
Short-Term Belowground Responses to Thinning and Burning Treatments in Southwestern Ponderosa Pine Forests of the USA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
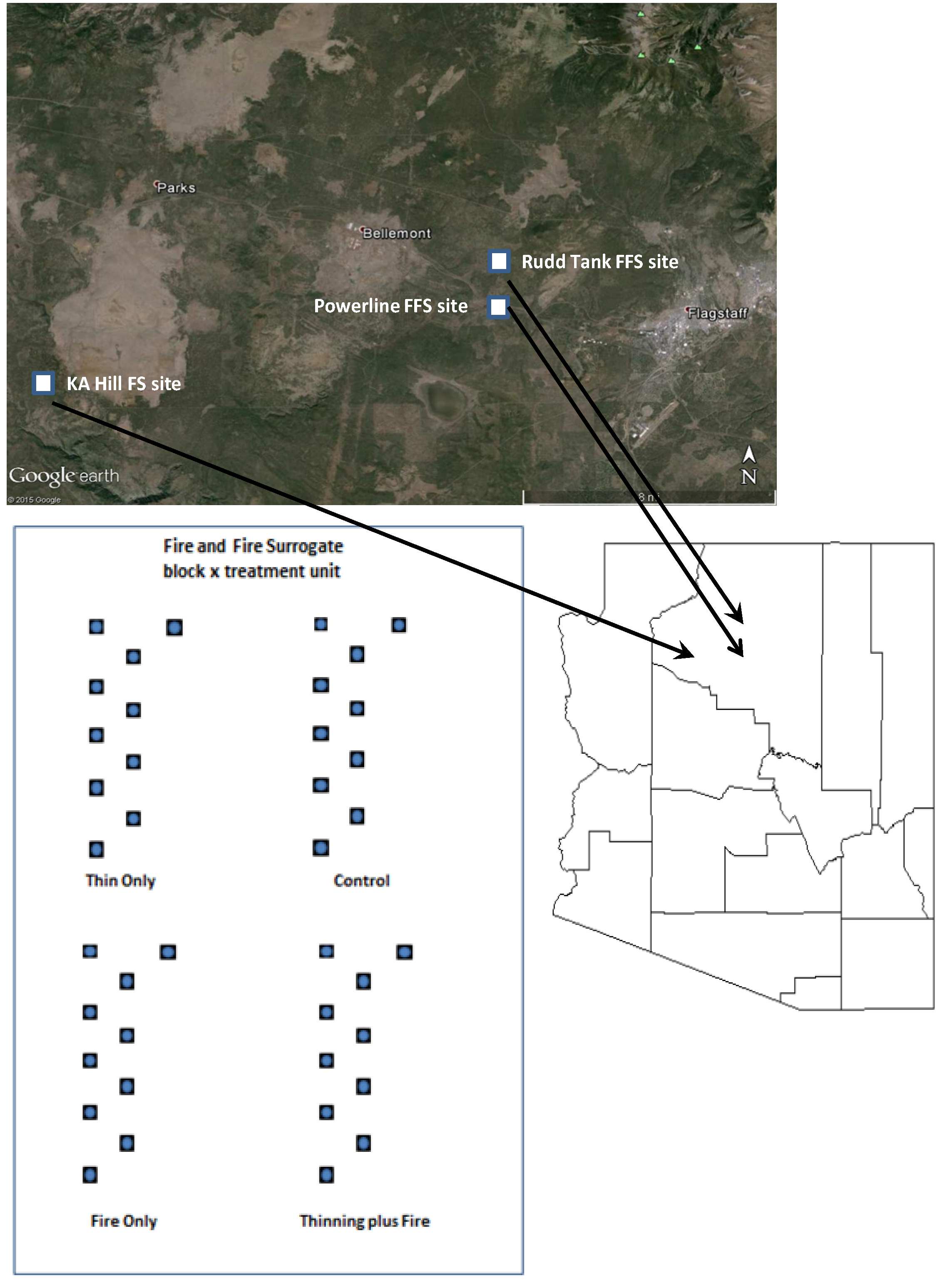
2.1 Study Site and Experimental Design
2.2 Soil Sampling and Nutrient Analyses
2.3 Microbial Community Analyses
2.4 Microbial Activity and Net N Transformations
2.5 Statistical methods
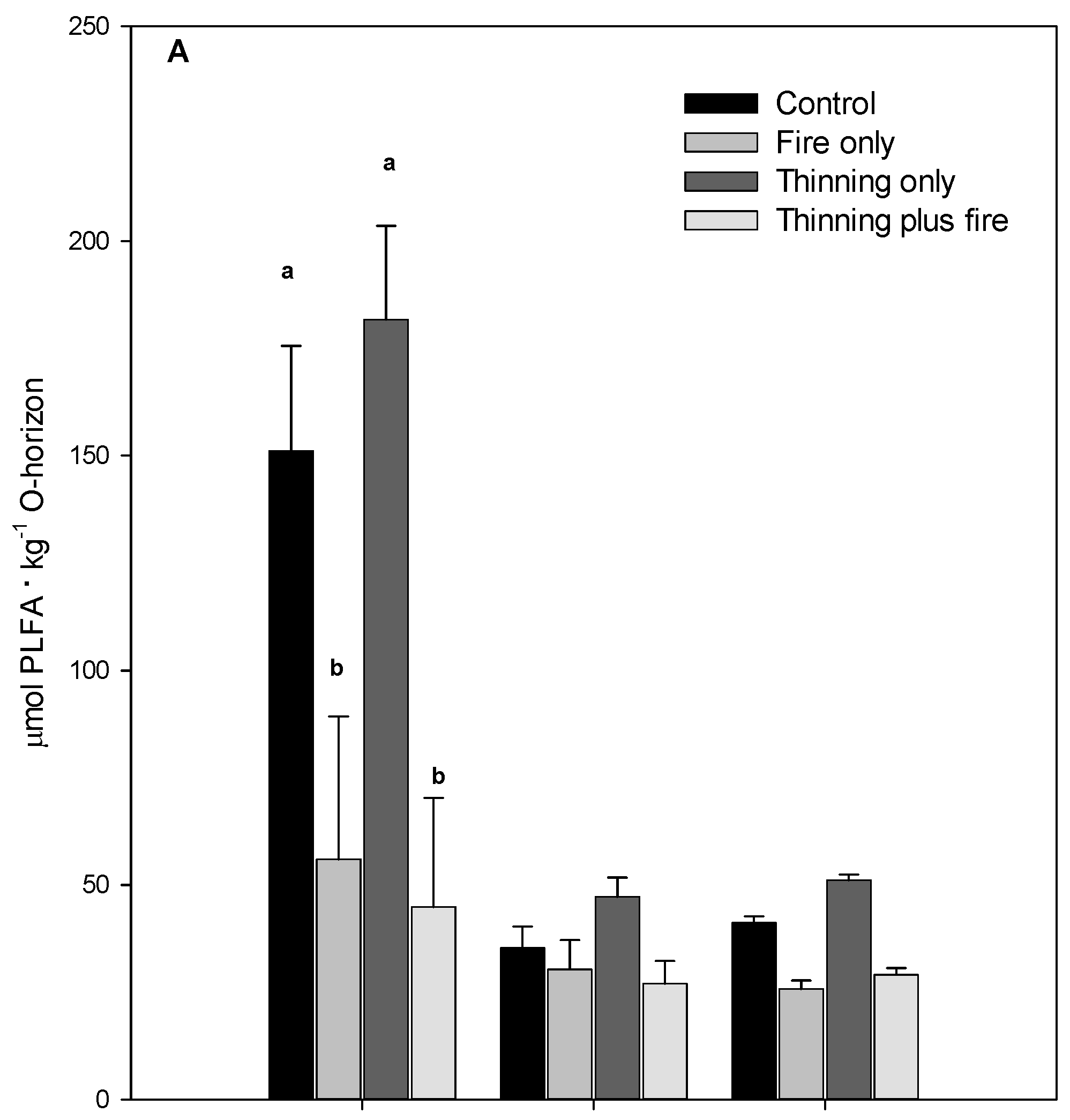
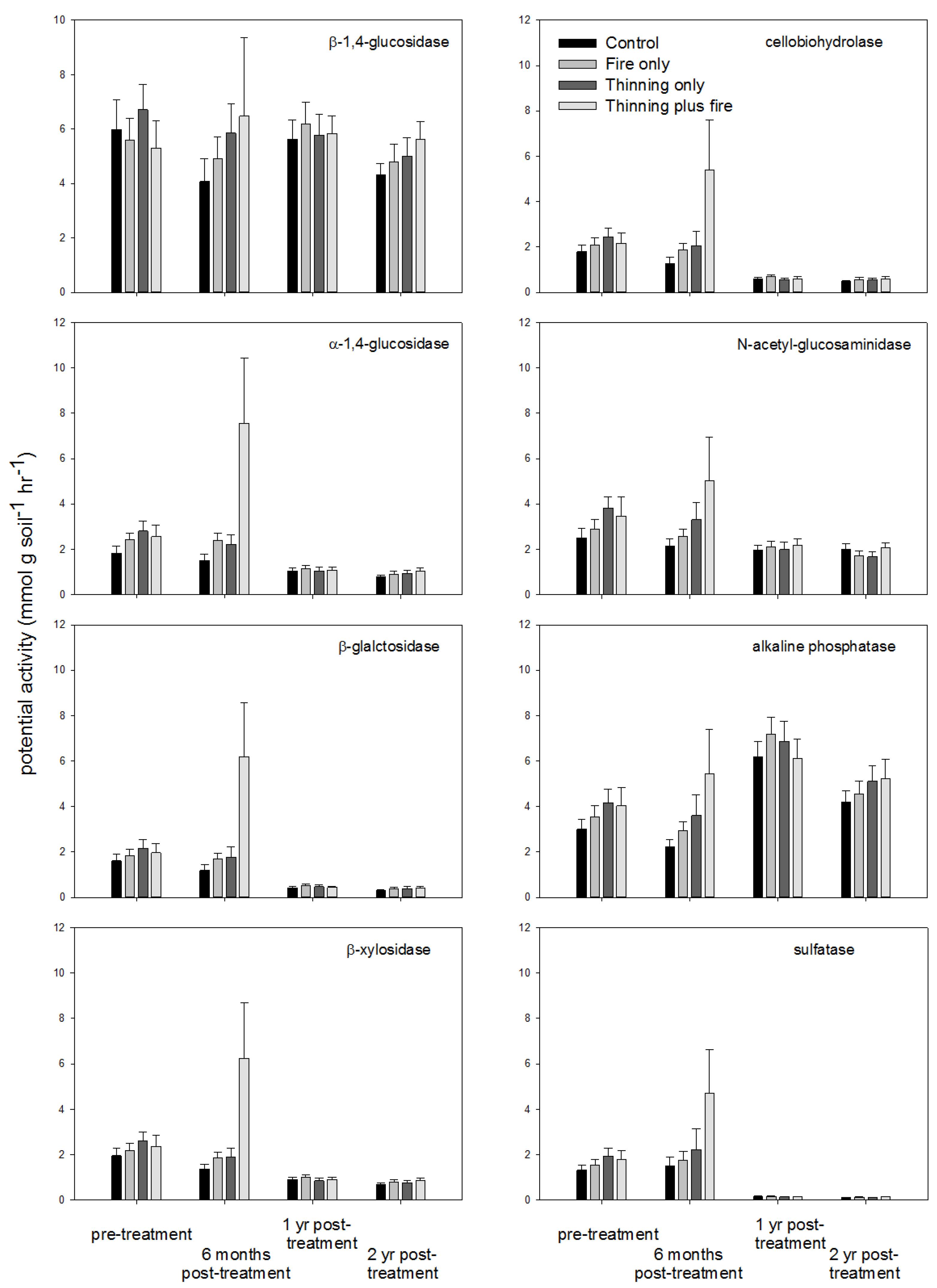
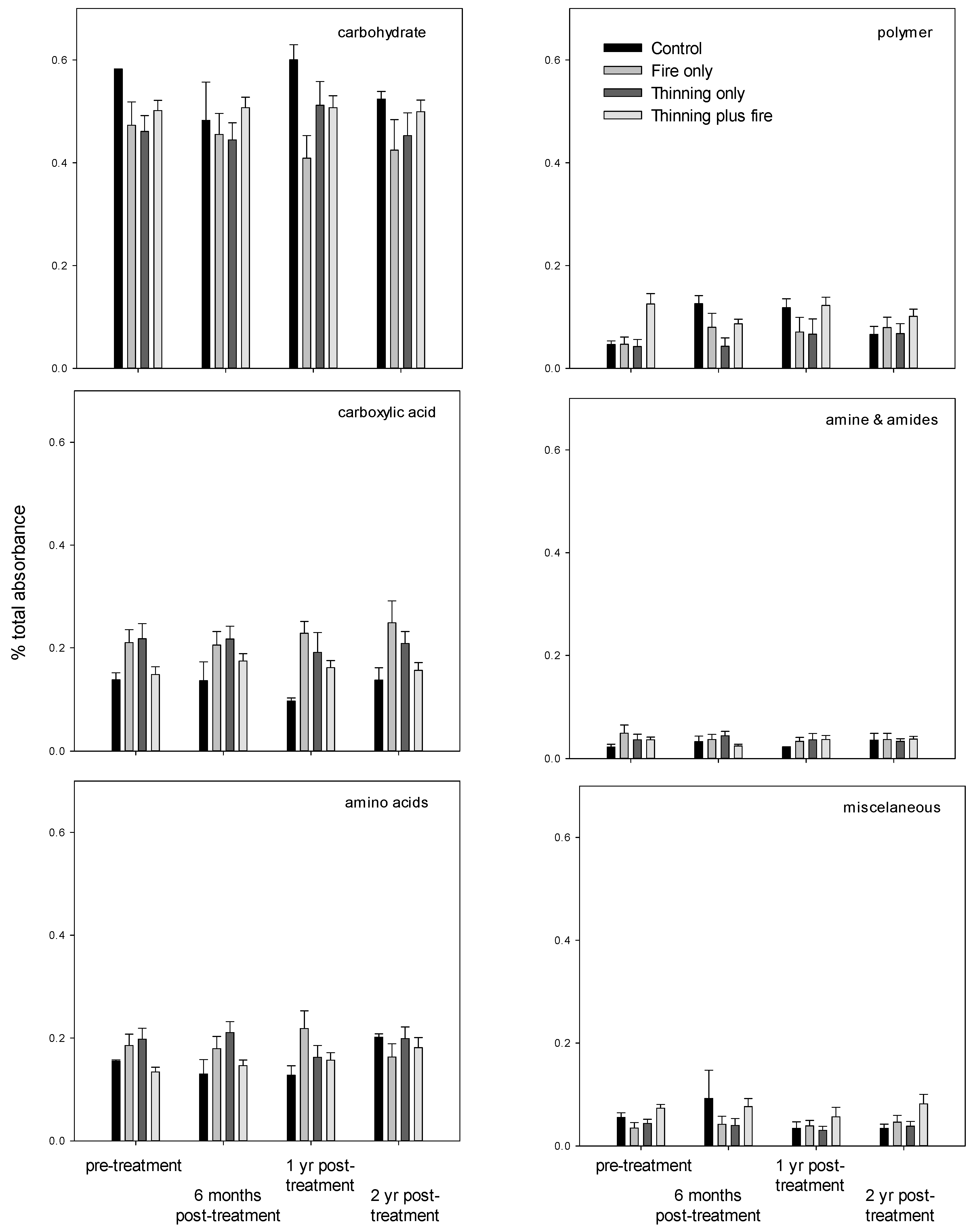
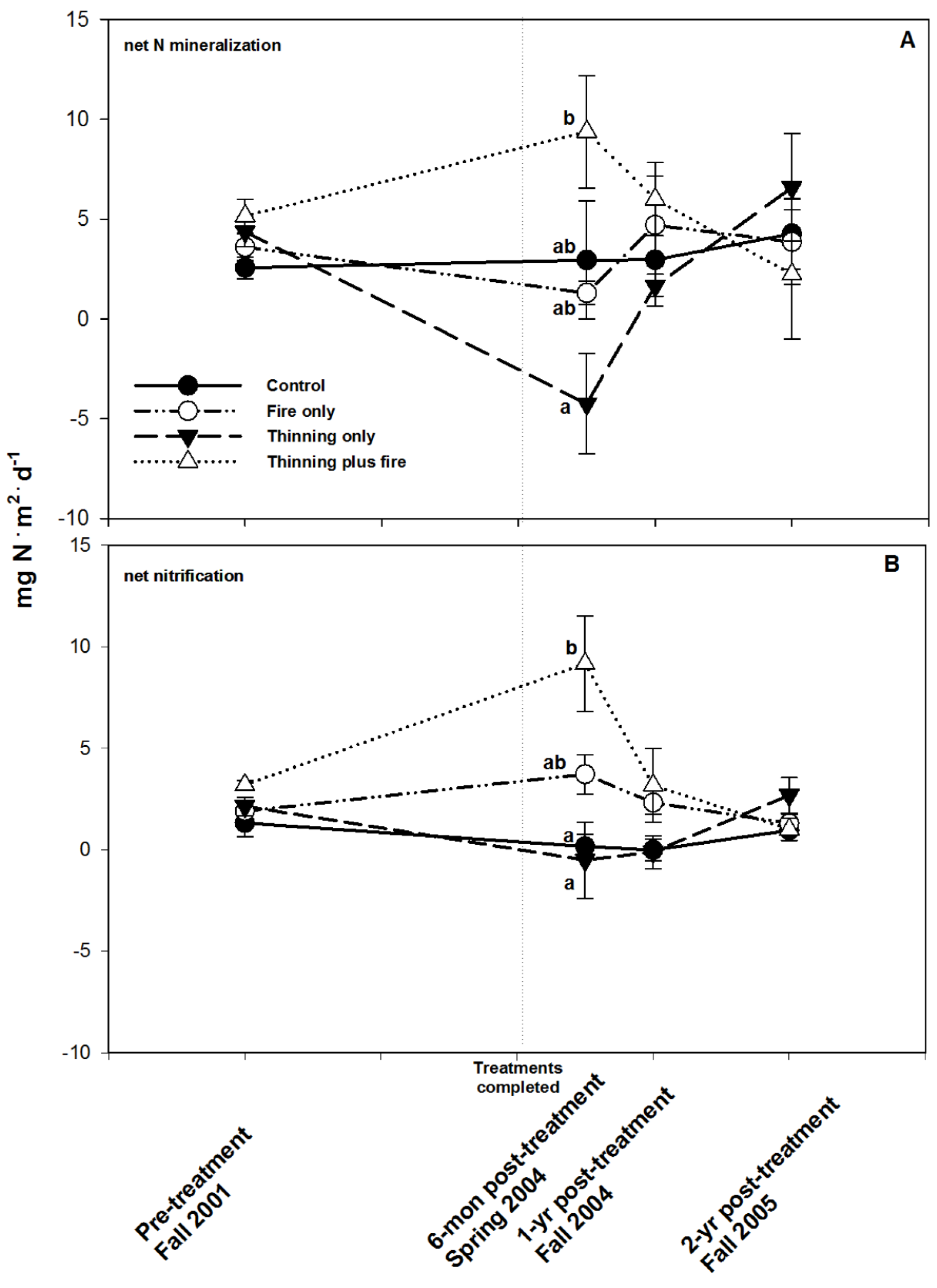
3. Results
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Covington, W.W.; Fulé, P.; Moore, M.M.; Hart, S.C.; Kolb, T.; Mast, J.; Sackett, S.S.; Wagner, M. Restoring ecosystem health in ponderosa pine forests of the Southwest. J. For. 1997, 95, 23–29. [Google Scholar]
- Fulé, P.Z.; Covington, W.W.; Moore, M.M. Determining reference conditions for ecosystem management of Southwestern ponderosa pine forests. Ecol. Appl. 1997, 7, 895–908. [Google Scholar] [CrossRef]
- White, A.S. Presettlement regeneration patterns in a Southwestern ponderosa pine stand. Ecology 1985, 66, 589–594. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Baisan, C.H. Historical fire regime patterns in the Southwestern United States since AD 1700. In Fire Effects in Southwestern Forests; Allen, C.D., Ed.; U.S. Department of Agriculture, Forest Service: Fort Collins, CO, USA, 1996; pp. 11–32. [Google Scholar]
- O’Brian, R.A. Arizona’s Forest Resources, 1999; U.S. Department of Agriculture, Forest Service: Ogden, UT, USA, 2002.
- Kaye, J.P.; Hart, S.C.; Fulé, P.Z.; Covington, W.W.; Moore, M.M.; Kaye, M.W. Initial carbon, nitrogen, and phosphorus fluxes following ponderosa pine restoration treatments. Ecol. Appl. 2005, 15, 1581–1593. [Google Scholar] [CrossRef]
- Bakker, J.D.; Moore, M.M. Controls on vegetation structure in Southwestern ponderosa pine forests, 1941 and 2004. Ecology 2007, 88, 2305–2319. [Google Scholar] [CrossRef] [PubMed]
- Pearson, G.A. A twenty-year record of changes in an Arizona ponderosa pine forest. Ecology 1933, 14, 272–285. [Google Scholar] [CrossRef]
- Savage, M.A.; Brown, P.M.; Feddema, J. The role of climate in a pine forest regeneration pulse in the southwestern United States. Ecoscience 1996, 3, 310–318. [Google Scholar]
- Covington, W.W.; Moore, M.M. Southwestern ponderosa forest structure. Changes since Euro-American settlement. J. For. 1994, 92, 39–47. [Google Scholar]
- Mast, J.N.; Fulé, P.Z.; Moore, M.M.; Covington, W.W.; Waltz, A.E.M. Restoration of presettlement age structure of an Arizona ponderosa pine forest. Ecol. Appl. 1999, 9, 228–239. [Google Scholar] [CrossRef]
- Pollet, J.; Omi, P.N. Effect of thinning and prescribed burning on crown fire severity in ponderosa pine forests. Int. J. Wildland Fire 2002, 11, 1–10. [Google Scholar] [CrossRef]
- Strom, R.A.; Fulé, P.Z. Pre-wildfire fuel treatments affect long-term ponderosa pine forest dynamics. Int. J. Wildland Fire 2007, 16, 128–138. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Giai, C.; Huang, J.; Miesel, J.R. Initial effect of fire and mechanical thinning on soil enzyme activity and nitrogen transformations in eight North American forest ecosystems. Soil Biol. Biochem. 2008, 40, 3076–3085. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Huang, J.; Hart, S.C. Fire, thinning, and the carbon economy, effects of the FFS treatments on carbon storage and sequestration rate. For. Ecol. Manag. 2008, 255, 3081–3097. [Google Scholar] [CrossRef]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Kobziar, L.N.; McBride, J.R.; Stephens, S.L. The efficacy of fire and fuels reduction treatments in a Sierra Nevada pine plantation. Int. J. Wildland Fire 2009, 18, 791–801. [Google Scholar] [CrossRef]
- DeBano, L.F.; Neary, D.G.; Ffolliott, P.F. Fire’s Effects on Ecosystems; John Wiley & Sons: New York, NY, USA, 1998; p. 333. [Google Scholar]
- Bardgett, R.D. The Biology of Soil, A Community and Ecosystem Approach; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Covington, W.W.; Sackett, S.S. Soil mineral nitrogen changes following prescribed burning in ponderosa pine. For. Ecol. Manag. 1992, 54, 18–191. [Google Scholar] [CrossRef]
- Kaye, J.P.; SHart, C. Ecological restoration alters nitrogen transformations in a ponderosa pine-bunchgrass ecosystem. Ecol. Appl. 1998, 8, 1052–1060. [Google Scholar]
- Frey, S.D.; Knorr, M.; Parrent, J.L.; Simpson, R.T. Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests. For. Ecol. Manag. 2004, 196, 159–171. [Google Scholar] [CrossRef]
- Hart, S.C.; DeLuca, T.H.; Newman, G.S.; MacKenzie, D.M.; Boyle, S.I. Post-fire vegetative dynamics as drivers of microbial community structure and function in forest soils. For. Ecol. Manag. 2005, 220, 166–184. [Google Scholar] [CrossRef]
- Simonin, K.; TKolb, E.; Montes-Helu, M.; Koch, G.W. The influence of thinning on components of stand water balance in a ponderosa pine forest stand during and after extreme drought. Agric. For. Meteorol. 2007, 143, 266–276. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Fritze, H. Clear-cutting and prescribed burning in coniferous forest, comparison of effects on soil fungal and total microbial biomass, respiration activity and nitrification. Soil Biol. Biochem. 1995, 27, 101–109. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Kiikkilä, O.; Fritze, H. Charcoal as a habitat for microbes and its effect on the microbial community underlying humus. Oikos 2000, 89, 231–242. [Google Scholar] [CrossRef]
- Cairney, J.W.G.; Bastias, B.A. Influences of fire on forest soil fungal communities. Can. J. For. Res. 2007, 37, 207–215. [Google Scholar] [CrossRef]
- Waldrop, M.P.; McColl, J.G.; Powers, R.F. Effects of forest postharvest management practices on enzyme activities in decomposing litter. Soil Sci. Soc. Am. J. 2003, 67, 1250–1256. [Google Scholar] [CrossRef]
- Grayston, S.J.; Renneberg, H. Assessing effects of forest management on microbial community structure in a central European beech forest. Can. J. For. Res. 2006, 40, 1901–1907. [Google Scholar] [CrossRef]
- Kaye, J.P.; Hart, S.C. Restoration and canopy-type effects on soil respiration in a ponderosa pine-bunchgrass ecosystem. Soil Sci. Soc. Am. J. 1998, 62, 1062–1072. [Google Scholar] [CrossRef]
- Grady, K.C.; Hart, S.C. Influences of thinning, prescribed burning, and wildfire on soil processes and properties in southwestern ponderosa pine forests, A retrospective study. For. Ecol. Manag. 2006, 234, 123–135. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Zouhar, K.L. Effects of selection harvest and prescribed fire on the soil nitrogen status of ponderosa pine forests. For. Ecol. Manag. 2000, 138, 263–271. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils, a review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- McIver, J.D.; Erickson, K.; Youngblood, A. Principal Short-Term Finding of the National Fire and Fire Surrogate Study; U.S. Department of Agriculture, Forest Service: Portland, OR, USA, 2012.
- Stephens, S.L.; Moghaddas, J.J.; Edminster, C.; Fiedler, C.E.; Haase, S.; Harrington, M.; Keeley, J.E.; Knapp, E.E.; McIver, J.D.; Metlen, K.; et al. Fire treatment effects on vegetation structure, fuels, and potential fire severity in western U.S. forest. Ecol. Appl. 2009, 19, 305–320. [Google Scholar] [CrossRef] [PubMed]
- McIver, J.D.; Weatherspoon, P.; Edminster, C. A Long-Term Study on the Effects of Alternative Ponderosa Pine Restoration Treatments. Ponderosa Pine Ecosystems Restoration and Conservation, Steps Toward Stewardship; U.S. Department of Agriculture, Forest Service: Ogden, UT, USA, 2001.
- Stephens, S.L.; McIver, J.D.; Boerner, R.E.J.; Fettig, C.J.; Fontaine, J.B.; Hartsough, B.R.; Kennedy, P.; Schwilk, D.W. Effects of forest fuel reduction treatments in the United States. BioScience 2012, 62, 549–560. [Google Scholar]
- Faiella, S.M.; Bailey, J.D. Fluctuations in fuel moisture across restoration treatments in semi-arid ponderosa pine forests of northern Arizona, USA. Int. J. Wildland Fire 2007, 16, 119–127. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture. Terrestrial Ecosystem Survey of the Coconino National Forest; U.S. Department of Agriculture, Forest Service, Southwestern Region: Albuquerque, NM, USA, 1995.
- U.S. Department of Agriculture. Terrestrial Ecosystem Survey of the Kaibab National Forest; U.S. Department of Agriculture, Forest Service, Southwestern Region: Albuquerque, NM, USA, 1991.
- Schwilk, D.W.; Keeley, J.E.; Knapp, E.E.; McIver, J.; Bailey, J.D.; Fettig, C.J.; Fiedler, C.E.; Harrod, R.J.; Moghaddas, J.J.; Outcalt, K.W.; et al. The national Fire and Fire Surrogate study, effects of fuel reduction methods on forest vegetation structure and fuels. Ecol. Appl. 2009, 19, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Youngblood, A. Thinning and burning in dry coniferous forests of the western United State, effectiveness in altering diameter distributions. For. Sci. 2010, 56, 46–59. [Google Scholar]
- Hendershot, W.H.; Lalande, H.; Duquette, M. Soil reaction and exchangeable acidity. In Soil Sampling and Methods of Analysis; Carter, M.R., Ed.; Canadian Society of Soil Science, Lewis Publishers: Boca Raton, FL, USA, 1993; pp. 141–145. [Google Scholar]
- Hassett, J.E.; Zak, D.R. Aspen harvest intensity decreases microbial biomass, extracellular enzyme activity, and soil nitrogen cycling. Soil Sci. Soc. Am. J. 2005, 69, 227–235. [Google Scholar] [CrossRef]
- Leckie, S. Methods of microbial community profiling and their application to forest soils. For. Ecol. Manag. 2005, 220, 88–106. [Google Scholar] [CrossRef]
- Frostegard, A.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Zelles, L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterization of microbial communities in soil, A review. Biol. Fertil. Soils 1999, 29, 111–129. [Google Scholar] [CrossRef]
- White, D.C.; Davis, W.M.; Nickels, J.S.; King, J.C.; Bobbie, R.J. Determination of the sedimentary microbial biomass by extractable lipid phosphate. Oecologia 1979, 40, 51–62. [Google Scholar] [CrossRef]
- Frostegard, A.; Tunlid, A.; Bååth, E. Microbial biomass measured as total lipid phosphate in soils of different organic contents. J. Microbiol. Methods 1991, 14, 151–163. [Google Scholar] [CrossRef]
- Schweitzer, J.A.; Bailey, J.K.; Fischer, D.G.; LeRoy, C.J.; Lonsdorf, E.V.; Whitham, T.G.; Hart, S.C. Soil microorganism—Plant interactions; heritable relationship between plant genotype and associated microorganisms. Ecology 2008, 89, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Eivazi, F.; Tabatabai, M.A. Phosphatases in soils. Soil Biol. Biochem. 1977, 9, 162–172. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Enzymatic analysis of microbial pattern and process. Biol. Fertil. Soils 1994, 17, 69–74. [Google Scholar] [CrossRef]
- Eivazi, F.; Bayan, M. Effects of long-term prescribed burning on the activity of select soil enzymes in an oak-hickory forest. Can. J. For. Res. 1996, 26, 1799–1804. [Google Scholar] [CrossRef]
- Olander, L.P.; Vitousek, P.M. Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 2000, 49, 18–191. [Google Scholar] [CrossRef]
- Ganeshamurthy, A.N.; Nielsen, N.E. Arylsulfatase and the biochemical mineralization of soil organic sulfur. Soil Biol. Biochem. 1990, 22, 1163–1165. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Antibus, R.K.; Linkins, A.E. An enzymatic approach to the analysis of microbial activity during plant litter decomposition. Agric. Ecosyst. Environ. 1991, 34, 43–54. [Google Scholar] [CrossRef]
- Classen, A.T.; Boyle, S.I.; Haskins, K.; Overby, S.; Hart, S.C. Community-level physiological profiles of bacteria and fungi, plate type and incubation temperature influences on contrasting soils. FEMS Microbiol. Ecol. 2003, 44, 319–328. [Google Scholar] [CrossRef]
- Garland, J.; Mills, A. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source-utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [PubMed]
- Kennedy, A.C. Carbon utilization and fatty acid profiles for characterization of bacteria. In Methods of Soil Analysis, Part 2 Microbiological and Biochemical Properties; SSSA Book Series No. 5; Weaver, R.W., Mickelson, S.H., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 548–551. [Google Scholar]
- Dobranic, J.K.; Zak, J.C. A microtiter plate procedure for evaluating functional diversity. Mycologia 1999, 91, 756–765. [Google Scholar] [CrossRef]
- Hart, S.C.; Stark, J.M.; Davidson, E.A.; Firestone, M.K. Nitrogen mineralization, immobilization, and nitrification. In Methods of Soil Analysis, Part 2 Microbiological and Biochemical Properties; SSSA Book Series No. 5; Weaver, R.W., Mickelson, S.H., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 985–1018. [Google Scholar]
- Lachat Instruments, Inc. QuickChem Method No. 12-107-06-1-B; Lachat Instruments, Inc.: Milwaukee, WI, USA, 2001. [Google Scholar]
- Lachat Instruments, Inc. QuickChem Method No. 10-107-04-1-C; Lachat Instruments, Inc.: Milwaukee, WI, USA, 2000. [Google Scholar]
- Schabenberger, O. Introducing the GLIMMIX procedure for generalized linear mixed models. In Proceedings of the Thirtieth Annual SAS® Users Group International Conference, Cary, NC, USA, 26–29 April 2005.
- Kenward, M.G.; Roger, J.H. Small Sample Inference for Fixed Effects from Restricted Maximum Likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S. Generalized linear mixed models, a practical guide for ecology and evolution. Trend Ecol. Evol. 2008, 2, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Mielke, P.W., Jr.; Berry, K.J. Permutation Methods, A Distance Function Approach; Springer-Verlag: New York, NY, USA, 2001. [Google Scholar]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Petrondas, D.A.; Gabriel, K.R. Multiple comparisons by rerandomization tests. J. Am. Stat. Assoc. 1983, 78, 949–957. [Google Scholar] [CrossRef]
- Milar, L.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future, managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Sala, A. Frequent fire alters nitrogen transformations in ponderosa pine stands of the Inland Northwest. Ecology 2006, 87, 2511–2522. [Google Scholar] [CrossRef]
- Wright, R.J.; Hart, S.C. Nitrogen and phosphorus status in a ponderosa pine forest after 20 years of interval burning. Ecoscience 1997, 4, 526–533. [Google Scholar]
- Boyle, S.I.; Hart, S.C.; Kaye, J.P.; Waldrop, M.P. Restoration and canopy type influence soil microflora in a ponderosa pine forest. Soil Sci. Soc. Am. J. 2005, 69, 1627–1638. [Google Scholar] [CrossRef]
- Maassen, S.; Fritze, F.; Wirth, S. Response of soil microbial biomass, activities, and community structure at a pine stand in northeastern Germany 5 years after thinning. Can. J. For. Res. 2006, 36, 1427–1434. [Google Scholar] [CrossRef]
- Siira-Pietikäinen, A.; Pietikäinen, J.; Fritze, H.; Haimi, J. Short-term responses of soil decomposer communities to forest management, clear felling versus alternative forest harvesting methods. Can. J. For. Res. 2001, 31, 88–99. [Google Scholar] [CrossRef]
- Hannam, K.D.; Quideau, S.A.; Kishcuk, B.E. Forest floor microbial communities in relation to stand composition and timber harvesting northern Alberta. Soil Biol. Biochem. 2006, 38, 2565–2575. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Tikka, P.J.; Valkonen, S.; Isomäki, A.; Fritze, H. Is the soil microbial community related to the basal area of trees in a Scots pine stand? Soil Biol. Biochem. 2007, 39, 1832–1834. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Huang, J.; Hart, S.C. Impacts of fire and fire surrogate treatments on forest soil properties, a meta-analytical approach. Ecol. Appl. 2009, 19, 338–358. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage, meta-analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]







© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Overby, S.T.; Hart, S.C. Short-Term Belowground Responses to Thinning and Burning Treatments in Southwestern Ponderosa Pine Forests of the USA. Forests 2016, 7, 45. https://doi.org/10.3390/f7020045
Overby ST, Hart SC. Short-Term Belowground Responses to Thinning and Burning Treatments in Southwestern Ponderosa Pine Forests of the USA. Forests. 2016; 7(2):45. https://doi.org/10.3390/f7020045
Chicago/Turabian StyleOverby, Steven T., and Stephen C. Hart. 2016. "Short-Term Belowground Responses to Thinning and Burning Treatments in Southwestern Ponderosa Pine Forests of the USA" Forests 7, no. 2: 45. https://doi.org/10.3390/f7020045