Dendrochronological Potential in a Semi-Deciduous Rainforest: The Case of Pericopsis elata in Central Africa

 ,
,
Abstract
:1. Introduction
2. Material and Methods
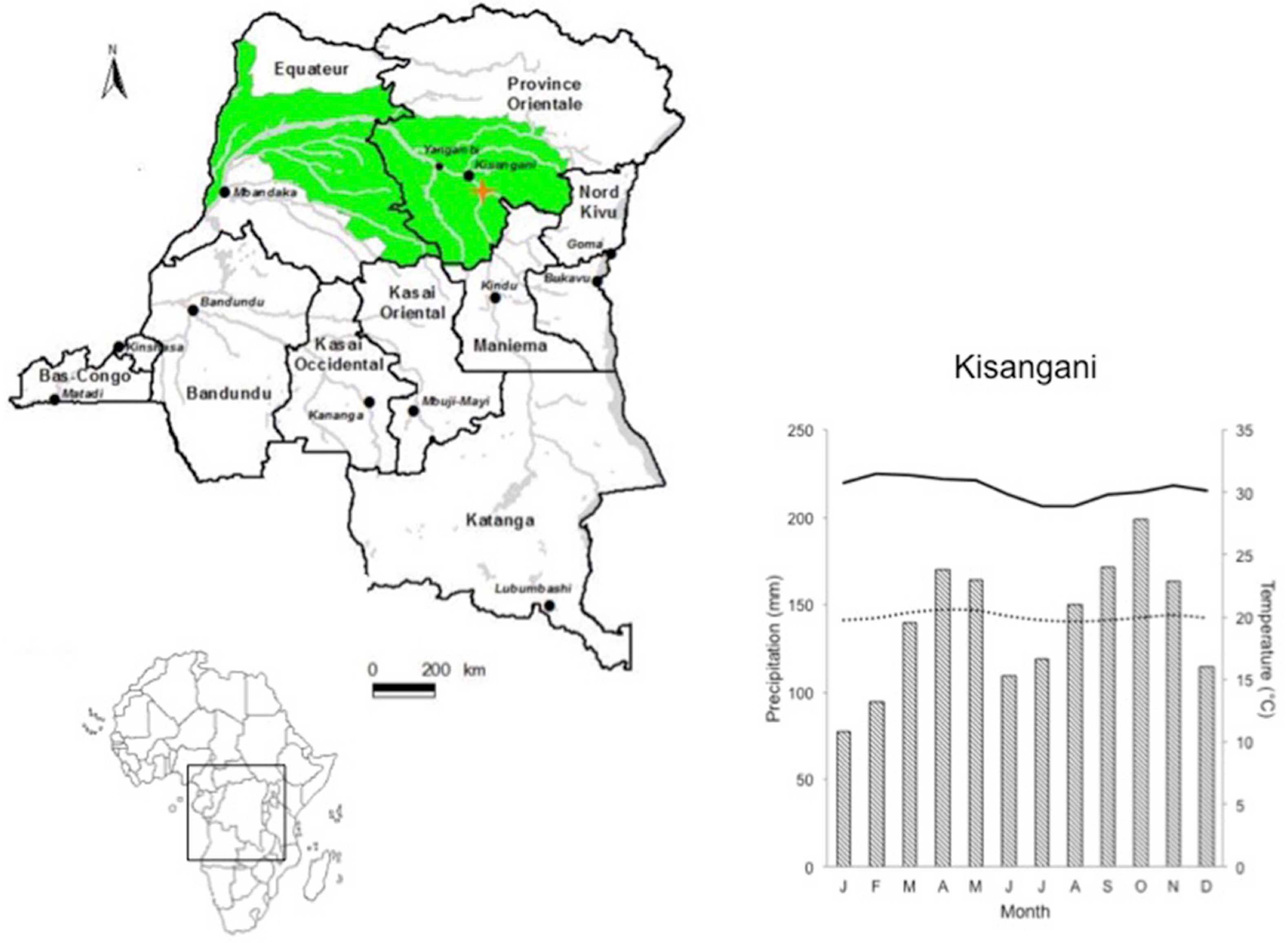
2.1. Sample Area

2.2. Sampling Stem Discs
2.3. Tree-Ring Measurements and Chronology Building
2.4. Climate-Growth Relationships
2.4.1. Growth Response to Interannual Climate Variability
2.4.2. Pointer Years and Extreme Climate Events
3. Results
3.1. General Tree-Rings Characteristics of Pericopsis Elata

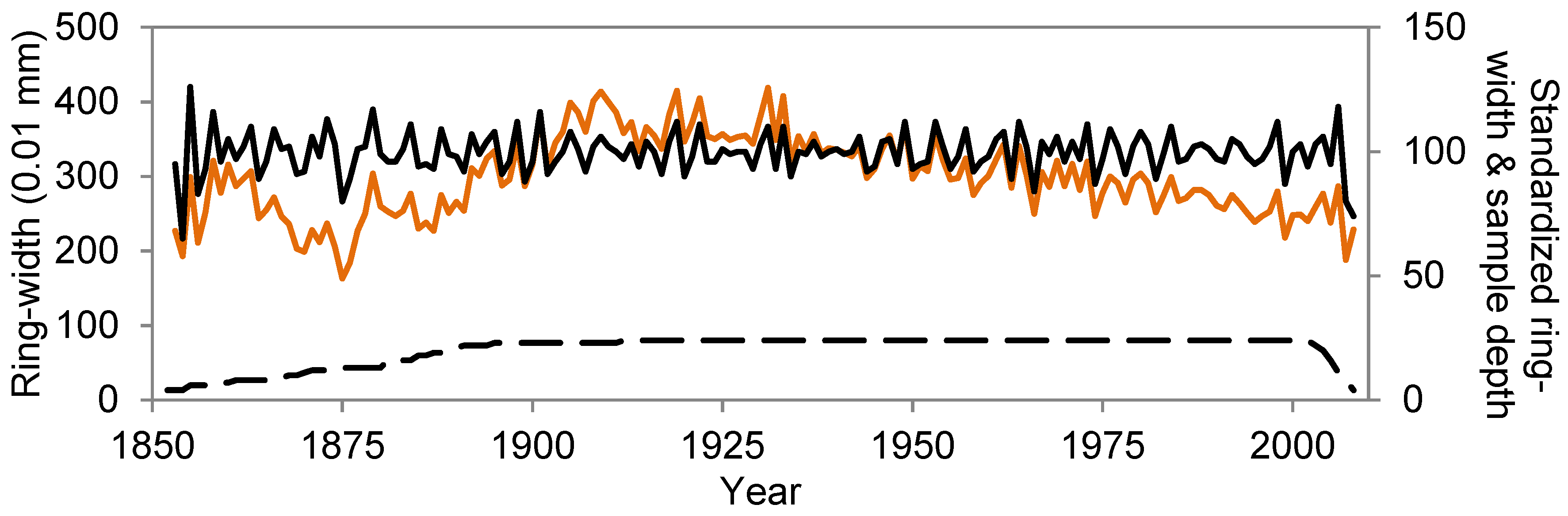
3.2. First Pericopsis Tree-Ring Chronology


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total n samples | 37 |
|---|---|
| n samples in chrono | 24 (65%) |
| Mean diameter (cm) | 82 ± 13 |
| Mean number of tree rings | 137 ± 23 |
| Time span chronology | 1852–2008 (157 years) |
| Mean ring width of chronology (mm) | 2.98 ± 0.54 |
| AC | 0.78 |
| Pearson r | 0.10 |
| EPS | 0.73 |
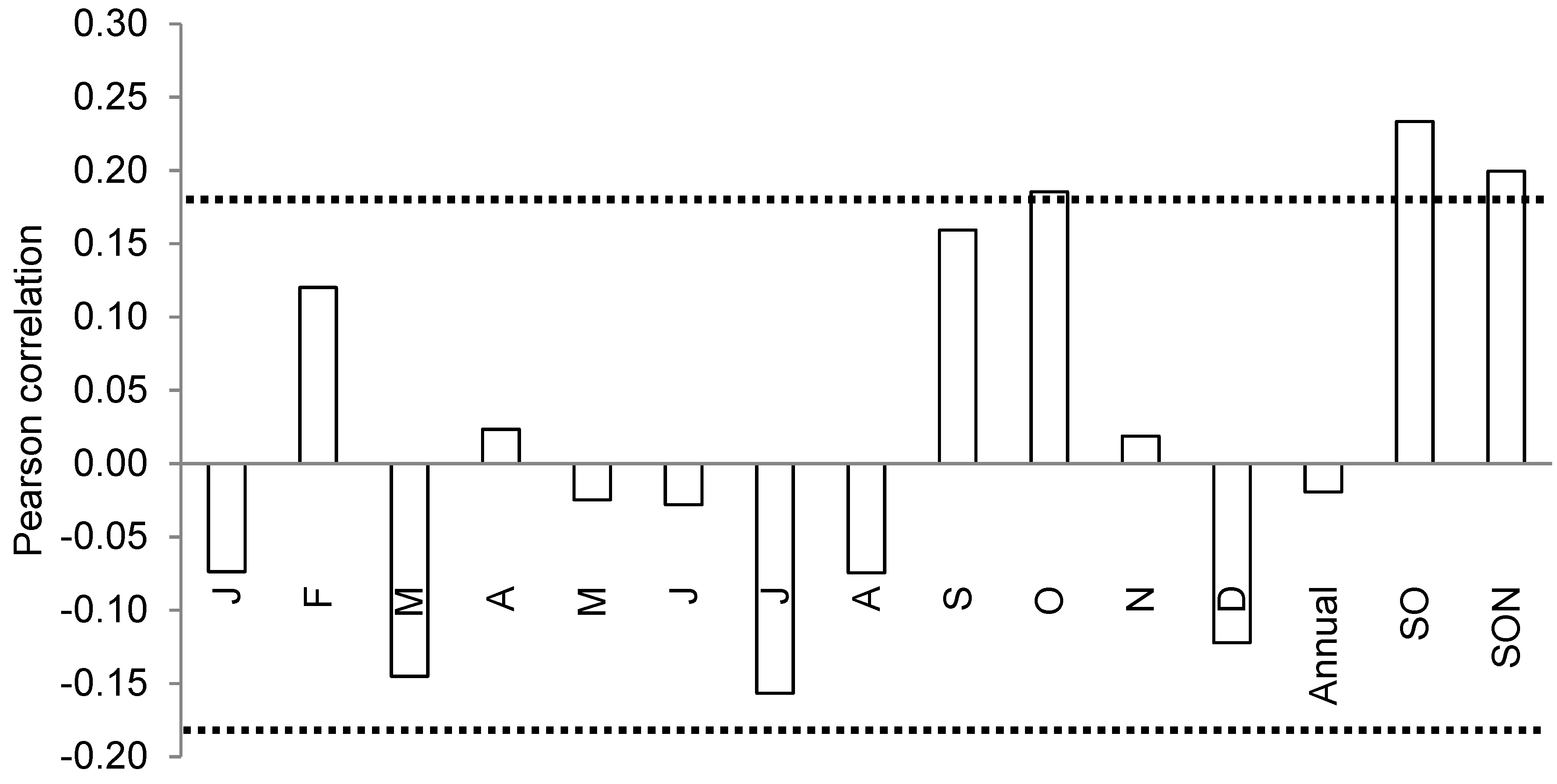
3.3. Climate-Growth Relationships
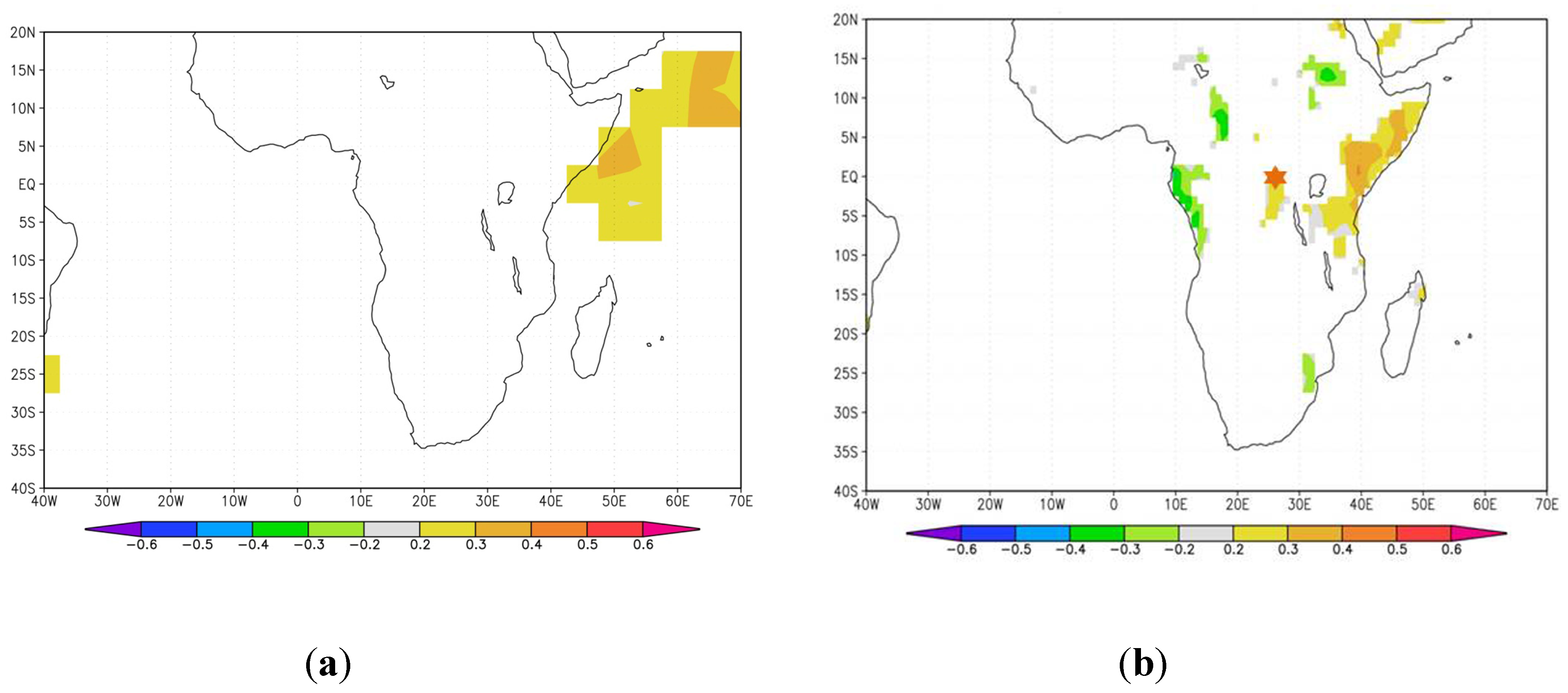
3.3.1. Growth Response to Interannual Climate Variability


3.3.2. Pointer Years and Extreme Climate Events
| Year | Annual_dry | SON_dry | Annual_wet | SON_wet | Dry_60 | El Niño | La Niña | PY | Chrono − 10% | Chrono + 10% |
|---|---|---|---|---|---|---|---|---|---|---|
| 1913 | X | |||||||||
| 1914 | X | X | X | X | ||||||
| 1921 | X | X | ||||||||
| 1927 | X | |||||||||
| 1929 | X | |||||||||
| 1930 | X | X | X | |||||||
| 1931 | X | X | ||||||||
| 1932 | X | X | ||||||||
| 1942 | X | |||||||||
| 1943 | X | X | ||||||||
| 1946 | X | |||||||||
| 1947 | X | |||||||||
| 1950 | X | X | ||||||||
| 1951 | X | X | X | |||||||
| 1952 | + | X | ||||||||
| 1961 | X | |||||||||
| 1962 | X | X | X | |||||||
| 1963 | X | + | X | |||||||
| 1967 | X | X | ||||||||
| 1971 | X | X | ||||||||
| 1972 | X | X | X | |||||||
| 1973 | X | - | X | |||||||
| 1977 | X | X | ||||||||
| 1978 | X | |||||||||
| 1982 | X | X | ||||||||
| 1983 | X | X | X | |||||||
| 1985 | X | X | ||||||||
| 1993 | X | X | ||||||||
| 1994 | X | X | ||||||||
| 1995 | X | |||||||||
| 1996 | X | X | X |
4. Discussion
4.1. Growth Patterns and Tree-Ring Chronology of Pericopsis elata
4.2. Climate-Growth Response of Pericopsis Elata
5. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Zuidema, P.A.; Baker, P.J.; Groenendijk, P.; Schippers, P.; van der Sleen, P.; Vlam, M.; Sterck, F. Tropical forests and global change: Filling knowledge gaps. Trends Plant Sci. 2013, 18, 413–419. [Google Scholar] [PubMed]
- Schweingruber, F.H.; Eckstein, D.; Serre-Bachet, F.; Bräker, O.U. Identification, presentation and interpretation of event years and pointer years in dendrochronology. Dendrochronologia 1990, 8, 9–38. [Google Scholar]
- Baker, P.J.; Bunyavejchewin, S.; Oliver, C.D.; Ashton, P.M.S. Disturbance history and historical stand dynamics of a seasonal tropical forest in western Thailand. Ecol. Monogr. 2005, 75, 317–343. [Google Scholar] [CrossRef]
- Schöngart, J.; Orthmann, B.; Hennenberg, K.J.; Porembski, S.; Worbes, M. Climate-growth relationships of tropical tree species in West Africa and their potential for climate reconstruction. Glob. Change Biol. 2006, 12, 1139–1150. [Google Scholar] [CrossRef]
- Therrell, M.D.; Stahle, D.W.; Ries, L.P.; Shugart, H.H. Tree-ring reconstructed rainfall variability in Zimbabwe. Clim. Dyn. 2006, 26, 677–685. [Google Scholar] [CrossRef]
- Schöngart, J. Growth-oriented logging (GOL): A new concept towards sustainable forest management in the vàrzea floodplains. For. Ecol. Manag. 2008, 256, 46–58. [Google Scholar] [CrossRef]
- Schöngart, J.; Arieira, J.; Fortes, C.F.; de Arruda, E.C.; da Cunha, C.N. Age-related and stand-wise estimates of carbon stocks and sequestration in the aboveground coarse wood biomass of wetland forests in the northern Pantanal, Brazil. Biogeosciences 2011, 8, 3407–3421. [Google Scholar] [CrossRef]
- Babst, F.; Moss, A.M.; Szejner, P.; Bouriaud, O.; Klesse, S.; Roden, J.; Ciais, P.; Poulter, B.; Frank, D.; Moore, D.J.P.; Trouet, V. A tree-ring perspective on the terrestrial carbon cycle. Oecologia. 2014. Available online: http://www.wsl.ch/fe/landschaftsdynamik/dendroclimatology/Publikationen/Babst_etal_Dendrochronologia_2014 (accessed on 28 October 2014). [CrossRef]
- De Ridder, M. Tree-ring analysis and stem biomass estimation of Limba (Terminalia superba Engl. & Diels) in the framework of sustainable forest management. Ph.D. Thesis, Ghent University, Ghent, Belgium, 26 June 2013. [Google Scholar]
- Schweingruber, F.H. Tree Rings: Basics and Applications of Dendrochronology, 1st ed.; Kluwer Academic Publishers: Dordrecht, the Netherlands, 1988. [Google Scholar]
- Worbes, M. One hundred years of tree-ring research in the tropic—A brief history and an outlook on future challenges. Dendrochronologia 2002, 20, 217–231. [Google Scholar] [CrossRef]
- Tarhule, A.; Hughes, M.K. Tree-ring research in semi-arid West Africa: Need and potential. Tree-Ring Res. 2002, 58, 31–46. [Google Scholar]
- Mbow, C.; Chhin, S.; Sambou, B.; Skole, D. Potential of dendrochronology to assess annual rates of biomass productivity in savanna trees of West Africa. Dendrochronologia 2013, 31, 41–51. [Google Scholar] [CrossRef]
- Pumijumnong, N. Dendrochronology in Southeast Asia. Trees. 2013. Available online: https://www.researchgate.net/profile/Nathsuda_Pumijumnong/publications (accessed on 28 October 2014). [CrossRef]
- Shah, S.K.; Bhattacharyya, A.; Chaudhury, V. Reconstruction of June-September precipitation based on tree-ring data of teak (Tectona grandis L.) from Hoshangabad, Madhya Pradesh, India. Dendrochronologia 2007, 25, 57–64. [Google Scholar] [CrossRef]
- Dünisch, O.; Montoia, V.R.; Bauch, J. Dendroecological investigations of Swietenia macrophylla King and Cedrela odorata L. (Meliaceae) in the central Amazon. Trees 2003, 17, 244–250. [Google Scholar]
- Worbes, M. How to measure growth dynamics in tropical trees. A review. IAWA J. 1995, 16, 337–351. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Zuidema, P.A. Relating tree growth to rainfall in Bolivian rain forests: A test for six species using tree ring analysis. Oecologia 2005, 146, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, E.A.; Baas, P.; Gasson, P.E. IAWA list of microscopic features for hardwood identification. IAWA J. 1989, 10, 219–332. [Google Scholar] [CrossRef]
- Douglass, A.E. Crossdating in dendrochronology. J. For. 1941, 39, 824–831. [Google Scholar]
- Bräuning, A.; Volland-Voigt, F.; Burchardt, I.; Ganzhi, O.; Nauss, T.; Peters, T. Climatic control of radial growth of Cedrela Montana in a humid mountain rainforest in southern Ecuador. Erdkunde 2009, 63, 337–345. [Google Scholar] [CrossRef]
- Worbes, M. Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropical trees from the Caparo Forest Reserve in Venezuela. J. Ecol. 1999, 87, 391–403. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Eckstein, D.; Sass, U. Tree-ring research on Tectona grandis in northern Thailand. IAWA J. 1995, 16, 385–392. [Google Scholar] [CrossRef]
- Berlage, H.P. Over het verband tusschen de dikte der jaarringen van Djatiboomen (Tectona grandis L.f.) en den regenval of Java. Tectona 1931, 24, 939–953. [Google Scholar]
- Krepkowski, J.; Bräuning, A.; Gebrekirstos, A. Growth dynamics and potential for cross-dating and multi-century climate reconstruction of Podocarpus falcatus in Ethiopia. Dendrochronologia 2012, 30, 257–265. [Google Scholar] [CrossRef]
- Trouet, V.; Esper, J.; Beeckman, H. Climate-growth relationships of Brachystegia spiciformis from the miombo woodland in south central Africa. Dendrochronologia 2010, 28, 161–171. [Google Scholar] [CrossRef]
- David, E.T.; Chhin, S.; Skole, D. Dendrochronological potential and productivity of tropical tree species in western Kenya. Tree-ring Res. 2014, 70, 119–135. [Google Scholar] [CrossRef]
- Détienne, P.; Mariaux, A. La périodicité de formation des cernes dans le bois d’okoumé. Bois For. Trop. 1970, 131, 37–50. [Google Scholar]
- Détienne, P.; Mariaux, A. Nature et périodicité des cernes dans le bois de Niangon. Bois For. Trop. 1975, 159, 29–37. [Google Scholar]
- Détienne, P.; Mariaux, A. Nature et périodicité des cernes dans le bois de samba. Bois For. Trop. 1976, 169, 29–35. [Google Scholar]
- Détienne, P.; Mariaux, A. Nature et périodicité des cernes dans les bois rouges de méliacées africaines. Bois For. Trop. 1977, 175, 52–61. [Google Scholar]
- Worbes, M.; Staschel, R.; Roloff, A.; Junk, W.J. Tree ring analysis reveals age structure, dynamics and wood production of a natural forest stand in Cameroon. For. Ecol. Manag. 2003, 173, 105–123. [Google Scholar] [CrossRef]
- De Ridder, M.; Trouet, V.; van den Bulcke, J.; Hubau, W.; van Acker, J.; Beeckman, H. A tree-ring based comparison of Terminalia superba climate-growth relationships in West and Central Africa. Trees 2013, 27, 1225–1238. [Google Scholar] [CrossRef]
- De Ridder, M.; van den Bulcke, J.; van Acker, J.; Beeckman, H. Tree-ring analysis of an African long-lived pioneer species as a tool for sustainable forest management. For. Ecol. Manag. 2013, 304, 417–426. [Google Scholar] [CrossRef]
- Groenendijk, P.; Sass-Klaassen, U.; Bongers, F.; Zuidema, P.A. Potential of tree-ring analysis in a wet tropical forest: A case study on 22 commercial tree species in Central Africa. For. Ecol. Manag. 2014, 323, 65–78. [Google Scholar] [CrossRef]
- Gebrekirstos, A.; Bräuning, A.; Sass-Klaassen, U.; Mbow, C. Opportunities and applications of dendrochronology in Africa. Curr. Opin. Environ. Sustain. 2014, 6, 48–53. [Google Scholar] [CrossRef]
- Kukachka, B.F. Krokodua—Afrormosia elata Harms, Leguminosae (Papilionaceae); Foreign Wood Series; Department of Agriculture, Forest Service: Washington, DC, USA, 1960. [Google Scholar]
- Bourland, N.; Kouadio, Y.L.; Fousséni, F.; Lejeune, P.; Doucet, J.-L. Ecology and management of Pericopsis elata (Harms) Meeuwen (Fabaceae) populations: A review. Biotechnol. Agron. Soc. Environ. 2012, 16, 486–498. [Google Scholar]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology. Applications in the Environmental Sciences, 1st ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990. [Google Scholar]
- Abensperg-Traun, M. CITES, sustainable use of wild species and incentive-driven conservation in developing countries, with an emphasis on Southern Africa. Biol. Conserv. 2009, 142, 948–963. [Google Scholar] [CrossRef]
- IUCN. IUCN Red List of Threatened Species; IUCN Species Survival Commission: Gland, Switzerland, 2001. [Google Scholar]
- White, F. The Vegetation of Africa, 1st ed; UNESCO: Gland, Switzerland, 1983. [Google Scholar]
- Van Oldenborgh, G.J.; Burgers, G. Searching for decadal variations in ENSO precipitation teleconnections. Geophys. Res. Lett. 2005, 32, L15701. [Google Scholar] [CrossRef]
- Boyemba, F.B. Ecologie de Pericopsis elata (Harms) Van Meeuwen (Fabaceae), arbre de forêt tropicale africaine à repartition agrégée. Ph.D. Thesis, Université Libre de Bruxelles, Brussels, Belgium, 2011. [Google Scholar]
- Calembert, J. Gestion des sols en Regions Chaudes, 1st ed.; Faculté Universitaire des Sciences Agronomiques: Gembloux, Belgium, 1995. [Google Scholar]
- Dickson, B.; Mathew, P.; Mickleburgh, S.; Oldfield, S.; Pouakouyou, D.; Suter, J. An Assessment of the Conservation Status, Management and Regulation of the Trade in Pericopsis elata, 1st ed.; Fauna and Flora International: Cambridge, UK, 2005. [Google Scholar]
- Rinn, F. TSAP-WinTM User Reference, 1st ed.; Rinntech: Heidelberg, Switzerland, 2003. [Google Scholar]
- Baillie, M.G.L.; Pilcher, J.R. A simple program for tree-ring research. Tree-Ring Bull. 1973, 33, 7–14. [Google Scholar]
- Eckstein, D.; Bauch, J. Beitrag zur Rationalisierung eines dendrochronologischen Verfahrens und zur Analyse seiner Aussagesicherheit. Forstwissenschaft Centralblatt 1969, 88, 230–250. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate, 1st ed.; Academic Press: London, UK, 1976. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Trouet, V.; van Oldenborgh, G.J. KNMI Climate Explorer: A web-based research tool for high-resolution paleoclimatology. Tree-Ring Res. 2013, 69, 3–13. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Stepaniak, D.P. Indices of El Niño evolution. J. Clim. 2001, 14, 1697–1701. [Google Scholar] [CrossRef]
- Mitchell, T.D.; Jones, P.D. An improved method of constructing a database of monthly climate observations and associated high-resolution grids. Int. J. Clim. 2005, 25, 693–712. [Google Scholar] [CrossRef]
- Schöngart, J.; Junk, W.J.; Piedade, M.T.F.; Ayres, J.M.; Hütterman, A.; Worbes, M. Teleconnection between tree growth in the Amazonian floodplains and the El Niño-Southern Oscillation effect. Glob. Change Biol. 2004, 10, 683–692. [Google Scholar] [CrossRef]
- Trenberth, K.E. The definition of El Niño. Bull. Amer. Meteor. Soc. 1997, 78, 2771–2777. [Google Scholar] [CrossRef]
- Worbes, M.; Fichtler, E. Wood anatomy and tree-ring structure and their importance for tropical dendrochronology. In Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management, 1st ed.; Junk, W.J., Piedade, M.T.F., Wittmann, F., Schöngart, J., Parolin, P., Eds.; Springer: Heidelberg, Switzerland, 2010; pp. 14–35. [Google Scholar]
- Stahle, D.W. Useful strategies for the development of tropical tree-ring chronologies. IAWA J. 1999, 20, 249–253. [Google Scholar] [CrossRef]
- Couralet, C.; Sterck, F.J.; Sass-Klaassen, U.; van Acker, J.; Beeckman, H. Species-specific growth responses to climate variations in understory trees of a Central African rain forest. Biotropica 2010, 42, 503–511. [Google Scholar] [CrossRef]
- Devall, M.S.; Parresol, B.R.; Wright, J.S. Dendroecological analysis of Cordia alliodora, Pseudobombax septenatum and Annona spraguei in Central Panama. IAWA J. 1995, 16, 411–424. [Google Scholar] [CrossRef]
- Bourland, N.; Kouadio, Y.L.; Lejeune, P.; Sonké, B.; Philippart, J.; Daïnou, K.; Fousséni, F.; Doucet, J.-L. Ecology of Pericopsis elata (Fabaceae), an endangered timber species in southeastern Cameroon. Biotropica 2012, 44, 840–847. [Google Scholar] [CrossRef]
- Balas, N.; Nicholson, S.E.; Klotter, D. The relationship of rainfall variability in West Central Africa to sea-surface temperature fluctuations. Int. J. Clim. 2007, 27, 1335–1349. [Google Scholar] [CrossRef]
- Camberlin, P.; Janicot, S.; Poccard, I. Seasonality and atmospheric dynamics of the teleconnection between African rainfall and tropical sea-surface temperature: Atlantic vs. ENSO. Int. J. Clim. 2001, 21, 973–1005. [Google Scholar] [CrossRef]
- Neuwirth, B.; Schweingruber, F.H.; Winiger, M. Spatial patterns of central European pointer years from 1901 to 1971. Dendrochronologia 2007, 24, 79–89. [Google Scholar]
- Wils, T.H.G.; Sass-Klaassen, U.G.W.; Eshetu, Z.; Bräuning, A.; Gebrekirstos, A.; Couralet, C.; Robertson, I.; Touchan, R.; Koprowski, M.; Conway, D.; Briffa, K.R.; Beeckman, H. Dendrochronology in the dry tropics: the Ethiopian case. Trees 2011, 25, 345–354. [Google Scholar] [CrossRef]
- Génova, M. Extreme pointer years in tree-ring records of Central Spain as evidence of volcanic eruptions (Huaynaputina, Peru, 1600 AC) and other climatic events. Clim. Past Discuss. 2011, 7, 4223–4259. [Google Scholar] [CrossRef]
- Bourland, N.; Cerisier, F.; Daïnou, K.; Smith, A.L.; Hubau, W.; Beeckman, H.; Brostaux, Y.; Fayolle, A.; Biwolé, A.B.; Fousséni, F.; et al. How tightly linked are Pericopsis elata (Fabaceae) patches to anthropogenic disturbances in south-eastern Cameroon? Implications for sustainable forest management. Forests 2014. submitted. [Google Scholar]
- Bourland, N. Dynamique d’une espèce ligneuse héliophile longévive dans un monde changeant: le cas de Pericopsis elata (Harms) Meeuwen (Fabaceae) au sud-est du Cameroun. Ph.D. Thesis, Université de Liège/Gembloux Agro-Bio Tech, Gembloux, Belgium, 6 December 2013. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Ridder, M.; Toirambe, B.; Van Den Bulcke, J.; Bourland, N.; Van Acker, J.; Beeckman, H. Dendrochronological Potential in a Semi-Deciduous Rainforest: The Case of Pericopsis elata in Central Africa. Forests 2014, 5, 3087-3106. https://doi.org/10.3390/f5123087
De Ridder M, Toirambe B, Van Den Bulcke J, Bourland N, Van Acker J, Beeckman H. Dendrochronological Potential in a Semi-Deciduous Rainforest: The Case of Pericopsis elata in Central Africa. Forests. 2014; 5(12):3087-3106. https://doi.org/10.3390/f5123087
Chicago/Turabian StyleDe Ridder, Maaike, Benjamin Toirambe, Jan Van Den Bulcke, Nils Bourland, Joris Van Acker, and Hans Beeckman. 2014. "Dendrochronological Potential in a Semi-Deciduous Rainforest: The Case of Pericopsis elata in Central Africa" Forests 5, no. 12: 3087-3106. https://doi.org/10.3390/f5123087