
Seasonal Variation in Soil Greenhouse Gas Emissions at Three Age-Stages of Dawn Redwood (Metasequoia glyptostroboides) Stands in an Alluvial Island, Eastern China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
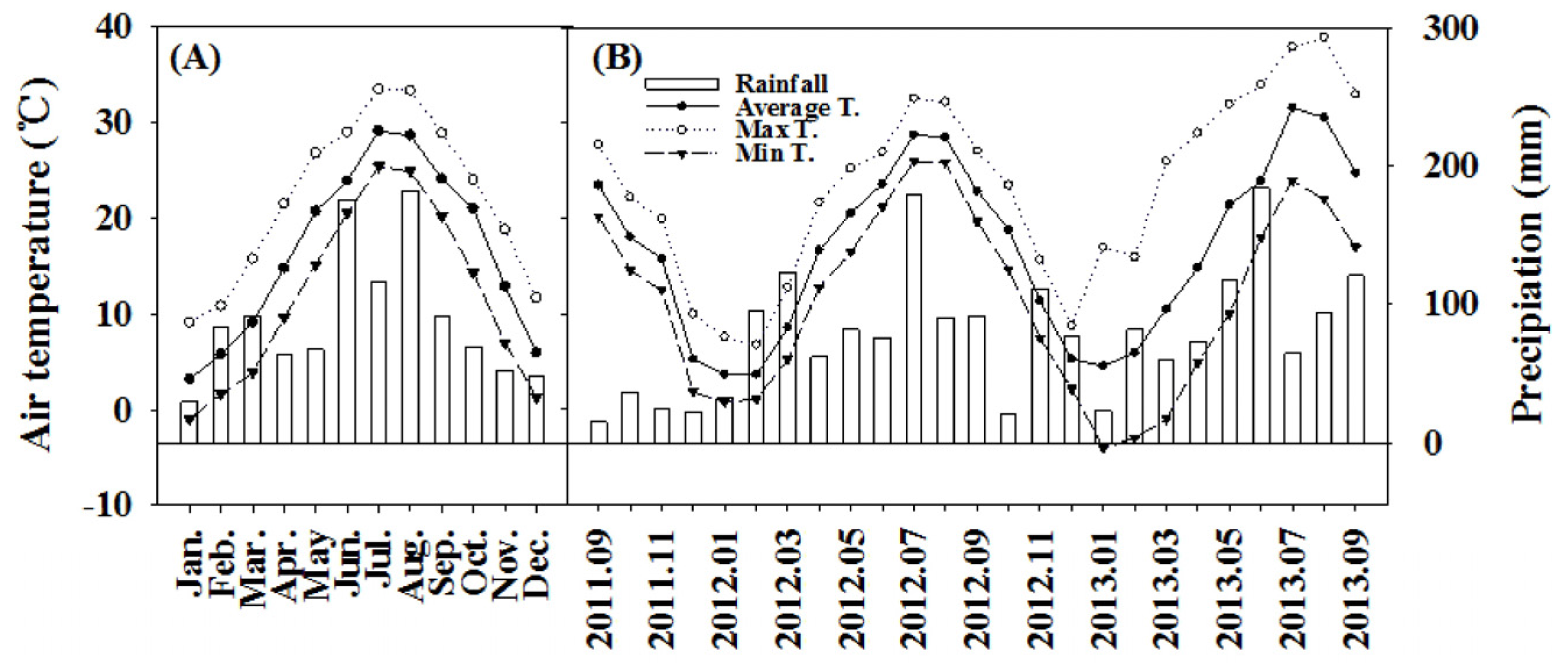
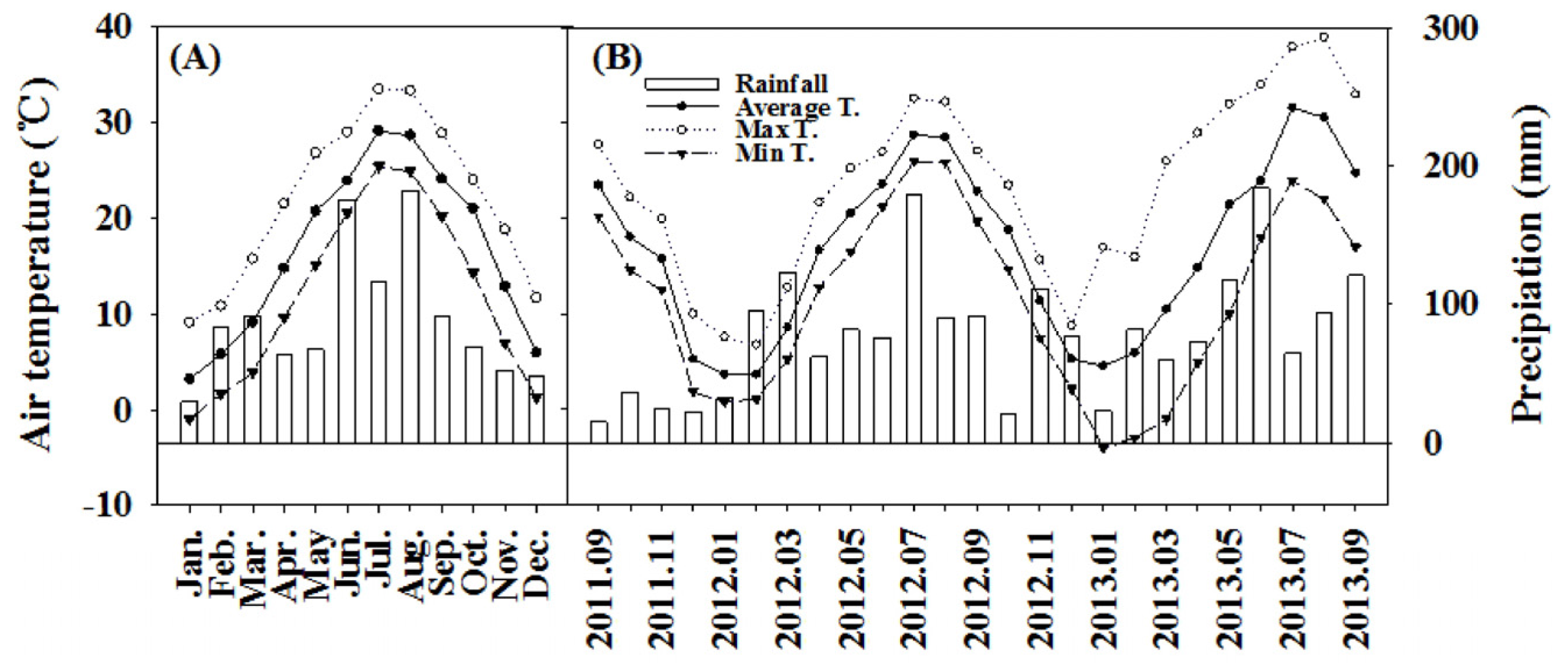
2.1. Site Description
2.2. Measurements
2.2.1. Soil Gas Emissions
2.2.2. Soil Temperature and Soil Water Content
2.3. Data Analysis
2.3.1. Q10 Values
2.3.2. The Relationship between GHG Emissions and Environmental Factors
3. Results
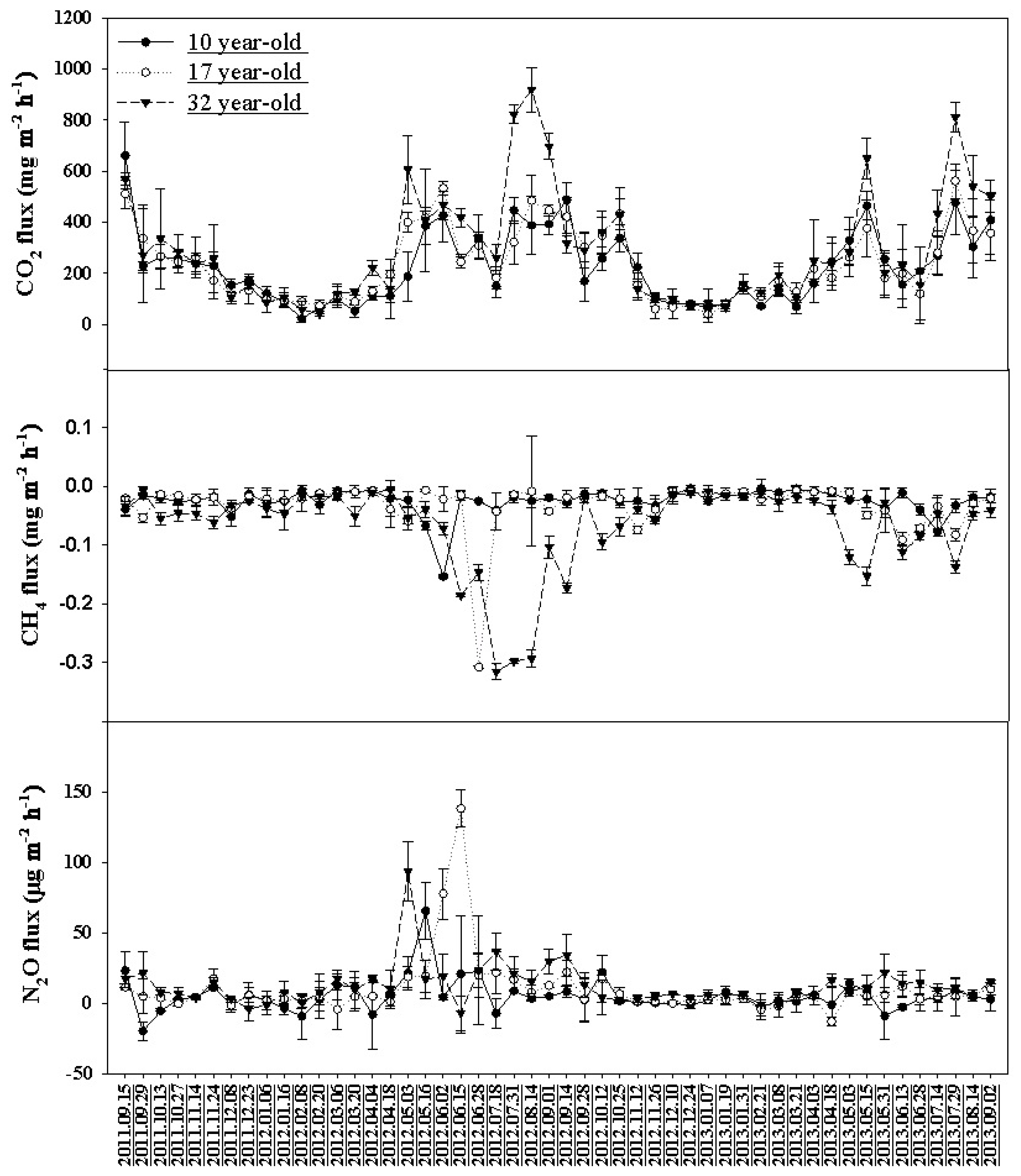
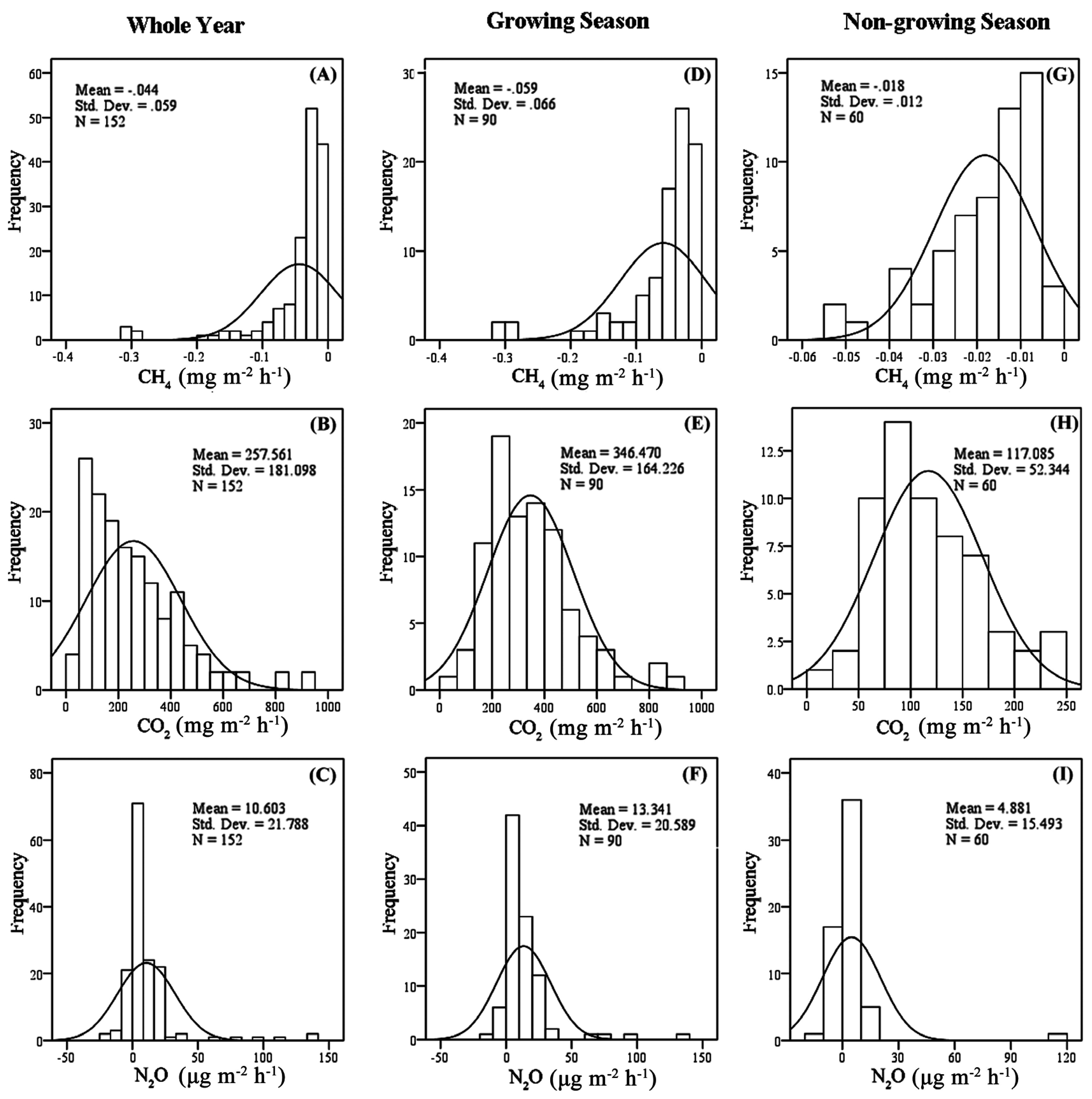
3.1. Soil Respiration Rate
3.2. Soil CH4 Uptake
3.3. Soil N2O Emission
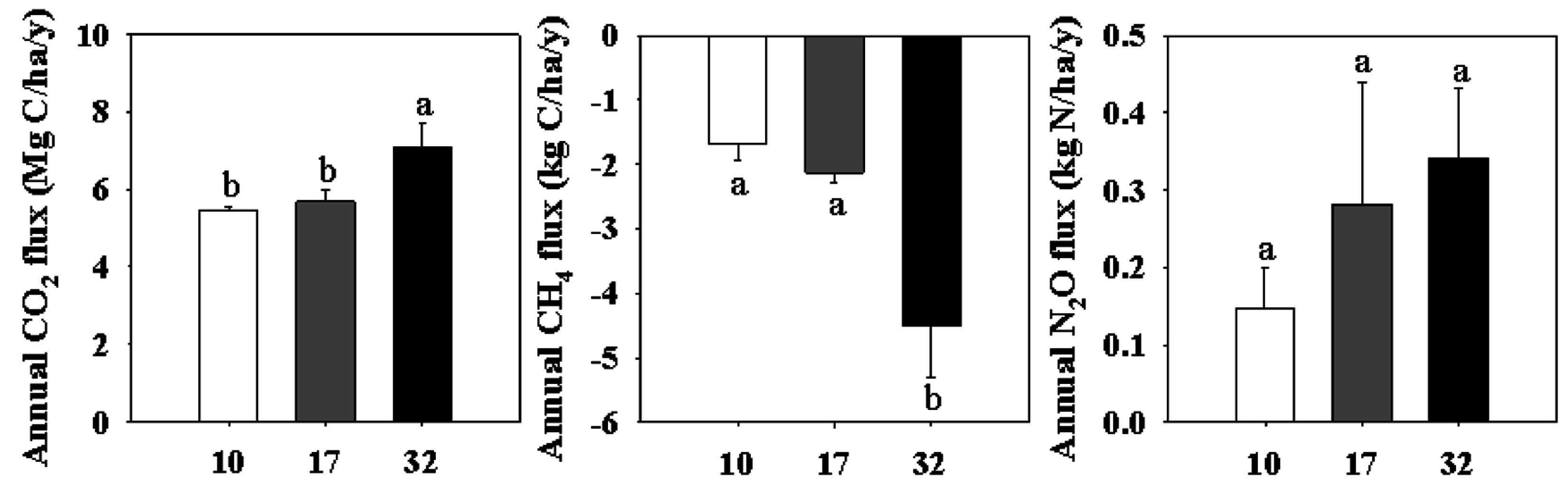
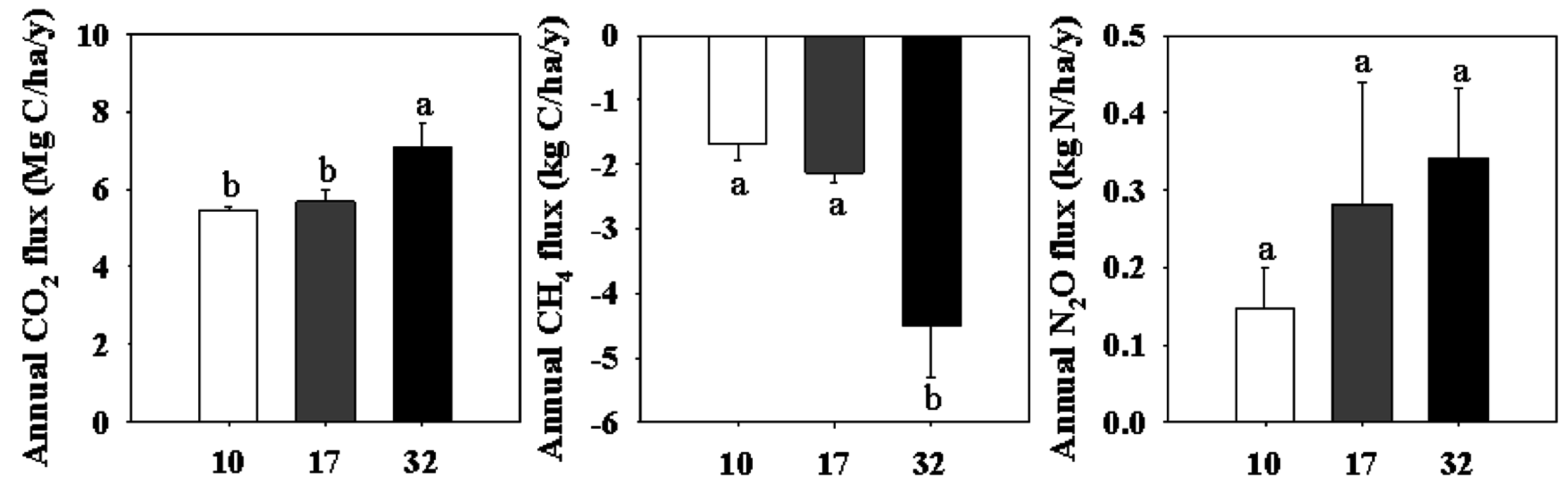
3.4. Annual GHG Emissions
3.5. The Effect of Soil Temperature on GHG Emissions
3.6. Effects of Soil Water Content on GHG Emissions
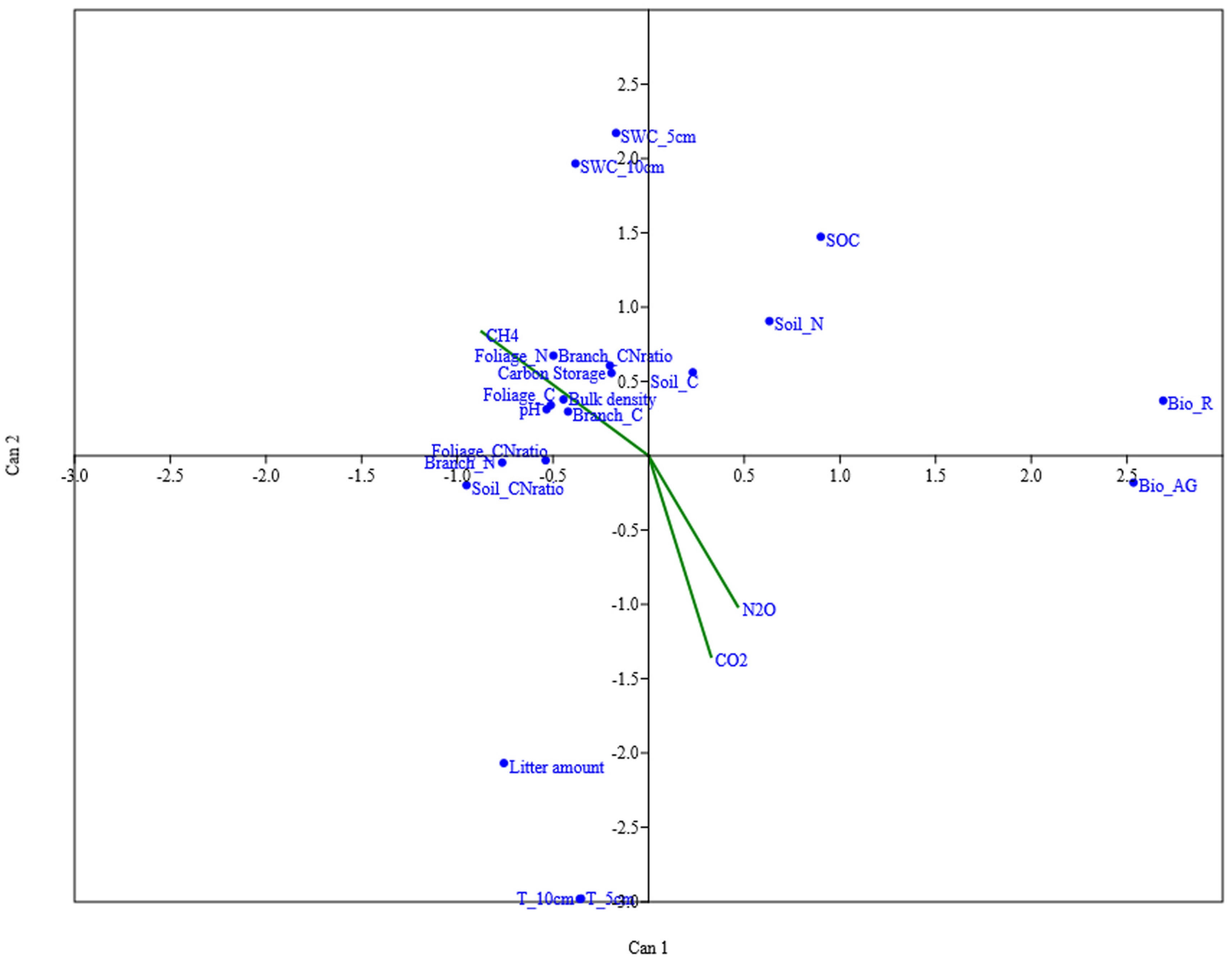
3.7. The Main Influencing Factors of Soil Greenhouse Gas Emissions
4. Discussion
4.1. Soil Carbon Dynamic in Different-Age Stands
4.2. Soil Nitrogen Dynamic in Different-Aged Stands
4.3. Factors Affecting Soil Greenhouse Gas Emissions
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Chapuis-Lardy, L.; Wrage, N.; Metay, A.; Chotte, J.; Bernoux, M. Soils, a sink for N2O? A review. Glob. Chang. Biol. 2007, 13, 1–17. [Google Scholar] [CrossRef]
- Carle, J.; Vuorinen, P.; Del Lungo, A. Status and trends in global forest plantation development. For. Prod. J. 2002, 52, 12–23. [Google Scholar]
- Dai, L.; Wang, Y.; Su, D.; Zhou, L.; Yu, D.; Lewis, B.J.; Qi, L. Major forest types and the evolution of sustainable forestry in China. Environ. Manag. 2011, 48, 1066–1078. [Google Scholar] [CrossRef] [PubMed]
- Central Committee of the Communist Party of China and China State Council. The Programming Outline for the Protection and Utilization of National Forestland (2010–2020); Central Committee of the Communist Party of China and China State Council: Beijing, China, 2010.
- Zhang, D.; Sun, X.; Zhou, G.; Yan, J.; Wang, Y.; Liu, S.; Zhou, C.; Liu, J.; Tang, X.; Li, J. Seasonal dynamics of soil CO2 effluxes with responses to environmental factors in lower subtropical forests of China. Sci. China Ser. D Earth Sci. 2006, 49, 139–149. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, G.; Guo, J.; Xie, J.; Wang, X. Soil respiration and carbon balance in a subtropical native forest and two managed plantations. Plant Ecol. 2007, 193, 71–84. [Google Scholar] [CrossRef]
- Iqbal, J.; Ronggui, H.; Lijun, D.; Lan, L.; Shan, L.; Tao, C.; Leilei, R. Differences in soil CO2 flux between different land use types in mid-subtropical China. Soil Biol. Biochem. 2008, 40, 2324–2333. [Google Scholar] [CrossRef]
- Rowlings, D.; Grace, P.; Kiese, R.; Weier, K. Environmental factors controlling temporal and spatial variability in the soil-atmosphere exchange of CO2, CH4 and N2O from an Australian subtropical rainforest. Glob. Chang. Biol. 2012, 18, 726–738. [Google Scholar] [CrossRef]
- Hashimoto, S.; Tanaka, N.; Suzuki, M.; Inoue, A.; Takizawa, H.; Kosaka, I.; Tanaka, K.; Tantasirin, C.; Tangtham, N. Soil respiration and soil CO2 concentration in a tropical forest, Thailand. J. For. Res. 2004, 9, 75–79. [Google Scholar] [CrossRef]
- Davidson, E.; Ishida, F.; Nepstad, D. Effects of an experimental drought on soil emissions of carbon dioxide, methane, nitrous oxide, and nitric oxide in a moist tropical forest. Glob. Chang. Biol. 2004, 10, 718–730. [Google Scholar] [CrossRef]
- Tang, X.; Liu, S.; Zhou, G.; Zhang, D.; Zhou, C. Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China. Glob. Chang. Biol. 2006, 12, 546–560. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, W.; Wang, K.; Qin, F.; Wang, W.; Dai, H.; Li, P. Responses of CO2, N2O and CH4 fluxes between atmosphere and forest soil to changes in multiple environmental conditions. Glob. Chang. Biol. 2014, 20, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Dutaur, L.; Verchot, L.V. A global inventory of the soil CH4 sink. Glob. Biogeochem. Cycles 2007, 21, 7949–7950. [Google Scholar] [CrossRef]
- Megonigal, J.P.; Guenther, A.B. Methane emissions from upland forest soils and vegetation. Tree Physiol. 2008, 28, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.; Flessa, H.; Veldkamp, E. Atmospheric methane uptake by tropical montane forest soils and the contribution of organic layers. Biogeochemistry 2012, 111, 469–483. [Google Scholar] [CrossRef]
- Smith, K.; Ball, T.; Conen, F.; Dobbie, K.; Massheder, J.; Rey, A. Exchange of greenhouse gases between soil and atmosphere: Interactions of soil physical factors and biological processes. Eur. J. Soil Sci. 2003, 54, 779–791. [Google Scholar] [CrossRef]
- Yan, Y.; Sha, L.; Cao, M.; Zheng, Z.; Tang, J.; Wang, Y.; Zhang, Y.; Wang, R.; Liu, G.; Wang, Y. Fluxes of ch4 and n2o from soil under a tropical seasonal rain forest in xishuangbanna, Southwest China. J. Environ. Sci. 2008, 20, 207–215. [Google Scholar] [CrossRef]
- Moore, K.; Fitzjarrald, D.; Sakai, R.; Goulden, M.; Munger, J.; Wofsy, S. Seasonal variation in radiative and turbulent exchange at a deciduous forest in central massachusetts. J. Appl. Meteorol. 1996, 35, 122–134. [Google Scholar] [CrossRef]
- Livesley, S.; Grover, S.; Hutley, L.; Jamali, H.; Butterbach-Bahl, K.; Fest, B.; Beringer, J.; Arndt, S. Seasonal variation and fire effects on CH4, N2O and CO2 exchange in savanna soils of northern Australia. Agric. For. Meteorol. 2011, 151, 1440–1452. [Google Scholar] [CrossRef]
- LePage, B.; Williams, C.; Yang, H. The Geobiology and Ecology of Metasequoia; Springer: Dordrecht, The Netherlands, 2005; Volume 22. [Google Scholar]
- Chu, K.; Cooper, W. An ecological reconnaissance in the native home of Metasequoia glyptostroboides. Ecology 1950, 31, 260–278. [Google Scholar] [CrossRef]
- Bajpai, V.; Rahman, A.; Kang, S. Chemical composition and anti-fungal properties of the essential oil and crude extracts of Metasequoia glyptostroboides miki ex hu. Ind. Crops Prod. 2007, 26, 28–35. [Google Scholar] [CrossRef]
- Polman, J.; Michon, S.; Militz, H.; Helmink, A. The wood of Metasequoia glyptostroboides (hu et cheng) of dutch origin. Holz als Roh-und Werkstoff 1999, 57, 215–221. [Google Scholar] [CrossRef]
- Bureau C.S.S. Chapter 1 Comprehensive. In Chongming Statistical Yearbook; Shanghai Statistics press: Chongming, Shanghai, China, 2016. [Google Scholar]
- SMC. Shanghai Meteorological data. Available online: http://www.smb.gov.cn/ (accessed on 27 Feberury 2014).
- Xiaver, B. Allometric Estimation of the Aboveground Biomass and Carbon in Metasequoia Glyptostroboide Plantations in Shanghai; Cranfield University: Bedfordshire, UK, 2009. [Google Scholar]
- Pedrotti, A.; Pauletto, E.; Crestana, S.; Holanda, F.; Cruvinel, P.; Vaz, C. Evaluation of bulk density of albaqualf soil under different tillage systems using the volumetric ring and computerized tomography methods. Soil Tillage Res. 2005, 80, 115–123. [Google Scholar] [CrossRef]
- Schofield, R.; Taylor, A. The measurement of soil pH. Soil Sci. Soc. Am. J. 1955, 19, 164–167. [Google Scholar] [CrossRef]
- Xiao, C. Characteristics and Carbon-Storage Estimation of Metasequoia glyptostroboides Plantation Ecosystems at Different Age Stages in Chongming Island. Master’ Dissertation, Shanghai Jiao Tong University, Shanghai, China, 2010. [Google Scholar]
- Wang, Y.; Wang, Y.; Ling, H. A new carrier gas type for accurate measurement of N2O by GC-ECD. Adv. Atmos. Sci. 2010, 27, 1322–1330. [Google Scholar] [CrossRef]
- Zheng, X.; Mei, B.; Wang, Y.; Xie, B.; Wang, Y.; Dong, H.; Xu, H.; Chen, G.; Cai, Z.; Yue, J. Quantification of n2o fluxes from soil–plant systems may be biased by the applied gas chromatograph methodology. Plant Soil 2008, 311, 211–234. [Google Scholar] [CrossRef]
- Yao, Z.; Wolf, B.; Chen, W.; Butterbach-Bahl, K.; Brüggemann, N.; Wiesmeier, M.; Dannenmann, M.; Blank, B.; Zheng, X. Spatial variability of N2O, CH4 and CO2 fluxes within the xilin river catchment of inner mongolia, China: A soil core study. Plant Soil 2010, 331, 341–359. [Google Scholar] [CrossRef]
- Hoff, J. Lectures on Theoretical and Physical Chemistry; Edward Arnold: London, UK, 1899. [Google Scholar]
- Davidson, E.; Janssens, I. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Janssens, I.; Carrara, A.; Ceulemans, R. Annual Q10 of soil respiration reflects plant phenological patterns as well as temperature sensitivity. Glob. Chang. Biol. 2004, 10, 161–169. [Google Scholar]
- Zimmermann, M.; Bird, M.; Zechmeister-Boltenstern, S. Q10 values of tropical forest soil respiration increases with ongoing decomposition. In Proceedings of the EGU General Assembly, Vienna, Austria, 22–27 April 2012; p. 4347.
- Werner, C.; Zheng, X.; Tang, J.; Xie, B.; Liu, C.; Kiese, R.; Butterbach-Bahl, K. N2O, CH4 and CO2 emissions from seasonal tropical rainforests and a rubber plantation in Southwest China. Plant Soil 2006, 289, 335–353. [Google Scholar] [CrossRef]
- Kiese, R.; Hewett, B.; Graham, A.; Butterbach-Bahl, K. Seasonal variability of N2O emissions and CH4 uptake by tropical rainforest soils of Queensland, Australia. Glob. Biogeochem. Cycles 2003, 17, 469–474. [Google Scholar] [CrossRef]
- Davidson, E.; Belk, E.; Boone, R. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Bowden, R.; Newkirk, K.; Rullo, G. Carbon dioxide and methane fluxes by a forest soil under laboratory-controlled moisture and temperature conditions. Soil Biol. Biochem. 1998, 30, 1591–1597. [Google Scholar] [CrossRef]
- Schaufler, G.; Kitzler, B.; Schindlbacher, A.; Skiba, U.; Sutton, M.; Zechmeister-Boltenstern, S. Greenhouse gas emissions from european soils under different land use: Effects of soil moisture and temperature. Eur. J. Soil Sci. 2010, 61, 683–696. [Google Scholar] [CrossRef]
- Tjoelker, M.; Oleksyn, J.; Reich, P. Modelling respiration of vegetation: Evidence for a general temperature-dependent Q10. Glob. Chang. Biol. 2001, 7, 223–230. [Google Scholar] [CrossRef]
- Högberg, P.; Read, D. Towards a more plant physiological perspective on soil ecology. Trends Ecol. Evol. 2006, 21, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Pavelka, M.; Acosta, M.; Marek, M.; Kutsch, W.; Janous, D. Dependence of the Q10 values on the depth of the soil temperature measuring point. Plant Soil 2007, 292, 171–179. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J. On the temperature dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Chen, C.; Yang, Z.; Xie, J.; Liu, X.; Zhong, X. Seasonal variations of soil CH4 uptake rate in castanopsis carlesii forest in mid-subtropical China. J. Appl. Ecol. 2012, 23, 17–22. [Google Scholar]
- Adamsen, A.; King, G. Methane consumption in temperate and subarctic forest soils: Rates, vertical zonation, and responses to water and nitrogen. Appl. Environ. Microbiol. 1993, 59, 485–490. [Google Scholar] [PubMed]
- Singh, J.; Singh, S.; Raghubanshi, A.; Singh, S.; Kashyap, A.; Reddy, V. Effect of soil nitrogen, carbon and moisture on methane uptake by dry tropical forest soils. Plant Soil 1997, 196, 115–121. [Google Scholar] [CrossRef]
- Luo, G.; Kiese, R.; Wolf, B.; Butterbach-Bahl, K. Effects of soil temperature and moisture on methane uptake and nitrous oxide emissions across three different ecosystem types. Biogeosciences 2013, 10, 3205–3219. [Google Scholar] [CrossRef]
- Yavitt, J.; Downey, D.; Lang, G.; Sexston, A. Methane consumption in two temperate forest soils. Biogeochemistry 1990, 9, 39–52. [Google Scholar] [CrossRef]
- Hanson, R.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [PubMed]
- Whalen, S.; Reeburgh, W. Moisture and temperature sensitivity of CH4 oxidation in boreal soils. Soil Biol. Biochem. 1996, 28, 1271–1281. [Google Scholar] [CrossRef]
- Von Arnold, K.; Nilsson, M.; Hånell, B.; Weslien, P.; Klemedtsson, L. Fluxes of CO2, CH4 and N2O from drained organic soils in deciduous forests. Soil Biol. Biochem. 2005, 37, 1059–1071. [Google Scholar] [CrossRef]
- Breuer, L.; Papen, H.; Butterbach-Bahl, K. N2O emission from tropical forest soils of Australia. J. Geophys. Res. Atmos. 2000, 105, 26353–26367. [Google Scholar] [CrossRef]
- Pihlatie, M.; Syväsalo, E.; Simojoki, A.; Esala, M.; Regina, K. Contribution of nitrification and denitrification to N2O production in peat, clay and loamy sand soils under different soil moisture conditions. Nutr. Cycl. Agroecosyst. 2004, 70, 135–141. [Google Scholar] [CrossRef]
- Morishita, T.; Aizawa, S.; Yoshinaga, S.; Kaneko, S. Seasonal change in N2O flux from forest soils in a forest catchment in japan. J. For. Res. 2011, 16, 386–393. [Google Scholar] [CrossRef]
- Gundersen, P.; Christiansen, J.; Alberti, G.; Brüggemann, N.; Castaldi, S.; Gasche, R.; Kitzler, B.; Klemedtsson, L.; Lobo-do-Vale, R.; Moldan, F. The response of methane and nitrous oxide fluxes to forest change in Europe. Biogeosciences 2012, 9, 3999–4012. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Wang, J.; Shi, Z.; Lu, L.; Zeng, J.; Ming, A.; Tang, J.; Yu, H. Effects of tree species mixture on soil organic carbon stocks and greenhouse gas fluxes in subtropical plantations in China. For. Ecol. Manag. 2013, 300, 4–13. [Google Scholar] [CrossRef]
- Hanson, P.; Edwards, N.; Garten, C.; Andrews, J. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Lichter, J. Limited carbon storage in soil and litter of experimental forest plots under increased atmospheric CO2. Nature 2001, 411, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, Z.; Zha, T.; Luo, Z.; Zheng, J.; Sun, O. Predicting soil respiration using carbon stock in roots, litter and soil organic matter in forests of Loess Plateau in China. Soil Biol. Biochem. 2013, 57, 135–143. [Google Scholar] [CrossRef]
- Chang, S.; Tseng, K.; Hsia, Y.; Wang, C.; Wu, J. Soil respiration in a subtropical montane cloud forest in taiwan. Agric. For. Meteorol. 2008, 148, 788–798. [Google Scholar] [CrossRef]
- Raich, J.; Tufekciogul, A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry 2000, 48, 71–90. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Koehler, B.; Corre, M.; Steger, K.; Well, R.; Zehe, E.; Sueta, J.; Veldkamp, E. An in-depth look into a tropical lowland forest soil: Nitrogen-addition effects on the contents of N2O, CO2 and CH4 and N2O isotopic signatures down to 2-m depth. Biogeochemistry 2012, 111, 695–713. [Google Scholar] [CrossRef]
- Fender, A.; Pfeiffer, B.; Gansert, D.; Leuschner, C.; Daniel, R.; Jungkunst, H. The inhibiting effect of nitrate fertilisation on methane uptake of a temperate forest soil is influenced by labile carbon. Biol. Fertil. Soils 2012, 48, 621–631. [Google Scholar] [CrossRef]
- Vanitchung, S.; Conrad, R.; Harvey, N.; Chidthaisong, A. Fluxes and production pathways of nitrous oxide in different types of tropical forest soils in Thailand. Soil Sci. Plant Nutr. 2011, 57, 650–658. [Google Scholar] [CrossRef]
- Heil, J.; Liu, S.; Vereecken, H.; Brüggemann, N. Mechanisms of inorganic nitrous oxide production in soils during nitrification and their dependence on soil properties. In Proceedings of the EGU General Assembly, Vienna, Austria, 27 April–2 May 2014; p. 4208.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 10-Year-Old Stand | 17-Year-Old Stand | 32-Year-Old Stand | |
|---|---|---|---|
| Tree Growth | |||
| Tree density (stems/ha) | 1050 | 725 | 550 |
| Average height (m) | 8.1 ± 1.5 | 16.2 ± 2.2 | 28.3 ± 3.4 |
| Average DBH (cm) | 10.5 ± 3.1 | 17.5 ± 3.4 | 27.2 ± 3.0 |
| Biomass carbon stock (t ha−1) | 13.96 | 29.76 | 64.93 |
| Litter (a) | |||
| Litter amount (t ha−1) | 1.04 ± 0.008 | 2.67 ± 0.012 | 3.87 ± 0.027 |
| Fallen leaf C (%) | 47.35 ± 0.61 | 48.11 ± 0.32 | 47.67 ± 0.40 |
| Fallen branch C (%) | 44.40 ± 0.33 | 44.86 ± 0.32 | 46.07 ± 0.37 |
| Fallen leaf N (%) | 1.60 ± 0.09 | 1.84 ± 0.05 | 1.69 ± 0.08 |
| Fallen branch N (%) | 0.75 ± 0.04 | 0.63 ± 0.04 | 0.64 ± 0.05 |
| Fallen leaf C:N ratio | 29.6 | 26.1 | 28.2 |
| Fallen branch C:N ratio | 59.2 | 71.2 | 72.0 |
| Soil Properties | |||
| Bulk density | 1.55 ± 0.01 | 1.62 ± 0.01 | 1.62 ± 0.01 |
| pH | 8.18 ± 0.068 | 8.19 ± 0.097 | 8.12 ± 0.063 |
| Total N (%) | 0.11 ± 0.014 | 0.19 ± 0.038 | 0.22 ± 0.002 |
| SOC (%) | 0.71 | 1.78 | 1.94 |
| Total C (%) | 1.40 ± 0.014 | 1.85 ± 0.036 | 2.11 ± 0.054 |
| C:N ratio | 13 | 10 | 10 |
| Soil carbon storage (t ha−1) | 31.87 ± 2.20 | 37.68 ± 1.07 | 40.01 ± 2.49 |
| Stand Age | 2011–2012 | 2012–2013 | 2011–2013 | |
|---|---|---|---|---|
| CH4 (mg m−2 h−1) | 10 | −0.030 ± 0.029 b | −0.021 ± 0.016 b | −0.026 ± 0.024 b |
| 17 | −0.035 ± 0.059 b | −0.030 ± 0.025 b | −0.032± 0.045 b | |
| 32 | −0.081 ± 0.093 a | −0.056 ± 0.049 a | −0.069 ± 0.075 a | |
| CO2 (mg m−2 h−1) | 10 | 233.35 ± 152.28 a | 223.25 ± 134.76 a | 228.30 ± 142.40 a |
| 17 | 250.42 ± 146.93 a | 225.86 ± 139.22 a | 238.14±142.20 a | |
| 32 | 322.40 ± 241.16 a | 273.01 ± 194.12 a | 297.71 ± 218.09 a | |
| N2O (μg m−2 h−1) | 10 | 7.17 ± 16.12 a | 3.40 ± 6.05 a | 5.29 ± 12.20 a |
| 17 | 15.79 ± 29.95 a | 4.38 ± 6.68 a | 10.09 ± 22.23 a | |
| 32 | 15.46 ± 19.23 a | 9.04 ± 7.56 b | 12.25 ± 14.82 a |
| Sites | 10-Year-Old Stand | 17-Year-Old Stand | 32-Year-Old Stand | Three Stands | |||||
|---|---|---|---|---|---|---|---|---|---|
| Soil Depth (cm) | 5 | 10 | 5 | 10 | 5 | 10 | 5 | 10 | |
| Whole Year | R2 | 0.58 | 0.65 | 0.72 | 0.66 | 0.79 | 0.78 | 0.68 | 0.69 |
| α | 67.61 | 59.71 | 62.01 | 65.99 | 58.23 | 60.60 | 62.78 | 61.89 | |
| β | 0.0659 | 0.0752 | 0.0732 | 0.0689 | 0.0872 | 0.0856 | 0.0752 | 0.0767 | |
| Q10 | 1.93 | 2.12 | 2.08 | 1.99 | 2.39 | 2.35 | 2.12 | 2.15 | |
| Growing Season | R2 | 0.24 | 0.23 | 0.36 | 0.23 | 0.52 | 0.51 | 0.36 | 0.69 |
| α | 132.36 | 129.40 | 89.30 | 113.75 | 77.02 | 84.09 | 96.82 | 61.89 | |
| β | 0.0383 | 0.0397 | 0.0569 | 0.0448 | 0.0746 | 0.0708 | 0.0568 | 0.0767 | |
| Q10 | 1.47 | 1.49 | 1.77 | 1.57 | 2.11 | 2.03 | 1.76 | 2.15 | |
| Non-growing Season | R2 | 0.18 | 0.33 | 0.59 | 0.54 | 0.61 | 0.57 | 0.35 | 0.69 |
| α | 71.56 | 51.86 | 55.85 | 55.52 | 53.15 | 52.70 | 65.24 | 61.89 | |
| β | 0.036 | 0.0811 | 0.0808 | 0.0824 | 0.0947 | 0.0986 | 0.0599 | 0.0767 | |
| Q10 | 1.43 | 2.25 | 2.24 | 2.28 | 2.58 | 2.68 | 1.82 | 2.15 | |
| CH4 | CO2 | N2O | T 5 cm | T 10 cm | SWC 0−10 cm | SWC 10–20 cm | |
|---|---|---|---|---|---|---|---|
| CH4 | 1.000 | −0.377 ** | −0.041 | −0.301 ** | −0.317 ** | −0.012 | 0.169 |
| CO2 | 1.000 | 0.380 ** | 0.765 ** | 0.776 ** | −0.211 * | −0.276 ** | |
| N2O | 1.000 | 0.274 ** | 0.274 ** | 0.141 | −0.047 | ||
| T 5 cm | 1.000 | 0.972 ** | −0.319 ** | −0.364 ** | |||
| T 10 cm | 1.000 | −0.324 ** | −0.385 ** | ||||
| SWC 0–10 cm | 1.000 | 0.671 ** | |||||
| SWC 10–20cm | 1.000 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, S.; Zhang, X.; Pumpanen, J.; Shen, G.; Xiong, F.; Liu, C. Seasonal Variation in Soil Greenhouse Gas Emissions at Three Age-Stages of Dawn Redwood (Metasequoia glyptostroboides) Stands in an Alluvial Island, Eastern China. Forests 2016, 7, 256. https://doi.org/10.3390/f7110256
Yin S, Zhang X, Pumpanen J, Shen G, Xiong F, Liu C. Seasonal Variation in Soil Greenhouse Gas Emissions at Three Age-Stages of Dawn Redwood (Metasequoia glyptostroboides) Stands in an Alluvial Island, Eastern China. Forests. 2016; 7(11):256. https://doi.org/10.3390/f7110256
Chicago/Turabian StyleYin, Shan, Xianxian Zhang, Jukka Pumpanen, Guangrong Shen, Feng Xiong, and Chunjiang Liu. 2016. "Seasonal Variation in Soil Greenhouse Gas Emissions at Three Age-Stages of Dawn Redwood (Metasequoia glyptostroboides) Stands in an Alluvial Island, Eastern China" Forests 7, no. 11: 256. https://doi.org/10.3390/f7110256