
Seasonal Variations of Carbon Dioxide, Water Vapor and Energy Fluxes in Tropical Indian Mangroves


Abstract
:1. Introduction
2. Materials and Methods
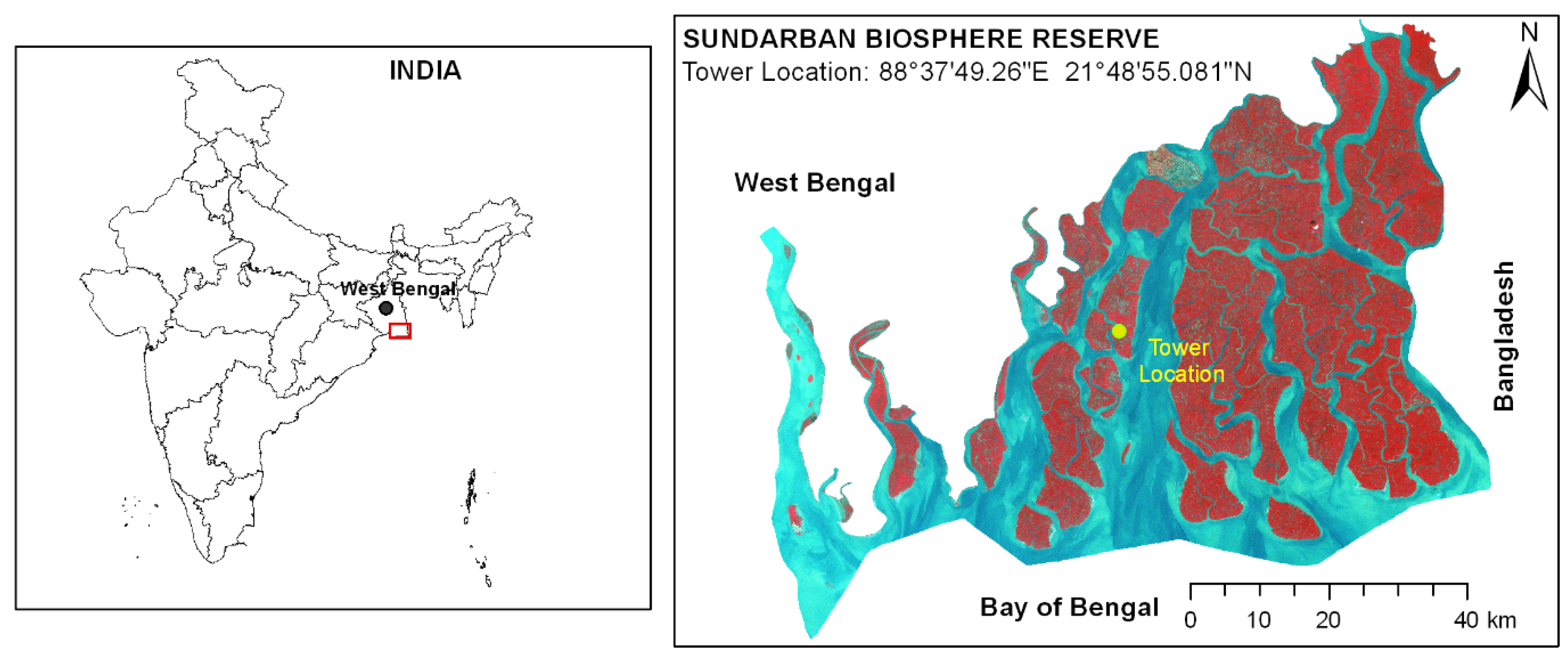
2.1. Study Area

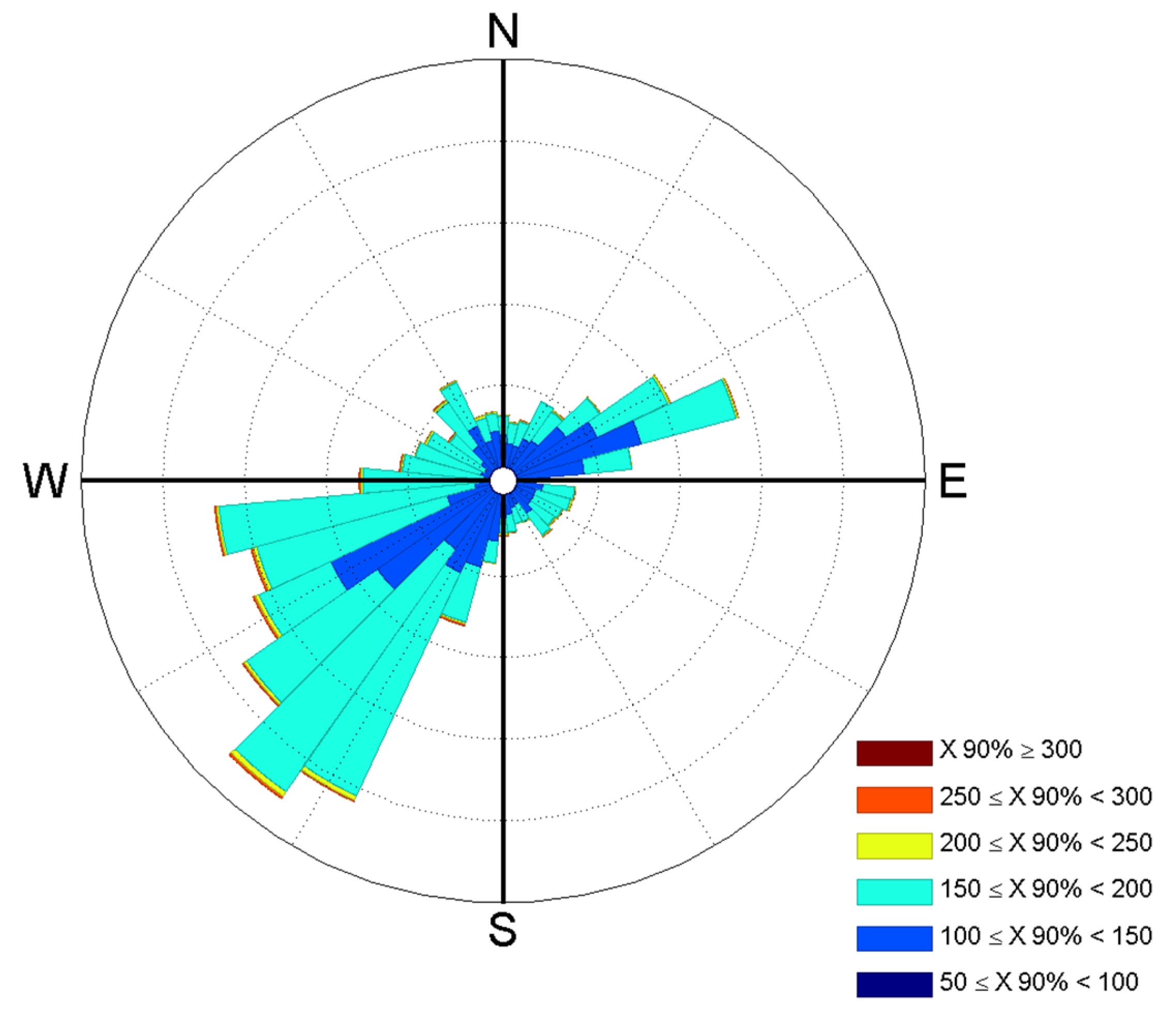
2.2. Eddy Covariance Flux Tower Setup

3. Flux Calculations and Corrections
3.1. Primary Raw Data Processing
3.2. Secondary Data Processing
3.2.1. CO2 Flux Spike Removal
3.2.2. LE and H Flux Spike Removal
3.2.3. Removal of Negative Nighttime CO2 Fluxes
3.2.4. Friction Velocity Correction
3.2.5. Gap-Filling and Uncertainty Analysis
3.3. Flux Partitioning
4. Results and Discussions
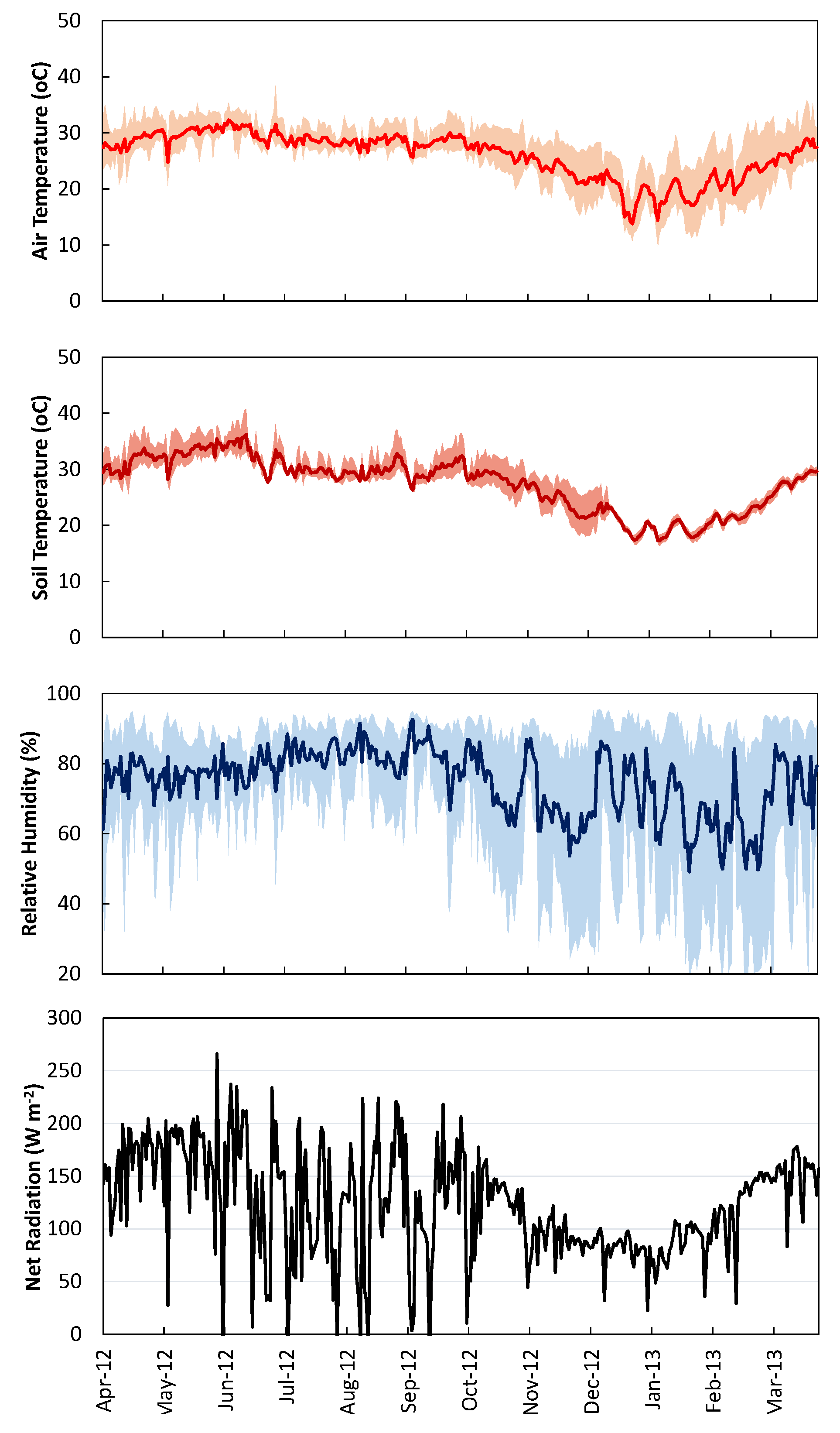
4.1. Meteorological Data


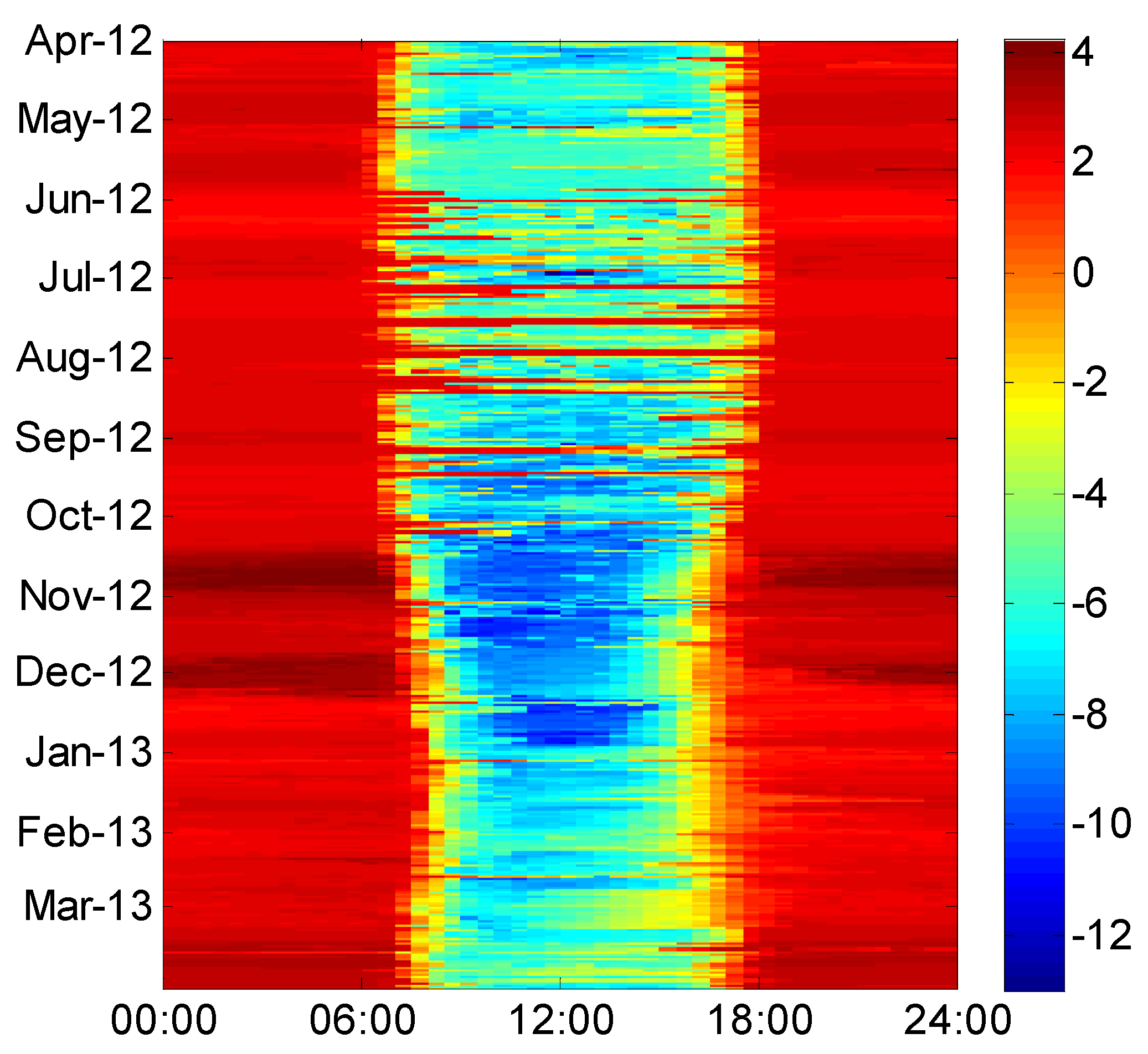
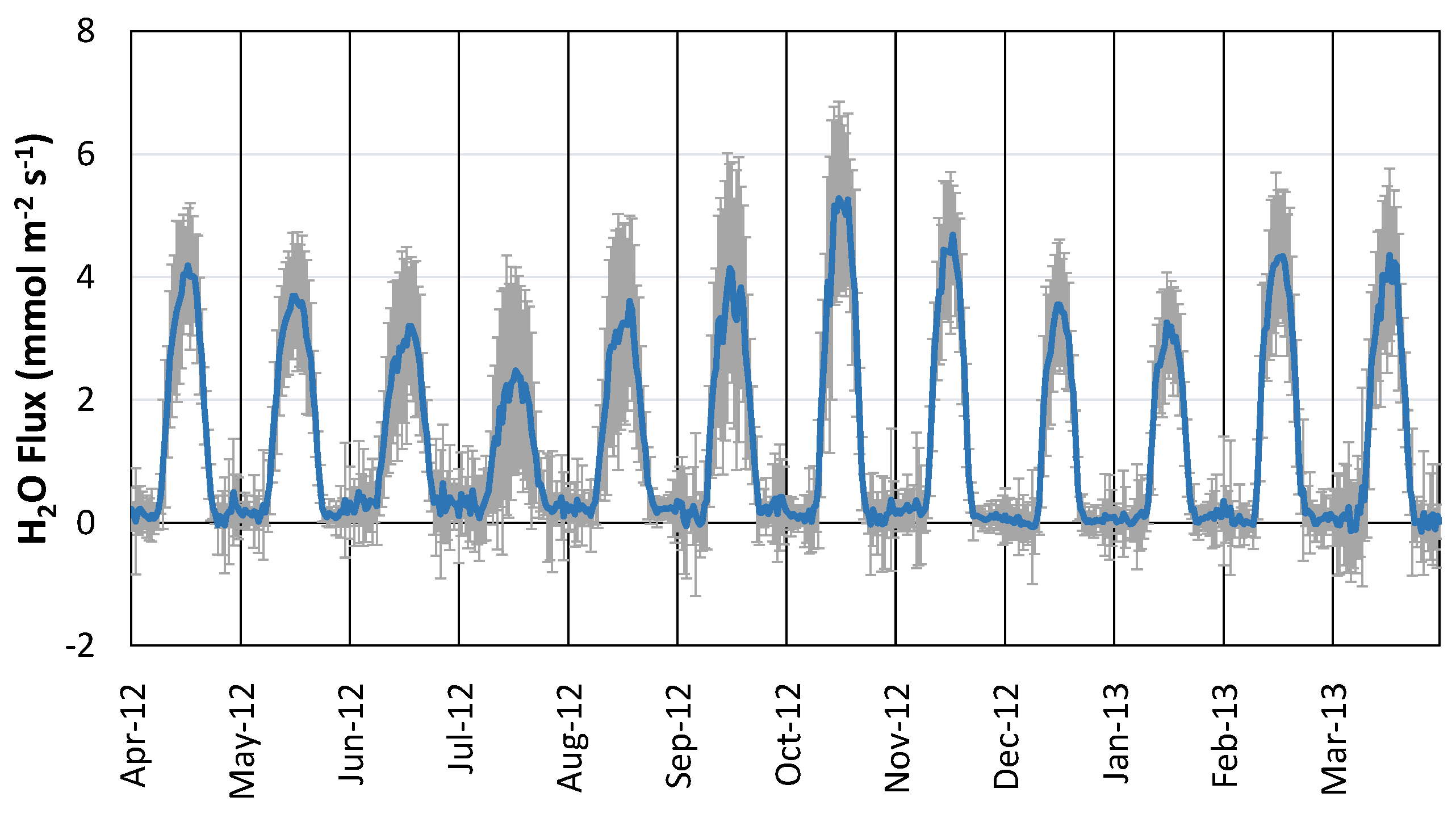
4.2. Variations in CO2 Flux and H2O Flux




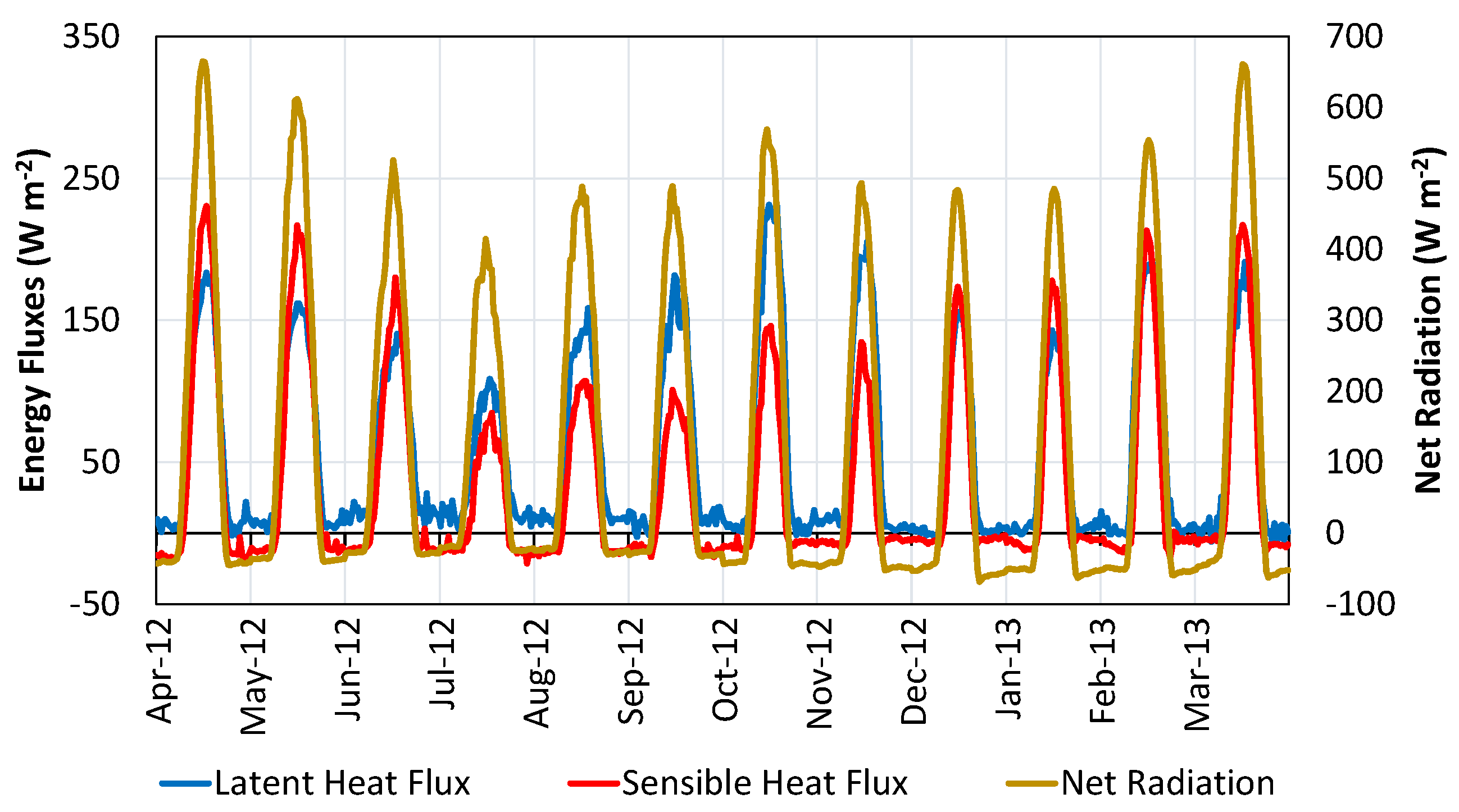
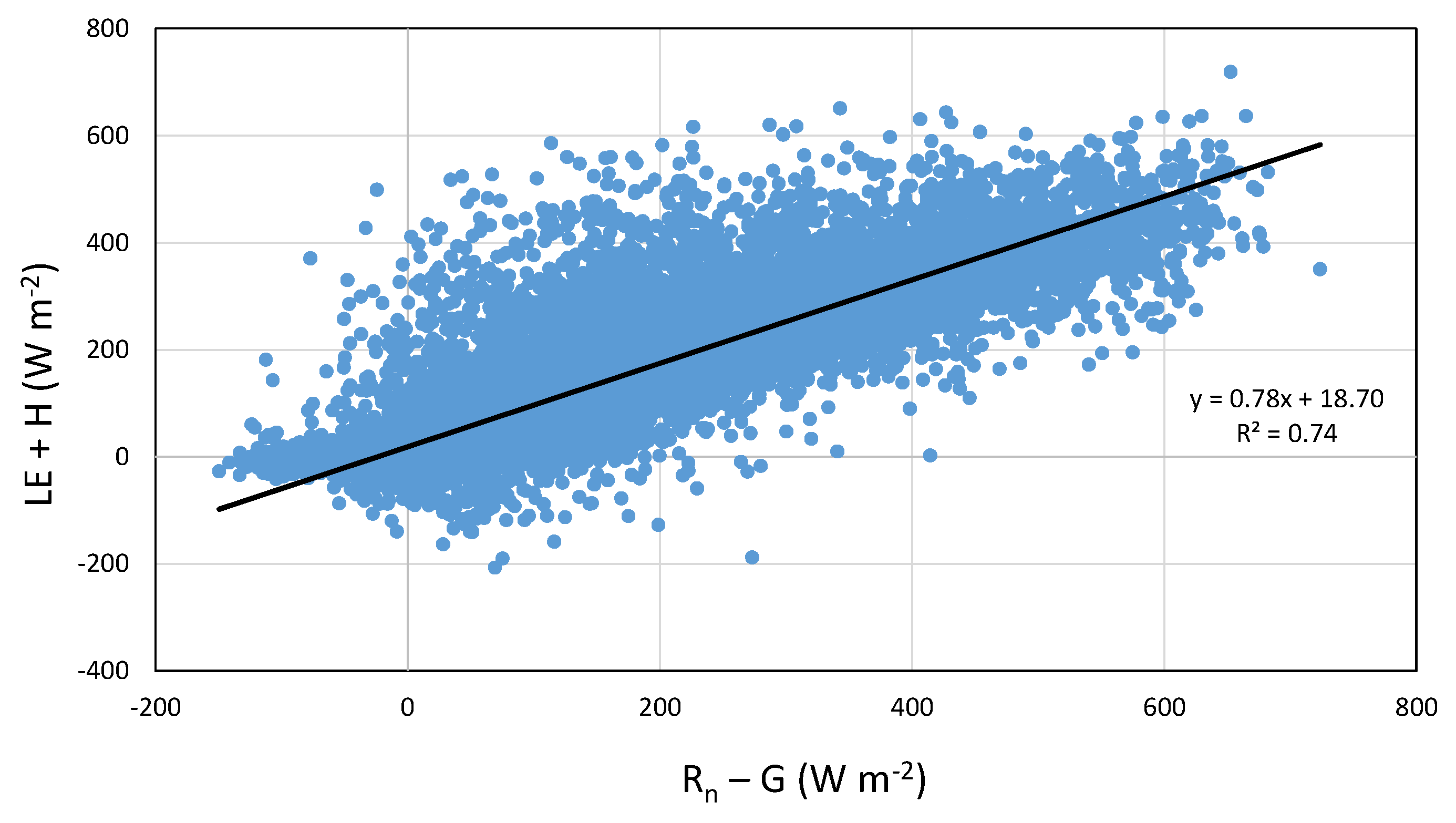
4.3. Heat Fluxes and Energy Balance


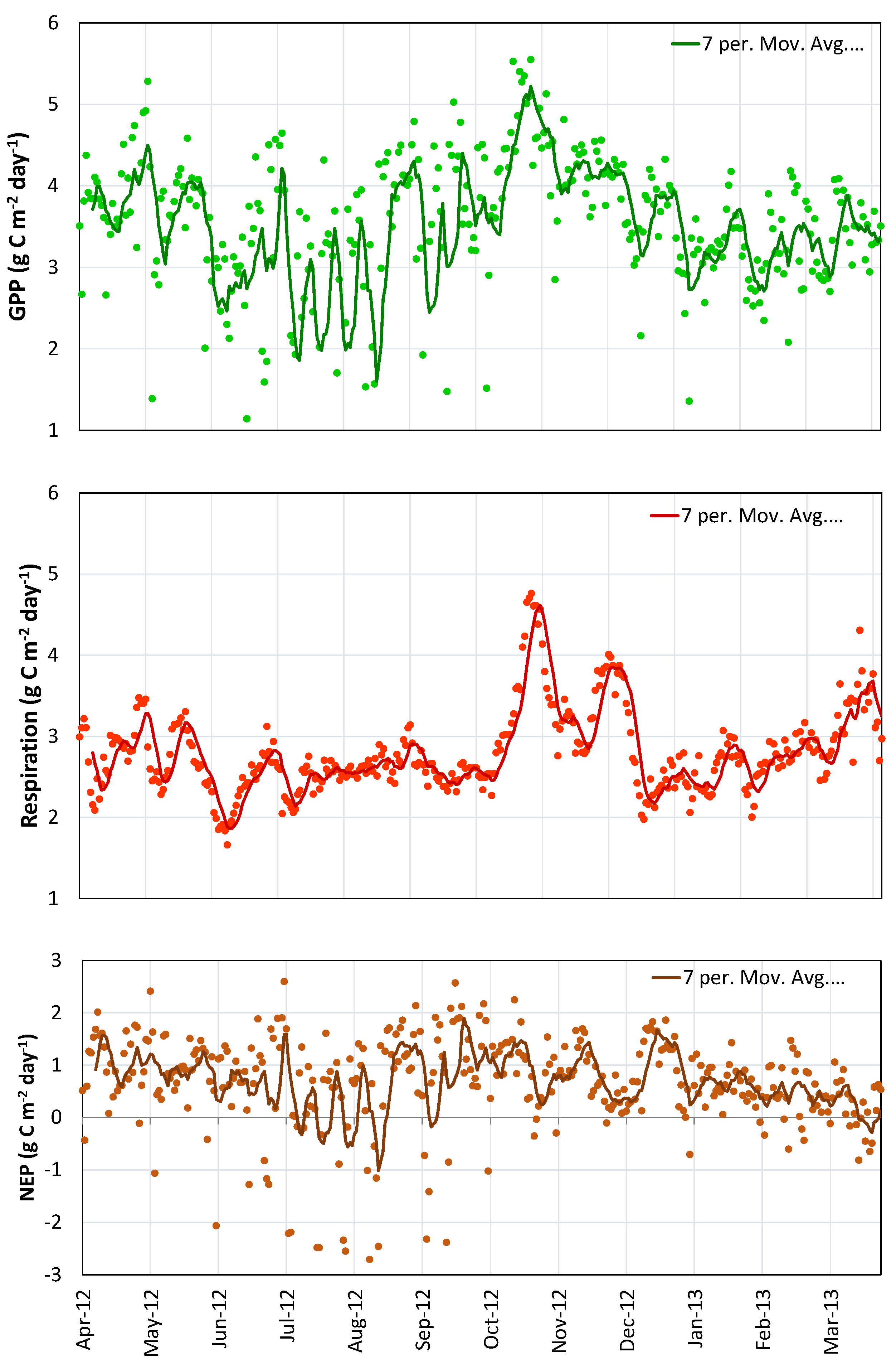
4.4. Net Carbon Sequestered


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimated Values | % Gap-Filled | |
|---|---|---|
| Bias Error (CO2 flux) | 0.13 ± 0.1 µmol·m−2·s−1 | |
| RMSE (CO2 flux) | 1.81 ± 0.24 µmol·m−2·s−1 | |
| Net Ecosystem Productivity (NEP) | 249 ± 20 g C m−2·year−1 | 35% |
| Gross Primary Productivity (GPP) | 1271 g C m−2·year−1 | 35% |
| Ecosystem Respiration (Reco) | 1022 g C m−2·year−1 | 35% |
| RMSE (H2O flux) | 0.73 ± 0.12 µmol·m−2·s−1 | |
| Evapotranspiration (ET) | 715 ± 120 mm·year−1 | 20% |
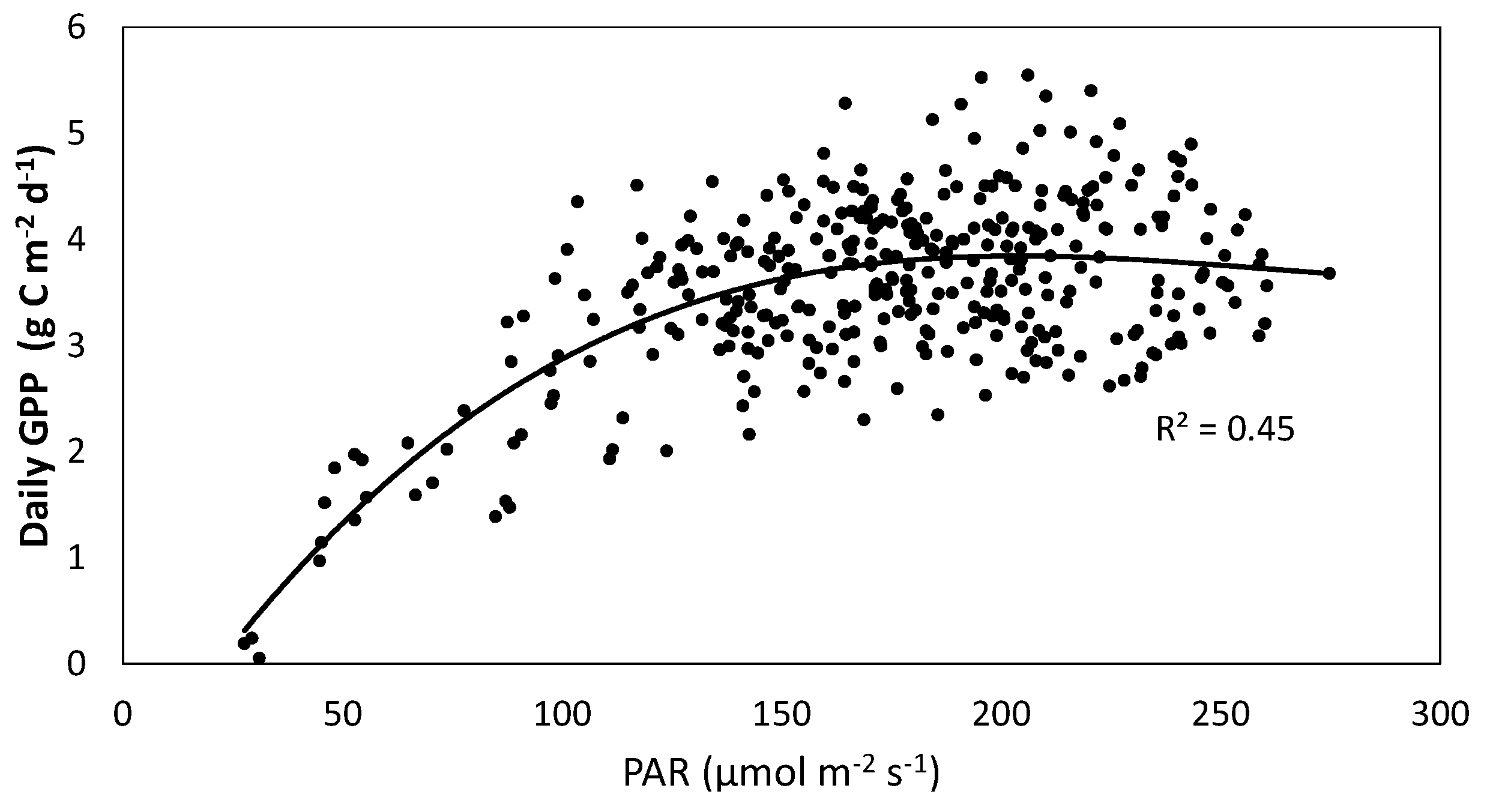
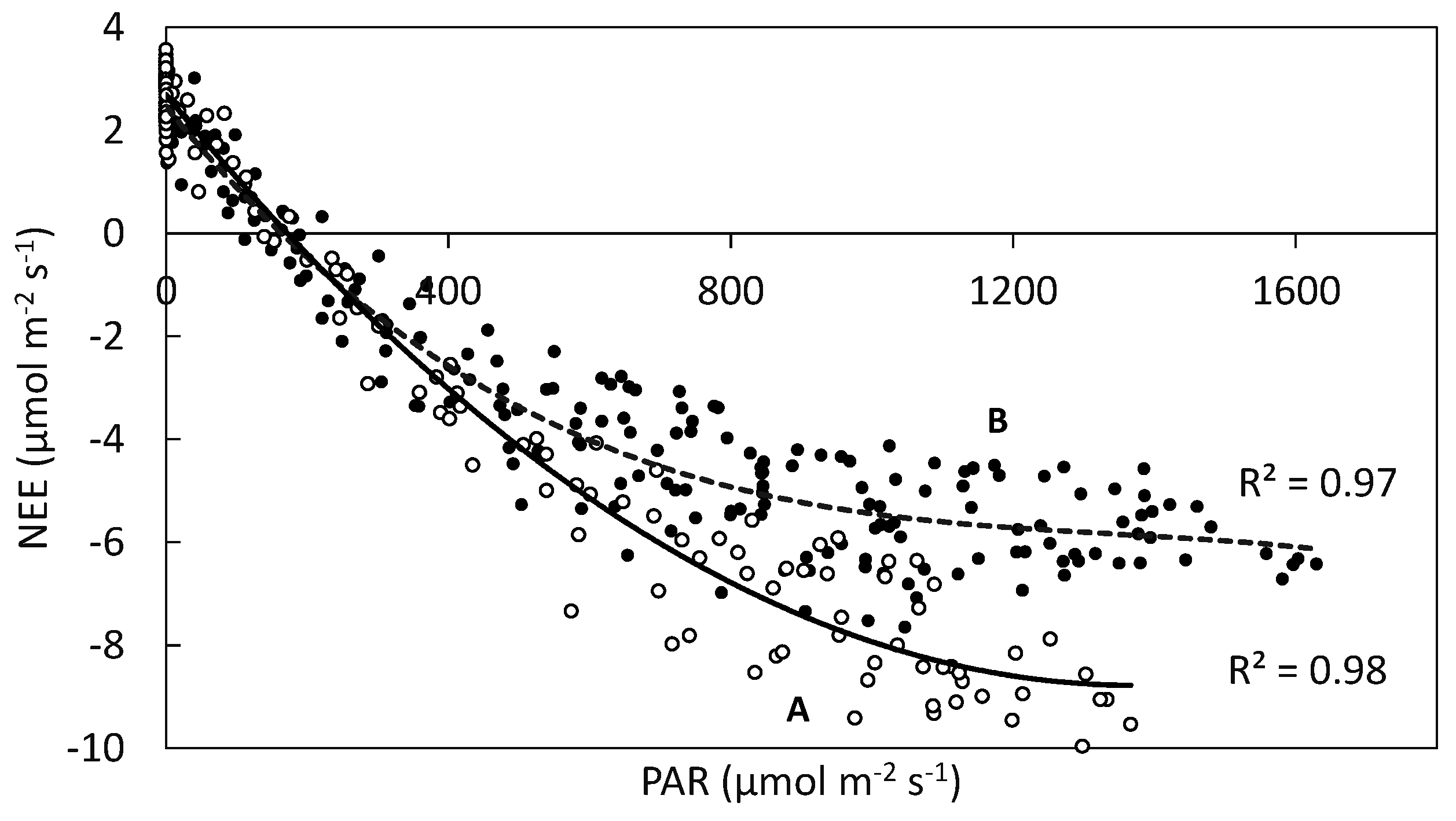
4.5. NEE and Photosynthetic Active Radiation (PAR)


| Month | Pmax (µmol CO2 m−2·s−1) | α (µmol CO2 µmol·photon−1) | Re (µmol CO2 m−2·s−1) | R2 |
|---|---|---|---|---|
| 12 April | 13.94 ± 0.45 | 0.0281 ± 0.0016 | 2.45 ± 0.08 | 0.86 |
| 12 May | 12.47 ± 0.55 | 0.0205 ± 0.0019 | 2.31 ± 0.16 | 0.71 |
| 12 June | 11.19 ± 0.33 | 0.0305 ± 0.0025 | 3.19 ± 0.12 | 0.77 |
| 12 July | 11.39 ± 0.26 | 0.0347 ± 0.0030 | 2.80 ± 0.17 | 0.76 |
| 12 August | 10.69 ± 0.23 | 0.0377 ± 0.0034 | 3.05 ± 0.18 | 0.75 |
| 12 September | 10.15 ± 0.28 | 0.0398 ± 0.0038 | 2.78 ± 0.16 | 0.68 |
| 12 October | 11.77 ± 0.42 | 0.0305 ± 0.0030 | 2.74 ± 0.19 | 0.68 |
| 12 November | 13.74 ± 0.35 | 0.0337 ± 0.0022 | 2.93 ± 0.13 | 0.84 |
| 12 December | 14.72 ± 0.40 | 0.0378 ± 0.0033 | 3.22 ± 0.22 | 0.75 |
| 13 January | 17.92 ± 0.55 | 0.0313 ± 0.0026 | 3.53 ± 0.24 | 0.78 |
| 13 February | 18.45 ± 0.68 | 0.0308 ± 0.0025 | 3.15 ± 0.22 | 0.79 |
| 13 March | 18.54 ± 0.84 | 0.0245 ±0.0015 | 2.42 ± 0.12 | 0.84 |
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. Intergovernmental Panel on Climate Change Working group Group I Contributions to IPCC Fifth Assessesment Report (AR5); Cambridge University Press: New York, NY, USA, 2013. [Google Scholar]
- Canadell, J.G.; le Quéré, C.; Raupach, M.R.; Field, C.B.; Buitenhuis, E.T.; Ciais, P.; Conway, T.J.; Gillett, N.P.; Houghton, R.A.; Marland, G. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proc. Natl. Acad. Sci. USA 2007, 104, 18866–18870. [Google Scholar] [CrossRef] [PubMed]
- Magnani, F.; Mencuccini, M.; Borghetti, M.; Berbigier, P.; Berninger, F.; Delzon, S.; Grelle, A.; Hari, P.; Jarvis, P.G.; Kolari, P.; et al. The human footprint in the carbon cycle of temperate and boreal forests. Nature 2007, 447, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Bouillon, S.; Borges, A.V.; Castaneda-Moya, E.; Diele, K.; Dittmar, T.; Duke, N.C.; Kristensen, E.; Lee, S.Y.; Marchand, C.; Middelburg, J.J.; et al. Mangrove production and carbon sinks: A revision of global budget estimates. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef] [Green Version]
- Chapman, V.J. Mangrove vegetation. Vaduz. J. Cramer 1976, 447–581. [Google Scholar]
- Tomlinson, P. The Botany of Mangroves; Cambridge University Press: Cambrige, United Kingdom, 1986. [Google Scholar]
- Robertson, A.I.; Alongi, D.M. Tropical Mangrove Ecosystems; American Geophysical Union: Washington, USA, 1992; Volume 41. [Google Scholar]
- Twilley, R.R. The exchange of organic carbon in basin mangrove forests in a southwest Florida estuary. Estuar. Coast. Shelf Sci. 1985, 20, 543–557. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Hincks, B.B.; Meyers, T.P. Measuring Biosphere-Atmosphere Exchanges of Biologically Related Gases with Micrometeorological Methods. Ecology 1988, 69, 1331–1340. [Google Scholar] [CrossRef]
- Lee, X.; Massman, W.J.; Law, B.E. Handbook of Micrometeorology: A Guide for Surface Flux Measurement and Analysis; Springer: New york, USA, 2006; Volume 29. [Google Scholar]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef]
- Verma, S.B. Micrometeorological methods for measuring surface fluxes of mass and energy. Remote Sens. Rev. 1990, 5, 99–115. [Google Scholar] [CrossRef]
- Baldocchi, D. TURNER REVIEW No. 15. “Breathing” of the terrestrial biosphere: Lessons learned from a global network of carbon dioxide flux measurement systems. Aust. J. Bot. 2008, 56, 1–26. [Google Scholar] [CrossRef]
- Wofsy, S.C.; Harris, R.C. The North American Carbon Program (NACP); Report of the NACP Committee of the U.S. Interagency Carbon Cycle Science Program: Washington, DC, USA, 2002. [Google Scholar]
- Barr, J.G.; Engel, V.; Fuentes, J.D.; Zieman, J.C.; O’Halloran, T.L.; Smith, T.J.; Anderson, G.H. Controls on mangrove forest-atmosphere carbon dioxide exchanges in western Everglades National Park. J. Geophys. Res. 2010, 115, G02020. [Google Scholar] [CrossRef]
- Food and Agriculture Organisation (FAO). The world’s mangroves 1980-2005. 2007. http://www.fao.org/docrep/010/a1427e/a1427e00.htm (accessed on 3 February 2016).
- Mukhopadhyay, S.K.; Jana, T.K.; De, T.K.; Sen, S. Measurement of exchange of CO2 in mangrove forest of Sunderbans using micrometeorological method. Trop. Ecol. 2000, 41, 57–60. [Google Scholar]
- Ganguly, D.; Dey, M.; Mandal, S.K.; De, T.K.; Jana, T.K. Energy dynamics and its implication to biosphere-atmosphere exchange of CO2, H2O and CH4 in a tropical mangrove forest canopy. Atmos. Environ. 2008, 42, 4172–4184. [Google Scholar] [CrossRef]
- Chanda, A.; Akhand, A.; Manna, S.; Dutta, S.; Hazra, S.; Das, I.; Dadhwal, V.K. Characterizing spatial and seasonal variability of carbon dioxide and water vapour fluxes above a tropical mixed mangrove forest canopy, India. J. Earth Syst. Sci. 2013, 122, 503–513. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Jha, C.S.; Rodda, S.R.; Thumaty, K.C.; Raha, A.K.; Dadhwal, V.K. Eddy covariance based methane flux in Sundarbans mangroves, India. J. Earth Syst. Sci. 2014, 123, 1089–1096. [Google Scholar] [CrossRef]
- Jha, C.S.; Thumaty, K.C.; Rodda, S.R.; Sonakia, A.; Dadhwal, V.K. Analysis of carbon dioxide, water vapour and energy fluxes over an Indian teak mixed deciduous forest for winter and summer months using eddy covariance technique. J. Earth Syst. Sci. 2013, 122, 1259–1268. [Google Scholar] [CrossRef]
- Watham, T.; Kushwaha, S.P.; Patel, N.R.; Dadhwal, V.K. Monitoring of carbon dioxide and water vapour exchange over a young mixed forest plantation using eddy covariance technique. Curr. Sci. 2014, 107, 858–866. [Google Scholar]
- Lloyd, C.R. Annual carbon balance of a managed wetland meadow in the Somerset Levels, UK. Agric. For. Meteorol. 2006, 138, 168–179. [Google Scholar] [CrossRef]
- Vickers, D.; Mahrt, L. Quality Control and Flux Sampling Problems for Tower and Aircraft Data. J. Atmos. Ocean. Technol. 1997, 14, 512–526. [Google Scholar] [CrossRef]
- Wilczak, J.M.; Oncley, S.P.; Stage, S.A. Sonic anemometer tilt correction algorithms. Bound.-Layer Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- Moncrieff, J.B.; Massheder, J.M.; de Bruin, H.; Elbers, J.; Friborg, T.; Heusinkveld, B.; Kabat, P.; Scott, S.; Soegaard, H.; Verhoef, A. A system to measure surface fluxes of momentum, sensible heat, water vapour and carbon dioxide. J. Hydrol. 1997, 188–189, 589–611. [Google Scholar] [CrossRef]
- Moncrieff, J.; Clement, R.; Finnigan, J.; Meyers, T. Averaging, detrending, and filtering of eddy covariance time series. In Handbook of Micrometeorology; Lee, X., Massman, W., Law, B., Eds.; Kluwer Academic: Dordrecht, Netherlands, 2004; pp. 7–31. [Google Scholar]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of flux measurements for density effects due to heat and water vapour transfer. Q. J. R. Meteorol. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Thomas, M.V.; Malhi, Y.; Fenn, K.M.; Fisher, J.B.; Morecroft, M.D.; Lloyd, C.R.; Taylor, M.E.; McNeil, D.D. Carbon dioxide fluxes over an ancient broadleaved deciduous woodland in southern England. Biogeosciences 2011, 8, 1595–1613. [Google Scholar] [CrossRef]
- Aubinet, M.; Feigenwinter, C.; Heinesch, B.; Bernhofer, C.; Canepa, E.; Lindroth, A.; Montagnani, L.; Rebmann, C.; Sedlak, P.; van Gorsel, E. Direct advection measurements do not help to solve the night-time CO2 closure problem: Evidence from three different forests. Agric. For. Meteorol. 2010, 150, 655–664. [Google Scholar] [CrossRef]
- Hirata, R.; Hirano, T.; Saigusa, N.; Fujinuma, Y.; Inukai, K.; Kitamori, Y.; Takahashi, Y.; Yamamoto, S. Seasonal and interannual variations in carbon dioxide exchange of a temperate larch forest. Agric. For. Meteorol. 2007, 147, 110–124. [Google Scholar] [CrossRef]
- Zhu, Z.; Sun, X.; Wen, X.; Zhou, Y.; Tian, J.; Yuan, G. Study on the processing method of nighttime CO2 eddy covariance flux data in ChinaFLUX. Sci. China Ser. D Earth Sci. 2006, 49, 36–46. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Moffat, A.M.; Papale, D.; Reichstein, M.; Hollinger, D.Y.; Richardson, A.D.; Barr, A.G.; Beckstein, C.; Braswell, B.H.; Churkina, G.; Desai, A.R.; et al. Comprehensive comparison of gap-filling techniques for eddy covariance net carbon fluxes. Agric. For. Meteorol. 2007, 147, 209–232. [Google Scholar] [CrossRef]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, H.; et al. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the Temperature Dependence of Soil Respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Department Biogeochemical Integration at the Max Planck Institute for Biogeochemistry. New REddyProcWeb Online Tool. https://www.bgc-jena.mpg.de/bgi/index.php/Services/REddyProcWeb (accessed on 2 Feburary 2016).
- Kljun, N.; Calanca, P.; Rotach, M.W.; Schmid, H.P. A simple parameterisation for flux footprint predictions. Bound.-Layer Meteorol. 2004, 112, 503–523. [Google Scholar] [CrossRef]
- Wilson, K.; Goldstein, A.; Falge, E.; Aubinet, M.; Baldocchi, D.; Berbigier, P.; Bernhofer, C.; Ceulemans, R.; Dolman, H.; Field, C.; et al. Energy balance closure at FLUXNET sites. Agric. For. Meteorol. 2002, 113, 223–243. [Google Scholar] [CrossRef]
- Goulden, M.L.; Munger, J.W.; Fan, S.-M.; Daube, B.C.; Wofsy, S.C. Measurements of carbon sequestration by long-term eddy covariance: Methods and a critical evaluation of accuracy. Glob. Chang. Biol. 1996, 2, 169–182. [Google Scholar] [CrossRef]
- Foken, T.; Oncley, S. Workshop on instrumental and methodical problems of land-surface flux measurements. Bull. Am. Meteorol. Soc. 1995, 76, 1191–1193. [Google Scholar]
- Baldocchi, D.; Falge, E.; Gu, L.H.; Olson, R.; Hollinger, D.; Running, S.; Anthoni, P.; Bernhofer, C.; Davis, K.; Evans, R.; et al. FLUXNET: A new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities. Bull. Am. Meteorol. Soc. 2001, 82, 2415–2434. [Google Scholar] [CrossRef]
- Hirata, R.; Saigusa, N.; Yamamoto, S.; Ohtani, Y.; Ide, R.; Asanuma, J.; Gamo, M.; Hirano, T.; Kondo, H.; Kosugi, Y.; et al. Spatial distribution of carbon balance in forest ecosystems across East Asia. Agric. For. Meteorol. 2008, 148, 761–775. [Google Scholar] [CrossRef]
- Luyssaert, S.; Inglima, I.; Jung, M.; Richardson, A.D.; Reichstein, M.; Papale, D.; Piao, S.L.; Schulze, E.D.; Wingate, L.; Matteucci, G.; et al. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Glob. Chang. Biol. 2007, 13, 2509–2537. [Google Scholar] [CrossRef]
- Joshi, H.G.; Ghose, M. Community structure, species diversity, and aboveground biomass of the Sundarbans mangrove swamps. Trop. Ecol. 2014, 55, 283–303. [Google Scholar]
- Simard, M.; Zhang, K.; Rivera-Monroy, V.H.; Ross, M.S.; Ruiz, P.L.; Castañeda-Moya, E.; Twilley, R.R.; Rodriguez, E. Mapping height and biomass of mangrove forests in Everglades National Park with SRTM elevation data. Photogramm. Eng. Remote Sens. 2006, 72, 299–311. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Verma, S.B.; Sims, P.L.; Meyers, T.P.; Bradford, J.A.; Burba, G.G.; Suyker, A.E. Gross primary production and light response parameters of four Southern Plains ecosystems estimated using long-term CO2-flux tower measurements. Glob. Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef]
- Ruimy, A.; Jarvis, P.G.; Baldocchi, D.D.; Saugier, B. CO2 fluxes over plant canopies and solar radiation: A review. Adv. Ecol. Res. 1995. [Google Scholar] [CrossRef]
- Ehleringer, J.; Björkman, O. Quantum yields for CO2 uptake in C3 and C4 plants dependence on temperature, CO2, and O2 concentration. Plant Physiol. 1977, 59, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Maher, D.T.; Santos, I.R.; Golsby-Smith, L.; Gleeson, J.; Eyre, B.D. Groundwater-derived dissolved inorganic and organic carbon exports from a mangrove tidal creek: The missing mangrove carbon sink? Limnol. Oceanogr. 2013, 58, 475–488. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodda, S.R.; Thumaty, K.C.; Jha, C.S.; Dadhwal, V.K. Seasonal Variations of Carbon Dioxide, Water Vapor and Energy Fluxes in Tropical Indian Mangroves. Forests 2016, 7, 35. https://doi.org/10.3390/f7020035
Rodda SR, Thumaty KC, Jha CS, Dadhwal VK. Seasonal Variations of Carbon Dioxide, Water Vapor and Energy Fluxes in Tropical Indian Mangroves. Forests. 2016; 7(2):35. https://doi.org/10.3390/f7020035
Chicago/Turabian StyleRodda, Suraj Reddy, Kiran Chand Thumaty, Chandra Shekhar Jha, and Vinay Kumar Dadhwal. 2016. "Seasonal Variations of Carbon Dioxide, Water Vapor and Energy Fluxes in Tropical Indian Mangroves" Forests 7, no. 2: 35. https://doi.org/10.3390/f7020035