Consequences of More Intensive Forestry for the Sustainable Management of Forest Soils and Waters

{kind=link}
{kind=link}
Abstract
: Additions of nutrients, faster growing tree varieties, more intense harvest practices, and a changing climate all have the potential to increase forest production in Sweden, thereby mitigating climate change through carbon sequestration and fossil fuel substitution. However, the effects of management strategies for increased biomass production on soil resources and water quality at landscape scales are inadequately understood. Key knowledge gaps also remain regarding the sustainability of shorter rotation periods and more intensive biomass harvests. This includes effects of fertilization on the long-term weathering and supply of base cations and the consequences of changing mineral availability for future forest production. Furthermore, because soils and surface waters are closely connected, management efforts in the terrestrial landscape will potentially have consequences for water quality and the ecology of streams, rivers, and lakes. Here, we review and discuss some of the most pertinent questions related to how increased forest biomass production in Sweden could affect soils and surface waters, and how contemporary forestry goals can be met while minimizing the loss of other ecosystem services. We suggest that the development of management plans to promote the sustainable use of soil resources and water quality, while maximizing biomass production, will require a holistic ecosystem approach that is placed within a broader landscape perspective.1. Introduction
Developing management strategies for the sustainable use of soil and water resources is recognized as a major environmental challenge for future generations [1,2]. Both soil and water resources are likely to become increasingly exploited and threatened by a growing human population, changes in biogeochemical cycles, and major land-use transitions at global scales [3]. Even in sparsely populated boreal regions, soils and waters are being affected by long-range transport and atmospheric deposition of pollutants, climate change, and land use activities, including forest management [4,5]. In order to manage forests in a sustainable way, decision making must be based on a sound understanding of the relationships between environmental change and ecosystem function. To develop this understanding, we need to improve our basic knowledge of the underlying mechanisms behind forest ecosystem dynamics and land-water interactions across the range of spatial and temporal scales relevant to land management.
Greater pressure is being placed on forests as the demand for wood products increases, and forest biomass becomes increasingly popular as an alternative to fossil fuels [6,7]. For example, the use of forest products in district heating in Sweden has dramatically increased from 1 PJ (∼0.15 million m3 tree biomass) in 1980 to 92 PJ (∼13.8 million m3 tree biomass) in 2008 [8]. In an effort to meet the rising demand for forest products, it has been suggested that forestry should be intensified by increasing the use of fertilizers and by using genetically improved seedling stocks to increase biomass production, and thereby decrease rotation periods. Whole-tree harvesting and full-tree harvesting (defined respectively as the removal of all above-stump tree biomass and the removal of all tree biomass including the stump) are two other forestry techniques that could potentially be used to meet the increased demands for forest biomass [9].
Intensified harvesting, in combination with other forest management practices, could have large ecological consequences for both terrestrial and aquatic ecosystems, including effects on the storage and availability of carbon (C), nitrogen (N), calcium (Ca), potassium (K) and other nutrients [10-12]. Because forestry operations typically affect the biological, physical and chemical properties of soils [9,13], it has been suggested that preserving forests in reserves is the best way to protect several ecosystem services, including the sequestration of soil C [14]. However, Berg et al. [15] used 41 years of soil survey data across all forested areas of Sweden and showed an average accumulation in the humus layer of 250 kgC·ha−1·yr−1. The corresponding accumulation in predominately managed forests in the northern part of the country estimated by Berg et al. [15] was actually higher than that estimated to accumulate in soils of the same region of undisturbed, old-growth forests during periods with no human or fire disturbance [16]. Thus, for Swedish forest soils, forestry may lead to an increased C accumulation, presumably by maintaining a lower average stand age, which is associated with greater rates of growth and detrital inputs to soil, when compared to old-growth forests. While conventional forest management may lead to increased carbon sequestration above and below ground, it is critical to understand how more intensive forestry approaches may influence the entire suite of ecosystem services that these landscapes provide.
The potential combined effect of new forest management approaches and climate change on the long-term sustainability of soil and water resources on soil and water resources also needs consideration. Of particular interest is whether and how the environmental impact of intensified biomass production and harvest will be affected by predicted climate changes that may alter both temperature and precipitation in Sweden. Soils and surface waters are closely linked by groundwater and runoff that transport dissolved nutrients and other solutes from terrestrial to aquatic ecosystems. As a consequence, climate-induced changes in plant productivity, soil biogeochemistry, and catchment hydrology will likely also affect the water quality and ecology of streams and lakes [17]. The potential downstream consequences of the combined effects of intensified biomass production and climate change will ultimately depend on how individual landscape elements or small catchments are affected, the distribution and arrangement of these affected subsystems, and how biogeochemical changes are propagated in time and space among hydrologically linked areas.
One critical management goal is to develop and implement strategies that increase tree biomass yield while maintaining the long-term sustainability of forested landscapes, which is defined here as the ability of the soil ecosystem to support desired rates of biomass production over several rotation periods without significantly affecting water chemistry and/or aquatic biodiversity. To better assess and predict the impacts of more intensive forestry in a changing climate on soils and streams, a more holistic ecosystem approach needs to be taken that includes perspectives from silviculture, ecology, hydrology, and biogeochemistry. Here we review and discuss some of the most pertinent questions, from a Swedish perspective, relevant to the future of both forest soils and waters. Additionally, we suggest approaches that can be used to provide much needed answers to questions about the long-term management of soil and surface water resources in the Swedish forested landscape. Our long-term objective is to move towards an enhanced mechanistic understanding that will lead the way to improved predictive models of ecosystem functioning under changing land use and climate scenarios. The more immediate goal is to provide knowledge that can direct future research efforts and help improve the decision support for forestry and land management communities in order to minimize environmental impacts on soils and waters associated with future biomass production.
2. Impacts of Nutrient Additions
Critical to the development of plans that promote the sustainable use of forest soil and water resources is an understanding of the controls over nutrient availability and retention within terrestrial ecosystems. One nutrient of particular importance is nitrogen (N), which is necessary for plant growth, and in many forests of boreal and temperate regions is the primary limiting factor for net primary production [18,19]. Because most N in soils is bound in relatively recalcitrant soil organic matter not easily accessed by the trees [20], the decomposition of large macromolecules to peptides, amino acids and inorganic N-forms, and the relationships between trees and their symbiotic mycorrhizal fungi are particularly important to the supply of N to primary producers [21,22]. Given the role of N as the primary limiting nutrient in boreal forests, one plan for intensified forestry in the future involves more frequent application of N-based fertilizers to specified production areas [23]. This includes frequent nutrient additions with a mineral balance based on the nutritional requirements of the forest stand [24]. This type of fertilization approach in Norway spruce stands of Sweden can increase stem wood production by more than 200% [25]. It remains unclear, however, whether achieving this increase in production is possible at broad spatial scales, and how such a management approach would affect other ecosystem services that these landscapes provide.
Over the past century, the N economy of forests in many parts of the globe has changed dramatically following increases in atmospheric N deposition linked to the combustion of fossil fuels and intensive agriculture [26]. These increases in N inputs have had major consequences for the ecological and biogeochemical dynamics of forest ecosystems. In general, for N-limited forests, small to moderate N additions (e.g., between 3 and 20 kgN·ha−1·yr−1) stimulate photosynthesis per unit foliage and increase the biomass of leaves and needles, as well as elevate above-ground biomass production relative to that below-ground [27]. Thus, moderate levels of N addition can positively stimulate tree growth, which could represent an important ecosystem C sink. Moreover, N addition may further increase ecosystem C sequestration as these inputs in some cases can have a retarding effect on the decomposition of soil organic matter [28,29].
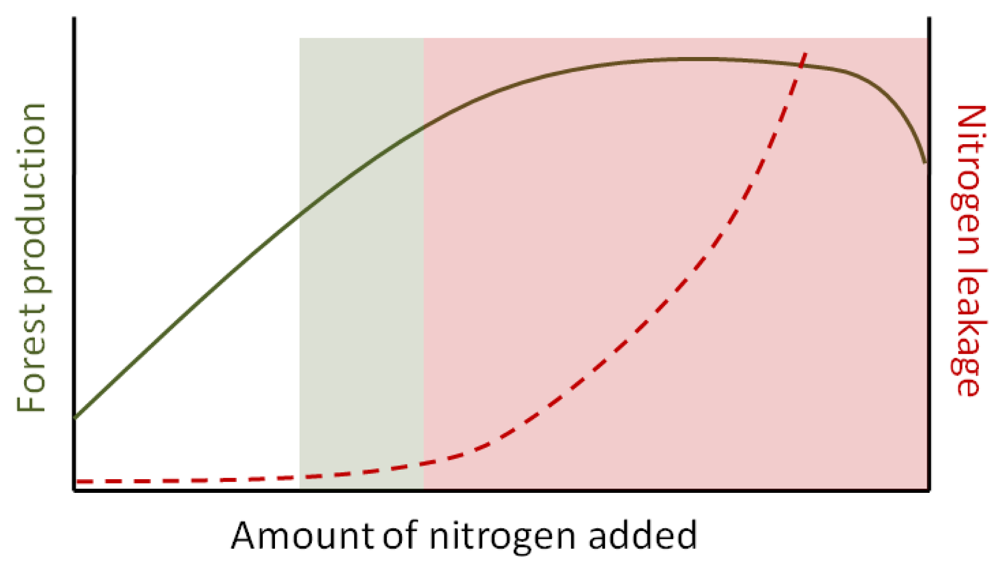
Data from Swedish forests typically show that N fertilization can increase C sequestration [4]. This relationship is non-linear, however, and at higher levels of fertilizer addition, forest ecosystems become saturated as the availability of mineral N exceeds the demand of plants and microbes [30,31]. Under these circumstances, forests lose retentive capacity and begin to leach N from soils to groundwater and streams (Figure 1), mainly in the form of nitrate (NO3−) as this is the most mobile form [32-34]. From a forest owner perspective, added N that is not captured by trees or retained in soil, and which is instead lost from the system, is a costly and unwanted effect of fertilization. Thus, finding a management formula for forest fertilization that will optimize biomass production, while minimizing N leaching, represents a major research priority and will be essential for plans to increase biomass yield and carbon sequestration without seriously affecting water quality (Figure 1).
Because NO3− can act as a pollutant in surface waters [35], an increase in stream N would be an undesirable consequence of more intensive forest fertilization. The potential loading of N to streams following forest fertilization will likely depend on how fertilizer is added (i.e., the timing and application rate), tree nutrient demand (phase of the growth cycle or rotation period), and soil processes that act to retain N in the terrestrial ecosystem [23]. Most terrestrial ecosystems have a large capacity to retain N, but the threshold at which forests begin to lose N, and the mechanisms by which N can be retained within landscapes are not fully understood. If fertilizers are added at low to moderate rates, at times of high plant demand, it is possible that long-term fertilization would have few consequences for nutrient loading in Swedish streams and downstream in the Baltic Sea. Forestry operations, specifically final felling, is presently a minor contributor to NO3− pollution in Swedish surface waters [36,37]. However, escalating the use of fertilizers may cause greater N pollution if this practice is not based on a solid, fundamental understanding of the factors that regulate nutrient retention in forest ecosystems. On the other hand, increased N retention in young, managed and productive stands may help reduce N leakage, compared to old-growth forests at “steady state” [38].
Nitrate leakage with increased fertilization intensity could contribute to the eutrophication and reduction of species diversity in surface waters [39]. Once in the stream, the fate of NO3− depends upon the strength and nature of nutrient limitation, the types and rates of biological processes that occur locally (e.g., assimilation vs. denitrification), and the balance between biological demand and hydrologic transport. There is evidence that in-stream processing can reduce the downstream losses of N from some landscapes [40], but understanding the role of streams and rivers in this regard remains an active area of research. Nutrient enrichment in streams is often associated with a suite of other land use changes (e.g., transitions to agriculture) that also alter habitat structure and the light regime, which together influence local communities and ecosystem processes. The effects of nitrogen enrichment in streams draining intact forests, however, are less clear. Evidence from a multi-year enrichment experiment (of N and P) in a temperate forest stream suggests that nutrient loading can increase invertebrate secondary production [41], alter detritus processing and the overall stream carbon balance [42], and result in unexpected, long-term changes in macroinvertebrate community composition [43]. It is not clear how similar enrichment would influence small, humic-rich boreal streams. There is, however, some evidence for N limitation of dissolved organic carbon (DOC) processing by microbes in streams and lakes in northern Sweden [44], but the broader potential consequences of enrichment at community and ecosystem levels are largely unknown. Of particular interest is whether and how changing inorganic nutrient dynamics in forests will alter the delivery, form and processing of organic nutrients in adjacent and downstream aquatic ecosystems.
Elevated levels of NO3− in streams and lakes also pose additional environmental concerns. For example, such increases in concentration are known to promote denitrification and associated production of N2O, a known greenhouse gas [45]. This risk is expected to be lower in forest, than agricultural and urban streams, which commonly are more enriched with anthropogenic NO3− [40]; however, these microbial processes remain to be evaluated in boreal forest streams subject to elevated N loading. Additionally, NO3− leaching may cause soil acidification. As NO3− ions percolate through the soil into the stream water, accompanying base cations (i.e., Ca2+, Mg2+ and K+) or other cations from forest soils move out of the soil into stream water as well. As more base cations are leached, the capacity of forest soils to buffer changes in pH will be reduced. This could lead to more acidification of susceptible soils and undesirable cation movements from the soil to stream water (i.e., H+, Al3+). An increasing proportion of the acidity in surface waters in North America and Europe is related to NO3− export [46], which can be especially problematic in areas where large amounts of N fertilizer have been added such as intensively managed forests or agricultural land [9,47]. Thus, increased N inputs over long time-scales could actually reduce the fertility of forest soils [47]. Although not an issue in Sweden so far, base cation depletion has been a major problem in parts of central Europe, where levels of atmospheric N deposition have been up to 30–40 kgN·ha−1·yr−1 [48,49], which is several times higher than anywhere in Sweden.
3. Impacts of Tree Harvesting
Tree harvesting has obvious and dramatic effects on the structure and function of terrestrial ecosystems [9]. With the introduction of whole-tree and full-tree harvesting, greater attention has been devoted to understanding the effects of felling operations on forest soils, including effects on biodiversity [50], and the potential for long-term sustainability of tree production owing to increased nutrient losses [51,52]. In addition to these effects on forest and soil properties, it is also long recognized that forest harvesting can influence the ecology and biogeochemistry of adjacent headwater streams [53,54]. Indeed, the general recognition of key linkages between terrestrial landscapes and their associated streams has a long history in ecology [55,56], and a rich scientific literature exists investigating the relationships between forest management practices, water chemistry, and stream discharge, illustrating how closely linked small streams are to their surrounding forests [57,58]. From this work, it is well established that clear-felling results in altered hydrology: decreased evapotranspiration, increased groundwater tables, and therefore greater runoff [59,60].
Given the central role of the hydrologic regime in the ecology and biogeochemistry of catchments, changes to the hydrologic cycle following forest harvesting are likely to have consequences for both terrestrial and aquatic ecosystems. The effects of altered flow may include changes in the rates of biogeochemical processes in both terrestrial and aquatic habitats (e.g., decomposition, nutrient cycling), and the timing and magnitude of downstream hydrologic transport of dissolved and particulate materials. For example, recent research has shown that clear-cutting can increase both the concentration and total flux of DOC in streams of boreal regions [61,62]. Such an increase is particularly important in boreal landscapes, not only because DOC represents a potentially important flux in the regional C cycle [63], but also because DOC acts as an important transport-vector for contaminants such as mercury [64] and persistent organic pollutants [65], and because it represents a major source of energy to foodwebs in downstream lakes [66].
In addition to these effects on DOC, forest harvesting also can result in losses of nutrients and other elements from forest soils with corresponding increases in the concentration of these materials in adjacent streams [54]. There is some evidence, however, that whole-tree harvesting may actually reduce short term NO3− losses [67]. According to this hypothesis, removing tops, branches, and needles substantially reduces logging residues and the corresponding detrital N pool, which could potentially reduce NO3− leaching immediately following harvesting [68-70]. However, base cations are also lost from sites during whole-tree harvesting [70]. Given the role of base cations for controlling pH in surface waters [71], the long-term consequences of whole-tree harvesting needs further evaluation before it is used as a means to reduce short-term N losses from catchments, particularly as an intensification of forestry in the future could have substantial effects on weakly-buffered streams in Sweden [72].
Forestry activities, including clear-felling and off-road transports, also have the potential to increase toxic methyl mercury concentrations in surface waters [73,74]. Further research is needed on the link between forestry and elevated levels of methyl mercury in surface waters. It is possible that the elevated levels of methyl mercury are related to the wetter ground conditions often seen after final harvest, or to soil disturbance associated with harvest and site preparation. More recent results have also suggested a large degree of variation in the sensitivity of different catchments to harvest operations with respect to mercury response [60]. Understanding the causes of the varying sensitivity may hold the key to more effective measures to mitigate the effects of forestry operations on mercury outputs.
Forest harvesting can also influence streams and rivers through alterations of key habitat features, which may persist for variable lengths of time [53]. For example, removal of streamside vegetation can temporarily increase incident light, which may elevate stream temperatures [75], and stimulate in-stream primary production and ecosystem respiration [76]. These effects of increasing light on benthic metabolism may be exacerbated when coincident with elevated nutrient concentrations [77]. Consistent with this, studies of clear-cutting in boreal streams show increases in benthic algal and bacterial productivity that correspond to both elevated incident light, and increases in stream nutrient availability [78,79]. Tree harvesting and near-stream management can also greatly impact sediment delivery and inputs of woody debris, both of which can have long-term effects on channel structure [80], and play a key role in the ecology and biogeochemistry of forest streams [81]. This effect of forestry on channel form has received little attention in boreal regions; however, a study of streams in Finland and Russia has demonstrated that forest management can affect the structure of stream channels, including a reduction in both the abundance of coarse woody debris and fine detritus [82].
As suggested above, the effects of forestry on aquatic resources at multiple time scales could be exaggerated by expected future increases in precipitation [83]. In Scandinavia, global climate models predict there will be shorter winters and precipitation events will likely be more variable, both in intensity and frequency [84]. Consequently, the length of the snow-covered season, the timing and magnitude of spring runoff, and the number of extreme events could be altered in the future [85]. Shorter periods with frozen ground could have profound influences on the physical effects of harvesting on forest soils. For example, forest machinery operating on unfrozen, wet ground will potentially have much larger and longer-lasting impacts on soil physical structure and surface water quality, when compared to these same activities on frozen ground. This could be further exaggerated when logging residues are harvested as part of a whole-tree or full-tree harvesting strategy instead of the conventional use as bedding on the skid road to avoid soil damages [86]. Overall, understanding the interactive effects of potential climate change and land management on aquatic resources represents an important research goal, and progress along these lines will require close collaboration among foresters and scientists.
4. Future Research Needs
To address the above problems and incorporate them into practical use, forest management has to be based on sound scientific understanding, and an improved dialogue between foresters and multiple scientific communities. Reaching this goal will require the integration of scientific approaches that span traditional disciplinary boundaries. This integration should include compiling available data from long-term descriptive and experimental studies, new innovative experimental manipulations, and both analytical and simulation modeling to evaluate questions across a range of scales, using multiple methods of inference. These research efforts need to be designed a priori to address questions relevant to forest management, and should also include interaction and feedback from stakeholders and managers according to an adaptive forest management approach. Such an approach may help to provide answers to questions required for the transition into a new era of environmental stewardship that focuses on maintaining, or even increasing, biomass production without jeopardizing soils or waters.
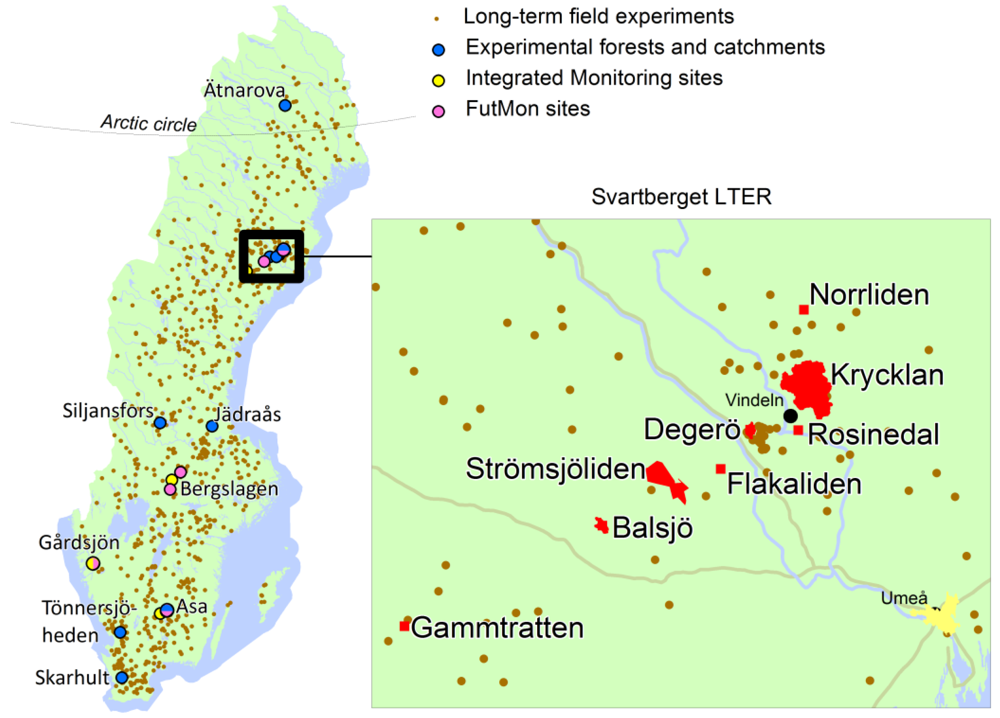
Historically, a main limitation to answering questions that integrate large spatial and temporal scales of forest, soils, and streams has been the availability of well organized, high-quality field data. There is, however, a unique opportunity emerging in Sweden to use recently compiled field data from hundreds of forest and soil experiments across the country, several established in the first half of the 20th century, and others more newly established within the Long-term Ecological Research (LTER) platform (Figure 2). The integration of research efforts among these sites would serve to establish an extensive network of geographically distributed experiments that also includes well-studied ecosystems within the network. One example of such an LTER platform is the Svartberget LTER site that constitutes the base for several large-scale, long-term research programs. Field research began in this area in the early 1920s and includes ecological studies on the effects of different forest management practices [27,58,87,88], climate change [89,90], and long-range transport of air pollutants on soils and water quality [65,91]. This area has also been used to develop a complete carbon budget of boreal mires and forests [92], and is home to long-term experiments on tree-soil interactions following conventional fertilization and nutrient optimization [27,93], and whole-tree harvesting [11,94-96]. As a result of these research activities, the Svartberget LTER infrastructure provides outstanding opportunities to study how different silvicultural practices influence soils and water quality in boreal forests.
Future forest management that includes an increased use of fertilizers will need to be based on a clear understanding of the underlying mechanisms of N cycling and retention in boreal forest ecosystems. A key aspect of this cycle involves symbiotic relationships between plants and mycorrhizal fungi, and determining the effects of N addition on this relationship remains an important research frontier. One testable hypothesis in this context is that the mycorrhizal fungi play a key role in N retention in boreal forests and that these groups are negatively affected by increased N supply [97]. An important question is whether functional populations of these fungi can be maintained under conditions of high N supply and if not, what happens to the capacity of forest ecosystems to retain nutrients? To move from merely speculation, based on individual studies, to an understanding of the long-term consequences of N fertilization there is a need to combine controlled, experimental case-studies with larger meta-analyses that involve in-depth reviews of published data. Together, these approaches can help deliver predictive models to forest managers that describe the effects of atmospheric N deposition and forest fertilization on mycorrhizal populations, and provide insight into how altering the activity of these soil fungi will influence broad-scale biogeochemical cycles.
To better understand how both conventional and future tree harvesting strategies will influence soil and water quality, we need a better mechanistic understanding of the processes controlling the delivery of materials from terrestrial to aquatic ecosystems, and the associated ecological and biogeochemical consequences of these transfers. One important question in this context centers on the availability of base cations in the soil. While there may not be an immediate threat to the long-term productivity in Sweden [98], little is known about how intensified forestry, in combination with potentially longer growing seasons, will affect soil base cations. Results from weathering models indicate that forest growth will be negatively affected by deficiencies in the supply of base cations in the soil following both conventional and whole-tree harvesting [99]. Furthermore these models predict that forest soils will take decades or centuries to recover from such losses [100]. Conversely, large-scale experimental manipulations suggest that decreases in the concentration of exchangeable base cations in soils do not necessarily limit forest growth in Sweden [98]. Recent results also suggest that the rate of base cation recovery following the termination of N addition is much more rapid than predicted by weathering models [93]. In a recent study from the Svartberget LTER, Klaminder et al. [101] showed that seven different weathering models produced contrasting results with a precision far from what is needed in order to predict any forestry related effects on the base cation pool. This type of result demonstrates the need for further research on this topic, the potential pitfalls associated with the over-reliance on individual models, and the benefits of using model ensembles to produce more realistic results and estimates of uncertainty [102].
Another research priority is to understand how proposed management practices will affect aquatic ecosystems, and how strategies can be developed that may mitigate these effects. For example, how might changes in water chemistry and habitat quality resulting from forest management be propagated through aquatic foodwebs, and downstream through drainage networks that also include wetlands and lakes? In this context, what is the role of landscape heterogeneity, and what are the ranges of spatial and temporal scales at which these management effects are manifested in aquatic ecosystems? Finally, to what degree are these land-water linkages buffered by riparian vegetation and organic soils? Indeed, refining our understanding of the role of riparian zones seems particularly important [103]. There is evidence that certain aspects of water quality may be maintained by riparian processes, which can include the retention of nutrients and sediments from upslope terrestrial habitats [104]. Given the many functions of the riparian zone, leaving buffer zones at harvesting has become a standard procedure along streams and rivers in Sweden. More could be done, however, to improve the functionality of these areas by better incorporating aspects related to riparian ecology, erosion control, biogeochemical hotspots, and groundwater discharge into the management decision.
Other management approaches that are promising from a water quality perspective involve organizing landscapes into areas that are highly sensitive to both traditional and intensive forestry and those locations that are less so. By applying this ‘landscape sensitivity’ approach, greater care can be devoted to protecting certain areas from (for example) off-road transport or intensive forest harvesting practices. One example of this is that areas at the interface of mineral and organic soils are hotspots of methyl mercury production. If these locations are also in close connection to streams, rutting by forest machinery could cause not only increased methylisation, but also rapid hydrological connection to adjacent surface waters. This general management approach will require more input from scientists related to how intrinsic properties of forest ecosystems (e.g., landscape position, slope, underlying geology, soil texture, etc.) influence this degree of sensitivity to various harvesting practices. Another way forward will include the use of planning tools designed to help managers develop strategies for maximizing forest yield in the long term, while maintaining particular water quality parameters below specific thresholds [105]. Furthermore, optimized use of fertilizers, differential management of sensitive forest stands, a better understanding of the functionality of riparian areas, and implementation of improved planning tools could lead the way towards more productive and sustainable forestry in the future.
5. Summary
Trees, soils and water are at the center for understanding how Swedish forests will be affected by intensified biomass production and climate change. While increased production of forest biomass will potentially help mitigate predicted climate changes, the negative effects on soils and/or the deterioration of water quality that may arise could influence other ecosystem services such as future forest productivity, biodiversity, and recreation. However, some of these potential negative impacts can be reduced by prudent forest management. While we are beginning to better understand how forestry affects forest soils and waters, the synergistic effects of climate change and land management are almost entirely unknown. Reliance on inadequate knowledge of the relationships among climate change, tree harvesting, and soil and water quality represents a profound risk of mismanaging two of the most abundant, but also most precious natural resources in Sweden. Future research therefore needs to provide applicable information about plant-soil and soil-water interactions that is valuable to society and forest managers and that ensures intensified forestry is performed without jeopardizing forest soils and water quality.


Acknowledgments
The research was funded through Future Forests, a multi-disciplinary research program supported by the Foundation for Strategic Environmental Research (MISTRA), the Swedish Forestry Industry, the Swedish University of Agricultural Sciences (SLU), Umeå University, and the Forestry Research Institute of Sweden.
References and Notes
- Gilliam, F.S.; Dick, D.A.; Kerr, M.L.; Adams, M.B. Effects of silvicultural practices on soil carbon and nitrogen in a nitrogen saturated central Appalachian (USA) hardwood forest ecosystem. Environ. Manage. 2004, 33, S108–S119. [Google Scholar]
- National Research Council. In Grand Challenges in Environmental Sciences; National Academy Press: Washington, DC, USA, 2001; p. 96.
- IPCC climate change 2007: The physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007.
- Hyvönen, R.; Ågren, G.I.; Linder, S.; Persson, T.; Cotrufo, M.F.; Ekblad, A.; Freeman, M.; Grelle, A.; Janssens, I.A.; Jarvis, P.G.; et al. The likely impact of elevated [CO2], nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: A literature review. New Phytol. 2007, 173, 463–480. [Google Scholar]
- Mack, M.C.; Schuur, E.A.G.; Bret-Harte, M.S.; Shaver, G.R.; Chapin, F.S., III. Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature 2004, 431, 440–443. [Google Scholar]
- Ericsson, K.; Huttunen, S.; Nilsson, L.J.; Svenningsson, P. Bioenergy policy and market development in Finland and Sweden. Energy Policy 2004, 32, 1707–1721. [Google Scholar]
- Egnell, G.; Laudon, H.; Rosvall, O. The role of forest biomass in future energy use—Potential and constraints. Forests 2011. submitted. [Google Scholar]
- Energy in Sweden; Sweden Energy Agency: Eskilstuna, Sweden, 2009.
- Fox, T.R. Sustained productivity in intensively managed forest plantations. For. Ecol. Manage. 2000, 138, 187–202. [Google Scholar]
- Hedwall, P.O.; Nordin, A.; Brunet, J.; Bergh, J. Compositional changes of forest-floor vegetation in young stands of Norway spruce as an effect of repeated fertilisation. For. Ecol. Manage. 2010, 259, 2418–2425. [Google Scholar]
- Olsson, B.A.; Bengtsson, J.; Lundkvist, H. Effects of different forest harvest intensities on the pools of exchangeable cations in coniferous forest soils. For. Ecol. Manage. 1996, 84, 135–147. [Google Scholar]
- Swank, W.T.; Waide, J.B. Interpretation of nutrient cycling research in a management context: Evaluating potential effects of alternative management strategies on site productivity. In Forests: Fresh Perspectives from Ecosystem Analysis; Waring, R.H., Ed.; Oregon State Universtiy Press: Corvallis, OR, USA, 1980; pp. 137–158. [Google Scholar]
- Briggs, R.D.; Hornbeck, J.W.; Smith, C.T.; Lemin, R.C.; McCormack, M.L. Long-term effects of forest management on nutrient cycling in spruce-fir forests. For. Ecol. Manage. 2000, 138, 285–299. [Google Scholar]
- Jonsson, M.; Wardle, D.A. Structural equation modelling reveals plant-community drivers of carbon storage in boreal forest ecosystems. Biol. Lett. 2010, 6, 116–119. [Google Scholar]
- Berg, B.; Johansson, M.B.; Nilsson, A.; Gundersen, P.; Norell, L. Sequestration of carbon in the humus layer of Swedish forests—Direct measurements. Can. J. For. Res. 2009, 39, 962–975. [Google Scholar]
- Wardle, D.A.; Hörnberg, G.; Zackrisson, O.; Kalela-Brundin, M.; Coomes, D.A. Long-term effects of wildfire on ecosystem properties across an island area gradient. Science 2003, 300, 972–975. [Google Scholar]
- Heino, J.; Virkkala, R.; Toivonen, H. Climate change and freshwater biodiversity: Detected patterns, future trends and adaptations in northern regions. Biol. Rev. 2009, 84, 39–54. [Google Scholar]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar]
- Tamm, C.O. Nitrogen in Terrestrial Ecosystems—Questions of Productivity, Vegetational Changes, and Ecosystem Stability; Ecological Studies 81; Springer Verlag: Berlin, Germany, 1991; p. 115. [Google Scholar]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar]
- Langley, J.A.; Hungate, B.A. Mycorrhizal controls on belowground litter quality. Ecology 2003, 84, 2302–2312. [Google Scholar]
- Treseder, K.K.; Turner, K.M.; Mack, M.C. Mycorrhizal responses to nitrogen fertilization in boreal ecosystems: Potential consequences for soil carbon storage. Glob. Change Biol. 2007, 13, 78–88. [Google Scholar]
- Nordin, A.; Bergström, A.-K.; Granberg, G.; Grip, H.; Gustafsson, D.; Gärdenäs, A.M.; Hyvönen-Olsson, R.; Jansson, P.-E.; Laudon, H.; Nilsson, M.B.; Svensson, M.; Öquist, M. Effekter av ett intensivare skogsbruk på skogslandskapets mark, vatten och växthusgaser; SLU: Uppsala, Sweden, 2009; p. 46. [Google Scholar]
- Linder, S. Foliar analysis for detecting and correcting nutrient imbalances in Norway spruce. Ecol. Bull. (Copenhagen) 1995, 44, 178–190. [Google Scholar]
- Bergh, J.; Linder, S.; Bergström, J. Potential production of Norway spruce in Sweden. For. Ecol. Manage. 2005, 204, 1–10. [Google Scholar]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.C.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar]
- Iivonen, S.; Kaakinen, S.; Jolkkonen, A.; Vapaavuori, E.; Linder, S. Influence of long-term nutrient optimization on biomass, carbon, and nitrogen acquisition and allocation in Norway spruce. Can. J. For. Res. 2006, 36, 1563–1571. [Google Scholar]
- Berg, B.; McClaugherty, C.; De Santo, A.V.; Johnson, D. Humus buildup in boreal forests: Effects of litter fall and its N concentration. Can. J. For. Res. 2001, 31, 988–998. [Google Scholar]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar]
- Aber, J.D.; McDonald, W.; Nadelhoffer, K.J.; Magill, A.; Bernston, G.; Kamakea, M.; McNulty, S.; Currie, W.S.; Rustad, L.; Fernandez, I. Nitrogen saturation in temperate forest ecosystems: Hypothesis revisited. BioScience 1998, 48, 921–934. [Google Scholar]
- Aber, J.D.; Nadelhoffer, K.J.; Steudler, P.; Melillo, J.M. Nitrogen saturation in northern forest ecosystems. BioScience 1989, 39, 378–386. [Google Scholar]
- Aber, J.D.; Goodale, C.L.; Ollinger, S.V.; Smith, M.-L.; Magill, A.; Martin, M.E.; Hallett, R.A.; Stoddard, J.L. Is nitrogen deposition altering the nitrogen status of northeastern forests? BioScience 2003, 53, 375–389. [Google Scholar]
- Dise, N.B.; Wright, R.F. Nitrogen leaching from European forest in relation to nitrogen deposition. For. Ecol. Manage. 1995, 71, 153–161. [Google Scholar]
- Beier, C.; Blanck, K.; Bredemeier, M.; Lamersdorf, N.; Rasmussen, L.; Xu, Y.J. Field-scale ‘clean rain’ treatments to two Norway spruce stands within the EXMAN project—Effects on soil solution chemistry, foliar nutrition and tree growth. For. Ecol. Manage. 1998, 101, 111–123. [Google Scholar]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar]
- Futter, M.N.; Ring, E.; Högbom, L.; Entenmann, S.; Bishop, K.H. Consequences of nitrate leaching following conventional harvesting of Swedish forests are dependent on spatial scale. Environ. Pollut. 2010, 158, 3552–3559. [Google Scholar]
- Brandt, M.; Ejhed, H.; Rapp, L. Nutrient loads to the Swedish marine environment in 2006. In Swedish Environmental Protection Agency Report 5815; Naturvårdsverket: Stockholm, Sweden, 2009; p. 93. [Google Scholar]
- Vitousek, P.M.; Reiners, W.A. Ecosystem succession and nutrient retention—Hypothesis. Bioscience 1975, 25, 376–381. [Google Scholar]
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar]
- Mulholland, P.J.; Helton, A.M.; Poole, G.C.; Hall, R.O.; Hamilton, S.K.; Peterson, B.J.; Tank, J.L.; Ashkenas, L.R.; Cooper, L.W.; Dahm, C.N.; et al. Stream denitrification across biomes and its response to anthropogenic nitrate loading. Nature 2008, 452, 202–205. [Google Scholar]
- Cross, W.F.; Wallace, J.B.; Rosemond, A.D.; Eggert, S.L. Whole-system nutrient enrichment increases secondary production in a detritus-based ecosystem. Ecology 2006, 87, 1556–1565. [Google Scholar]
- Benstead, J.P.; Rosemond, A.D.; Cross, W.F.; Wallace, J.B.; Eggert, S.L.; Suberkropp, K.; Gulis, V.; Greenwood, J.L.; Tant, C.J. Nutrient enrichment alters storage and fluxes of detritus in a headwater stream ecosystem. Ecology 2009, 90, 2556–2566. [Google Scholar]
- Davis, J.M.; Rosemond, A.D.; Eggert, S.L.; Cross, W.F.; Wallace, J.B. Long-term nutrient enrichment decouples predator and prey production. PNAS 2010, 107, 121–126. [Google Scholar]
- Berggren, M.; Laudon, H.; Jansson, M. Landscape regulation of bacterial growth efficiency in boreal freshwaters. Glob. Biogeochem. Cycle 2007, 21, 2844–2852. [Google Scholar]
- Beaulieu, J.J.; Tank, J.L.; Hamilton, S.K.; Wollheim, W.M.; Hall, R.O.; Mulholland, P.J.; Peterson, B.J.; Ashkenas, L.R.; Cooper, L.W.; Dahm, C.N.; et al. Nitrous oxide emission from denitrification in stream and river networks. PNAS 2011, 108, 214–219. [Google Scholar]
- Stoddard, J.L.; Jeffries, D.S.; Lükewille, A.; Clair, T.A.; Dillon, P.J.; Driscoll, C.T.; Forsius, M.; Johannessen, M.; Kahl, J.S.; Kellogg, J.H.; et al. Regional trends in aquatic recovery from acidification in North America and Europe. Nature 1999, 401, 575–578. [Google Scholar]
- Gundersen, P.; Schmidt, I.K.; Raulund-Rasmussen, K. Leaching of nitrate from temperate forests—Effects of air pollution and forest management. Environ. Rev. 2006, 14, 1–57. [Google Scholar]
- Hofmeister, J.; Oulehle, F.; Kram, P.; Hruska, J. Loss of nutrients due to litter raking compared to the effect of acidic deposition in two spruce stands, Czech Republic. Biogeochem. 2008, 88, 139–151. [Google Scholar]
- Jandl, R.; Alewell, C.; Prietzel, J. Calcium loss in Central European forest soils. Soil Sci. Soc. Am. J. 2004, 68, 588–595. [Google Scholar]
- Martikainen, P.; Siitonen, J.; Punttila, P.; Kaila, L.; Rauh, J. Species richness of Coleoptera in mature managed and old-growth boreal forests in southern Finland. Biol. Conserv. 2000, 94, 199–209. [Google Scholar]
- Johnson, D.W.; Todd, D.E. Harvesting effects on long-term changes in nutrient pools of mixed oak forest. Soil Sci. Soc. Am. J. 1998, 62, 1725–1735. [Google Scholar]
- Walmsley, J.; Godfold, D. Stump harvesting for Bioenergy—A review of the environmental impacts. Forestry 2010, 83, 17–38. [Google Scholar]
- Webster, J.R.; Golladay, S.W.; Benfield, E.F.; Meyer, J.L.; Swank, W.T.; Wallace, J.B. Catchment disturbance and stream response: An overview of stream research at Coweeta Hydrologic Laboratory. In River Conservation and Management; Boon, P.J., Calow, P., Petts, G.E., Eds.; John Wiley and Sons: Chichester, UK, 1992; pp. 231–253. [Google Scholar]
- Kreutzweiser, D.P.; Hazlett, P.W.; Gunn, J.M. Logging impacts on the biogeochemistry of boreal forest soils and nutrient export to aquatic systems: A review. Environ. Rev. 2008, 16, 157–179. [Google Scholar]
- Hynes, H. The stream and its valley. Verhandlungen der Internationalen Vereinigung fur theoretische und angewandte Limnologie 1975, 19, 1–15. [Google Scholar]
- Allan, J.D. Landscape and riverscapes: The influence of land use on river ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar]
- Likens, G.E.; Bormann, F.H.; Johnson, N.M.; Fisher, D.W.; Pierce, R.S. Effects of forest cutting and herbicide treatment on nutrient budgets in Hubbard Brook watershed-ecosystem. Ecol. Monogr. 1970, 40, 23–47. [Google Scholar]
- Löfgren, S.; Ring, E.; von Brömssen, C.; Sørensen, R.; Högbom, L. Short-term effects of clear-cutting on the water chemistry of two boreal streams in northern Sweden: A paired catchment study. Ambio 2009, 38, 347–356. [Google Scholar]
- Andreassian, V. Waters and forests: From historical controversy to scientific debate. J. Hydrol. 2004, 291, 1–27. [Google Scholar]
- Sørensen, R.; Ring, E.; Meili, M.; Högbom, L.; Seibert, J.; Grabs, T.; Laudon, H.; Bishop, K. Forest harvest increases runoff most during low flows in two boreal streams. Ambio 2009, 38, 357–363. [Google Scholar]
- Cummins, T.; Farrell, E.P. Biogeochemical impacts of clearfelling and reforestation on blanket-peatland streams—II. Major ions and dissolved organic carbon. For. Ecol. Manage. 2003, 180, 557–570. [Google Scholar]
- Laudon, H.; Hedtjärn, J.; Schelker, J.; Bishop, K.; Sørensen, R.; Ågren, A. Response of dissolved organic carbon following forest harvesting in a boreal forest. Ambio 2009, 38, 381–386. [Google Scholar]
- Cole, J.J.; Prairie, Y.T.; Caraco, N.F.; McDowell, W.H.; Tranvik, L.J.; Striegl, R.G.; Duarte, C.M.; Kortelainen, P.; Downing, J.A.; Middelburg, J.J.; Melack, J. Plumbing the global carbon cycle: Integrating inland waters into the terrestrial carbon budget. Ecosystems 2007, 10, 171–184. [Google Scholar]
- Porvari, P.; Verta, M.; Munthe, J.; Haapanen, M. Forestry practices increase mercury and methyl mercury output from boreal forest catchments. Environ. Sci. Technol. 2003, 37, 2389–2393. [Google Scholar]
- Bergknut, M.; Laudon, H.; Wiberg, K. Dioxins, PCBs, and HCB in soil and peat profiles from a pristine boreal catchment. Environ. Pollut. 2010, 158, 2518–2525. [Google Scholar]
- Berggren, M.; Ström, L.; Laudon, H.; Karlsson, J.; Jonsson, A.; Giesler, R.; Bergström, A.K.; Jansson, M. Lake secondary production fueled by rapid transfer of low molecular weight organic carbon from terrestrial sources to aquatic consumers. Ecol. Lett. 2010, 13, 870–880. [Google Scholar]
- Lundborg, A. Reducing the nitrogen load: Whole-tree harvesting—A literature review. Ambio 1997, 26, 387–393. [Google Scholar]
- Slesak, R.A.; Schoenholtz, S.H.; Harrington, T.B.; Strahm, B.D. Dissolved carbon and nitrogen leaching following variable logging-debris retention and competing-vegetation control in Douglas-fir plantations of western Oregon and Washington. Can. J. For. Res. 2009, 39, 1484–1497. [Google Scholar]
- Staaf, H.; Olsson, B.A. Effects of slash removal and stump harvesting on soil water chemistry in a clearcutting in SW Sweden. Scand. J. Forest Res. 1994, 9, 305–310. [Google Scholar]
- Wall, A. Effect of removal of logging residue on nutrient leaching and nutrient pools in the soil after clearcutting in a Norway spruce stand. For. Ecol. Manage. 2008, 256, 1372–1383. [Google Scholar]
- Reuss, J.O.; Johnson, D.W. Effect of soil processes on the acidification of water by acid deposition. J. Environ. Qual. 1985, 14, 26–31. [Google Scholar]
- Ågren, A.; Buffam, I.; Bishop, K.; Laudon, H. Sensitivity of pH in a boreal stream network to a potential decrease in base cations caused by forest harvest. Can. J. Fish. Aquat. Sci. 2010, 67, 1116–1125. [Google Scholar]
- Munthe, J.; Hultberg, H. Mercury and methylmercury in runoff from a forested catchment— Concentrations, fluxes, and their response to manipulations. Water Air Soil Pollut. Focus 2004, 4, 607–618. [Google Scholar]
- Bishop, K.; Allan, C.; Bringmark, L.; Garcia, E.; Hellsten, S.; Högbom, L.; Johansson, K.; Lomander, A.; Meili, M.; Munthe, J.; et al. The effects of forestry on Hg bioaccumulation in nemoral/boreal waters and recommendations for good silvicultural practice. Ambio 2009, 38, 373–380. [Google Scholar]
- Burton, T.M.; Likens, G.E. Effect of strip-cutting on stream temperatures in Hubbard Brook Experimental Forest, New Hampshire. BioScience 1973, 23, 433–435. [Google Scholar]
- Clapcott, J.E.; Barmuta, L.A. Forest clearance increases metabolism and organic matter processes in small headwater streams. J. North Am. Benthol. Soc. 2010, 29, 546–561. [Google Scholar]
- Bernot, M.J.; Sobota, D.J.; Hall, R.O.; Mulholland, P.J.; Dodds, W.K.; Webster, J.R.; Tank, J.L.; Ashkenas, L.R.; Cooper, L.W.; Dahm, C.N.; et al. Inter-regional comparison of land-use effects on stream metabolism. Freshw. Biol. 2010, 55, 1874–1890. [Google Scholar]
- Holopainen, A.L.; Huttunen, P. Effects of forest clear-cutting and soil disturbance on the biology of small forest brooks. Hydrobiologia 1992, 243, 457–464. [Google Scholar]
- McKie, B.G.; Malmqvist, B. Assessing ecosystem functioning in streams affected by forest management: increased leaf decomposition occurs without changes to the composition of benthic assemblages. Freshw. Biol. 2009, 54, 2086–2100. [Google Scholar]
- Harding, H.E.; Benfield, J.S.; Bolstad, E.F.; Helfman, P.V.; Jones, E.B.D., III. Stream biodiversity: The ghost of land use past. PNAS 1998, 95, 14843–14847. [Google Scholar]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Liljaniemi, P.; Vuori, K.M.; Ilyashuk, B.; Luotonen, H. Habitat characteristics and macroinvertebrate assemblages in boreal forest streams: Relations to catchment silvicultural activities. Hydrobiologia 2002, 474, 239–251. [Google Scholar]
- Weslien, J.; Finer, L.; Jonsson, J.A.; Koivusalo, H.; Lauren, A.; Ranius, T.; Sigurdsson, B.D. Effects of increased forest productivity and warmer climates on carbon sequestration, run-off water quality and accumulation of dead wood in a boreal landscape: A modelling study. Scand. J. For. Res. 2009, 24, 333–347. [Google Scholar]
- Kjellström, E.; Nikulin, G.; Hannson, U.; Strandberg, G.; Ullerstig, A. 21st century changes in the European climate: Uncertainties derived from an ensemble of regional climate model simulations. Tellus 2010, 63, 24–40. [Google Scholar]
- Nikulin, G.; Kjellström, E.; Hannson, U.; Strandberg, G.; Ullerstig, A. Evaluation and future projections of temperature, precipitation and wind extremes over Europe in an ensemble of regional climate simulations. Tellus 2010, 63, 41–55. [Google Scholar]
- Eliasson, L.; Wästerlund, I. Effects of slash reinforcement of strip roads on rutting and soil compaction on a moist fine-grained soil. For. Ecol. Manage. 2007, 252, 118–123. [Google Scholar]
- Tamm, O. The Experimental Forests of Kulbäcksliden and Svartberget in North Sweden; Centraltryckeriet: Stockholm, Sweden, 1926; p. 87. [Google Scholar]
- Tamm, C.O.; Aronsson, A.; Popovic, B. Optimum nutrition and nitrogen saturation in Scots pine stands. Stud. For. Suec. 1999, 206, 1–126. [Google Scholar]
- Köhler, S.J.; Buffam, I.; Seibert, J.; Bishop, K.H.; Laudon, H. Dynamics of stream water TOC concentrations in a boreal headwater catchment: Controlling factors and implications for climate scenarios. J. Hydrol. 2009, 373, 44–56. [Google Scholar]
- Mellander, P.E.; Ottosson, M.; Laudon, H. Climate change impact on snow and soil temperature in boreal Scots pine stands. Clim. Change 2007, 85, 179–193. [Google Scholar]
- Klaminder, J.; Bindler, R.; Laudon, H.; Bishop, K.; Emteryd, O.; Renberg, I. Flux rates of atmospheric lead pollution within soils of a small catchment in northern Sweden and their implications for future stream water quality. Environ. Sci. Technol. 2006, 40, 4639–4645. [Google Scholar]
- Nilsson, M.; Sagerfors, J.; Buffam, I.; Laudon, H.; Eriksson, T.; Grelle, A.; Klemedtsson, L.; Weslien, P.; Lindroth, A. Contemporary carbon accumulation in a boreal oligotrophic minerogenic mire—A significant sink after accounting for all C-fluxes. Glob. Change Biol. 2008, 14, 2317–2332. [Google Scholar]
- Högberg, P.; Fan, H.B.; Quist, M.; Binkley, D.; Tamm, C.O. Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest. Glob. Change Biol. 2006, 12, 489–499. [Google Scholar]
- Olsson, B.A.; Lundkvist, H.; Staaf, H. Nutrient status in needles of Norway spruce and Scots pine following harvesting of logging residues. Plant Soil 2000, 223, 161–173. [Google Scholar]
- Egnell, G.; Leijon, B. Survival and growth of planted seedlings of Pinus sylvestris and Picea abies after different levels of biomass removal in clear-felling. Scand. J. For. Res. 1999, 14, 303–311. [Google Scholar]
- Egnell, G.; Valinger, E. Survival, growth, and growth allocation of planted Scots pine trees after different levels of biomass removal in clear-felling. For. Ecol. Manage. 2003, 177, 65–74. [Google Scholar]
- Högberg, P.; Johannisson, C.; Yarwood, S.; Callesen, I.; Näsholm, T.; Myrold, D.D.; Högberg, M.N. Recovery of ectomycorrhiza after ‘nitrogen saturation’ of a conifer forest. New Phytol. 2011, 189, 515–525. [Google Scholar]
- Binkley, D.; Högberg, P. Does atmospheric deposition of nitrogen threaten Swedish forests? For. Ecol. Manage. 1997, 92, 119–152. [Google Scholar]
- Akselsson, C.; Westling, O.; Sverdrup, H.; Gundersen, P. Nutrient and carbon budgets in forest soils as decision support in sustainable forest management. For. Ecol. Manage. 2007, 238, 167–174. [Google Scholar]
- Sverdrup, H.; Rosén, K. Long-term base cation mass balances for Swedish forests and the concept of sustainability. For. Ecol. Manage. 1998, 110, 221–236. [Google Scholar]
- Klaminder, J.; Lucas, R.W.; Futter, M.N.; Bishop, K.H.; Köhler, S.J.; Egnell, G.; Laudon, H. Silicate mineral weathering rate estimates: Are they currently precise enough to be useful when predicting the recovery of nutrient pools after harvesting? For. Ecol. Manage. 2011, 261, 1–9. [Google Scholar]
- Viney, N.R.; Bormann, H.; Breuer, L.; Bronstert, A.; Croke, B.F.W.; Frede, H.; Graff, T.; Hubrechts, L.; Huisman, J.A.; Jakeman, A.J.; et al. Assessing the impact of land use change on hydrology by ensemble modelling (LUCHEM) II: Ensemble combinations and predictions. Adv. Water Resour. 2009, 32, 147–158. [Google Scholar]
- Gundersen, P.; Lauren, A.; Finér, L.; Ring, E.; Koivusalo, H.; Saetersdal, M.; Weslien, J.O.; Sigurdsson, B.D.; Högbom, L.; Laine, J.; Hansen, K. Environmental services provided from Riparian Forests in the Nordic countries. Ambio 2010, 39, 555–566. [Google Scholar]
- Peterjohn, W.T.; Correll, D.L. Nutrient dynamics in an agricultural watershed: Observations on the role of a riparian forest. Ecology 1984, 65, 1466–1475. [Google Scholar]
- Öhman, K.; Seibert, J.; Laudon, H. An approach for including consideration of stream water dissolved organic carbon in long term forest planning. Ambio 2009, 38, 387–393. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Laudon, H.; Sponseller, R.A.; Lucas, R.W.; Futter, M.N.; Egnell, G.; Bishop, K.; Ågren, A.; Ring, E.; Högberg, P. Consequences of More Intensive Forestry for the Sustainable Management of Forest Soils and Waters. Forests 2011, 2, 243-260. https://doi.org/10.3390/f2010243
Laudon H, Sponseller RA, Lucas RW, Futter MN, Egnell G, Bishop K, Ågren A, Ring E, Högberg P. Consequences of More Intensive Forestry for the Sustainable Management of Forest Soils and Waters. Forests. 2011; 2(1):243-260. https://doi.org/10.3390/f2010243
Chicago/Turabian StyleLaudon, Hjalmar, Ryan A. Sponseller, Richard W. Lucas, Martyn N. Futter, Gustaf Egnell, Kevin Bishop, Anneli Ågren, Eva Ring, and Peter Högberg. 2011. "Consequences of More Intensive Forestry for the Sustainable Management of Forest Soils and Waters" Forests 2, no. 1: 243-260. https://doi.org/10.3390/f2010243