Spatial Distribution and Volume of Dead Wood in Unmanaged Caspian Beech (Fagus orientalis) Forests from Northern Iran


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
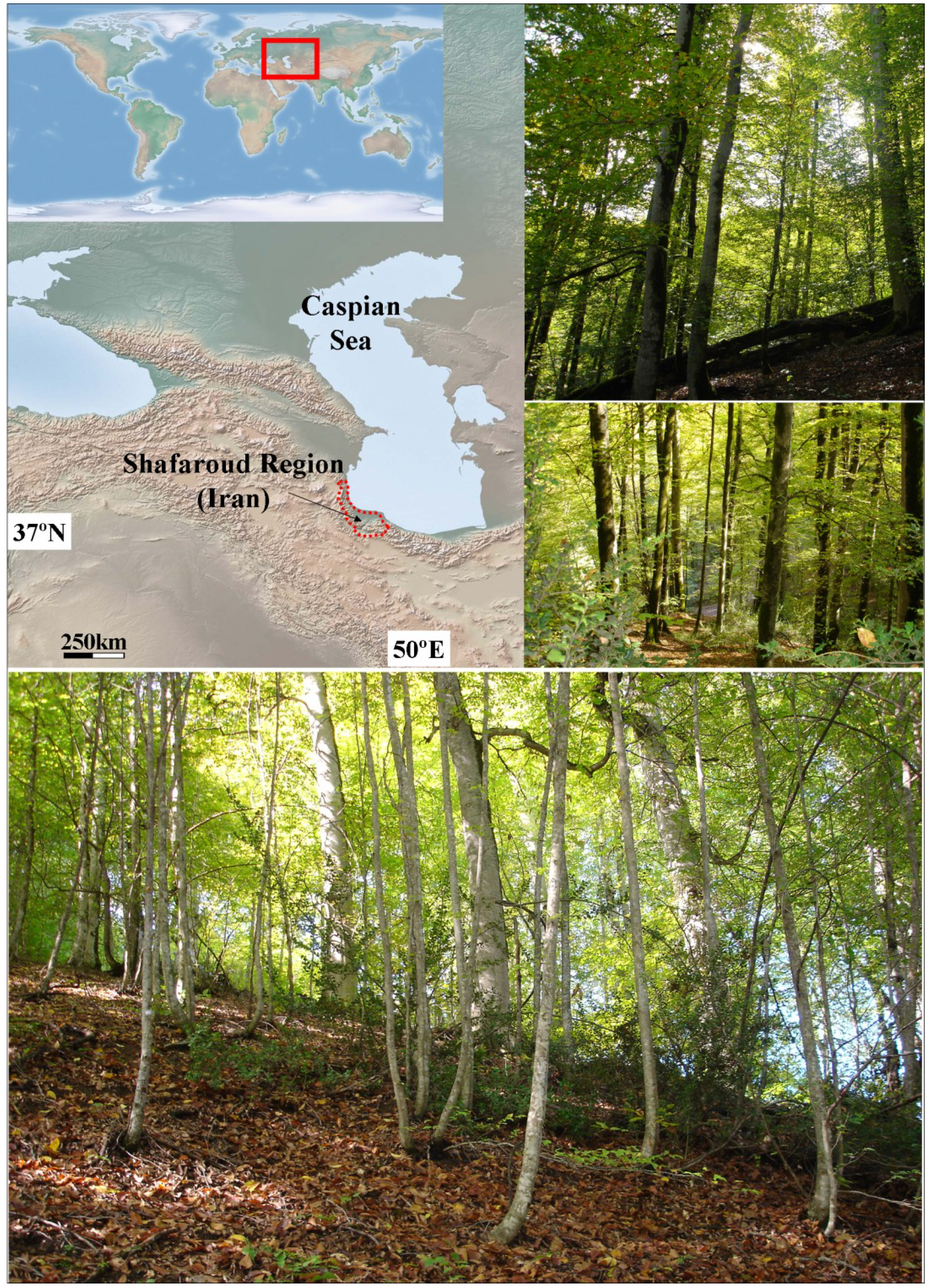
2.1. Study Site and Field Sampling


2.2. Point Pattern Analyses of Living and Dead Beech Trees

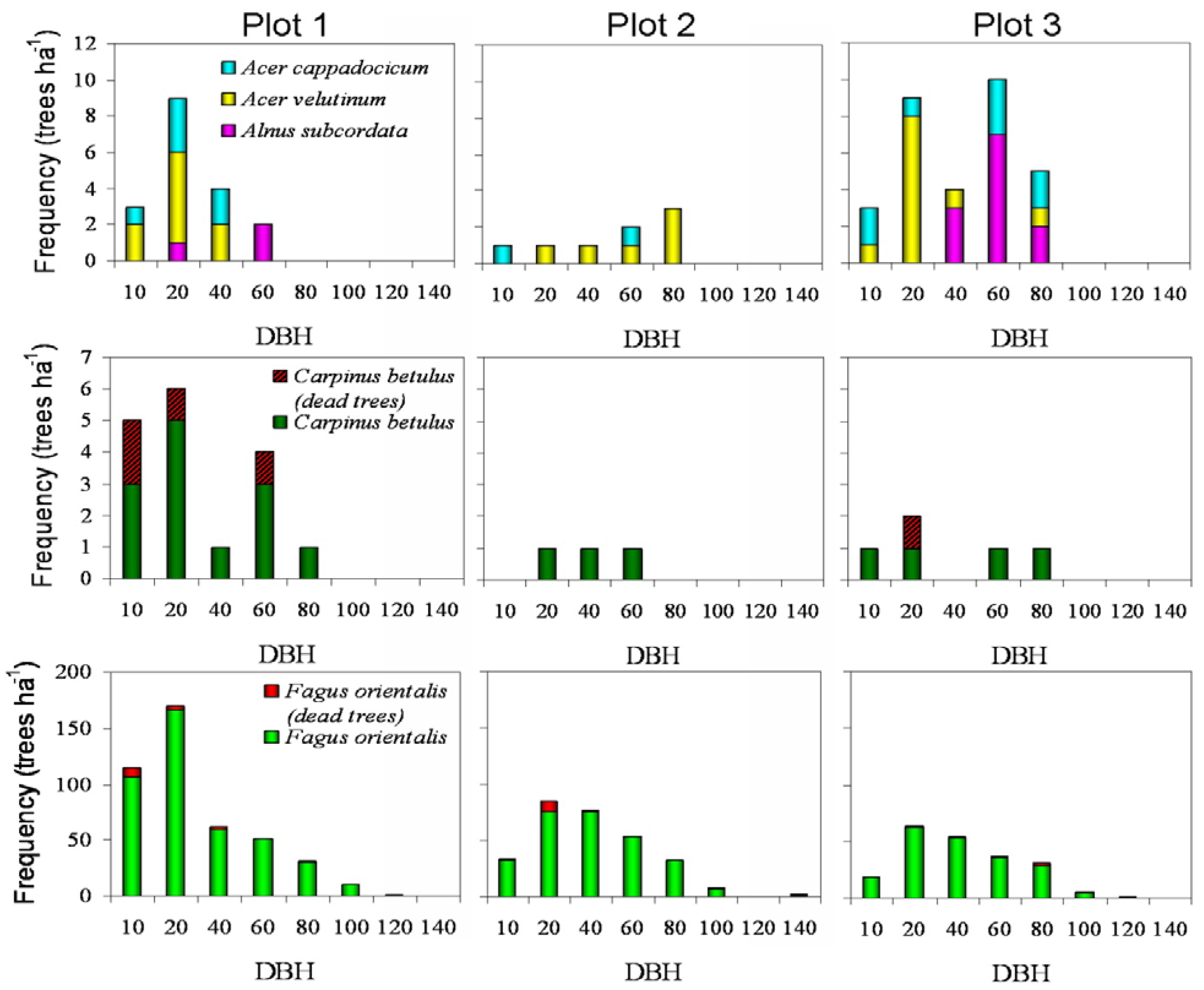
3. Results
3.1. Standing Dead Trees

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Living Trees | |||||||
| Plot | F. orientalis | A. velutinum | A. subcordata | C. betulus | A. cappadocicum | Total | |
| Tree Density | 1 | 424 | 9 | 3 | 13 | 6 | 455 |
| 2 | 277 | 6 | 0 | 3 | 2 | 288 | |
| (ind. ha−1) | |||||||
| 3 | 206 | 11 | 12 | 4 | 8 | 241 | |
| Basal Area | 1 | 36.5 | 0.2 | 0.4 | 1.2 | 0.1 | 38.4 |
| (m2 ha−1) | 2 | 34.7 | 1.4 | 0 | 0.2 | 0.1 | 36.5 |
| 3 | 27.5 | 0.6 | 2.5 | 0.5 | 1.2 | 32.3 | |
| Standing Dead Trees | |||||||
| Plot | F. orientalis | A. velutinum | A. subcordata | C. betulus | A. cappadocicum | Total | |
| Tree Density | 1 | 16 | 0 | 0 | 4 | 0 | 20 |
| 2 | 15 | 0 | 0 | 0 | 0 | 15 | |
| (ind. ha−1) | |||||||
| 3 | 6 | 0 | 0 | 1 | 0 | 7 | |
| Basal Area | 1 | 0.8 | 0 | 0 | 0.3 | 0 | 1.1 |
| 2 | 2.3 | 0 | 0 | 0 | 0 | 2.3 | |
| (m2 ha−1) | |||||||
| 3 | 1.8 | 0 | 0 | 0 | 0 | 1.8 | |
3.2. Fallen Dead Trees
| Tree Species | Number | Tree Diameter (cm) | Length (m) | Volume (m3) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| min | max | mean | min | max | mean | min | max | mean | |||
| Plot 1 | F. orientalis | 9 | 7 | 90 | 42 | 5 | 28 | 16.4 | 0.04 | 27.5 | 3.1 |
| A. velutinum | 1 | 18 | 18 | 18 | 12 | 12 | 12 | 0.31 | 0.31 | 0.31 | |
| Plot 2 | F. orientalis | 12 | 14 | 100 | 44.9 | 8 | 34 | 15.1 | 0.21 | 18.1 | 3.5 |
| Plot 3 | F. orientalis | 22 | 13 | 94 | 44.4 | 6 | 32 | 19.4 | 0.21 | 19.4 | 4.3 |
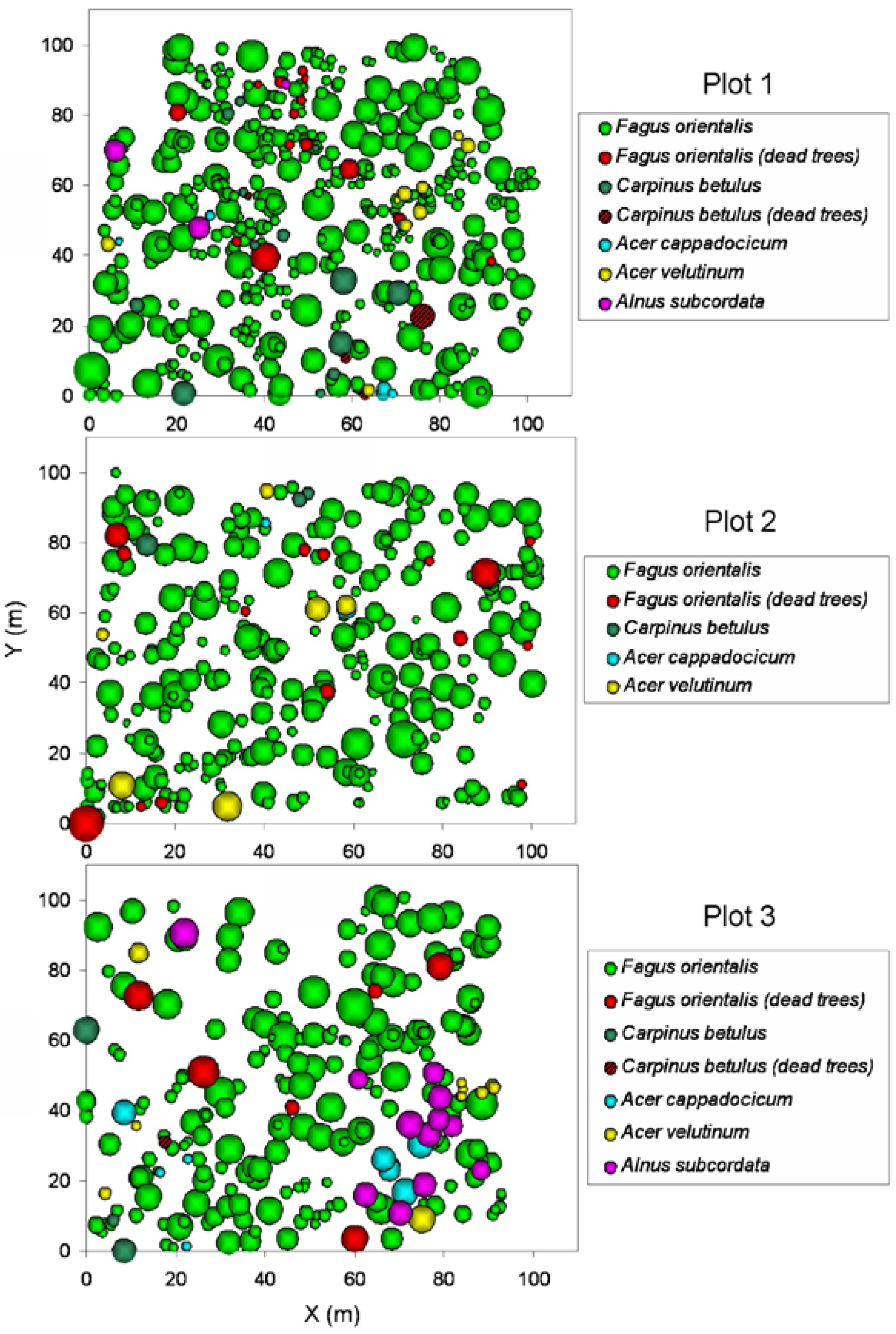
3.3. Spatial Analysis of Dead Trees


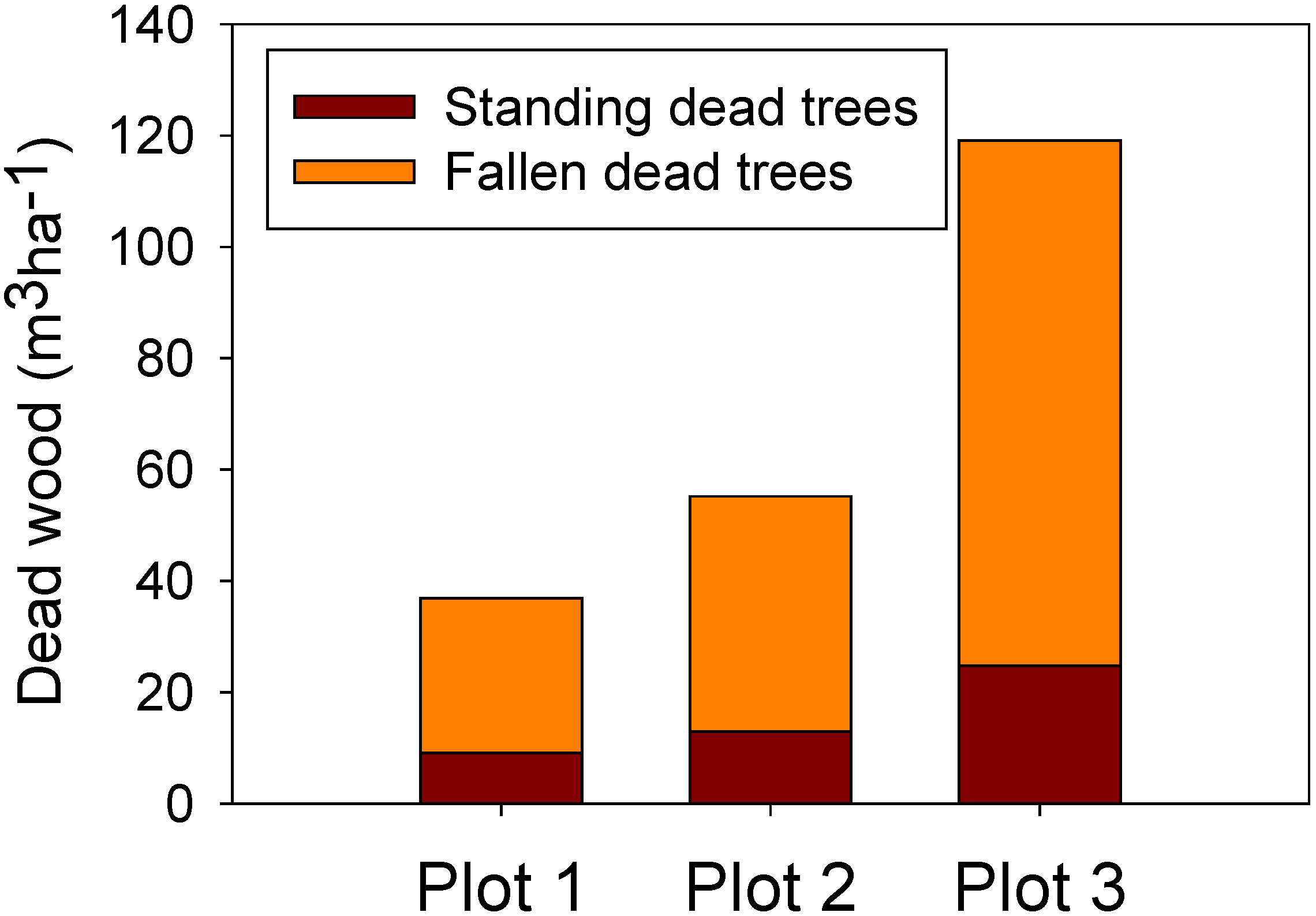
3.4. Volume of Dead Wood

| Plot | Living trees wood volume (m3 ha−1) | Standing dead wood volume (m3 ha−1) | Fallen dead wood volume (m3 ha−1) | Dead wood volume to alive wood volume ratio (%) |
|---|---|---|---|---|
| 1 | 490.6 | 9.1 | 27.8 | 7.5 |
| 2 | 486.0 | 12.9 | 42.3 | 11.4 |
| 3 | 417.0 | 24.7 | 94.4 | 29.0 |
| Rot class (%) | ||||
|---|---|---|---|---|
| I | II | III | IV | |
| Plot 1 | 20 | 10 | 30 | 40 |
| Plot 2 | 0 | 0 | 83 | 17 |
| Plot 3 | 4 | 41 | 23 | 32 |
4. Discussion
5. Conclusions
Conflicts of Interest
Acknowledgements
References
- Peterken, G.F. Natural Woodland: Ecology and Conservation in Northern Temperate Regions; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Gossner, M.M.; Lachat, T.; Brunet, J.; Isacsson, G.; Bouget, C.; Brustel, H.; Brandl, R.; Weisser, W.W.; Müller, J. Current near-to-nature forest management effects on functional trait composition of saproxylic beetles in Beech Forests. Conserv. Biol. 2013, 27, 605–614. [Google Scholar] [CrossRef]
- Standovár, T.; Kenderes, K. A review on natural stand dynamics in beech woods of east central Europe. Appl. Ecol. Env. Res. 2003, 1, 19–46. [Google Scholar]
- Linares, J.C.; Carreira, J.A.; Ochoa, V. Human impacts drive forest structure and diversity. Insights from Mediterranean mountain forest dominated by Abies pinsapo. Eur. J. For. Res. 2011, 130, 533–542. [Google Scholar] [CrossRef]
- Kirby, K.J.; Reid, C.M.; Thomas, R.C.; Goldsmith, F.B. Preliminary estimates of fallen dead wood and standing dead trees in managed and unmanaged forests in Britain. J. Appl. Ecol. 2007, 35, 148–155. [Google Scholar]
- Lassauce, A.; Paillet, Y.; Jactel, H.; Bouget, C. Deadwood as a surrogate for forest biodiversity: Meta-Analysis of correlations between deadwood volume and species richness of saproxylic organisms. Ecol. Indic. 2011, 11, 1027–1039. [Google Scholar] [CrossRef]
- Christensen, M.; Hahn, K.P.; Mountford, E.; Odor, M. Dead wood in European beech (Fagus sylvatica) forest reserves. For. Ecol. Manag. 2005, 210, 267–282. [Google Scholar] [CrossRef]
- Stokland, J.N.; Siitonen, J. Biodiversity in Dead Wood; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Heilmann-Clausen, J.; Christensen, M. Does size matter? On the importance of various dead wood fractions for fungal diversity in Danish beech forests. For. Ecol. Manag. 2004, 201, 105–117. [Google Scholar]
- Brunet, J.; Fritz, O.; Richnau, G. Biodiversity in European beech forests—A review with recommendations for sustainable forest management. Ecol. Bull. 2010, 53, 77–94. [Google Scholar]
- Diggle, P.J. Statistical Analysis of Point Patterns; Arnold: London, UK, 2003. [Google Scholar]
- Arevalo, J.R.; Fernandez-Palacios, J.M. Spatial patterns of trees and juveniles in a laurel forest of Tenerife, Canary Islands. Plant Ecol. 2003, 165, 1–10. [Google Scholar] [CrossRef]
- Tilman, D. Competition and biodiversity in spatially structured habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Szwagrzyk, J.; Szewczyk, J. Tree mortality and effects of release from competition in an old-growth Fagus-Abies-Picea stand. J. Veg. Sci. 2001, 12, 621–626. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; Wiley: New York, NY, USA, 1996. [Google Scholar]
- Lorimer, C.G.; White, A.S. Scale and frequency of natural disturbances in the Northeastern United States: Implications for early successional forest habitats and regional age distributions. For. Ecol. Manag. 2003, 185, 41–64. [Google Scholar] [CrossRef]
- Saniga, M.; Schutz, J.P. Relation of dead wood course within the development cycle of selected virgin forests in Slovakia. For. Sci. 2002, 48, 513–528. [Google Scholar]
- Mountford, E. Fallen and dead wood levels in the near-nature beech at La Tillaie Reserve, Fontainebleau, France. Oxf. For. Inst. For. 2002, 75, 203–208. [Google Scholar]
- Aakala, T.; Kuuluvainen, T.; Gauthier, S.; Grandpré, L. Standing dead trees and their decay-class dynamics in the north eastern boreal old-growth forests of Quebec. For. Ecol. Manag. 2008, 255, 410–420. [Google Scholar] [CrossRef]
- Parhizkar, P.; Sagheb-Talebi, K.; Mataji, A.; Namiranian, M. Influence of gap size and development stages on the silvicultural characteristics of oriental beech (Fagus orientalis Lipsky) regeneration. Caspian J. Env. Sci. 2011, 9, 55–65. [Google Scholar]
- Akhavan, R.; Sagheb-Talebi, Kh.; Zenner, E.K.; Safavimanesh, F. Spatial patterns in different forest development stages of an intact old-growth Oriental beech forest in the Caspian region of Iran. Eur. J. For. Res. 2012, 131, 1355–1366. [Google Scholar] [CrossRef]
- Sefidi, K.; Marvie-Mohadjer, M.R. Characteristics of coarse woody debris in successional stages of natural beech (Fagus orientalis) forests of Northern Iran. J. For. Sci. 2010, 56, 7–17. [Google Scholar]
- Zolfaghari, E.; Marvi Mohajer, M.R.; Namiranian, M. Impact of dead trees on natural regeneration in forest stands (Chelir District, Kheiroudkenar, Now-shahr). Iran. J. For. Poplar Res. 2007, 15, 234–240. [Google Scholar]
- Habashi, H. Study of Ecological and Silvicultural Importance of Dead Trees in Nour Forests. Master Thesis, University of Tarbiat Modarres, Tehran, Iran, 1998. [Google Scholar]
- Muller-Using, S.; Bartsch, N. Dynamics of Woody Debris in A Beech (Fagus sylvatica L.) Forest in Centeral Germany. In Proceeding of the 7th International Beech Symposium, Tehran, Iran, 10–20 May 2004; pp. 83–89.
- Nordén, B.; Ryberg, M.; Götmark, F.; Olausson, B. Relative importance of coarse and fine woody debris for the diversity of wood-inhabiting fungi in temperate broadleaf forests. Biol. Conserv. 2004, 117, 1–10. [Google Scholar] [CrossRef]
- Goreaud, F.; Pèlissier, R. Avoiding misinterpretation of biotic interactions with the intertype K12-function: Population independence vs. random labelling hypotheses. J. Veg. Sci. 2004, 14, 681–692. [Google Scholar]
- Stoyan, D.; Stoyan, H. Fractals, Random Shapes and Point Fields: Methods of Geometrical Statistics; Wiley: Chichester, UK, 1994. [Google Scholar]
- Wiegand, T.; Moloney, K.A. Ring, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- He, F.; Legendre, P.; LaFrankie, J.V. Distribution patterns of tree species in a Malaysian tropical rain forest. J. Veg. Sci. 1997, 8, 105–114. [Google Scholar] [CrossRef]
- Gray, L.; He, F. Spatial point-pattern analysis for detecting density-dependent competition in a boreal chronosequence of Alberta. For. Ecol. Manag. 2009, 259, 98–106. [Google Scholar] [CrossRef]
- He, F.; Duncan, R.P. Density-dependent effects on tree survival in old-growth Douglas fir forest. J. Ecol. 2000, 88, 676–688. [Google Scholar] [CrossRef]
- Little, L.R. Investigating competitive interactions from spatial patterns of tree in multi-space boreal forests: The random mortality hypothesis revisited. Can. J. Bot. 2002, 80, 93–100. [Google Scholar] [CrossRef]
- Getzin, S.; Dean, C.; He, F.; Trofymow, J.A.; Wiegand, K.; Wiegand, T. Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island. Ecography 2006, 29, 671–682. [Google Scholar] [CrossRef]
- Duncan, R.P. Competition and the coexistence of species in a mixed podocarp stand. J. Ecol. 1991, 79, 1073–1084. [Google Scholar] [CrossRef]
- Rozas, V.; Zas, R.; Solla, A. Spatial structure of deciduous forest stands with contrasting human influence in northwest Spain. Eur. J. For. Res. 2009, 128, 273–285. [Google Scholar] [CrossRef]
- Veblen, T.T.; Ashton, D.H.; Schlegel, F.M. Tree regeneration strategies in a lowland Nothofagus-dominated forest in southcentral Chile. J. Biogeo. 1979, 6, 329–340. [Google Scholar] [CrossRef]
- Salas, C.; LeMay, V.; Nunez, P.; Pacheco, P.; Espinosa, A. Spatial patterns in an old-growth Nothofagus oblique forest in south central Chile. For. Ecol. Manag. 2006, 231, 38–46. [Google Scholar] [CrossRef]
- Peters, R.; Poulson, T.L. Stem growth and canopy dynamics in a world-wide range of Fagus forests. J. Veg. Sci. 1994, 5, 421–432. [Google Scholar] [CrossRef]
- Canham, C.D. Growth and canopy architecture of shade tolerant trees. Response to canopy gaps. Ecology 1988, 69, 786–795. [Google Scholar] [CrossRef]
- Canham, C.D. Suppression and release during canopy recruitment in Fagus grandifolia. Bull. Torrey Bot. Club 1990, 117, 1–7. [Google Scholar] [CrossRef]
- Attiwill, P.M. The disturbance of forest ecosystems: The ecological basis for conservation management. For. Ecol. Manage. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Olano, J.M.; Laskurain, N.A.; Escudero, A.; de La Cruz, M. Why and where do adult trees die in a young secondary temperate forest? The role of neighbourhood. Ann. For. Sci. 2009, 66, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–276. [Google Scholar] [CrossRef]
- Beneke, C.; Manning, D. Coarse Woody Debris (CWD) in the Weberstedter Holz, A near Natural Beech Forest in Central Germany. Presented at Part of Deliverable 20 of the Nat-Man project (Nature based Management of Beech in Europe); Albert-Ludwigs University: Thuringia, Germany, 2003; p. 18. Available online: http://sl.life.ku.dk/English/research/forest_and_ecology/silviculture/natman/natural_beech_forests/~/media/Sl/Forskning_Research/Skov_oekologi/natman/reports/deliverable_20_part_beneke_christian.ashx (accessed on 6 May 2013).
- Castro-Gil, A. Evolution and structure of Artikutza, an 80-year old beech forest in Navarra (northern Spain). Munibe (Cienc. Nat. Nat. Zientziak) 2009, 57, 257–281. [Google Scholar]
- Motta, R.; Berretti, R.; Lingua, E.; Piussi, P. Coarse woody debris, forest structure and regeneration in the Valbona Forest Reserve, Paneveggio. Ital. Alps. For. Ecol. Manag. 2006, 235, 155–163. [Google Scholar] [CrossRef]
- Piovesan, G.; di Filippo, A.; Alessandrini, A.; Biondi, F.; Schirone, B. Structure, dynamics and dendroecology of an old-growth Fagus forest in the Apennines. J. Veg. Sci. 2005, 16, 13–28. [Google Scholar]
- Atici, E.; Colak, A.H.; Rotherhm, I.D. Coarse dead wood volume of managed oriental beech (Fagus orientalis Lipsky) stands in Turkey. For. Syst. 2008, 17, 216–227. [Google Scholar]
- Colak, A.; Tokcan, M.; Rotherham, I.; Atici, E. The amount of coarse dead wood and associated decay rates in forest reserves and managed forests, northwest Turkey. For. Syst. 2009, 18, 350–359. [Google Scholar]
- Wisdom, M.J.; Bate, L.J. Snag density varies with intensity of timber harvest and human access. For. Ecol. Manag. 2008, 255, 2085–2093. [Google Scholar] [CrossRef]
- Russell, M.B.; Kenefic, L.S.; Weiskittel, A.R.; Puhlick, J.J.; Brissette, J.C. Assessing and modeling standing deadwood attributes under alternative silvicultural regimes in the Acadian Forest region of Maine, USA. Can. J. For. Res. 2012, 42, 1873–1883. [Google Scholar] [CrossRef]
- Russell, M.B.; Woodall, C.W.; Fraver, S.; D’Amato, A.W. Estimates of downed woody debris decay class transitions for forests across the eastern United States. Ecol. Model. 2013, 251, 22–31. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Amanzadeh, B.; Sagheb-Talebi, K.; Foumani, B.S.; Fadaie, F.; Camarero, J.J.; Linares, J.C. Spatial Distribution and Volume of Dead Wood in Unmanaged Caspian Beech (Fagus orientalis) Forests from Northern Iran. Forests 2013, 4, 751-765. https://doi.org/10.3390/f4040751
Amanzadeh B, Sagheb-Talebi K, Foumani BS, Fadaie F, Camarero JJ, Linares JC. Spatial Distribution and Volume of Dead Wood in Unmanaged Caspian Beech (Fagus orientalis) Forests from Northern Iran. Forests. 2013; 4(4):751-765. https://doi.org/10.3390/f4040751
Chicago/Turabian StyleAmanzadeh, Beitollah, Khosro Sagheb-Talebi, Bahman Sotoudeh Foumani, Farhad Fadaie, Jesús Julio Camarero, and Juan Carlos Linares. 2013. "Spatial Distribution and Volume of Dead Wood in Unmanaged Caspian Beech (Fagus orientalis) Forests from Northern Iran" Forests 4, no. 4: 751-765. https://doi.org/10.3390/f4040751