Slow Lives in the Fast Landscape: Conservation and Management of Plethodontid Salamanders in Production Forests of the United States

Abstract
:1. Introduction
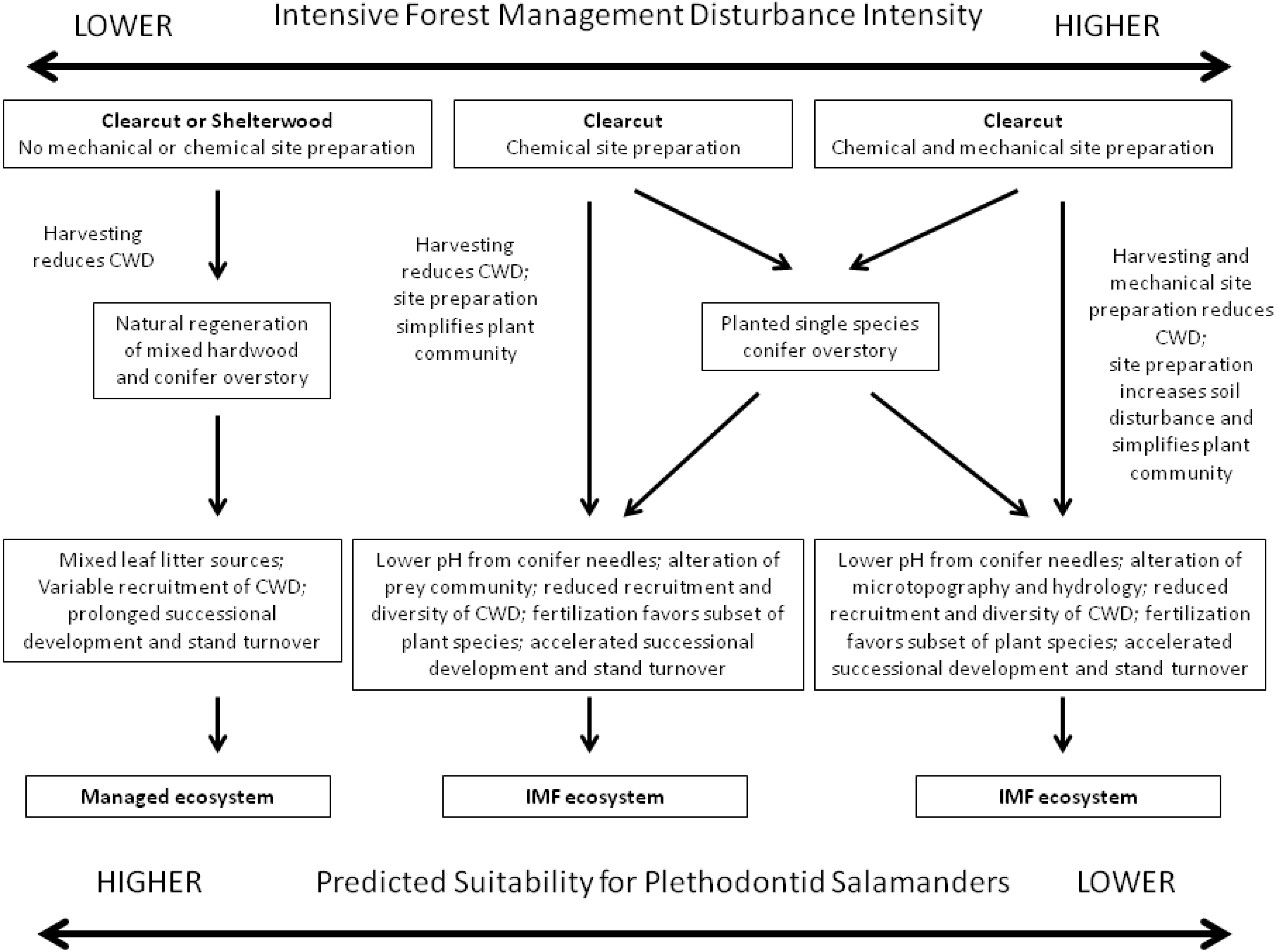
2. IMF Ecosystems in Time and Space

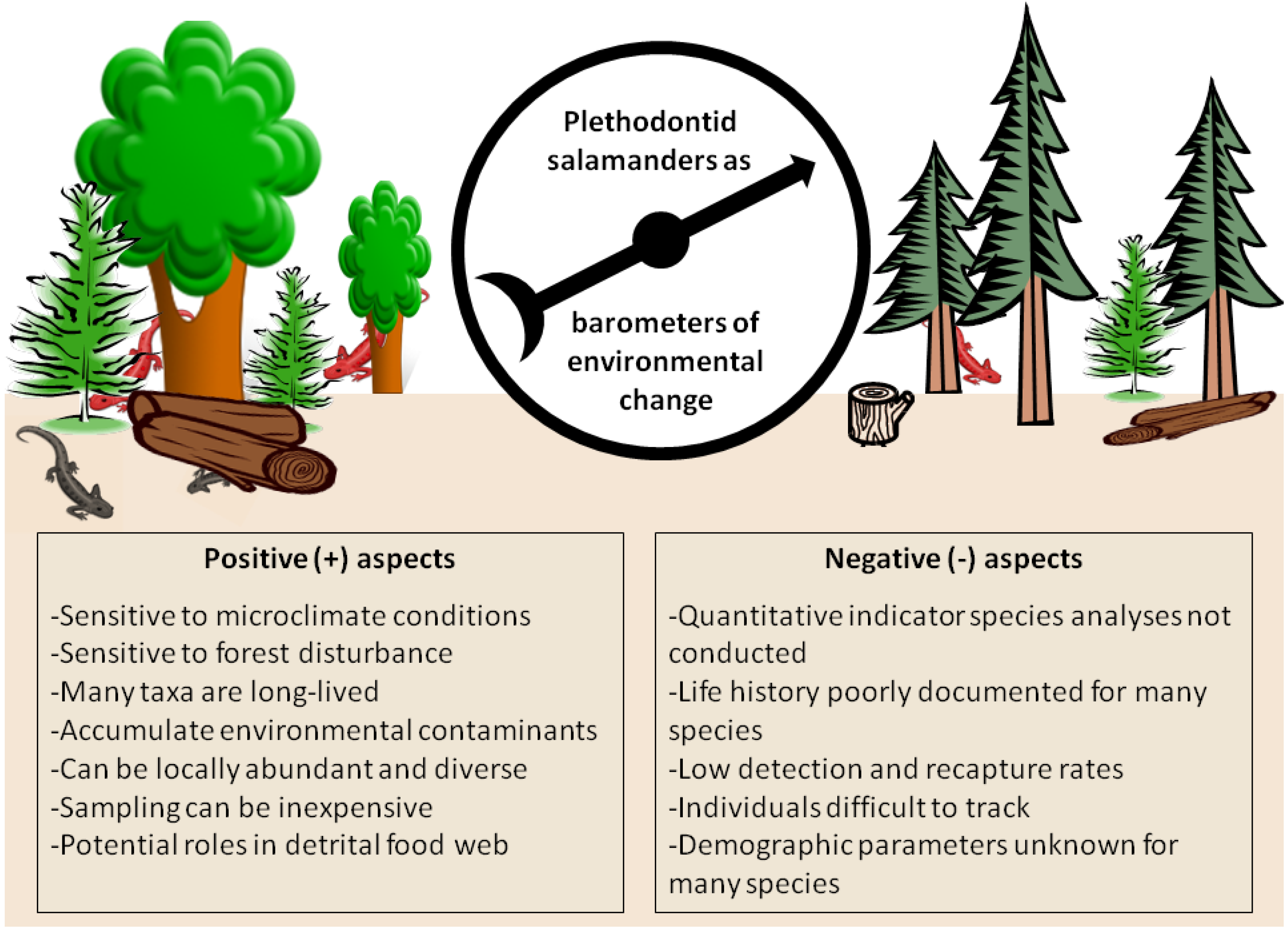
3. Salamanders as Barometers of Forest Condition

4. Case Studies of Salamanders in Managed Forests
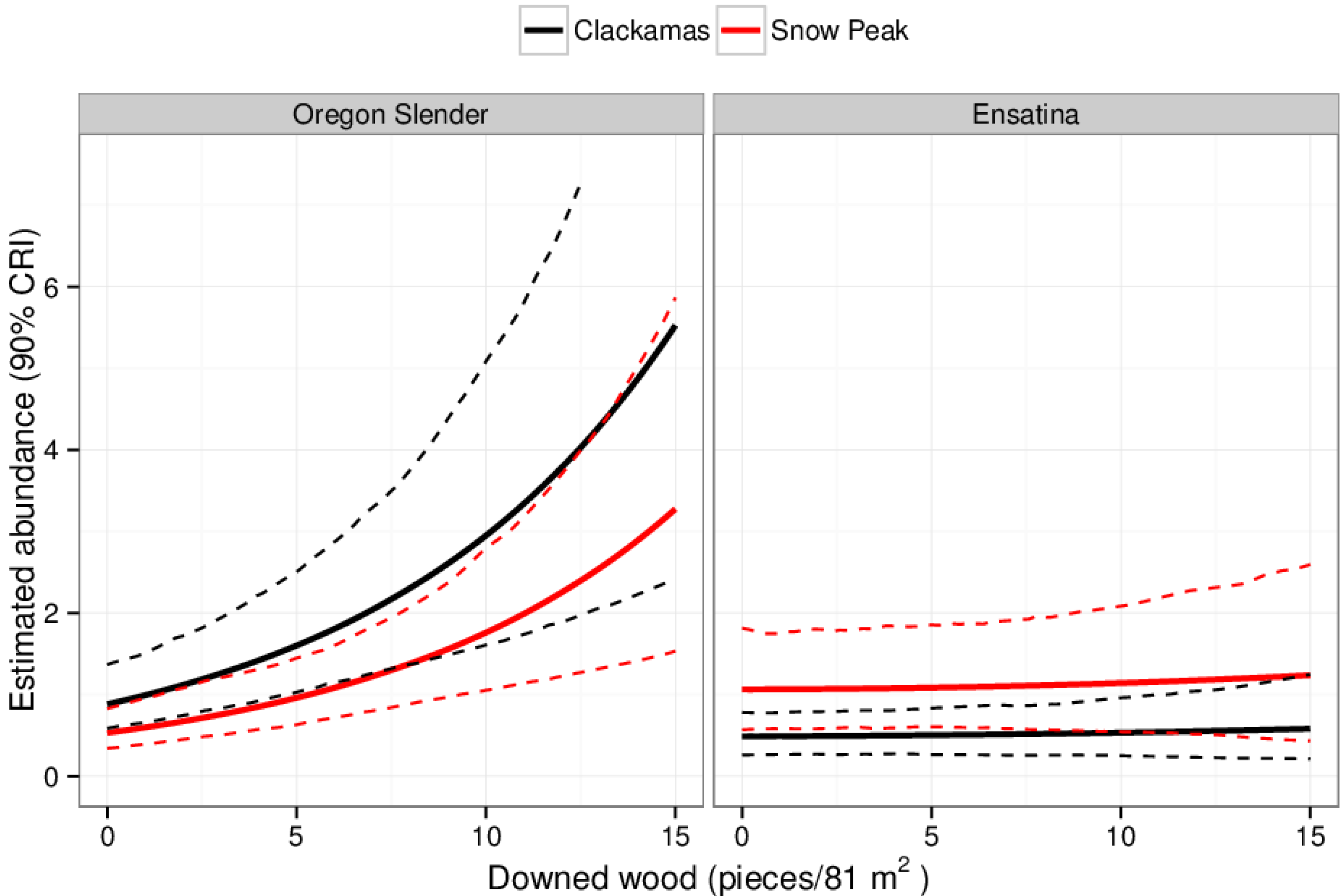
4.1. Douglas-Fir Regeneration and Plethodontid Salamanders in Cascades Forest


{kind=link}
{kind=link}
{kind=link}
| Species | Block | Year | Occupancy Estimate | 95% CI | Abundance Estimate | 95% CI |
|---|---|---|---|---|---|---|
| Oregon slender | Snow Peak | 2013 | 0.59 | 0.45, 0.75 | 0.91 | 0.60, 1.38 |
| Clackamas | 2013 | 0.76 | 0.63, 0.90 | 1.5 | 0.99, 2.32 | |
| Snow Peak | 2014 | 0.45 | 0.33, 0.63 | 0.62 | 0.41, 0.99 | |
| Clackamas | 2014 | 0.63 | 0.48, 0.80 | 1.03 | 0.66, 1.62 | |
| Ensatina | Snow Peak | 2013 | 0.61 | 0.41, 0.82 | 1.0 | 0.52, 1.7 |
| Clackamas | 2013 | 0.36 | 0.21, 0.53 | 0.46 | 0.24, 0.76 | |
| Snow Peak | 2014 | 0.66 | 0.47, 0.86 | 1.15 | 0.64, 1.95 | |
| Clackamas | 2014 | 0.25 | 0.34, 0.56 | 0.53 | 0.28, 0.83 |
4.2. Relationships of Plethodontid Salamanders with Woody Debris in Appalachian Forest
| Model | ∆AIC | Model Weight (wi) | R2 | β | SE | Direction |
|---|---|---|---|---|---|---|
| Overhead Canopy Conditions and Foraging (30-m2 scale) | 0 | 0.58 | 0.29 | |||
| Intercept | 2.724 | 1.039 | ||||
| Basal Area | 0.146 | 0.030 | + | |||
| Understory Density | 0.003 | 0.014 | + | |||
| Leaf Litter Depth | −0.410 | 0.147 | − | |||
| Percent Herbaceous Cover | 0.061 | 0.029 | + | |||
| Large-Scale Habitat Conditions (30-m2 scale) | 1.40 | 0.29 | 0.20 | |||
| Intercept | 1.435 | 0.536 | ||||
| Basal Area | 0.158 | 0.025 | + | |||
| Large-Scale Habitat Conditions (10-m2 scale) | 0.00 | 0.59 | 0.15 | |||
| Intercept | 0.421 | 0.246 | ||||
| Basal Area | 0.059 | 0.011 | + | |||
| Overhead Canopy Conditions (10-m2 scale) | 1.98 | 0.22 | 0.15 | |||
| Intercept | 0.371 | 0.359 | ||||
| Basal Area | 0.060 | 0.013 | + | |||
| Understory Density | 0.003 | 0.015 | + |
| Category | Species | CWD/FWD a | Leaf Litter | Tree/Sapling | Herbaceous | Rock | Bare Ground | Other | Number of Captures |
|---|---|---|---|---|---|---|---|---|---|
| All Treatments | Total Salamanders | 10.0 | 75.0 | 6.1 | 1.3 | 2.3 | 0.8 | 4.5 | 2480 |
| No Harvest Treatments | 7.5 | 77.0 | 6.3 | 1.7 | 2.3 | 0.9 | 4.4 | 1311 | |
| Harvest Treatments | 12.8 | 72.7 | 5.9 | 0.9 | 2.4 | 0.8 | 4.5 | 1169 | |
| All Treatments | Plethodon cinereus | 7.7 | 75.9 | 6.7 | 1.5 | 3.2 | 1.5 | 3.5 | 1107 |
| No Harvest Treatments | 6.3 | 76.9 | 6.3 | 1.9 | 3.7 | 1.8 | 3.0 | 567 | |
| Harvest Treatments | 9.1 | 74.8 | 7.0 | 1.1 | 2.6 | 1.3 | 4.1 | 540 | |
| All Treatments | Desmognathus spp. | 9.4 | 75.5 | 7.0 | 1.7 | 0.2 | 0.2 | 6.1 | 641 |
| No Harvest Treatments | 7.3 | 77.3 | 7.6 | 1.7 | 0.0 | 0.0 | 6.1 | 423 | |
| Harvest Treatments | 13.3 | 72.0 | 6.0 | 1.8 | 0.5 | 0.5 | 6.0 | 218 | |
| All Treatments | P. glutinosus complex | 16.8 | 69.3 | 3.2 | 0.4 | 4.1 | 0.6 | 5.4 | 463 |
| No Harvest Treatments | 13.2 | 69.5 | 3.6 | 0.6 | 4.2 | 1.2 | 7.8 | 167 | |
| Harvest Treatments | 13.3 | 72.0 | 6.0 | 1.8 | 0.5 | 0.5 | 6.0 | 218 | |
| All Treatments | P. richmondi | 5.2 | 86.0 | 4.1 | 1.6 | 0.0 | 0.0 | 3.1 | 193 |
| No Harvest Treatments | 5.4 | 89.3 | 0.9 | 2.7 | 0.0 | 0.0 | 1.8 | 112 | |
| Harvest Treatments | 4.9 | 81.5 | 8.6 | 0.0 | 0.0 | 0.0 | 4.9 | 81 | |
| All Treatments | P. wehrlei | 22.2 | 51.1 | 17.8 | 0.0 | 6.7 | 0.0 | 2.2 | 45 |
| No Harvest Treatments | 7.4 | 59.3 | 25.9 | 0.0 | 7.4 | 0.0 | 0.0 | 27 | |
| Harvest Treatments | 44.4 | 38.9 | 5.6 | 0.0 | 5.6 | 0.0 | 5.6 | 18 |
5. Conclusions
5.1. Beyond CWD
5.2. Opportunities for Future Research
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pan, Y.; Birdsey, R.A.; Phillips, O.L.; Jackson, R.B. The structure, distribution, and biomass of the world’s forests. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 593–622. [Google Scholar]
- State of the World’s Forests 2012; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012.
- Berlik, M.M.; Kittredge, D.B.; Foster, D.R. The illusion of preservation: A global environmental argument for the local production of natural resources. J. Biogeogr. 2002, 29, 1557–1568. [Google Scholar] [CrossRef]
- Abernethy, K.A.; Coad, L.; Taylor, G.; Lee, M.E.; Maisels, F. Extent and ecological consequences of hunting in Central African rainforests in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef]
- Berger, A.L.; Palik, B.; D'Amato, A.W.; Fraver, S.; Bradford, J.B.; Nislow, K.; King, D.; Brooks, R.T. Ecological impacts of energy-wood harvests: Lessons from whole-tree harvesting and natural disturbance. J. For. 2013, 111, 139–153. [Google Scholar]
- Stein, S.M.; McRoberts, R.E.; Alig, R.J.; Nelson, M.D.; Theobald, D.M.; Eley, M.; Dechter, M.; Carr, M.A. Forests on the Edge: Housing Development on America’s Private Forests; Gen. Tech. Rep. PNW-636; Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2009; p. 15. [Google Scholar]
- Smail, R.A.; Lewis, D.J. Forest-Land Conversion, Ecosystem Services, and Economic Issues for Policy: A Review; PNW-GTR-797; Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2009; p. 40. [Google Scholar]
- Forests Sourcebook: Practical Guidance for Sustaining Forests in Development Cooperation; World Bank: Washington, DC, USA, 2008.
- Constanza, R.; de Groot, R.; Sutton, P.; van der Ploeg, S.; Anderson, S.J.; Kubidzewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Binkley, C.S.; Aronow, M.E.; Washburn, C.L.; New, D. Global perspectives on intensively managed plantations: Implications for the Pacific Northwest. J. For. 2005, 103, 61–64. [Google Scholar]
- Paquette, A.; Messier, C. The role of plantations in managing the world’s forests in the Anthropocene. Front. Ecol. Environ. 2010, 8, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Sedjo, R.A. The potential of high-yield plantation forestry for meeting timber needs. New For. 1999, 17, 339–359. [Google Scholar] [CrossRef]
- Carnus, J.-M.; Parrotta, J.; Brockerhoff, E.; Arbez, M.; Jactel, H.; Kremer, A.; Lamb, D.; O'Hara, K.; Walters, B. Planted forests and biodiversity. J. For. 2006, 104, 65–77. [Google Scholar]
- Hayes, J.P.; Schoenholtz, S.H.; Hartley, M.J.; Murphy, G.; Powers, R.F.; Berg, D.; Radosevich, S.R. Environmental consequences of intensively managed forest plantations in the Pacific Northwest. J. For. 2005, 103, 83–87. [Google Scholar]
- Fox, T.R.; Jokela, E.J.; Allen, H.L. The development of pine plantation silviculture in the southern United States. J. For. 2007, 105, 337–347. [Google Scholar]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; Della Sala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The forgotten stage of forest succession: Early-successional ecosystems on forest sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef]
- Moore, R.D.; Richardson, J.S. Natural disturbance and forest management in riparian zones: Comparison of effects at reach, catchment, and landscape scales. Freshw. Sci. 2012, 31, 239–247. [Google Scholar] [CrossRef]
- Linden, D.W.; Roloff, G.J.; Kroll, A.J. Conserving avian richness through structure retention in managed forests of the Pacific Northwest. For. Ecol. Manag. 2012, 284, 174–184. [Google Scholar] [CrossRef]
- Welsh, H.H., Jr.; Droege, S. A case for using plethodontid salamanders for monitoring biodiversity and ecosystem integrity of North American forests. Conserv. Biol. 2001, 15, 558–569. [Google Scholar] [CrossRef]
- Petranka, J.W. Salamanders of the United States and Canada; Smithsonian Institution: Washington, DC, USA, 1998. [Google Scholar]
- Pough, F.H. The advantages of ecothermy for tetrapods. Am. Nat. 1980, 115, 92–112. [Google Scholar] [CrossRef]
- Burton, T.M.; Likens, G.E. Energy flow and nutrient cycling in salamander populations in the Hubbard Brook Experimental Forest, New Hampshire. Ecology 1975, 56, 1068–1080. [Google Scholar] [CrossRef]
- Hocking, D.J.; Babbitt, K.J. Amphibian contributions to ecosystem services. Herpetol. Conserv. Biol. 2014, 9, 1–17. [Google Scholar]
- Davic, R.D.; Welsh, H.H., Jr. On the ecological role of salamanders. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 405–434. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Todd, B.D.; Blomquist, S.M.; Calhoun, A.J.K.; Gibbons, J.W.; Gibbs, J.P.; Graeter, G.J.; Harper, E.B.; Hocking, D.J.; Hunter, M.L.J.; Patrick, D.A.; Rittenhouse, T.A.G.; Rothermel, B.B. Effects of timber harvest on amphibian populations: Understanding mechanisms from forest experiments. BioScience 2009, 50, 853–862. [Google Scholar] [CrossRef]
- Welsh, H.H.; Hodgson, G.R. Woodland salamanders as metrics of forest ecosystem recovery: A case study from California’s redwoods. Ecosphere. 2013, 4, p. art59. Available online: http://dx.doi.org/10.1890/ES12-00400.1. [CrossRef]
- Tilghman, J.M.; Ramee, S.W.; Marsh, D.M. Meta-analysis of the effects of canopy removal on terrestrial salamander populations in North America. Biol. Conserv. 2012, 152, 1–9. [Google Scholar] [CrossRef]
- Demaynadier, P.G.; Hunter, M.L.J. The relationship between forest management and amphibian ecology: A review of the North American literature. Environ. Rev. 1995, 3, 230–261. [Google Scholar] [CrossRef]
- Homyack, J.A.; Haas, C.A. Long-term effects of experimental forest harvesting on abundance and reproductive demography of terrestrial salamanders. Biol. Conserv. 2009, 142, 110–121. [Google Scholar] [CrossRef]
- Destefano, S. Regional and national issues for forest wildlife research and management. For. Sci. 2002, 48, 181–189. [Google Scholar]
- Kerby, J.L.; Richards-Hrdlicka, K.L.; Storfer, A.; Skelly, D.L. An examination of amphibian sensitivity to environmental contaminants: Are amphibians poor canaries? Ecol. Lett. 2010, 13, 60–67. [Google Scholar]
- Niemi, G.J.; McDonald, M.E. Application of ecological indicators. Annu. Rev. Evol. Ecol. Syst. 2004, 35, 89–111. [Google Scholar] [CrossRef]
- Bergoffen, W.W. 100 Years of Federal Forestry; Agriculture Information Bulletin No. 402; USDA Forest Service: Washington, DC, USA, 1976; p. 199. [Google Scholar]
- McCleery, D.W. American Forests: A History of Resiliency and Recovery; USDA Forest Service and Forest History Society: Durham, NC, USA, 1992. [Google Scholar]
- Forest Ecosystem Management: An Ecological, Economic and Social Assessment; Report of the Forest Ecosystem Management Assessment Team, USA; Department of Agriculture Forest Service: Portland, OR, USA, 1993.
- Moore, S.E.; Cubbage, F.; Eicheldinger, C. Impacts of Forest Stewardship Council (FSC) and Sustainable Forestry Initiative (SFI) forest certification in North America. J. For. 2012, 110, 79–88. [Google Scholar]
- Zhang, D.; Butler, B.J.; Nagubadi, R.V. Institutional timberland ownership in the US South: magnitude, location, dynamics, and management. J. For. 2012, 110, 355–361. [Google Scholar]
- Wear, N.D.; Greis, J.G. The Southern Forests Futures Project; Wear, D.N., Greis, J.G., Eds.; USDA-Forest Service, Southern Research Station: Asheville, NC, USA, 2013; p. 552. [Google Scholar]
- Atwood, C.; Fox, TR.; Loftis, D.L. Effects of alternative silviculture on stump sprouting in the southern Appalachians. For. Ecol. Manag. 2009, 257, 1305–1313. [Google Scholar] [CrossRef]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practive of Silviculture: Applied Forest Ecology, 9th ed.; John Wiley & Sons: New York, NY, USA, 1997. [Google Scholar]
- Enge, M.K.; Marion, W. Effects of clearcutting and site preparation on herpetofauna of a north Florida flatwoods. Conserv. Biol. 1986, 14, 177–192. [Google Scholar]
- Ramovs, B.V.; Roberts, M.R. Understory vegetation and environment responses to tillage, forest harvesting, and conifer plantation development. Ecol. Appl. 2003, 13, 1682–1700. [Google Scholar] [CrossRef]
- Aust, M.W.; Blinn, C.R. Forestry best management practices for timber harvesting and site preparation in the eastern United States: An overview of water quality and productivity research during the past 20 years (1982–2002). Water Air Soil Pollut. 2004, 4, 5–36. [Google Scholar] [CrossRef]
- Forest Practices Illustrated; Washington State Department of Natural Resources, Forest Practices Division: Olympia, WA, USA, 2007; p. 152.
- Requirements for the SFI 2010–2014 Program; Sustainable Forestry Initiative: Washington, DC, USA, 2010; p. 123.
- FSC Principles and Criteria for Forest Stewardship; Forest Stewardship Council: Bonn, Germany, 2012; p. 37.
- Bunnell, F.L.; Boyland, M.; Wind, E. How should we spatially distribute dead and dying wood? In Proceedings of the Symposium on the Ecology and Management of Dead Wood in Western Forests; Laudenslayer, W.F., Jr., Shea, P.J., Valentine, B.F., Weatherspoon, C.P., Lisle, T.E., Eds.; Gen. Tech. Rep. PSW-GTR-181; Pacific Southwest Research Station; Forest Service. U.S. Department of Agriculture: Albany, CA, USA, 2002; pp. 739–752. [Google Scholar]
- Olson, D.H.; Kluber, M.R. Plethodontid salamander distributions in managed forest headwaters in western Oregon, USA. Herpetol. Conserv. Biol. 2014, 9, 76–96. [Google Scholar]
- Ohmann, J.L.; Gregory, M.J.; Spies, T.A. Influence of environment, disturbance, and ownership on forest vegetation of coastal Oregon. Ecol. Appl. 2007, 17, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Underwood, E.C.; Viers, J.H.; Quinn, J.F.; North, M. Using topography to meet wildlife and fuels treatment objectives in fire-suppressed landscapes. Environ. Manag. 2010, 46, 809–819. [Google Scholar] [CrossRef]
- Suming, J.; Sader, S.A. Effects of forest ownership and change on forest harvest rates, types and trends in northern Maine. For. Ecol. Manag. 2006, 228, 177–186. [Google Scholar] [CrossRef]
- Miller, D.A.; Wigley, T.B.; Miller, K.V. Managed forests and conservation of terrestrial biodiversity in the southern United States. J. For. 2009, 107, 197–203. [Google Scholar]
- Perera, A.J.; Buse, L.J.; Weber, M.G. Emulating Natural Forest Landscape Disturbances; Columbia University Press: New York, NY, USA, 2004. [Google Scholar]
- Fischer, J.; Abson, D.J.; Butsic, V.; Chappell, M.J.; Ekroos, J.; Hanspach, J.; Kuemmerle, T.; Smith, H.G.; Wehrden, H.V. Land sparing versus land sharing: Moving forward. Conserv. Lett. 2013, 7, 149–157. [Google Scholar] [CrossRef]
- Phalan, B.; Onial, M.; Balmford, A.; Green, A.W. Reconciling food production and biodiversity conservation: Land sharing and land sparing compared. Science 2011, 333, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Drever, C.R.; Peterson, G.; Messier, C.; Bergeron, Y.; Flannigan, M. Can forest management based on natural disturbance maintain ecological resilience? Can. J. For. Res. 2006, 36, 2285–2299. [Google Scholar]
- Tittler, R.; Messier, C.; Fall, A. Concentrating anthropogenic disturbance to balance ecological and economic values: Applications to forest management. Ecol. Appl. 2012, 22, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, D.; Sait, S.M.; Kunin, W.E.; Benton, T.G. Food production vs. biodiversity: Comparing organic and conventional agriculture. J. Appl. Ecol. 2013, 50, 355–364. [Google Scholar] [CrossRef]
- Swanson, F.J.; Franklin, J.F. New forestry principles from ecosystem analysis of Pacific Northwest forests. Ecol. Appl. 1992, 2, 262–274. [Google Scholar]
- Green, R.E.; Cornell, S.J.; Scharlemann, J.P.W.; Balmford, A. Farming and the fate of wild nature. Science 2005, 307, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Lindenmayer, D. Future directions for biodiversity conservation in managed forests: Indicator species, impact studies and monitoring programs. For. Ecol. Manag. 1999, 115, 277–287. [Google Scholar] [CrossRef]
- Kremen, C. Assessing the indicator properties of species assemblages for natural areas monitoring. Ecol. Appl. 1992, 2, 203–217. [Google Scholar]
- Sayler, A. The reproductive ecology of the red-backed salamander, Plethodon cinereus, in Maryland. Copeia 1966, 1966, 183–193. [Google Scholar]
- Angleberger, M.A.P.; Chinnici, J.P. Dimorphism in the red-backed salamander Plethodon cinereus (Green) at Mountain Lake, Virginia. Va. J. Sci. 1975, 26, 153–158. [Google Scholar]
- Ruben, J.A.; Boucot, A.J. The origin of the lungless salamanders (Amphibia: Plethodontidae). Am. Nat. 1989, 134, 161–169. [Google Scholar] [CrossRef]
- Homyack, J.A.; Haas, C.A.; Hopkins, W.A. Influence of temperature and body mass on standard metabolic rate of eastern red-backed salamanders (Plethodon cinereus). J. Therm. Biol. 2010, 35, 143–146. [Google Scholar] [CrossRef]
- Stefanski, M.; Gatten, R.E.; Pough, F.H. Activity metabolism of salamanders: Tolerance to dehydration. J. Herpetol. 1989, 23, 45–50. [Google Scholar] [CrossRef]
- Gatten, R.E.; Miller, K.; Full, R.J. Energetics at Rest and During Locomotion, in Environmental Physiology of the Amphibians; Feder, M.E., Burggren, W.W., Eds.; University of Press: Chicago, IL, USA, 1992; pp. 314–377. [Google Scholar]
- Homyack, J.A.; Haas, C.A.; Hopkins, W.A. Effects of experimental forest harvesting on energetics of terrestrial salamanders. J. Wildl. Manag. 2011, 7, 1267–1278. [Google Scholar]
- Grant, E.H.C.; Nichols, J.D.; Lowe, W.H.; Fagan, W.F. Use of multiple dispersal pathways facilitates amphibian persistence in stream networks. Proc. Natl. Acad. Sci. USA 2010, 107, 6936–6940. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.H.; Leirness, J.B.; Cunningham, P.B.; Steel, E.A. Riparian buffers and forest thinning: Effects on headwater vertebrates 10 years after thinning. For. Ecol. Manag. 2014, 321, 81–93. [Google Scholar] [CrossRef]
- Ash, A.N. Disappearance and return of plethodontid salamanders to clearcut plots in the southern Blue Ridge Mountains. Conserv. Biol. 1997, 11, 983–989. [Google Scholar] [CrossRef]
- Petranka, J.W.; Eldridge, M.E.; Haley, K.E. Effects of timber harvesting on southern Appalachian salamanders. Conserv. Biol. 1993, 7, 363–370. [Google Scholar] [CrossRef]
- Wyman, R.L.; Hawksley-Lescault, D.S. Soil acidity affects distribution, behavior, and physiology and the salamander, Plethodon cinereus. Ecology 1987, 68, 1819–1827. [Google Scholar] [CrossRef]
- Burke, J.N.; Bergeron, C.M.; Todd, B.D.; Hopkins, W.A. Effects of mercury on behavior and performance of northern two-lined salamanders (Eurycea bislineata). Environ. Pollut. 2010, 158, 3546–3551. [Google Scholar] [CrossRef] [PubMed]
- Bank, M.S.; Loftin, C.S.; Jung, R.E. Mercury bioaccumulation in northern two-lined salamanders from streams in the northeastern United States. Ecotoxicology 2005, 14, 181–191. [Google Scholar] [CrossRef]
- Bergeron, C.M.; Bodinof, C.M.; Unrine, J.M.; Hopkins, W.A. Bioaccumulation and maternal transfer of mercury and selenium in amphibians. Environ. Toxicol. Chem. 2010, 29, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Corn, P.S.; Bury, R.B. Sampling Methods for Terrestrial Amphibians and Reptiles; United States Department of Agriculture; Forest Service; Pacific Southwest Research Station: Portland, OR, USA, 1990. [Google Scholar]
- Pearce, J.; Venier, L. Are salamanders good bioindicators of sustainable forest management in boreal forests? Can. J. For. Res. 2009, 39, 169–179. [Google Scholar]
- Walton, B.M.; Steckler, S. Contrasting effects of salamanders on forest-floor macro- and mesofauna in laboratory microcosms. Pedobiologia 2005, 49, 51–60. [Google Scholar] [CrossRef]
- Best, M.L.; Welsh, H.H. The trophic role of a forest salamander: Impacts on invertebrates, leaf litter retention, and the humification process. Ecosphere 2014, 5, 16. [Google Scholar] [CrossRef]
- Wyman, R.L. Experimental assessment of salamanders as predators of detrital food webs: Effects on invertebrates, decomposition, and the carbon cycle. Biodivers. Conserv. 1998, 7, 641–650. [Google Scholar] [CrossRef]
- Hocking, D.J.; Babbitt, K.J. Effects of red-backed salamanders on ecosystem functions. PLoS One 2014, 9, e86854. [Google Scholar] [CrossRef] [PubMed]
- Homyack, J.A.; Sucre, E.B.; Haas, C.A.; Fox, T.R. Does Plethodon cinereus affect leaf litter decomposition and invertebrate abundances in mixed oak forest? J. Herpetol. 2010, 44, 447–456. [Google Scholar] [CrossRef]
- Walton, B.M. Salamanders in forest-floor food webs: Environmental heterogeneity affects the strength of top-down effects. Pedobiologia 2005, 49, 381–393. [Google Scholar] [CrossRef]
- Landres, P.B.; Verner, J.; Thomas, J.W. Ecological uses of vertebrate indicator species: A critique. Conserv. Biol. 1988, 2, 316–328. [Google Scholar] [CrossRef]
- Andelman, S.J.; Fagan, W.F. Umbrellas and flagships: Efficient conservation surrogates or expensive mistakes? Proc. Natl. Acad. Sci. USA 2000, 97, 5954–5959. [Google Scholar] [CrossRef]
- Murtaugh, P.A. The statistical evaluation of ecological indicators. Ecol. Appl. 1996, 6, 132–139. [Google Scholar] [CrossRef]
- Carignan, V.; Villard, M.-A. Selecting indicator species to monitor ecological integrity. Environ. Monit. Assess. 2002, 78, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Belote, R.T.; Jones, R.H.; Wieboldt, T. Compositional stability and diversity of vascular plant communities following logging disturbance in Appalachian forests. Ecology 2012, 22, 502–516. [Google Scholar]
- Homyack, J.; Weyerhaeuser Company, Timberlands Technology, Vanceboro, NC, USA. Unpublished work. 2014.
- Kroll, A.J.; Runge, J.P.; MacCracken, J.G. Unreliable amphibian population metrics obfuscate more than they reveal. Biol. Conserv. 2009, 142, 2802–2806. [Google Scholar] [CrossRef]
- Bailey, L.L.; Simons, T.R.; Pollock, K.H. Estimating site occupancy and species detection probability parameters for terrestrial salamanders. Ecol. Appl. 2004, 14, 692–702. [Google Scholar] [CrossRef]
- Mazerolle, M.J.; Bailey, L.L.; Kendall, W.L.; Royale, J.A.; Converse, S.J.; Nichols, J.D. Making great leaps forward: Accounting for detectability in herpetological field studies. J. Herpetol. 2007, 41, 672–689. [Google Scholar] [CrossRef]
- Otto, C.R.V.; Roloff, G.J.; Thames, R.E. Comparing population patterns to processes: Abundance and survival of a forest salamander following habitat degradation. PLoS One 2014, 9, e93859. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.R.V.; Kroll, A.J.; McKenny, H.C. Amphibian response to downed wood retention in managed forests: A prospectus for future biomass harvest in North America. For. Ecol. Manag. 2013, 304, 275–285. [Google Scholar] [CrossRef]
- Harpole, D.N.; Haas, C.A. Effects of seven silvicultural treatments on terrestrial salamanders. For. Ecol. Manag. 1999, 114, 349–356. [Google Scholar] [CrossRef]
- Brin, A.; Meredieu, C.; Piou, D.; Brustel, H.; Jactel, H. Changes in quantitative patterns of dead wood in maritime pine plantations over time. For. Ecol. Manag. 2008, 256, 913–921. [Google Scholar] [CrossRef]
- Homyack, J.A.; Paxton, B.J.; Wilson, M.D.; Watts, B.D.; Miller, D.A. Snags and cavity-nesting birds within intensively managed pine stands in eastern North Carolina, USA. South. J. Appl. For. 2011, 35, 148–154. [Google Scholar]
- Hartley, M.J. Rationale and methods for conserving biodiversity in plantation forests. For. Ecol. Manag. 2002, 155, 81–95. [Google Scholar] [CrossRef]
- Forest Biomass Retention and Harvesting Guidelines for the Southeast; Forest Guild Southeast Working Group: Santa Fe, NM, USA, 2012.
- Pavlacky, D.C., Jr.; Blakesley, J.A.; White, G.C.; Hanni, D.J.; Lukacs, P.M. Hierarchical multi-scale occupancy estimation for monitoring wildlife populations. J. Wildl. Manag. 2012, 76, 154–162. [Google Scholar] [CrossRef]
- Royle, J.A.; Nichols, J.D. Estimating abundance from repeated presence-absence data or point counts. Ecology 2003, 84, 777–790. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence; Elsevier: San Diego, CA, USA, 2006. [Google Scholar]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Rubin, D.B. Bayesian Data Analysis, 2nd ed.; Chapman & Hall/CRC: Boca Raton, FL, USA, 2004. [Google Scholar]
- Nichols, J.D.; Bailey, L.L.; O’Connell, A.F.; Talancy, N.W., Jr.; Grant, E.H.C.; Gilbert, A.T.; Annand, E.M.; Husband, T.P.; Hines, J.E. Multi-scale occupancy estimation and modelling using multiple detection methods. J. Appl. Ecol. 2008, 45, 1321–1329. [Google Scholar] [CrossRef]
- Jones, L.L.C.; Leonard, W.P. Amphibians of the Pacific Northwest; Olson, D.H., Ed.; Seattle Audubon Society: Seattle, DC, USA, 2005. [Google Scholar]
- Spies, T.A.; Franklin, J.F.; Thomas, T.B. Coarse woody debris in Douglas-Fir forests of Western Oregon and Washington. Ecology 1988, 69, 1689–1702. [Google Scholar] [CrossRef]
- Homyack, J.A. Effects of Forest Regeneration Methods On Salamander Populations in Central Appalachia. Ph.D. Thesis, Department of Fisheries and Wildlife Sciences, Virginia Tech., Blacksburg, VA, USA, 2009; p. 196. [Google Scholar]
- Maser, C.; Anderson, R.G.; Cromack, K., Jr.; Williams, J.T.; Martin, R.E. Dead and Down Woody Material, in Wildlife Habitats in Managed Forests: The Blue Mountains of Oregon and Washington; Thomas, J.W., Ed.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1979; pp. 79–85. [Google Scholar]
- O’Donnell, K.; Thompson, F., III; Semlitsch, R.D. Predicting variation in microhabitat utilization of terrestrial salamanders. Herpetologica 2014, 70, 259–265. [Google Scholar] [CrossRef]
- Owens, A.K.; Moseley, K.R.; McCay, T.S.; Castleberry, S.B.; Kilgo, J.C.; Ford, W.M. Amphibian and reptile community response to coarse woody debris manipulations in upland loblolly pine (Pinus taeda) forests. For. Ecol. Manag. 2008, 256, 2078–2083. [Google Scholar] [CrossRef]
- Homyack, J.A.; Aardweg, Z.; Gorman, T.A.; Chalcraft, D.R. Initial effects of woody biomass removal and intercropping of switchgrass (Panicum virgatum) on herpetofauna in eastern North Carolina. Wildl. Soc. Bull. 2013, 248, 1–9. [Google Scholar]
- Greenberg, C.H. Response of reptile and amphibian communities to canopy gaps created by wind disturbance in the southern Appalachians. For. Ecol. Manag. 2001, 148, 135–144. [Google Scholar] [CrossRef]
- Hunter, M.L.J. A mesofilter conservation strategy to complement fine and coarse filters. Conserv. Biol. 2005, 19, 1025–1029. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Hobbs, R.J.; Montague-Drake, R.; Alexandra, R.; Bennett, A.; Burgman, M.; Cale, P.; Calhoun, A.; Cramer, V.; Cullen, P.; et al. A checklist for ecological management of landscapes for conservation. Ecol. Lett. 2008, 11, 78–91. [Google Scholar] [PubMed]
- Walls, S.C. Identifying monitoring gaps for amphibian populations in a North American biodiversity hotspot, the southeastern USA. Biodivers. Conserv 2014, in press. [Google Scholar]
- Otto, C.R.V.; Bailey, L.L.; Roloff, G.J. Improving species occupancy estimation when sampling violates the closure assumption. Ecography 2013, 36, 1299–1309. [Google Scholar] [CrossRef]
- Homyack, J.A. Evaluating habitat quality of vertebrates using conservation physiology tools. Wildl. Res. 2010, 37, 332–342. [Google Scholar] [CrossRef]
- Homyack, J.A.; Haas, C.A. Effects of repeated-stand entries on terrestrial salamanders and their habitats. Southeast. Nat. 2013, 12, 353–366. [Google Scholar] [CrossRef]
- Neu, J.; Jones, P.D.; Demarais, S.; Ezell, A.; Wigley, T.B. Retained woody structure in 1- to 2-year-old loblolly pine (Pinus taeda L.) plantations in Mississippi, Louisiana, and Arkansas: Implications for wildlife conservation. J. Sustain. For. 2014, 33, 152–172. [Google Scholar] [CrossRef]
- Bunnell, F.L.; Kremsater, L.L.; Wind, E. Managing to sustain vertebrate richness in forests of the Pacific Northwest: Relationships within stands. Environ. Rev. 1999, 7, 97–146. [Google Scholar] [CrossRef]
- Riffell, S.; Verschuyl, J.; Miller, D.; Wigley, T.B. Biofuel harvests, coarse woody debris, and biodiversity—A meta-analysis. For. Ecol. Manag. 2011, 261, 878–887. [Google Scholar] [CrossRef]
- Bunnell, F.L.; Houde, I. Downed wood and biodiversity—Implication to forest practices. Environ. Manag. 2010, 18, 397–421. [Google Scholar]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Homyack, J.A.; Kroll, A.J. Slow Lives in the Fast Landscape: Conservation and Management of Plethodontid Salamanders in Production Forests of the United States. Forests 2014, 5, 2750-2772. https://doi.org/10.3390/f5112750
Homyack JA, Kroll AJ. Slow Lives in the Fast Landscape: Conservation and Management of Plethodontid Salamanders in Production Forests of the United States. Forests. 2014; 5(11):2750-2772. https://doi.org/10.3390/f5112750
Chicago/Turabian StyleHomyack, Jessica A., and Andrew J. Kroll. 2014. "Slow Lives in the Fast Landscape: Conservation and Management of Plethodontid Salamanders in Production Forests of the United States" Forests 5, no. 11: 2750-2772. https://doi.org/10.3390/f5112750