 284, 28865–28873. [Google Scholar] [CrossRef]
- Marsili, E.; Baron, D.B.; Shikhare, I.D.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Shewanella secretes flavins that mediate extracellular electron transfer. Proc. Natl. Acad. Sci. USA 2008, 105, 3968–3973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanthier, M.; Gregory, K.B.; Lovley, D.R. Growth with high planktonic biomass in Shewanella oneidensis fuel cells. FEMS Microbiol. Lett. 2008, 278, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Bard, A.J.; Faulkner, L.R. Electrochemical Methods: Fundamentals and Applications, 2nd ed.; John Wiley & Sons Inc.: New York, NY, USA, 2001. [Google Scholar]
- Torres, C.I.; Marcus, A.K.; Parameswaran, P.; Rittmann, B.E. Kinetic experiments for evaluating the nernst-monod model for anode-respiring bacteria (ARB) in a biofilm anode. Environ. Sci. Technol. 2008, 42, 6593–6597. [Google Scholar] [CrossRef] [PubMed]
- Marcus, A.K.; Torres, C.I.; Rittmann, B.E. Conduction-based modeling of the biofilm anode of a microbial fuel cell. Biotechnol. Bioeng. 2007, 98, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Pavithra, D.; Mukesh, D. Biofilm formation, bacterial adhesion and host response on polymeric implants—Issues and prevention. Biomed. Mater. 2008, 3, 034003. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; He, G.; Carmona-Martinez, A.A.; Agarwal, S.; Greiner, A.; Hou, H.; Schröder, U. Electrospun carbon fiber mat with layered architecture for anode in microbial fuel cells. Electrochem. Commun. 2011, 13, 1026–1029. [Google Scholar] [CrossRef]
- Park, H.I.; Mushtaq, U.; Perello, D.; Lee, I.; Cho, S.K.; Star, A.; Yun, M. Effective and low-cost platinum electrodes for microbial fuel cells deposited by electron beam evaporation. Energy Fuels 2007, 21, 2984–2990. [Google Scholar] [CrossRef]
- Bretschger, O.; Obraztsova, A.; Sturm, C.A.; Chang, I.S.; Gorby, Y.A.; Reed, S.B.; Culley, D.E.; Reardon, C.L.; Barua, S.; Romine, M.F.; et al. Current production and metal oxide reduction by Shewanella oneidensis MR-1 wild type and mutants. Appl. Environ. Microbiol. 2007, 73, 7003–7012. [Google Scholar] [CrossRef] [PubMed]
- Thormann, K.M.; Saville, R.M.; Shukla, S.; Pelletier, D.A.; Spormann, A.M. Initial phases of biofilm formation in Shewanella oneidensis MR-1. J. Bacteriol. 2004, 186, 8096–8104. [Google Scholar] [CrossRef] [PubMed]
- Hirst, J. Elucidating the mechanisms of coupled electron transfer and catalytic reactions by protein film voltammetry. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 225–239. [Google Scholar] [CrossRef]
- Epstein, A.K.; Hochbaum, A.I.; Kim, P.; Aizenberg, J. Control of bacterial biofilm growth on surfaces by nanostructural mechanics and geometry. Nanotechnology 2011, 22, 494007:1–494007:8. [Google Scholar] [CrossRef]
- Malvankar, N.S.; Tuominen, M.T.; Lovley, D.R. Biofilm conductivity is a decisive variable for high-current-density Geobacter sulfurreducens microbial fuel cells. Energy Environ. Sci. 2012, 5, 5790–5797. [Google Scholar] [CrossRef]
- McDonough, J.R.; Choi, J.W.; Yang, Y.; la Mantia, F.; Zhang, Y.G.; Cui, Y. Carbon nanofiber supercapacitors with large areal capacitances. Appl. Phys. Lett. 2009, 95, 243109:1–243109:3. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, X.; Ju, H. Detection of NADH and ethanol based on catalytic activity of soluble carbon nanofiber with low overpotential. Anal. Chem. 2006, 79, 453–458. [Google Scholar] [CrossRef]
- Ra, E.J.; Raymundo-Piñero, E.; Lee, Y.H.; Béguin, F. High power supercapacitors using polyacrylonitrile-based carbon nanofiber paper. Carbon 2009, 47, 2984–2992. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchez, D.V.P.; Jacobs, D.; Gregory, K.; Huang, J.; Hu, Y.; Vidic, R.; Yun, M. Changes in Carbon Electrode Morphology Affect Microbial Fuel Cell Performance with Shewanella oneidensis MR-1. Energies 2015, 8, 1817-1829. https://doi.org/10.3390/en8031817
Sanchez DVP, Jacobs D, Gregory K, Huang J, Hu Y, Vidic R, Yun M. Changes in Carbon Electrode Morphology Affect Microbial Fuel Cell Performance with Shewanella oneidensis MR-1. Energies. 2015; 8(3):1817-1829. https://doi.org/10.3390/en8031817
Chicago/Turabian StyleSanchez, David V. P., Daniel Jacobs, Kelvin Gregory, Jiyong Huang, Yushi Hu, Radisav Vidic, and Minhee Yun. 2015. "Changes in Carbon Electrode Morphology Affect Microbial Fuel Cell Performance with Shewanella oneidensis MR-1" Energies 8, no. 3: 1817-1829. https://doi.org/10.3390/en8031817