Tree Species Selection in the Face of Drought Risk—Uncertainty in Forest Planning


Northwest German Forest Research Institute, Grätzelstr. 2, D-37079 Göttingen, Germany
*
Author to whom correspondence should be addressed.
Forests 2017, 8(10), 363; https://doi.org/10.3390/f8100363
Submission received: 21 August 2017
/
Revised: 20 September 2017
/
Accepted: 23 September 2017
/
Published: 26 September 2017
Abstract
:Rapid climate change leads to significant shifts in the site-productivity relationship of tree species and alters abiotic and biotic risks well beyond classical rotation ages on many forest sites worldwide. Forest conversion may be an adequate measure to counter possible negative effects of climate change. Unfortunately, climate-driven changes in abiotic and biotic risks bear a significant source of intrinsic uncertainty inherent in climate projections. It is our goal to appraise uncertainty in species selection under drought stress, one of the most important risk factors for many forests. We derive a method to assess drought restrictions and demonstrate the uncertainty in the process of species selection by applying three climate scenarios. Furthermore, we interpret the consequences of climate uncertainty in the light of different management goals, i.e., a business-as-usual silviculture, a climate protection strategy favoring CO2 sequestration and a biodiversity strategy increasing diversity. The methods are applied to two representative regions in the North German Plain. The results clearly show the strong need for adaptive planning when drought restrictions are considered. However, different silvicultural management objectives may alter the extent of adaptive planning. The uncertainty in the planning process arising from different underlying climate projections strongly depends on the regional site characteristics and on forest management strategy. In conclusion, it is most important in forest planning to clearly state the management goals and to carefully explore if the goals can be met under climate change and if the uncertainty due to climate projections significantly affects the results of species selection.
1. Introduction
Strategic forest planning decisions such as species selection in regeneration are strongly impaired by changing climate conditions because of their long-term commitment. Climate change alters the growing conditions of forests well beyond a stand’s lifespan [1]. In central Europe, projected rising temperatures, the most likely seasonal shift in precipitation and an accumulation of extreme weather events such as droughts, torrential rain and storms [2,3,4] will change tree growth and will have a considerable impact on survival rate (e.g., [5,6,7,8]). Ultimately, stand productivity and thus forest enterprise economics will be affected (e.g., [9,10,11,12]).
Forest conversion is seen as one adaptive measure to counter or at least diminish the negative effects of climate change (e.g., [13,14]). Formerly, good forestry practice based species selection on constant site characteristics, i.e., soil nutrients and site water budget. Under climate change, site water budget is no longer constant but varies according to altered climatic water balances, i.e., the difference between precipitation and evapotranspiration. Consequently, forest planning now has to consider the dynamics in site water budget. This, in fact, poses a new challenge. While forest planning always had to decide on a basis of incomplete information, projections of climate change bear an intrinsic uncertainty, where climate scenarios cannot be assigned any probability of occurrence [15], thus termed ‘deep uncertainty’. The uncertainty in climate projections when used in forest planning raises two concerns, i.e., to build confidence in the projections [16,17] and to communicate the degree of uncertainty for proposed recommendations [18]. Several studies point out methods of how to account for deep uncertainty (e.g., [19,20]). In order to successfully adapt forest ecosystems and forest management practices to climate change uncertainties, robust decision making approaches are promoted [21]. Furthermore, sensitivity analysis should be applied to examine how robust a decision is to adapt to climate change [22]. Our approach to deal with climate uncertainty in this study is straightforward as we use a set of three climate projections to set up a range of potential climate developments to assess their impact on planning decisions.
This study on tree species selection under climate change considering drought risk puts the above-mentioned methods and procedures into practice for two forest regions in the North German Plain. Most importantly, we present an approach suitable for area-wide forest planning encompassing climate projections, their uncertainty, stand type planning rule-sets and drought risk assessment. Risk in general is defined as the probability of expected loss or damage due to a particular hazard for a given area and reference period [23]. Drought risk, in particular, is the reduction in forest productivity, loss of vitality and eventually even mortality induced by temporal shifts towards dryer site conditions (e.g., [24,25,26,27]). Already nowadays and even more under future climate conditions, water availability limits species selection on many sites in the North German Plain [28,29]. Generally, the direct effect of drought on trees is hard to quantify (e.g., [30,31]). Thus, we account for drought vulnerability, i.e., the degree to which forests are susceptible to negative impacts of drought [32], rather than quantifying actual risk, when assessing future species’ site suitability.
This study pursues three goals: (1) to illustrate the options in stand type planning based on site characteristics and implemented silvicultural management strategy and to point out the alternatives in adaptive planning under drought restrictions; (2) to quantify the uncertainty in the planning process arising from different climate projections; and (3) to demonstrate the consequences of climate uncertainty for forest planning based on the findings in two typical regions, representative for larger growth zones in the North German Plain, i.e., whether the choice of one particular climate scenario is necessary as it strongly affects the stand type planning or if the differences between stand type planning under the different climate scenarios are acceptably small to desist from considering climate uncertainty.
As the results are based on forest inventory data, with three underlying management strategies and three climate projections in two representative regions, this study allows for detailed analysis and specific conclusions. We draw conclusions on the interaction between management scenario and climate projection and its consequence for forest planning under uncertainty. This fosters the learning process and aids forest managers to cope with uncertainty.
2. Materials and Methods
This study investigates how different climate scenarios and silvicultural management strategies influence species selection in forest planning. Therefore, the combinations of species occurrence and site characteristics in two representative regions are identified. The site-specific foundations, i.e., soil and climate properties, are presented and the rule-set for species selection considering drought restrictions in regard to different management strategies is introduced.
2.1. Forest Data
The primary research question, to account for uncertainty in forest planning, is examined based on forest stand data in two regions, Uelzen with a total of 48,682 ha of forests (33.5% forest cover) and Fläming with 90,903 ha of forests (42.3% forest cover), in the North German Plain (Figure 1). Both regions are representative in regard to site characteristics and forest structure for larger growth zones in the North German Plain.
In each region a representative number of stands was sampled from all forest stands with available inventory data (n = 999 in Uelzen and n = 1008 in Fläming). These two study regions were selected, because they comprise different starting conditions concerning species occurrence (Table 1), regional climate and soil conditions (Table 2).
In both regions Scots pine is the dominant species but the spectrum in Uelzen is much more diverse with considerable proportions of Norway spruce, other broadleaves, pedunculate and sessile oak, European beech and Douglas-fir. In Fläming only beech, oak and other broadleaves contribute considerably.
The climate values indicate slightly warmer and much dryer conditions in Fläming compared to Uelzen. Especially in the growing season the water deficit, expressed by the climatic water balance, is obvious in Fläming although the mean available soil moisture is marginally larger than in Uelzen.
2.2. Climate Scenarios
We characterize the range of potential future climate from 2011 to 2070 using three climate projections all based on the emission scenario RCP 8.5 [33,34]. The Potsdam Institute for Climate Impact Research created three regional scale climate projections using the regional climate model STARS (Statistical Analogue Resampling Scheme) [35] by downscaling the global circulation models INM-CM4 [36], ECHAM6 [37] and ACCESS1.0 [38]. Although under debate [39,40] the climate model STARS is still recommended for climate impact assessments (e.g., [41]). Climate parameters for all 1218 weather stations of the German Weather Service from 2011 to 2070 were obtained in a daily resolution. The basic climate parameters used in this research are precipitation sum in the growing season and mean temperature in the growing season. Regarding the projected annual mean temperature, the projections predict a 1.1 °C increase for the North German Plain comparing current values (mean in period 1991 to 2010) with values in period 2051 to 2070 under the INM-CM4 run while ECHAM6 projects a 1.6 °C increase and ACCESS1.0 even a 2.7 °C increase. With regard to the pre-industrial age an additional increase of 0.8 °C until 2010 needs to be added resulting in an overall increase of approx. 1.9 °C to 3.5 °C. Throughout this study these three climate projections are labelled minimum, median and maximum climate run.
The projected daily mean temperature and precipitation sum of all climate stations within a 20-km distance of the respective location are interpolated to obtain climate values for each selected stand using WaSiM-ETH’s distance-weighted regression model [42,43].
The key climate variable in the process of selecting species in forest planning is site water budget. Site water budget is defined according to [44], i.e., the sum of climatic water balance in the growing season (difference of precipitation and potential evapotranspiration for grass reference [45]) and the available soil moisture as defined by [46]. The start of the growing season is defined according to the model LNVAR by [47] (p. 52) and the ending is determined applying suggestions by [48,49,50], i.e., either the temperature or the short day criterion, whichever is met first.
Figure 2 displays the development as mean values of all selected sites in Uelzen and Fläming under minimum, median and maximum climate run.
Obviously, in Uelzen throughout time and across climate scenarios, site water budget values are on a higher level than in Fläming. Annual fluctuations are generally higher than trends over time. Both in Uelzen and Fläming under the minimum run the trend line indicates hardly any deviation from the current situation. The median run clearly shows a negative trend to much dryer conditions in both regions at the end of the projection period. Even worse is the decrease in site water budget under the maximum climate scenario, which projects values as low as approximately −250 mm in Uelzen and −400 mm in Fläming in 2070.
2.3. Soil Data
Decisions on site suitability of species in this context rely, first of all, on soil nutrients and site water budget. For each selected stand the necessary variables, soil nutrients and available soil moisture (asm) calculated for 1 m soil depth, are assessed using the soil map of scale 1:1 Mio [51]. Soil nutrients are organized in five classes according to the information of the soil map and model functions by [52].
A heatmap gives a summary of the proportions of sites according to soil nutrients and site water budget (Figure 3).
Currently, the majority of sites in Uelzen (65%) exhibit a medium soil nutrient status and a positive site water budget up to 100 mm (Figure 3, top heatmaps). A considerable proportion of sites (12%) are rich in nutrients and well water supplied with more than 100 mm. In Fläming soil nutrients and site water budget are generally much lower under status quo conditions. Almost all sites are characterized by a negative site water budget in the growing season. In the category −100 to 0 mm 23% show a good soil nutrient status and another 29% are classified in the medium soil nutrient category. A combined amount of 45% are represented by the site water budget class of −200 to −100 mm, divided into 20% medium and 25% poor soil nutrient supply. The lower three rows of heatmaps display the distributions of sites according to site water budget conditions in the period 2051 to 2070 applying three climate scenarios of minimum, median and maximum. Obviously, the sites in Uelzen and Fläming face dryer conditions in the future. The shift of site proportions towards more negative site water budget classes is intensified the more extreme the underlying climate scenario. While under minimum climate scenario still 28% of all sites in Uelzen exhibit a positive site water budget in the period 2051 to 2070, almost the entire area suffers negative balances under median and maximum climate scenarios. In Fläming the shift is even more severe as large proportions of sites are categorized into the site water budget class −100 to −200 mm (67% under minimum climate scenario), into the class −200 to −300 mm (61% under median and 24% under maximum climate scenario) or even below −300 mm (35% under median and 75% under maximum climate scenario).
2.4. Rule-Set for Stand Type Planning
As a starting point in our stand type planning analysis we assume that all sampled stands in the two regions are final harvested in 2011 and a new forest generation is planned on these sites. The climatic conditions of the period 2051 to 2070 under the climate scenarios of minimum, median and maximum specify drought restrictions considered in the planning process. Other than simulating a progressive harvesting and planning over time taking actual stand development into account, this approach bears the advantage that all forest stands will be 50 years old in mid-period 2051–2070. This is crucial as the rejection of species in the planning process because of drought restrictions needs to be oriented towards future climate conditions. With 50 years into stand development this should be sufficient to warrant at least stand survival refraining from other risk factors. Furthermore, this approach enhances the comparability between the different forest management concepts as it only applies the different species preferences in the course of stand type planning and ignores the different rotation ages. We justify this theoretical construct—to ignore the actual stand ages and instead assume immediate planning of new forest generations on all sites in 2011—because the sampled forest data from the two regions is representative for large growth zones. Therefore, we assume that in the growth zones a sufficient number of stands having the same site and species entities as in the analyzed sample are close to rotation age and are pending for final harvest in 2011. Consequently, the results derived from the study areas in Uelzen and Fläming are valid for their respective growth zones.
Forest management concepts predetermine decisions on species selection for the next forest generation. In order to evaluate the interaction between species selection priorities, changing climatic conditions and drought restrictions, we implement three contrasting forest management concepts: (1) a business-as-usual strategy which mimics currently-applied silvicultural practices on public forest land in Germany; (2) a climate protection strategy which aims to maximize CO2 sequestration in standing volume as well as wood products; and (3) a biodiversity strategy which focusses on increasing the area of potential natural vegetation (for more information see [53]). The selection of future tree species always has to be in line with species site-suitability for all management strategies. The business-as-usual strategy prefers deciduous species and mixed stand types as defined by the principles of close-to-nature silviculture. In the climate protection strategy a high CO2 sequestration can best be realized by fast-growing conifer species. The biodiversity strategy consequently selects tree species of potential natural vegetation.
In the first stage of the planning process for each forest site the next generation of species is selected according to common methodology based on a matrix of soil nutrients and current site water budget (Figure 4) [54].
The ecogram concept illustrated in Figure 4 is further refined to meet forest management goals and adjusted to the regional climate conditions of growth zones. Thus, we obtain one matrix for each applied management strategy and study region (cf. Tables S1–S6 in the supplement). Stand types are assigned based on 7 soil nutrient classes and 14 site water budget classes as is customary in forest planning in the state forests of Lower Saxony [54]. In many cases several stand types are site-suitable. Therefore, following rules are applied to determine the final decision. Under the business-as-usual strategy, the stand type among the recommended is selected which is closest to the current stocking, i.e., planning the same stand type or including the leading or mixed species if possible. In case none of the current species is among the suggested stand types, a completely different stand type is planned. The reasoning behind this approach is to keep the costs of forest conversion as low as possible, i.e., to utilize natural regeneration instead of planting. The concept of the climate protection strategy as well as the biodiversity strategy is to reverse the current management practices and to stress the new objectives. Thus, regardless of the current stocking, under climate protection and biodiversity strategy a stand type is randomly selected from the recommended according to predefined probabilities in any case (probabilities are given in Table S7 in the supplement).
Although current regional climate conditions are considered in this planning approach as site water budget is a function of precipitation and temperature, explicit changes of the climate conditions are not taken into account. Thus, in this study we refer to the first stage as planning under current (constant) site conditions. The species assignment is motivated mainly by silvicultural management decisions always respecting site suitability.
Climate projections strongly suggest a significant shift in site water budget within a time horizon well less than rotation ages. Therefore, in a second planning stage we account for climate change by assessing the potential future site water budget under different climate scenarios. This second stage is termed planning under future site conditions or adaptive species planning. Specifically, at each site the suggested species of the first stage is further checked for drought restrictions. Thus, species assignment is motivated by silvicultural management decisions respecting current site suitability and future drought restrictions.
In forest planning the decision on species’ site-suitability has to rely on certain thresholds which evaluate drought vulnerability. Ecosystem vulnerability in the context of climate change is commonly defined as the impact of adverse climatic effects on ecosystem services [55]. Considering the available data, which needs to be at hand or could easily be generated for each forest site in the planning process, we propose to use site water budget during growing season, i.e., the sum of the climatic water balance in the growing season plus the available soil moisture, as an indicator in the drought vulnerability assessment [28,56]. The continuous parameter site water budget is converted into vulnerability classes following the threshold theory according to [44]. For five major tree species Table 3 presents the applied vulnerability classes and their interpretation in the planning process.
According to the specified thresholds in Table 3 Norway spruce is the most drought-prone species, followed by European beech, the oak and Douglas-fir group and Scots pine as the most drought-resistant species. For a complete planning foundation other species are assigned to one of the four groups according to Table S8. As an adaptation measure to future dryer conditions we recommend to establish species only as primary species when classified in the low drought vulnerability class. Species should be planned as admixture species when medium drought vulnerability is projected. And finally, no cultivation should be envisaged under high drought vulnerability conditions.
In the stage of adaptive forest planning new stand types are assigned if drought restrictions are exceeded according to the rule-sets provided in Tables S9 and S10. The adaptive planning stage operates stepwise, i.e., if the leading or the admixture species of stage 1 does not meet the drought threshold the first alternative stand type is tested. If the first alternative also misses the requirements the second alternative stand type is examined, etc. In some cases two stand types are recommended as alternatives, which are then planned in equal proportions if selected. In order to guarantee stocking with forests even under severe drought conditions some stand types are taken as last planning alternatives; these are pure Scots pine, Scots pine-oak mixtures, Scots pine-birch mixtures and birch-Scots pine mixtures.
For a more lucid analysis of results simplified stand types are formulated, i.e., species mixtures, for instance, Scots pine with beech or with birch are aggregated to the stand type Scots pine-deciduous species mixture. Table 4 lists the simplified stand types and their abbreviations which are further used throughout the text.
2.5. Adaptation Effort
It is important to define a sound reference in order to evaluate the adaptation effort in the second stage. Intuitively, it seems to make a big difference in terms of adaptation effort whether only the admixture species has to be planned differently, e.g., instead of a Scots pine-European beech stand a Scots pine-oak stand, or if the leading species does not meet the drought restrictions, e.g., instead of a European beech-Scots pine stand an oak-Scots pine stand, or even the leading and admixture species have to be changed, e.g., instead of a European beech-Norway spruce stand a birch-Scots pine stand. Therefore, we assign following scores according to the species proportion that needs to be changed, always assuming for mixed stands that the total splits into 70% leading species and 30% admixture species: (1) 0.7 if the leading species is changed and the former does not become the admixture species; (2) 0.4 if the leading species is changed and the former becomes the admixture species; (3) 0.3 if the admixture species is changed. The sum of all scores within a study region comprises the overall adaptation effort score (AES). The AES has a natural range of values between 0 (no adaptation in stage 2 is necessary at all) and 100 (absolutely no realizability of stage 1 planned species). Thus, the AES can be interpreted as the species proportion that needs adaptation to meet the drought restrictions.
3. Results
3.1. Stand Type Planning under Constant Site Conditions
Based on the distributions of current stand types pending for potential species conversion until 2070 in the two regions of Uelzen and Fläming under the three management scenarios (business-as-usual, climate protection and biodiversity), Figure 5 shows the resulting distributions planned under constant site conditions (stage 1). In addition, Figure 6 and Figure 7 present the distributions applying adaptive planning (stage 2). The main focus of this analysis lies on the identification of uncertainty arising in the planning process due to different climate projections.
The proportions of stand types pending for potential conversion in 2011 are, of course, identical for each forest management strategy (Figure 5, upper row, dark blue bars). In Uelzen, pure Scots pine stands make up the highest proportion of the stands to be potentially converted. The proportion of Scots pine-conifer mixtures follows second. Norway spruce stands, Scots pine-deciduous species mixtures and broadleaves of low rotation age contribute considerably as well. The succeeding distributions planned under constant site conditions (stage 1), however, show significant differences between business-as-usual, climate protection and biodiversity management (Figure 5, upper row, light blue bars). Obviously, these differences reflect the different silvicultural objectives of the management scenarios. In Uelzen, no pure Scots pine and Norway spruce stands are planned in stage 1 no matter which management scenario is applied. Under business-as-usual management there is a strong shift towards Scots pine-conifer mixtures. Beech mixtures, oak mixtures, Douglas-fir mixtures as well as pure beech and broadleaves of high value are also planned to a considerable extent. Under the climate protection strategy stand types are mainly transferred to Grand fir mixtures, Douglas-fir mixtures, beech mixtures and Scots pine-deciduous species mixtures. The biodiversity strategy favors oak mixtures, Scots pine-deciduous species mixtures and beech mixtures.
In Fläming, more than 80% of the stands pending for potential conversion are pure Scots pine in 2011. Scots pine-deciduous species mixtures, oak mixtures and broadleaves of low rotation age contribute to a much lesser degree (Figure 5, bottom row, dark blue bars). In accordance with the findings in Uelzen, the distribution of succeeding stand types planned under constant site conditions (stage 1) mirrors management goals (Figure 5, bottom row, light blue bars). Under business-as-usual management mainly pure Scots pine and Scots pine-deciduous species mixtures and minor proportions of oak mixtures and very few beech mixtures are planned. Under the climate protection strategy, there is a shift towards highly productive conifers, i.e., Douglas-fir mixtures, Grand fir mixtures, and Scots pine-deciduous species mixtures as well as a few European larch-beech mixtures. Species planning under the biodiversity strategy strongly promotes beech mixtures, Scots pine-deciduous species mixtures and oak mixtures.
These resulting stand type distributions planned under constant site conditions are subject to examination under climate change conditions, in particular drought stress, in adaptive stand type planning (stage 2).
3.2. Adaptive Stand Type Planning under Drought Restrictions
The need to adjust the stand type selection under constant site conditions when considering drought stress restrictions (cf. Table 3) demonstrates in part the impact of climate change on forest planning. The results can be analyzed looking at three aspects: (1) the change in selected stand types, where a decrease indicates drought vulnerability and an increase is the result of the assignment of more drought tolerant stand types (Figure 6 and Figure 7); (2) the absolute need of adjustment expressed by the adaptation effort score (AES) which is the percentage of necessary change in stand type area between planning under constant site conditions (stage 1) and under adaptive planning (stage 2) in regard to the three climate projections of minimum, median and maximum (Table 5); and (3) the uncertainty arising in the planning process due to different climate projections, characterized by the spread between the necessary adjustments in stand type selection, i.e., the difference between the AES values under the minimum and the maximum climate scenario (Table 5).
The detailed analysis of changes on stand type level in Figure 6 (top) reveals that in Uelzen under business-as-usual management the planned pure beech and beech mixtures as well as Douglas-fir-beech mixtures are most vulnerable. Pure beech and Douglas-fir-beech mixtures almost entirely exceed the drought restrictions under median and maximum climate scenario. Oak mixtures and Douglas-fir mixtures are seen as alternative stand types with increasing proportions in stage 2. Clearly, the proportions of the most drought-tolerant stand types, Scots pine-conifer and Scots pine-deciduous species mixtures, increase with intensifying drier conditions. Under climate protection management the proportions of the highly productive stand types Grand fir mixture, Douglas-fir-beech mixture decrease when more extreme climate projections (median and maximum) are assumed (Figure 6, middle). Also beech mixtures, European larch-beech mixtures and Norway spruce mixtures increasingly miss the drought limits under median and maximum climate run. Oak mixtures and Douglas-fir mixtures benefit under minimum and median climate projections as other species suffer from dryer conditions. But under the maximum climate scenario these stand types almost disappear. On the other hand, the proportion of Scots pine mixtures increases drastically. Under the biodiversity strategy the area of beech mixtures strongly decreases with intensifying drier conditions (Figure 6, bottom). The oak mixtures gain proportions under the minimum and median climate scenarios. Under maximum climate scenario also a large proportion of the planned oak mixtures exceed the drought restrictions. While the small proportion of broadleaves of low rotation age stays more or less constant, the already large proportion of Scots pine-deciduous species mixtures (28% under stage 1) increases to 73% under maximum climate projection.
The results for the Fläming region in Figure 7 show that under business-as-usual management the high proportion of pure Scots pine (53%) is constant over the planning stages and climate scenarios. A sharp decrease in the proportions to almost absence is observed for oak mixtures and beech mixtures. Scots pine-deciduous species mixtures receive a considerable gain. Under the climate protection strategy drought restrictions notably reduce especially Douglas-fir mixtures, Scots pine-deciduous species mixtures, Grand fir mixtures, larch-beech mixtures and even oak mixtures. On the other hand, pure Scots pine and, most prominently, Scots pine-conifer mixtures profit in the adaptive planning process (from 1% in stage 1 to 30% in stage 2 under minimum climate scenario and over 40% under median and maximum climate scenario). Under biodiversity management beech mixtures and oak mixtures lose large proportions with increasingly drier conditions. The in the first planning stage rarely represented European larch-beech mixtures, Norway spruce mixtures, Douglas-fir-beech mixtures and broadleaves of high value are entirely disregarded when applying drought restrictions. Scots pine-deciduous species mixtures are the alternative stand types in adaptive planning with increases of 35 percentage points under minimum climate scenario and 42 percentage points under median and maximum climate scenario. To a much lesser extent also broadleaves of low rotation age profit from the drought restrictions.
Obviously, the effort for adaptive planning increases the more extreme the underlying climate projection, for all management strategies in both regions (Table 5). The AES values, i.e., the species proportion that needs adaptation to meet the drought restrictions in stage 2, indicate a negligibly small effort under the minimum climate scenario for all management scenarios (between 0.3 and 3.2) in the Uelzen region. However, already assuming the median climate scenario the AES jumps up to considerable values of 17.9 under business-as-usual, 31.5 under climate protection and 48.5 under biodiversity management. The need to adjust is even more dramatic under the maximum climate scenario especially in regard to climate protection and biodiversity management, reaching AES values of 54.8 and 74.1, respectively.
In terms of planning uncertainty, business-as-usual is rather robust comparing the management strategies with an uncertainty range of 19.8 percentage points and only a 3.9 percentage point difference between median and maximum climate run (Table 5, last column). Quite differently, under climate protection and biodiversity management the planning uncertainty is extremely large (range of 54.5 and 70.9 percentage points, respectively). Consequently, under these management strategies it is of high importance which climate scenario is assumed during the planning process as it has a strong impact on stand type distribution.
In contrast to the findings in the Uelzen region the effort for adaption applying the drought restrictions is already quite extensive under the minimum climate scenario for all management strategies in Fläming (Table 5). While under business-as-usual the proportion of species which need adaptive planning is 12% and 28% under climate protection, more than half of the species planned in stage 1 need to be replaced in stage 2. However, the increase in the AES with intensifying drier conditions is not as pronounced as in Uelzen, reaching AES values of 15.1%, 40.6% and 56.2% respectively under business-as-usual, climate protection and biodiversity management in regard to the maximum climate scenario.
This relatively small climate effect on stand type planning in Fläming becomes even more obvious, when looking at the inherent uncertainty. Under business-as-usual management the range between minimum and maximum climate scenarios amounts to almost negligible 2.9 percentage points and under climate protection to 12.4 percentage points. The effect of climate uncertainty on the biodiversity strategy is only just 5.7 percentage points.
The changes on the stand type level explain why already the minimum climate scenario causes a high need for adaptive planning and the rather low differences between the impacts of the three climate projections in Fläming. The drought restrictions of the site-adapted species under constant site conditions, for instance oak and beech under business-as-usual and biodiversity management and additionally Douglas-fir and Grand fir under climate protection, are exceeded on many sites in Fläming under the minimum climate scenario already. While there are more site-adapted alternatives for stand types under adaptive planning graduated to climate scenarios in Uelzen, in Fläming the most drought-tolerant stand types, pure pine and pine mixtures, make up the alternative in many cases already under minimum climate scenario. Thus, just little further adaptive planning is possible under even more severe climate projections.
4. Discussion
The findings and the knowledge gained depend on several assumptions and methodological approaches applied. Thus, before we discuss the results in more detail we elaborate on three issues, i.e., (1) tree species selection in the light of changing environmental conditions, (2) the method to assess drought vulnerability and its implications on the planning process and (3) the approach to incorporate climate uncertainty when selecting tree species in the context of forest conversion.
4.1. Tree Species Selection in Light of Changing Environmental Conditions
Forest conversion strives to optimize the achievement of forest enterprise targets in general. This specifically encompasses adaptation measures to changing environmental conditions (e.g., [13,57,58]). Forest conversion involves changes in tree species in the course of regeneration as well as in stand structure objectives, i.e., pure stands versus mixed and even-aged stands versus uneven-aged structures. This study exclusively deals with species selection under changing environmental conditions. It is not considered, whether regeneration is introduced after final harvest of the current stocking or as advance planting under shelter. Also forest management practices to convert forests from even-aged to uneven-aged structures are not presented. Then again, potential changes from pure stands to species mixtures are indeed incorporated in the rule-set for stand type planning.
While many studies develop important theoretical foundations for forest conversion based on model stands or model forest enterprises (e.g., [13,57,59,60,61,62]), we implement a forest planning procedure to select tree species for regeneration based on actual forest inventory data and subject to site information and climate projections on a regional level. As is the aim of the study we focus on the effect of changes in drought risk on future species’ site suitability, and we neglect other environmental aspects such as potential reduction in nitrogen deposition [63,64], the influence of sulphur deposition [65] or rising CO2 concentrations [66].
The presented rule-set for species selection is based on forest planning procedures which are applied in Germany today [54,67,68]. The assignment of site-suitable species according to soil nutrients and site water budget takes regional climate properties into account as precipitation and temperature determine site water budget. Furthermore, to warrant flexibility in the planning process several suitable species or species mixtures are listed, if possible. Thus, we explicitly assigned stand types for regeneration based on site-suitability and according to the respective silvicultural management strategy (cf. Tables S9 and S10). Consequently, under business-as-usual management deciduous species and mixtures are preferred, under the climate protection strategy fast-growing conifer species are favored, and the biodiversity strategy emphasizes potentially natural species compositions. These first stage assignments result in a strong predisposition for a need of adaptive planning in the second stage. The entire species selection procedure in both stages abstracts from other particulars to clarify the impact of forest management targets and the effect of climate change.
It is important to note that we did not impose any restrictions other than site-suitability and respecting the drought vulnerability thresholds on the selection of stand types. Of course, large forest enterprises will formulate targets for species distributions [69]. For example, the forest enterprise of Lower Saxony strives to establish deciduous species on 65% of the forest area [54]. Some forest areas will be excluded from forest conversion due to nature protection regulations. And on the regional level different forest owners will pursue different forest management strategies. Beyond that it is also plausible that new stand types need to be defined to meet adaptation demands. For example, the Douglas-fir-beech mixture is converted to conifer mixtures or oak mixtures if drought restrictions apply (cf. Tables S9 and S10). Just as well an alternative to meet drought thresholds could be the newly formulated stand types Douglas-fir-Red Oak or Douglas-fir-Littleleaf Linden.
Furthermore, other risk factors will hamper some species from regeneration as well. Especially in the North German Plain, biotic risk factors such as several Scots pine-related insect pests, nun moth, bark beetle, jewel beetle and root rot may cause severe damage to forest stands. These boundary conditions of additional restrictions and further risk factors need to be considered when species are selected for regeneration under real-life conditions. Extensive knowledge on the environmental conditions for cultivation, growth potential of tree species and possible risk factors as well as awareness of uncertainties are imperative, as species choice for regeneration is a long-term commitment. For example, Schou et al. [70] show how climate change-related uncertainty and the risk of stand destruction influence decision making on converting Norway spruce stands in Denmark.
By virtue of focusing solely on drought restrictions and three management strategies, our analysis adds insights to the topic of species selection, necessary adaptation measures on a regional level and, above all, the interaction between management strategies and climate change in connection with inherent uncertainty.
4.2. Drought Vulnerability Assessment in Forest Planning
It is crucial in forest planning that the information necessary for well-founded decisions is ubiquitously available. And as drought vulnerability very much depends on local site conditions (e.g., [71,72,73]), consequentially thresholds for species selection considering drought risk should be based on easily available site parameters. Our approach to integrate a drought vulnerability assessment in forest planning is straightforward: we employ species-specific site water budget thresholds based on observed species occurrence subject to their productivity and classify three options for cultivation, i.e., as primary species, as admixture species and as an exclusion criteria. The ranking of species with respect to drought vulnerability coincides with various other studies in central Europe (e.g., [74,75,76,77]). Our method is in line with the often applied relative transpiration index (RTI) [78,79] which accentuates absolute water availability. Müller [78] describes how to derive the RTI based on climatic water balance and available soil moisture, whose sum equals the site water budget used in this study. Furthermore, it seems plausible to use the long-term climatic trend of site water budget as a fuzzy decision criteria for forest planning. On the other hand, short-term singular but extreme dry spells might contradict species selection recommendations (e.g., [80,81,82,83,84,85,86,87]). Choat et al. [30] conclude that these singular deviations from the long-term climatic trend are crucial for species-specific drought vulnerability. However, assessing species- and site-specific drought stress currently still lacks universal and established findings [30,31]. Allen et al. [88] extensively compiled findings of lesser vulnerability as well as more rapid mortality due to drought and they identified six generally accepted global vulnerability drivers, i.e., droughts can eventually occur everywhere, droughts become hotter with warmer climates, non-linear relationships exist between atmospheric moisture demand and temperature during drought, hotter droughts trigger mortality faster, higher frequency of short droughts will eventually trigger mortality, and time-span for forest recovery after drought is long compared to relatively rapid mortality. This underlines the complex nature of evaluating the effect of drought on forests, hence, it is important to provide a robust method for forest planning which can be refined when new insights are available.
4.3. Climate Uncertainty in the Context of Forest Conversion
Many aspects contribute to uncertainty when future developments are anticipated. Walker et al. [89] provide a conceptual framework to classify the various dimensions of uncertainty in model-based decision support. In the case of climate impact research Wilby and Dessai [90] illustrate how uncertainty amplifies in cascades, beginning with assumptions on future societies, the resulting emission scenarios, the applied global circulation models for climate projections, which are further downscaled using regional climate models, and finally the uncertainties associated with impact models and the transfer of projected results to adaptation measures.
Uncertainty in the climate projection itself arises from three distinct sources, i.e., natural fluctuations in the climate system, climate model uncertainty and scenario uncertainty [91]. As different climate models project different changes in climate variables in response to the same radiative forcing, the presented climate impact analysis substantially depends on the selected climate model STARS. STARS projects future climate based on a temperature-driven resampling scheme (TCR). The temperature trend is usually predefined by the gradient in the results of global climate models, in this case INM-CM4, ECHAM6 and ACCESS1.0. STARS’ resampling scheme rearranges annual climate values in a first step and subsequently reorders 12-day-weather sequences to achieve the temperature trend. Wechsung and Wechsung [39,40] critically evaluate this methodology. They quantify a negative precipitation bias of approx. 5% per 1 K temperature increase. Despite the model limitations, Wechsung and Wechsung [40] recommend STARS for vulnerability and uncertainty studies. In any case, the potential precipitation bias introduced by STARS should be taken into account when interpreting the presented results. Overall, Hawkins and Sutton [91] conclude that the uncertainty in regional climate projections for the next few decades is strongly affected by model uncertainty and natural fluctuations, whereas for the far-future, scenario uncertainty contributes dominantly to overall uncertainty.
In our study we follow proposals by, e.g., [15,92,93,94] using three climate scenario projections to at least partially account for climate uncertainty in our analysis. On that part Jones [92] emphasizes that any conclusion derived from impact analysis must remain highly conditional on the underlying ranges of climate projections. Furthermore, the full range of climate uncertainty is unknown [89] and even in the framework of large climate projection ensembles the uncertainty cannot be precisely quantified [18,95]. Therefore, as our model-based decision support on species selection heavily relies on the three assumed climate projections and as the uncertainty associated with these climate projections is only a rough approximation, any recommendations based on our results must be robust, i.e., at best species selection complies with each underlying climate scenario.
According to Dittrich et al. [96] robust decisions achieve the aspired goals under a broad range of future developments, which makes them particularly suited for deep uncertainty. Among the many strategies to adapt to uncertain futures some seem especially consistent with forest planning demands, e.g., selecting “no-regret” strategies, preferring flexible options, anticipating “safety margins” or reducing the decision time horizon (cf. [97]). Concerning our analysis of species selection under drought vulnerability, a “no-regret” strategy could be to opt for the most drought-resistant species, i.e., Scots pine, which yields benefits even under the most extreme climate scenario as well as under current conditions. Paraphrased, Scots pine has a low sensitivity to false assumptions regarding future climate conditions but, on the other hand, this security is traded for potentially higher productivity by other species. Flexible options and “safety margins” could encompass species mixtures such as Scots pine and European beech and/or oak, i.e., two or three major species spread the risk between drought-resistant but less-productive and more drought-vulnerable but also more-productive species. Furthermore, in the course of time, forest management can be adjusted to new insights and the changing climate conditions. Finally, reducing the decision time horizon can easily be achieved by adjusting the silvicultural expectation, i.e., for example, reducing the target diameter for final harvest in Douglas-fir from 70 cm to 60 cm shortens the time horizon by approximately 20 years. This could very well determine whether or not to select Douglas-fir for regeneration.
In this study we clearly show that the amount of uncertainty due to climate change concerning species selection strongly depends on the forest management strategy chosen. While it is easy for forest practitioners to opt for a certain forest management strategy, it is much more difficult for them to base their decision for species selection on a chain of model predictions, the underlying assumptions and the inherent uncertainty. Although decision support provides information on different scenarios and their implications on forests and forest enterprises, in the end one forest planning alternative has to be set into practice. As is the case with risk assessment, where decision-makers can act in a risk-averse, risk-neutral or risk-taking manner [98,99], the attitude towards uncertainty in general and climate-induced projection variability specifically is highly person-specific with a strong subjective share. Thus, although one cannot assign probabilities to climate change scenarios for theoretical reasons [15], the decision-maker ultimately needs to select one scenario as the most probable in his personal assessment, converting belief into subjective probability (cf. [21]). Yousefpour et al. [100] take decision-making in the context of adaptive forest management one step further as they show how forest managers’ beliefs about the effects of climate change are altered by new insights and how this additional knowledge influences their decisions.
4.4. The Findings and Their Implications in a Broader Context
The two most important issues to be discussed concerning the results of this study are (1) whether or not the specific management goals can be realized under drought restrictions and (2) how a forest manager’s belief in a certain climate scenario influences adaptive planning.
First of all, the results and their implications for forest planning are very specific for each growth zone, represented by the two study regions Uelzen and Fläming. In Uelzen, favored by site, climate and higher species diversity forests are less vulnerable to drought than in Fläming. In Uelzen, under the minimum climate scenario, the management goals of each silvicultural strategy, business-as-usual, climate protection and biodiversity, are realizable without concessions. Under median and maximum climate scenario and business-as-usual management, about one fifth of species proportions need to be adjusted to drought restrictions. Basically this involves beech mixtures being exchanged for oak mixtures and Scots pine mixtures; thus, it is feasible to deem the management goals achieved under drought restrictions as well. Therefore, in the case of business-as-usual silviculture in Uelzen, the uncertainty in climate projections plays a minor role in achieving the management goals, although adaptive planning is necessary for the median and maximum climate scenario. Under the biodiversity strategy, beech mixtures are replaced by oak mixtures and Scots pine-deciduous species mixtures. As birch-oak mixtures and pine mixtures are potentially natural forest ecosystems in Uelzen [101], this shift does not contradict the biodiversity strategy at all. Thus, the forest manager’s belief in a certain climate change scenario is insignificant for biodiversity goal achievement in Uelzen. Nevertheless, the proportion of adaptive planning rises up to 74% under the maximum climate scenario. However, adaptive planning under the climate protection strategy is not in line with management goals when applying the median or maximum climate scenario. The most productive Grand fir mixtures and Douglas-fir mixtures will not meet drought restriction thresholds and need to be exchanged for Scots pine mixtures and oak mixtures if no new alternative stand types as, for example, Douglas-fir-Red Oak or Douglas-fir-Littleleaf Linden are considered. Depending on the decision-makers’ subjective choice of climate scenario, putting aside the minimum climate run, the management goals will be missed with 32% replacement under median and 55% replacement under the maximum climate scenario.
Most striking about the analysis for the Fläming region is the perception that climate uncertainty is practically meaningless for adaptive planning decisions. So no matter what the forest manager’s belief and which silvicultural management strategy pursued, the maximum deviation in stand type replacement is 12 percentage points. On the other hand, even under minimum climate scenario, the necessary replacement of species proportions under adaptive planning is between 12% (business-as-usual management) and 50% (biodiversity management). For biodiversity management goals the shift from beech mixtures and oak mixtures to Scots pine-deciduous species is in line with potentially natural forest ecosystems in Fläming [102], thus the management goals are realizable. Under business-as-usual strategy oak mixtures and beech mixtures have to be exchanged for Scots pine mixed either with deciduous or with other conifer species. Therefore, concessions to meet the desired close-to-nature management goals, are necessary, especially when looking at the aim to increase the area of deciduous species. The climate protection strategy is the most crucial in respect to goal achievement. Like in Uelzen, the most productive stand types of Douglas-fir mixtures and Grand fir mixtures are replaced by pure Scots pine and Scots pine-conifer mixtures under adaptive planning to meet drought restrictions. This clearly contradicts the management goals to select fast-growing conifer tree species to a great extent.
In summary, it is important to note that adaptive planning is potentially sub-optimal in regard to forest management goals, i.e., concessions might be unavoidable concerning ecological and/or economic targets [11]. Moreover, in some special cases adaptive planning may be independent from climate uncertainty and equates to robust decision making, provided the decision maker is aware of the determining factors (e.g., [103,104,105]).
Altogether, in Uelzen under the minimum climate scenario and averaging the three management scenarios, 2% of the species proportions need to be exchanged to meet drought vulnerability thresholds. Under the median climate scenario the proportion is 33% and under maximum run is 50%. In Fläming the proportions for replacement range from 30% to 37%. This assessment is comparable to findings by Schelhaas et al. [13], who report that between 36% and 40% of the forest area in Europe is expected not to meet species suitability criteria by 2070 because of climate change. However, the reservation must be made that the forest area considered for active species conversion in this study is hypothetical, as financial restrictions or silvicultural principles might prevent planting. Nevertheless, the proportions of necessary stand type replacements indicate the need for adaptation measures.
In a broader context Spathelf et al. [106] conclude that close-to-nature silviculture falls short to be an adequate concept to adapt forests to climate change, when taken seriously about the two key elements of preference of natural regeneration and low-impact interventions. They argue that silvicultural concepts which allow for small-scale disturbances to promote a broad range of species to regenerate and thus to enhance stand resilience are more suitable. However, as site conditions rapidly change, rigid conceptions on the unrestricted advantageousness of natural regeneration need revision [14]. Additionally, in this study we just selected stands for potential conversion based on the framework conditions of three different management strategies without implicitly assuming the according principles for conversion.
The presented findings rely on the applied method to assign stand types according to site properties related to soil nutrients and site water budget. Drought restrictions are considered in site water budget and are modelled dynamically over time. Bolte et al. [62] introduce an alternative method to select species according to site suitability. They propose to evaluate the regional risk for species based on their retrospective performance, to apply climate envelope analysis and to use climate matching analysis. This sophisticated procedure will certainly yield comprehensive science-based recommendations for species selection. We believe, however, that implementing a slightly advanced species selection procedure which is based on traditional methods has an increased likelihood of adoption in many forest planning practice.
5. Conclusions
As Yousefpour et al. [104] point out, it is of outmost importance for successful adaptive forest planning that decision-makers are aware of the interactive nature of management goals and limitations imposed by changing climate conditions and their intrinsic uncertainty. Complementary to more theoretical or large-scale approaches (e.g., [13,21,57,61,62]) which clarify tendencies and draw general conclusions, our regional-scale approach reveals specific management options for the next generation of forests based on given forest stands on defined sites in a region under climate change. Furthermore, we promote a method for decision support which builds on easily and ubiquitously available data, i.e., site water budget under climate change. Moreover, the presented procedure to assess site-suitability of species under climate change is an advancement of the well-established current method. Thus, acceptability in forest practice is assumed high.
Four main conclusions are drawn: (1) Obviously, the need for adaptive planning considering drought vulnerability strongly depends on the climate change scenario subjectively deemed most probable and the underlying forest management goals; (2) Adaptive planning might force a compromise between responsible forest planning decisions and forest management goals; (3) Uncertainty in climate change scenarios and, thus, the decision-makers subjective choice of a certain climate projection may influence species selection considerably, e.g., climate protection management in Uelzen, or in some other case may be irrelevant, e.g., biodiversity management in Fläming; (4) Recommendations for species selection in Uelzen imply that the need for adaptive planning is clearly lower than in Fläming and consequently, most management goals are realizable with some minor concessions. However, the choice of climate scenario has a considerable effect on the assessment of species suitability. In Fläming there is an extensive need for adaptive planning and, as a result under business-as-usual and climate protection management strategies, fairly strong adjustments to management goals are necessary. On the other hand, climate uncertainty is not an issue in Fläming.
Future research should focus on better methods to quantify the effect of drought on productivity and tree vitality, ultimately in interaction with other abiotic and biotic risk factors. To further enhance the scientific basis for forest planning, further risk factors need to be incorporated into the planning process.
Supplementary Materials
The following are available online at www.mdpi.com/1999-4907/8/10/363/s1, Table S1: Assignment of stand types in regard to soil nutrient and site water budget matrix under business-as-usual management in Uelzen, Germany, Table S2: Assignment of stand types in regard to soil nutrient and site water budget matrix under climate protection management in Uelzen, Table S3: Assignment of stand types in regard to soil nutrient and site water budget matrix under biodiversity management in Uelzen, Table S4: Assignment of stand types in regard to soil nutrient and site water budget matrix under business-as-usual management in Fläming, Germany, Table S5: Assignment of stand types in regard to soil nutrient and site water budget matrix under climate protection management in Fläming, Table S6: Assignment of stand types in regard to soil nutrient and site water budget matrix under biodiversity management in Fläming, Table S7: Selection probabilities for random draw of stand types, Table S8: Assignment of other species (limited to species relevant in the two study regions) to species groups according to drought vulnerability (leading species in group in italics), Table S9: Assignment of stand types in Uelzen according to the silvicultural management strategy under constant site conditions (1. stage) and applying drought restrictions (2. stage), Table S10: Assignment of stand types in Fläming according to the silvicultural management strategy under constant site conditions (1. stage) and applying drought restrictions (2. stage).
Acknowledgments
We gratefully acknowledge the critical remarks and helpful comments by three reviewers. We thank our colleague Bernd Ahrends for processing the soil data. The climate data was provided by the Potsdam Institute for Climate Impact Research (PIK). We also thankfully acknowledge Peter C. Werner, PIK, for processing the climate projections data. This study was carried out within the project “Nachhaltiges Landmanagement im Norddeutschen Tiefland”, which was funded by the German Federal Ministry of Education and Research under reference number 033L029. Funding to cover the costs to publish in open access was provided by the Northwest German Forest Research Institute.
Author Contributions
M.A. conducted the data analysis and wrote the first draft of the paper. R.-V.N., R.S.N. and H.S. formulated the forest management rule sets. R.-V.N. and H.S. customized the assignment of stand types in regard to soil nutrient and site water budget matrix. R.S.N. processed the raw data. J.S. and H.S. derived the method for drought vulnerability assessment. All authors jointly formulated the research questions and worked on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smith, S.J.; Edmonds, J.; Hartin, C.A.; Mundra, A.; Calvin, K. Near-term acceleration in the rate of temperature change. Nat. Clim. Chang. 2015, 5, 333–336. [Google Scholar] [CrossRef]
- Ballester, J.; Rodó, X.; Giorgi, F. Future changes in Central Europe heat waves expected to mostly follow summer mean warming. Clim. Dyn. 2010, 35, 1191–1205. [Google Scholar] [CrossRef]
- Beniston, M.; Stephenson, D.B.; Christensen, O.B.; Ferro, C.A.T.; Frei, C.; Goyette, S.; Halsnaes, K.; Holt, T.; Jylhä, K.; Koffi, B.; et al. Future extreme events in European climate: An exploration of regional climate model projections. Clim. Chang. 2007, 81, 71–95. [Google Scholar] [CrossRef]
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, R.; Jones, R.; Kolli, R.K.; Kwon, W.; Laprise, R.; et al. Regional climate projections. In Climate Change, 2007: The Physical Science Basis; Contribution of Working Group I to the Fourth Assessment Report of theIntergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K., Tignor, M., Miller, H., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 847–940. [Google Scholar]
- Neumann, M.; Mues, V.; Moreno, A.; Hasenauer, H.; Seidl, R. Climate variability drives recent tree mortality in Europe. Glob. Chang. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Hummel, S.; Cullmann, D.A. Modelling and economic evaluation of forest biome shifts under climate change in Southwest Germany. For. Ecol. Manage. 2010, 259, 710–719. [Google Scholar] [CrossRef]
- Zubizarreta-Gerendiain, A.; Garcia-Gonzalo, J.; Strandman, H.; Jylha, K.; Peltola, H. Regional effects of alternative climate change and management scenarios on timber production, economic profitability and carbon stocks in Norway spruce forests in Finland. Can. J. For. Res. 2015, 283, 274–283. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.; Nabuurs, G.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2012, 3, 203–207. [Google Scholar] [CrossRef]
- Thiele, J.C.; Nuske, R.S.; Ahrends, B.; Panferov, O.; Albert, M.; Staupendahl, K.; Junghans, U.; Jansen, M.; Saborowski, J. Climate change impact assessment—A simulation experiment with Norway spruce for a forest district in Central Europe. Ecol. Model. 2017, 346, 30–47. [Google Scholar] [CrossRef]
- Schelhaas, M.J.; Nabuurs, G.J.; Hengeveld, G.; Reyer, C.; Hanewinkel, M.; Zimmermann, N.E.; Cullmann, D. Alternative forest management strategies to account for climate change-induced productivity and species suitability changes in Europe. Reg. Environ. Chang. 2015, 15, 1581–1594. [Google Scholar] [CrossRef]
- Brang, P.; Bugmann, H.; Bürgi, A.; Mühlethaler, U.; Rigling, A.; Schwitter, R. Klimawandel als waldbauliche Herausforderung. Schweiz. Z. Forstwes. 2008, 159, 362–373. [Google Scholar] [CrossRef]
- Foley, A.M. Uncertainty in regional climate modelling: A review. Prog. Phys. Geogr. 2010, 34, 647–670. [Google Scholar] [CrossRef]
- Knutti, R.; Sedlácek, J. Robustness and uncertainties in the new CMIP5 climate model projections. Nat. Clim. Chang. 2013, 3, 369–373. [Google Scholar] [CrossRef]
- Baumberger, C.; Knutti, R.; Hirsch Hadorn, G. Building confidence in climate model projections: An analysis of inferences from fit. Wiley Interdiscip. Rev. Clim. Chang. 2017, 8, e454. [Google Scholar] [CrossRef]
- Kandlikar, M.; Risbey, J.; Dessai, S. Representing and communicating deep uncertainty in climate-change assessments. C. R. Geosci. 2005, 337, 443–455. [Google Scholar] [CrossRef]
- Singh, R.; Reed, P.M.; Keller, K. Many-objective robust decision making for managing an ecosystem with a deeply uncertain threshold response. Ecol. Soc. 2015, 20, 12. [Google Scholar] [CrossRef]
- Lempert, R.J.; Collins, M.T. Managing the Risk of Uncertain Threshold Responses: Comparison of Robust, Optimum, and Precautionary Approaches. Risk Anal. 2007, 27, 1009–1026. [Google Scholar] [CrossRef] [PubMed]
- Yousefpour, R.; Augustynczik, A.L.D.; Hanewinkel, M. Pertinence of reactive, active, and robust adaptation strategies in forest management under climate change. Ann. For. Sci. 2017, 1–10. [Google Scholar] [CrossRef]
- Dessai, S.; Hulme, M. Assessing the robustness of adaptation decisions to climate change uncertainties: A case study on water resources management in the East of England. Glob. Environ. Chang. 2007, 17, 59–72. [Google Scholar] [CrossRef]
- United Nations Department of Humanitarian Affairs. Internationally Agreed Glossary of Basic Terms Related to Desaster Management. Available online: http://reliefweb.int/sites/reliefweb.int/files/resources/004DFD3E15B69A67C1256C4C006225C2-dha-glossary-1992.pdf (accessed on 11 May 2017).
- Van Heerden, K.; Yanai, R.D. Effects of stresses on forest growth in models applied to the Solling spruce site. Ecol. Model. 1995, 83, 273–282. [Google Scholar] [CrossRef]
- Hanson, P.J.; Weltzin, J.F. Drought disturbance from climate change: Response of United States forests. Sci. Total Environ. 2000, 262, 205–220. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.G.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.T.; et al. A global overview of drought and heat induced tree mortality reveals emerging climate change risk for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Birdsey, R.; Pan, Y. Drought and dead trees. Nat. Clim. Chang. 2011, 1, 444–445. [Google Scholar] [CrossRef]
- Spellmann, H.; Albert, M.; Schmidt, M.; Sutmöller, J.; Overbeck, M. Waldbauliche Anpassungsstrategien für veränderte Klimaverhältnisse. AFZ-Derwald 2011, 66, 19–23. [Google Scholar]
- Spathelf, P.; Van Der Maaten, E.; Van Der Maaten-Theunissen, M.; Campioli, M.; Dobrowolska, D. Climate change impacts in European forests: The expert views of local observers. Ann. For. Sci. 2014, 71, 131–137. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, L.D.L.; Anderegg, W.R.L.; Berry, J.A. Not all droughts are created equal: Translating meteorological drought into woody plant mortality. Tree Physiol. 2013, 33, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Adger, W.N. Vulnerability. Glob. Environ. Chang. 2006, 16, 268–281. [Google Scholar] [CrossRef]
- Moss, R.H.; Edmonds, J.A.; Hibbard, K.A.; Manning, M.R.; Rose, S.K.; Van Vuuren, D.P.; Carter, T.R.; Emori, S.; Kainuma, M.; Kram, T.; et al. The next generation of scenarios for climate change research and assessment. Nature 2010, 463, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5. [Google Scholar] [CrossRef]
- Orlowsky, B.; Gerstengarbe, F.W.; Werner, P.C. A resampling scheme for regional climate simulations and its performance compared to a dynamical RCM. Theor. Appl. Climatol. 2008, 92, 209–223. [Google Scholar] [CrossRef]
- Volodin, E.M.; Dianskii, N.A.; Gusev, A.V. Simulating present-day climate with the INMCM4.0 coupled model of the atmospheric and oceanic general circulations. Atmos. Ocean. Phys. 2010, 46, 414–431. [Google Scholar] [CrossRef]
- Stevens, B.; Giorgetta, M.; Esch, M.; Mauritsen, T.; Crueger, T.; Rast, S.; Salzmann, M.; Schmidt, H.; Bader, J.; Block, K.; et al. Atmospheric component of the MPI-M earth system model: ECHAM6. J. Adv. Model. Earth Syst. 2013, 5, 146–172. [Google Scholar] [CrossRef]
- Bi, D.; Dix, M.; Marsland, S.J.; O’Farrell, S.; Rashid, H.A.; Uotila, P.; Hirst, A.C.; Kowalczyk, E.; Golebiewski, M.; Sullivan, A.; et al. The ACCESS coupled model: Description, control climate and evaluation. Aust. Meteorol. Oceanogr. J. 2013, 63, 41–64. [Google Scholar] [CrossRef]
- Wechsung, F.; Wechsung, M. A methodological critique on using temperature-conditioned resampling for climate projections as in the paper of Gerstengarbe et al. (2013) winter storm- and summer thunderstorm-related loss events in Theoretical and Applied Climatology (TAC). Theor. Appl. Climatol. 2016, 126, 611–615. [Google Scholar] [CrossRef]
- Wechsung, F.; Wechsung, M. Short Communication Dryer years and brighter sky—The predictable simulation outcomes for Germany’s warmer climate from the weather resampling model STARS. Int. J. Climatol. 2014, 35, 3691–3700. [Google Scholar] [CrossRef]
- Bloch, R.; Wechsung, F.; Bachinger, J. Climate change impacts of legume-grass swards: Implications for organic farming in the Federal State of Brandenburg, Germany. Reg. Environ. Chang. 2015, 15, 405–414. [Google Scholar] [CrossRef]
- Schulla, J. Model Description WaSIM Completely Revised Version of 2012 with 2013 and 2015 Extensions; Hydrology Software Consulting J. Schulla: Zürich, Switzerland, 2015. [Google Scholar]
- Schulla, J.; Jasper, K. Model Description Wasim-Eth; Institute for Atmospheric and Climate Science, Swiss Federal Institute of Technology: Zurich, Switzerland, 2007; p. 181. [Google Scholar]
- Grier, C.C.; Running, S.W. Leaf area of mature northwestern coniferous forests: Relation to site water balance. Ecology 1977, 58, 893–899. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO Irrigation; Food and Agriculture Organization (FAO): Rome, Italy, 1998. [Google Scholar]
- Overbeck, M.; Schmidt, M.; Fischer, C.; Evers, J.; Schulze, A.; Hövelmann, T.; Spellmann, H. Ein statistisches Modell zur Regionalisierung der nutzbaren Feldkapazität von Waldstandorten in Niedersachsen. Forstarchiv 2011, 82, 92–100. [Google Scholar]
- Menzel, A. Phänologie von Waldbäumen unter sich Ändernden Klimabedingungen—Auswertung der Beobachtungen in den Internationalen Phänologischen Gärten und Möglichkeiten der Modellierung von Phänodaten; LMU München: München, Germany, 1997. [Google Scholar]
- Von Wilpert, K. Die Jahrringstruktur von Fichten in Abhängigkeit vom Bodenwasserhaushalt auf Pseudogley und Parabraunerde: Ein Methodenkonzept zur Erfassung Standortspezifischer Wasserstreßdisposition; Freiburger bodenkundliche Abhandlungen 24; Institut für Bodenkunde und Waldernährungslehre der Albert-Ludwigs-Universität Freiburg: Freiburg, Germany, 1990. [Google Scholar]
- Walther, A.; Linderholm, H.W. A comparison of growing season indices for the Greater Baltic Area. Int. J. Biometeorol. 2006, 51, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Frich, P.; Alexander, L.V.; Della-Marta, P.; Gleason, B.; Haylock, M.; Tank Klein, A.M.G.; Peterson, T. Observed coherent changes in climatic extremes during the second half of the twentieth century. Clim. Res. 2002, 19, 193–212. [Google Scholar] [CrossRef]
- Richter, A.; Adler, G.H.; Fahrak, M.; Eckelmann, W. Erläuterungen zur Nutzungsdifferenzierten Bodenübersichtskarte der Bundesrepublik Deutschland im Maßstab 1:1.000.000 (BÜK1000N, Version 2.3); Bundesanstalt für Geowissenschaften und Rohstoffe: Hannover, Germany, 2007. [Google Scholar]
- Ahrends, B. Ableitung der Nährstoffziffer Anhand von Bodenphysikalischen und Bodenchemischen Kenngrößen. Available online: http://www.dss-wuk.de/hintergrund/modelle/naehrstoffmodell/ (accessed on 15 October 2016).
- Albert, M.; Hansen, J.; Nagel, J.; Schmidt, M.; Spellmann, H. Assessing risks and uncertainties in forest dynamics under different management scenarios and climate change. For. Ecosyst. 2015, 2, 14. [Google Scholar] [CrossRef]
- Niedersächsisches Forstplanungsamt. Langfristige Ökologische Waldentwicklung—Richtlinien zur Baumartenwahl; Niedersächsisches Ministerium für den ländlichen Raum, Ernährung, Landwirtschaft und Verbraucherschutz: Hannover, Germany, 2004. [Google Scholar]
- Füssel, H.-M.; Klein, R.J.T. Climate Change Vulnerability Assessments: An Evolution of Conceptual Thinking. Clim. Chang. 2006, 75, 301–329. [Google Scholar] [CrossRef]
- Spellmann, H.; Sutmöller, J.; Meesenburg, H. Risikovorsorge im Zeichen des Klimawandels. AFZ-DerWald 2007, 23, 1246–1249. [Google Scholar]
- Yousefpour, R.; Hanewinkel, M. Modelling of forest conversion planning with an adaptive simulation-optimization approach and simultaneous consideration of the values of timber, carbon and biodiversity. Ecol. Econ. 2009, 68, 1711–1722. [Google Scholar] [CrossRef]
- Spittlehouse, D.L.; Stewart, R.B. Adaptation to climate change in forest management. J. Ecosyst. Manag. 2003, 4, 1–11. [Google Scholar]
- Yousefpour, R.; Hanewinkel, M.; Moguédec, G. Le Evaluating the Suitability of Management Strategies of Pure Norway Spruce Forests in the Black Forest Area of Southwest Germany for Adaptation to or Mitigation of Climate Change. Environ. Manag. 2010, 45, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Lafond, V.; Lagarrigues, G.; Cordonnier, T.; Courbaud, B. Uneven-aged management options to promote forest resilience for climate change adaptation: Effects of group selection and harvesting intensity. Ann. For. Sci. 2014, 71, 173–186. [Google Scholar] [CrossRef]
- Lindner, M. Developing adaptive forest management strategies to cope with climate change. Tree Physiol. 2000, 20, 299–307. [Google Scholar] [CrossRef]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Alveteg, M.; Walse, C.; Warfvinge, P. Reconstructing Historic Atmospheric Deposition and Nutrient Uptake from Present Day Values Using MAKEDEP. Water Air Soil Pollut. 1998, 104, 269–283. [Google Scholar] [CrossRef]
- Albert, M.; Schmidt, M. Climate-sensitive modelling of site-productivity relationships for Norway spruce (Picea abies (L.) Karst.) and common beech (Fagus sylvatica L.). For. Ecol. Manag. 2010, 259, 739–749. [Google Scholar] [CrossRef]
- Hauck, M.; Zimmermann, J.; Jacob, M.; Dulamsuren, C.; Bade, C.; Ahrends, B.; Leuschner, C. Rapid recovery of stem increment in Norway spruce at reduced SO 2 levels in the Harz Mountains, Germany. Environ. Pollut. 2012, 164, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Plant CO2 responses: An issue of definition, time and resource supply. New Phytol. 2006, 172, 393–411. [Google Scholar] [CrossRef] [PubMed]
- Böckmann, T.; Spellmann, H.; Hüsing, F. Neukonzeption und Weiterentwicklung der Forsteinrichtung in den Niedersächsischen Landesforsten. Forst Holz 1998, 53, 298–302. [Google Scholar]
- Spellmann, H. Sicherung einer nachhaltigen Waldentwicklung auf überbetrieblichen Ebenen. Forstwiss. Cbl 2003, 122, 250–257. [Google Scholar] [CrossRef]
- Spiecker, H. Silvicultural management in maintaining biodiversity and resistance of forests in Europe—Temperate zone. J. Environ. Manag. 2003, 67, 55–65. [Google Scholar] [CrossRef]
- Schou, E.; Jellesmark, B.; Bredahl, J. Regeneration decisions in forestry under climate change related uncertainties and risks: Effects of three different aspects of uncertainty. For. Policy Econ. 2015, 50, 11–19. [Google Scholar] [CrossRef]
- Lévesque, M.; Saurer, M.; Siegwolf, R.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway spruce and European larch. Glob. Chang. Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, M.; Rigling, A.; Bugmann, H.; Weber, P.; Brang, P. Growth response of five co-occurring conifers to drought across a wide climatic gradient in Central Europe. Agric. For. Meteorol. 2014, 197, 1–12. [Google Scholar] [CrossRef]
- Lévesque, M.; Walthert, L.; Weber, P. Soil nutrients influence growth response of temperate tree species to drought. J. Ecol. 2016, 104, 377–387. [Google Scholar] [CrossRef]
- Bredemeier, M.; Lamersdorf, N.; Schulte-Bisping, H.; von Lüpke, B. Risk appraisal for forest management with respect to site quality and environmental changes. In Risk Analysis in Forest Management; von Gadow, K., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Roloff, A.; Grundmann, B.M. Waldbaumarten und ihre Verwendung im Klimawandel. Arch. Forstwes. Landsch. Oekol. 2008, 42, 97–109. [Google Scholar]
- Zang, C.; Rothe, A.; Weis, W.; Pretzsch, H. Zur Baumarteneignung bei Klimawandel: Ableitung der Trockenstress-Anfälligkeit wichtiger Waldbaumarten aus Jahrringbreiten. Allg. Forst Jagdztg. 2011, 182, 98–112. [Google Scholar]
- Kätzel, R.; Zander, M.; Löffler, S. Wenn der Durst kommt—Eiche, Buche, Kiefer und Douglasie im Trockenstresstest. Eberswalder Forstl. Schriftenr. 2015, 59, 40–54. [Google Scholar]
- Müller, J. Effects of drought on the water balance of selected forest sites and the evaluation of a drought risk. In Schriften aus der Forstlichen Fakultät Göttingen und der Niedersächsischen Forstlichen Versuchsanstalt; Sauerländer’s Verlag: Bad Orb, Germany, 2006; pp. 142–148. [Google Scholar]
- Wellpott, A.; Imbry, F.; Schindler, D.; Mayer, H. Simulation of drought for a Scots pine forest (Pinus sylvestris L.) in the southern upper Rhine plain. Meteorol. Z. 2005, 14, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- Adams, H.D.; Kolb, T.E. Tree growth response to drought and temperature in a mountain landscape in northern Arizona, USA. J. Biogeogr. 2005, 32, 1629–1640. [Google Scholar] [CrossRef]
- Klos, R.J.; Wang, G.G.; Bauerle, W.L.; Rieck, J.R. Drought impact on forest growth and mortality in the southeast USA: An analysis using Forest Health and Monitoring data. Ecol. Appl. 2009, 19, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Zingg, A.; Bürgi, A. Trockenperioden seit 1900 und Waldwachstum: Eine Analyse langfristiger Datenreihen. Schweiz. Z. Forstwes. 2008, 159, 352–361. [Google Scholar] [CrossRef]
- Hogg, E.H.T.; Brandt, J.P.; Michaelian, M. Impacts of a regional drought on the productivity, dieback, and biomass of western Canadian aspen forests. Can. J. For. Res. 2008, 38, 1373–1384. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Gazol, A.; Camarero, J.J.; Anderegg, W.R.L.; Vincente-Serrano, S.M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Glob. Ecol. Biogeogr. 2017, 26, 166–176. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Walker, W.E.; Harresmoes, P.; Rotmans, J.; van der Sluijs, J.P.; van Asselt, M.B.; Janssen, P.; Krayer von Krauss, M.P. Defining Uncertainty: A Conceptual Basis for Uncertainty Management. Integr. Assess. 2003, 4, 5–17. [Google Scholar] [CrossRef]
- Wilby, R.L.; Dessai, S. Robust adaptation to climate change. Weather 2010, 65, 176–180. [Google Scholar] [CrossRef]
- Hawkins, E.; Sutton, R. The potential to narrow unceratinty in regional climate predictions. Am. Meteorol. Soc. 2009, 1095–1107. [Google Scholar] [CrossRef]
- Jones, R. Managing uncertainty in climate change projections—Issues for impact assessment. Clim. Chang. 2000, 45, 403–419. [Google Scholar] [CrossRef]
- Knutti, R. The end of model democracy? Clim. Chang. 2010, 102, 395–404. [Google Scholar] [CrossRef]
- Seidl, R.; Lexer, M.J. Forest management under climatic and social uncertainty: Trade-offs between reducing climate change impacts and fostering adaptive capacity. J. Environ. Manag. 2013, 114, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Booth, B.B.B.; Harris, G.R.; Murphy, J.M.; Sexton, D.M.H.; Webb, M.J. Towards quantifying uncertainty in transient climate change. Clim. Dyn. 2006, 27, 127–147. [Google Scholar] [CrossRef]
- Dittrich, R.; Wreford, A.; Moran, D. A survey of decision-making approaches for climate change adaptation: Are robust methods the way forward? Ecol. Econ. 2016, 122, 79–89. [Google Scholar] [CrossRef]
- Hallegatte, S. Strategies to adapt to an uncertain climate change. Glob. Environ. Chang. 2009, 19, 240–247. [Google Scholar] [CrossRef]
- Plattner, T.M. Risikoaversion als Relevanter Faktor der Risikobewertung von Naturgefahren; ETH Zürich: Zürich, Switzerland, 2006. [Google Scholar]
- Hanewinkel, M.; Hummel, S.; Albrecht, A. Assessing natural hazards in forestry for risk management: A review. Eur. J. For. Res. 2011, 130, 329–351. [Google Scholar] [CrossRef]
- Yousefpour, R.; Temperli, C.; Bugmann, H.; Elkin, C.; Hanewinkel, M.; Meilby, H.; Jacobsen, J.B.; Thorsen, B.J. Updating beliefs and combining evidence in adaptive forest management under climate change: A case study of Norway spruce (Picea abies L. Karst ) in the Black Forest, Germany. J. Environ. Manag. 2013, 122, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, T.; Zacharias, D. PNV-Karten für Niedersachsen auf Basis der BÜK50; Niedersächsisches Landesamt für Ökologie: Hannover, Germany, 2003. [Google Scholar]
- Bundesamt für Naturschutz. Karte der Potentiellen Natürlichen Vegetation Deutschlands; BfN: Bonn, Germany, 2011. [Google Scholar]
- Cheaib, A.; Badeau, V.; Boe, J.; Chuine, I.; Delire, C.; Dufrene, E.; Francois, C.; Gritti, E.S.; Legay, M.; Page, C.; et al. Climate change impacts on tree ranges: Model intercomparison facilitates understanding and quantification of uncertainty. Ecol. Lett. 2012, 15, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Yousefpour, R.; Jacobsen, J.B.; Thorsen, B.J.; Meilby, H.; Hanewinkel, M.; Oehler, K. A review of decision-making approaches to handle uncertainty and risk in adaptive forest management under climate change. Ann. For. Sci. 2012, 69, 1–15. [Google Scholar] [CrossRef]
- Yousefpour, R.; Hanewinkel, M. Climate Change and Decision-Making under Uncertainty. Curr. For. Rep. 2016, 2, 143–149. [Google Scholar] [CrossRef]
- Spathelf, P.; Bolte, A.; van der Maaten, E. Is Close-to-Nature Silviculture (CNS) an adequate concept to adapt forests to climate change? Landbauforschung 2015, 65, 161–170. [Google Scholar]
Figure 1.
The two study regions Uelzen and Fläming in the North German Plain near the cities of Hamburg and Berlin (land cover data in the overview map © ESA/ESA GlobCorine Project, led by Université Catholique de Louvain).
Figure 1.
The two study regions Uelzen and Fläming in the North German Plain near the cities of Hamburg and Berlin (land cover data in the overview map © ESA/ESA GlobCorine Project, led by Université Catholique de Louvain).
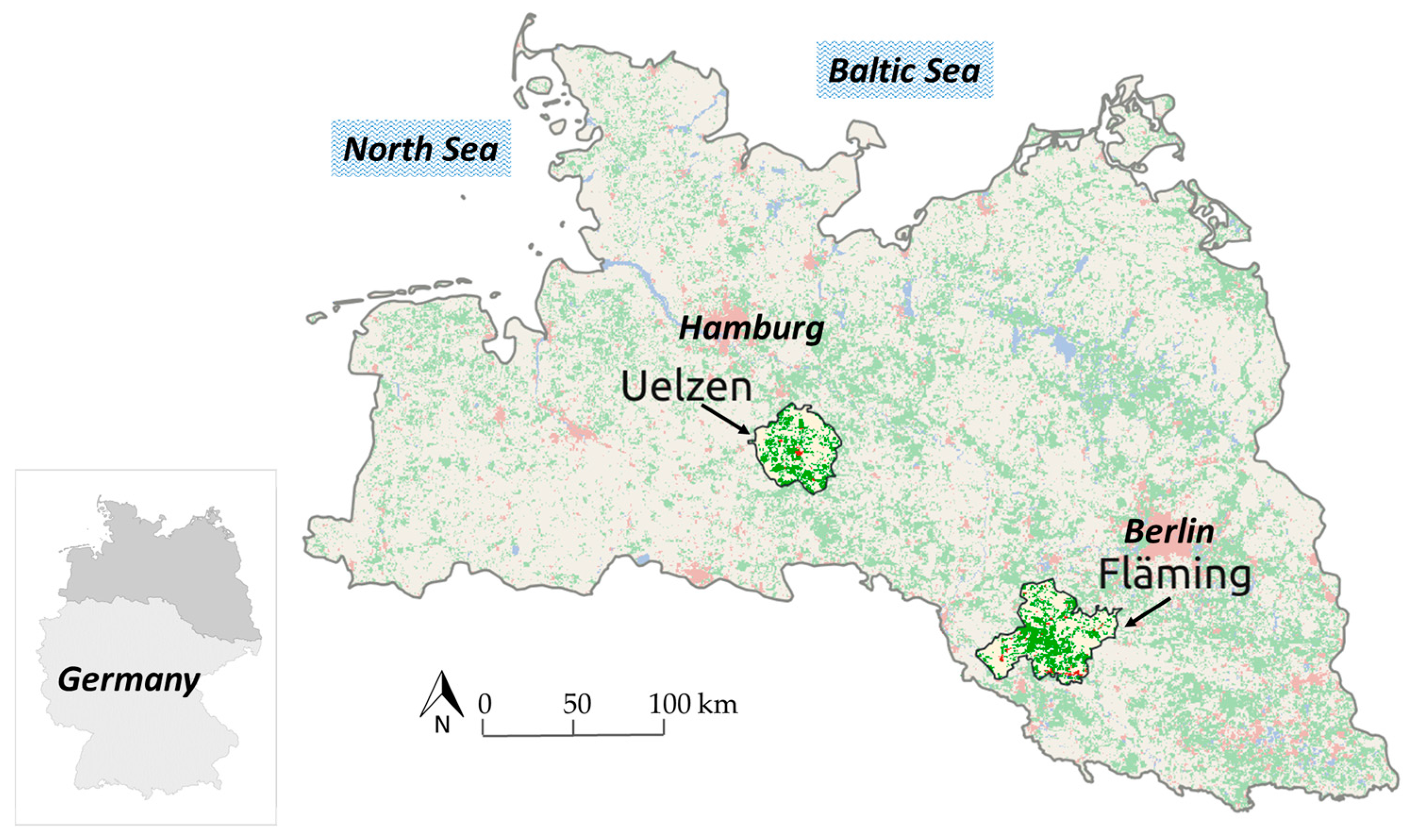
Figure 2.
Site water budget over time projected with STARS using RCP 8.5 under minimum, median and maximum climate scenarios in Uelzen (left) and Fläming (right). The values are means of all individual sites.
Figure 2.
Site water budget over time projected with STARS using RCP 8.5 under minimum, median and maximum climate scenarios in Uelzen (left) and Fläming (right). The values are means of all individual sites.
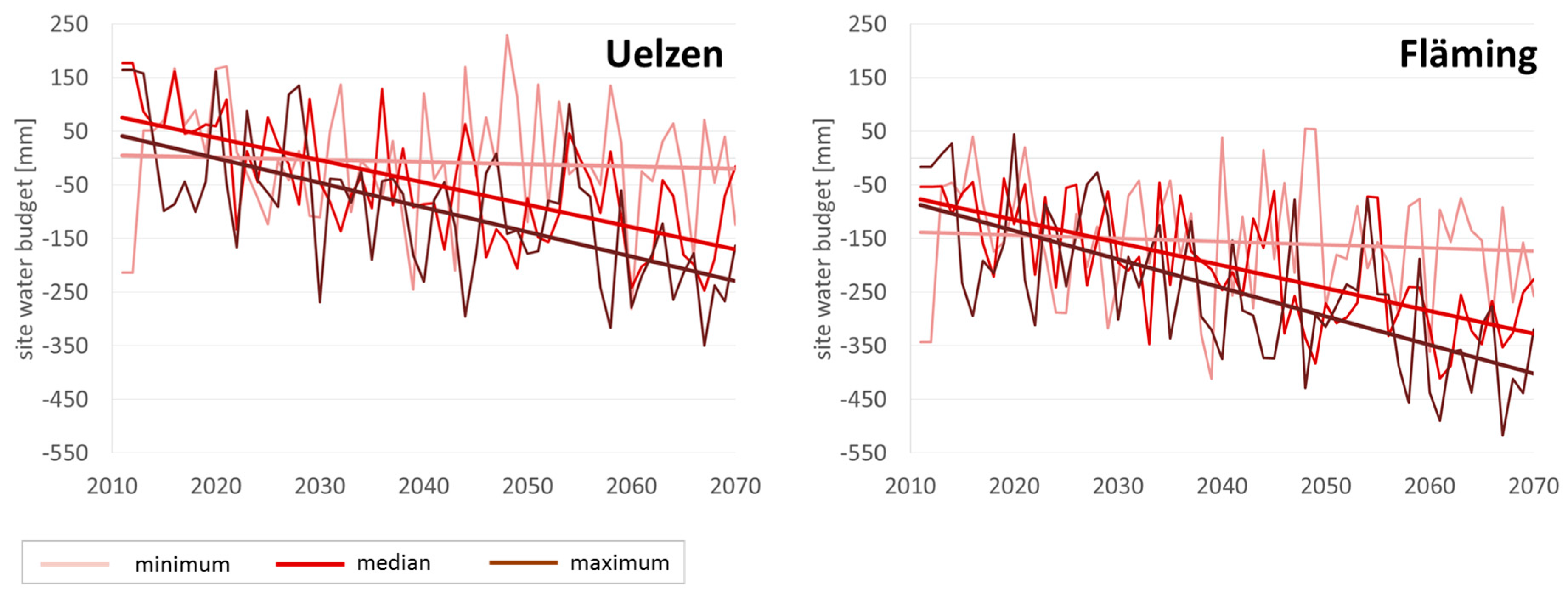
Figure 3.
Proportion of sites according to soil nutrient classes and site water budget in growing season classes in Uelzen and Fläming (top heatmaps for the period 1991 to 2010; three rows of heatmaps below for site water budget conditions in the period 2051 to 2070 under three climate scenarios of minimum, median and maximum; only classes with proportions greater than 1% are listed).
Figure 3.
Proportion of sites according to soil nutrient classes and site water budget in growing season classes in Uelzen and Fläming (top heatmaps for the period 1991 to 2010; three rows of heatmaps below for site water budget conditions in the period 2051 to 2070 under three climate scenarios of minimum, median and maximum; only classes with proportions greater than 1% are listed).
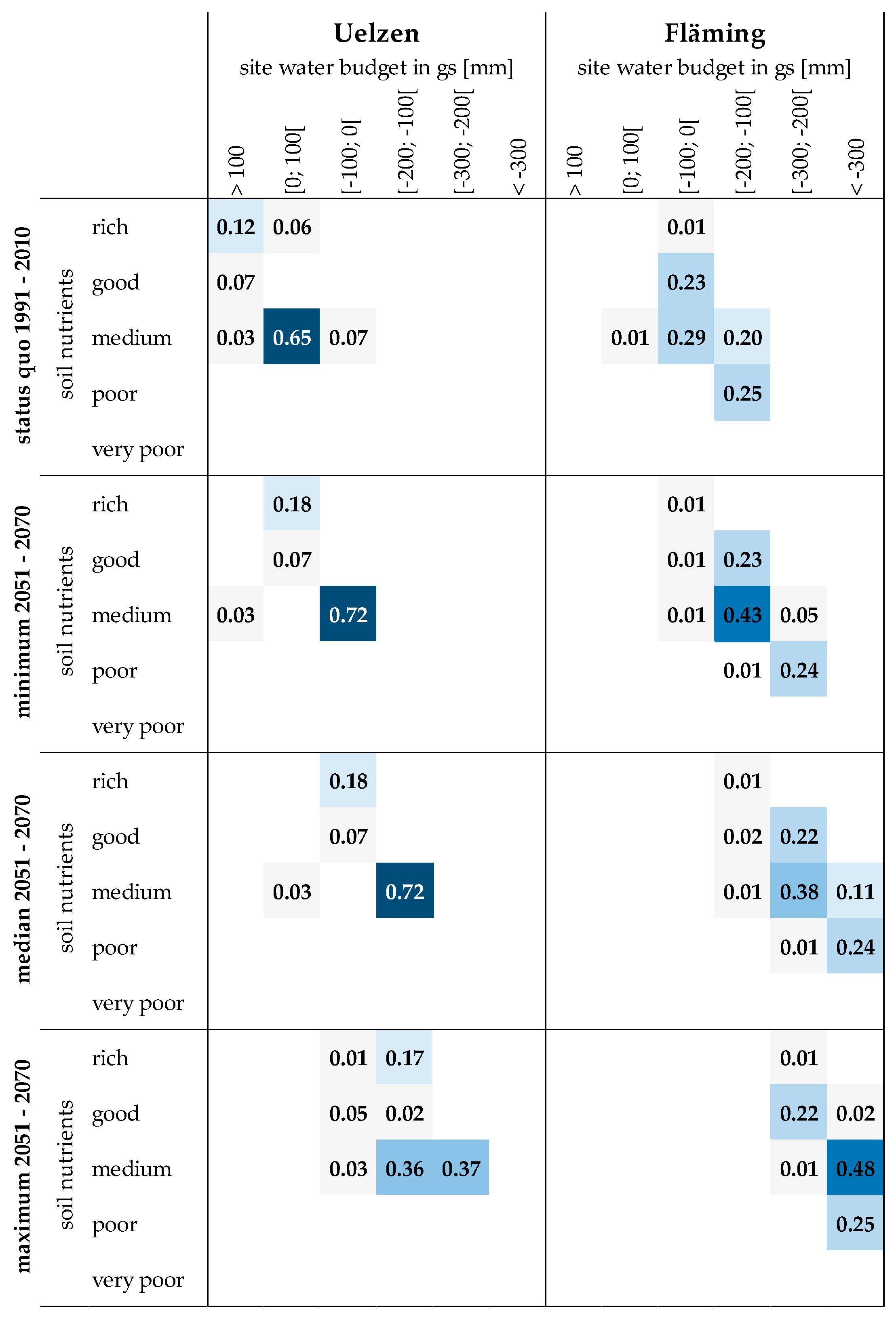
Figure 4.
Simplified ecogram to assign species based on a matrix of soil nutrients and site water budget in the North German Plain (BLHV: broadleaves of high value; EB: European beech; O: oak species; EL: European larch; DF: Douglas-fir; NS: Norway spruce; SP: Scots pine).
Figure 4.
Simplified ecogram to assign species based on a matrix of soil nutrients and site water budget in the North German Plain (BLHV: broadleaves of high value; EB: European beech; O: oak species; EL: European larch; DF: Douglas-fir; NS: Norway spruce; SP: Scots pine).

Figure 5.
Frequency distribution of current stand type area (dark blue) and succeeding stand type area planned under constant site conditions (stage 1) in 2070 (light blue) under three silvicultural management strategies in Uelzen (top row) and Fläming (bottom row).
Figure 5.
Frequency distribution of current stand type area (dark blue) and succeeding stand type area planned under constant site conditions (stage 1) in 2070 (light blue) under three silvicultural management strategies in Uelzen (top row) and Fläming (bottom row).
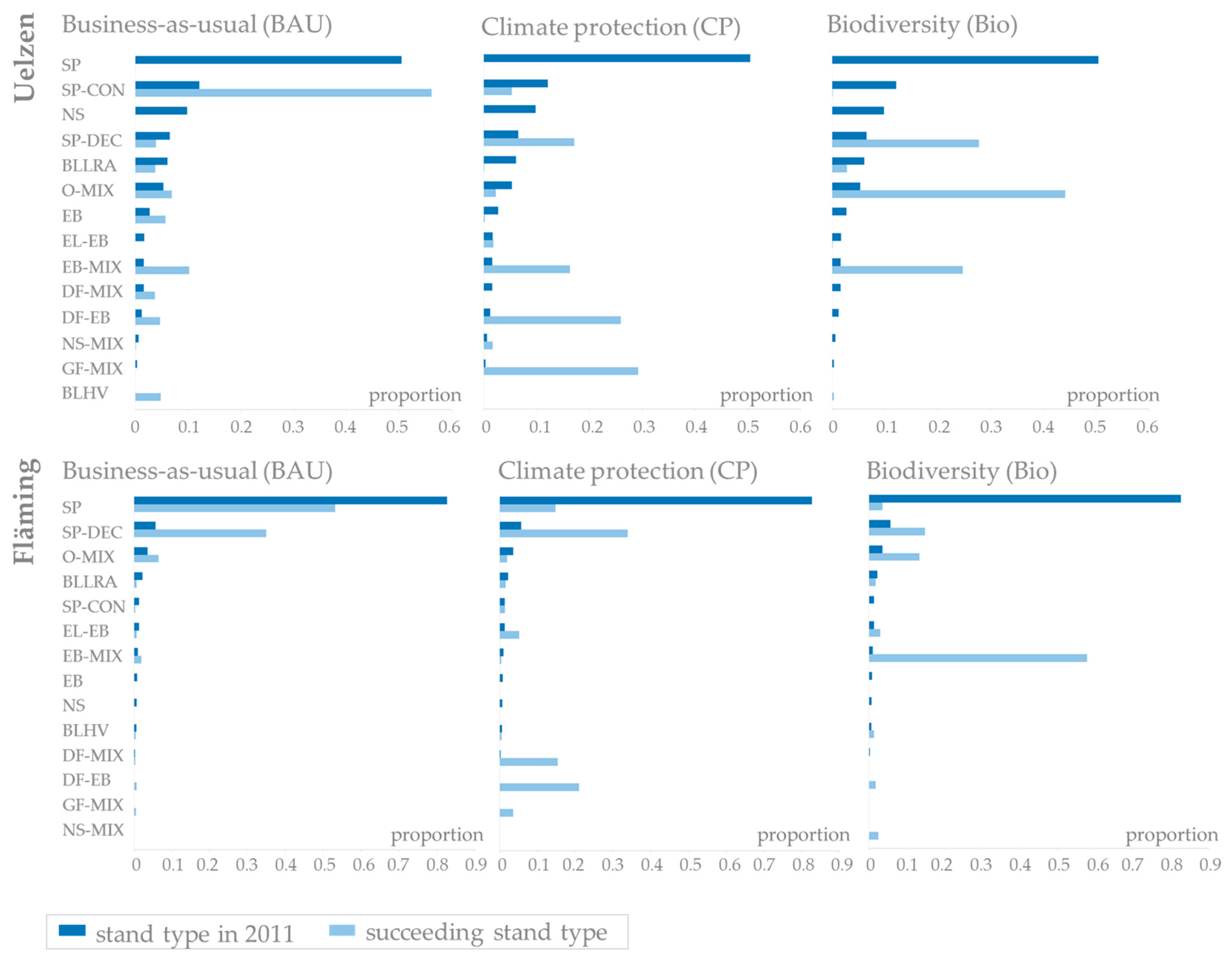
Figure 6.
Frequency distribution of succeeding stand type area under constant site conditions in 2070 (light blue) and adaptive planning (stage 2) considering drought restrictions under minimum (light gray), median (gray) and maximum (dark gray) climate scenarios under three silvicultural management strategies in Uelzen.
Figure 6.
Frequency distribution of succeeding stand type area under constant site conditions in 2070 (light blue) and adaptive planning (stage 2) considering drought restrictions under minimum (light gray), median (gray) and maximum (dark gray) climate scenarios under three silvicultural management strategies in Uelzen.
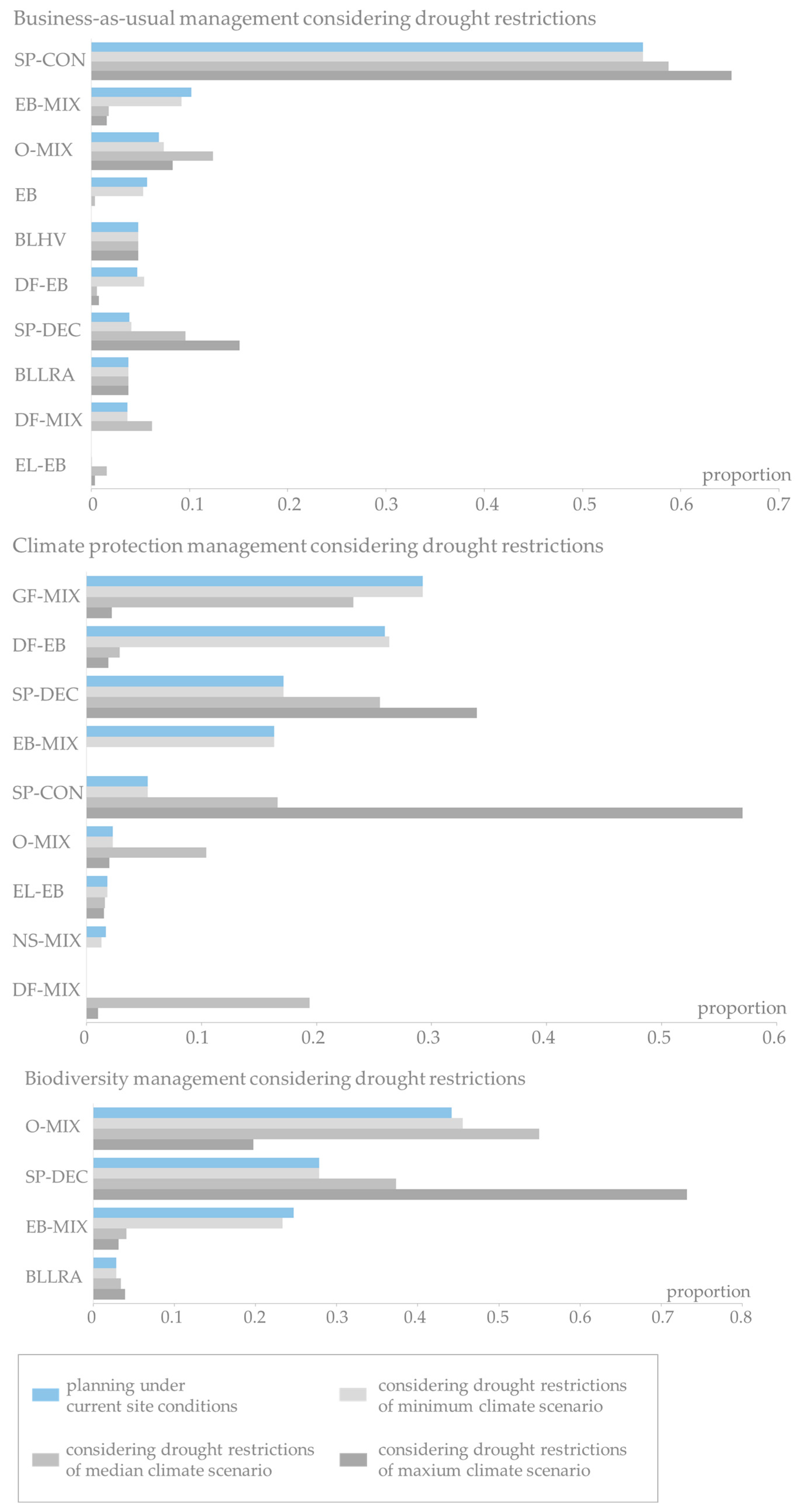
Figure 7.
Frequency distribution of succeeding stand type area under constant site conditions in 2070 (light blue) and adaptive planning (stage 2) considering drought restrictions under minimum (light gray), median (gray) and maximum (dark gray) climate scenarios under three silvicultural management strategies in Fläming.
Figure 7.
Frequency distribution of succeeding stand type area under constant site conditions in 2070 (light blue) and adaptive planning (stage 2) considering drought restrictions under minimum (light gray), median (gray) and maximum (dark gray) climate scenarios under three silvicultural management strategies in Fläming.
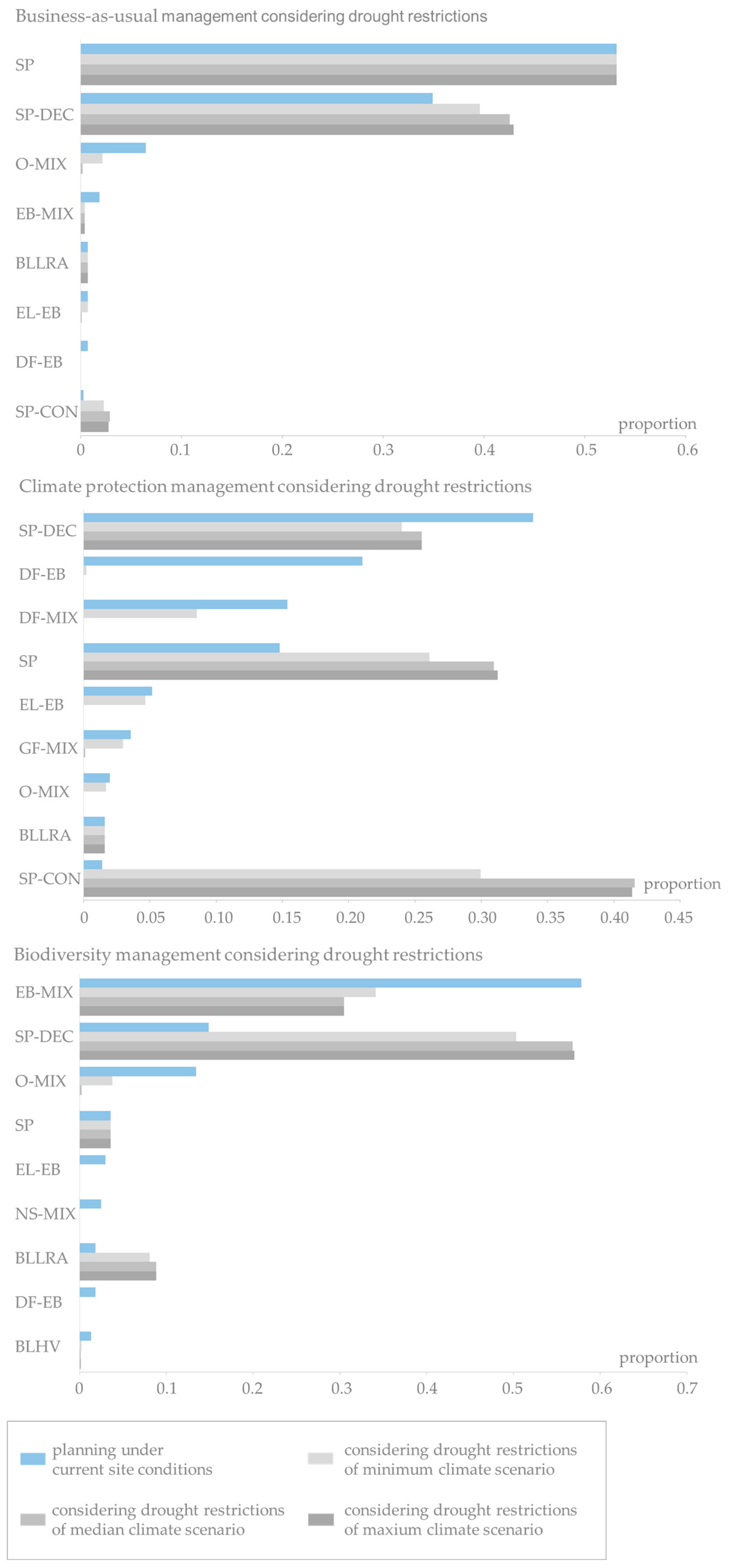

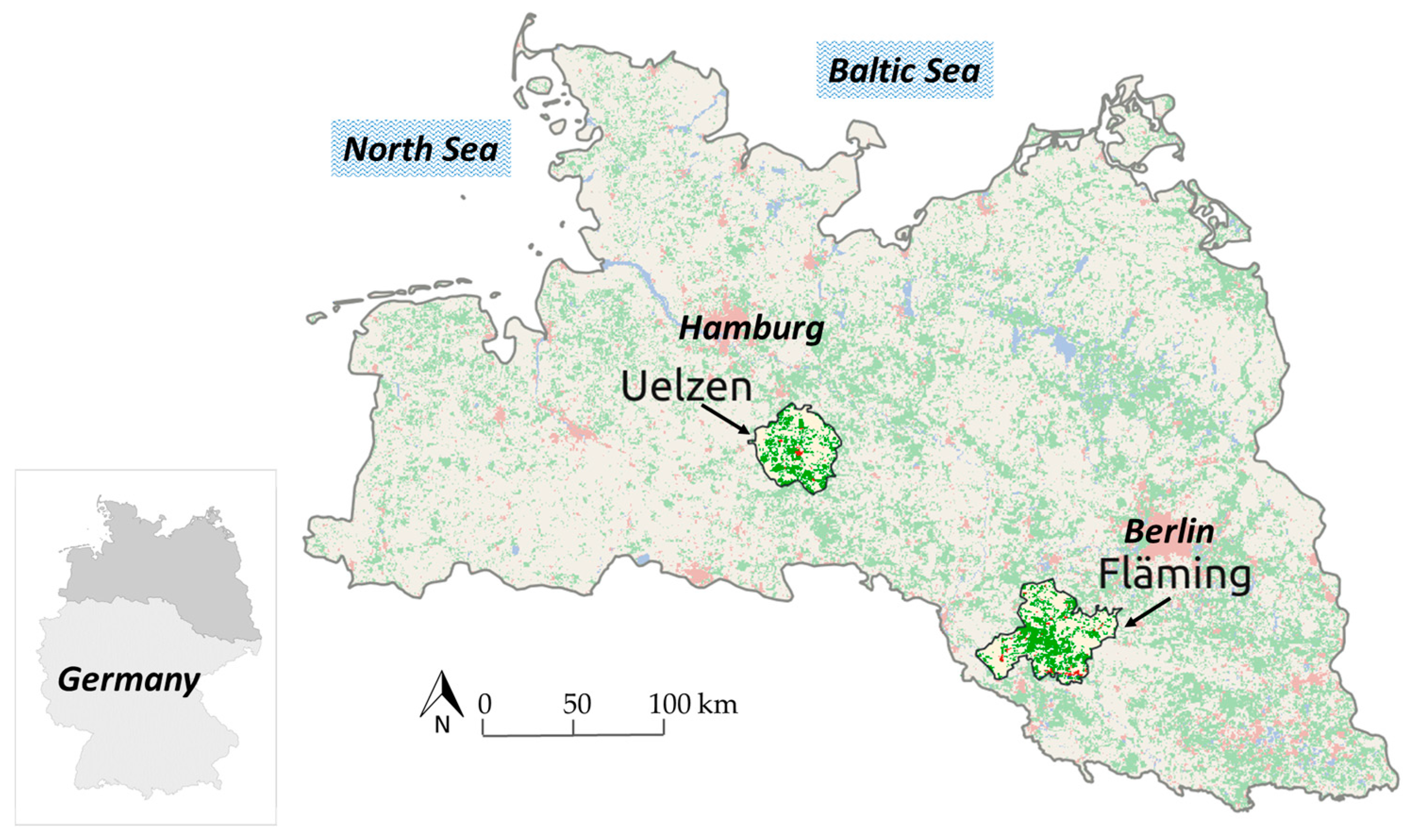{kind=link}
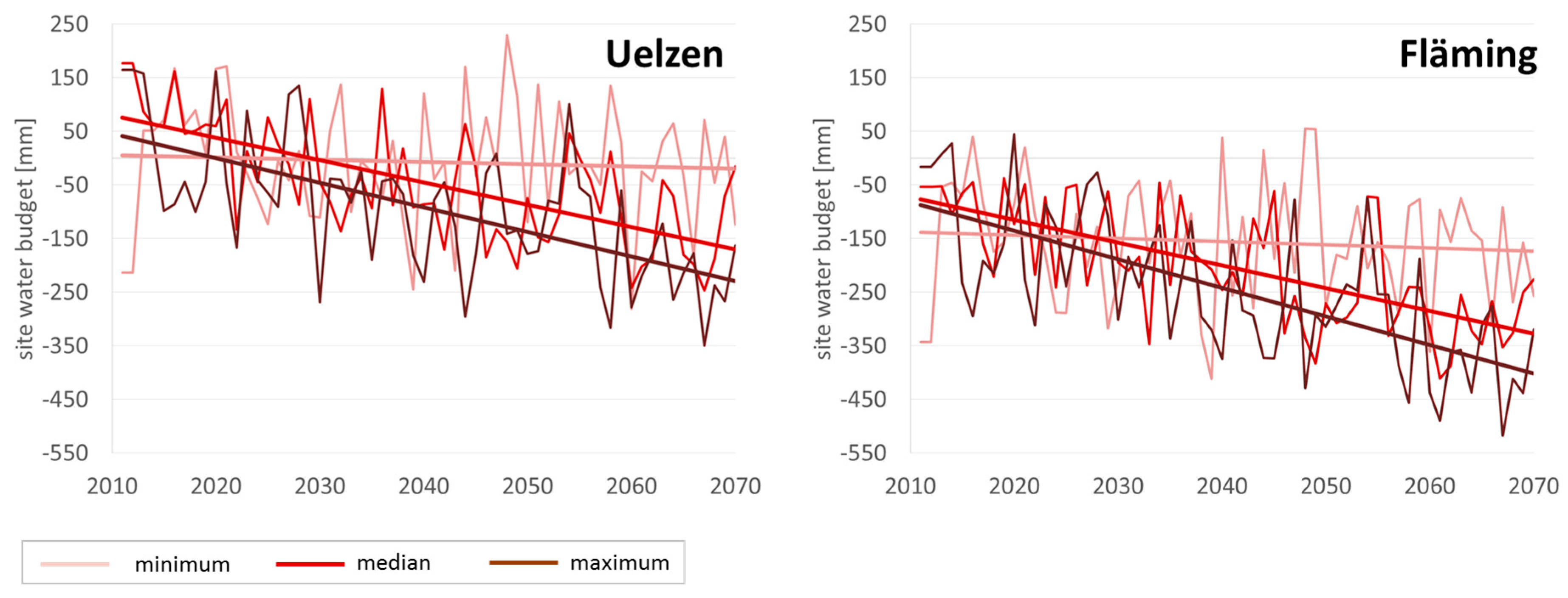{kind=link}
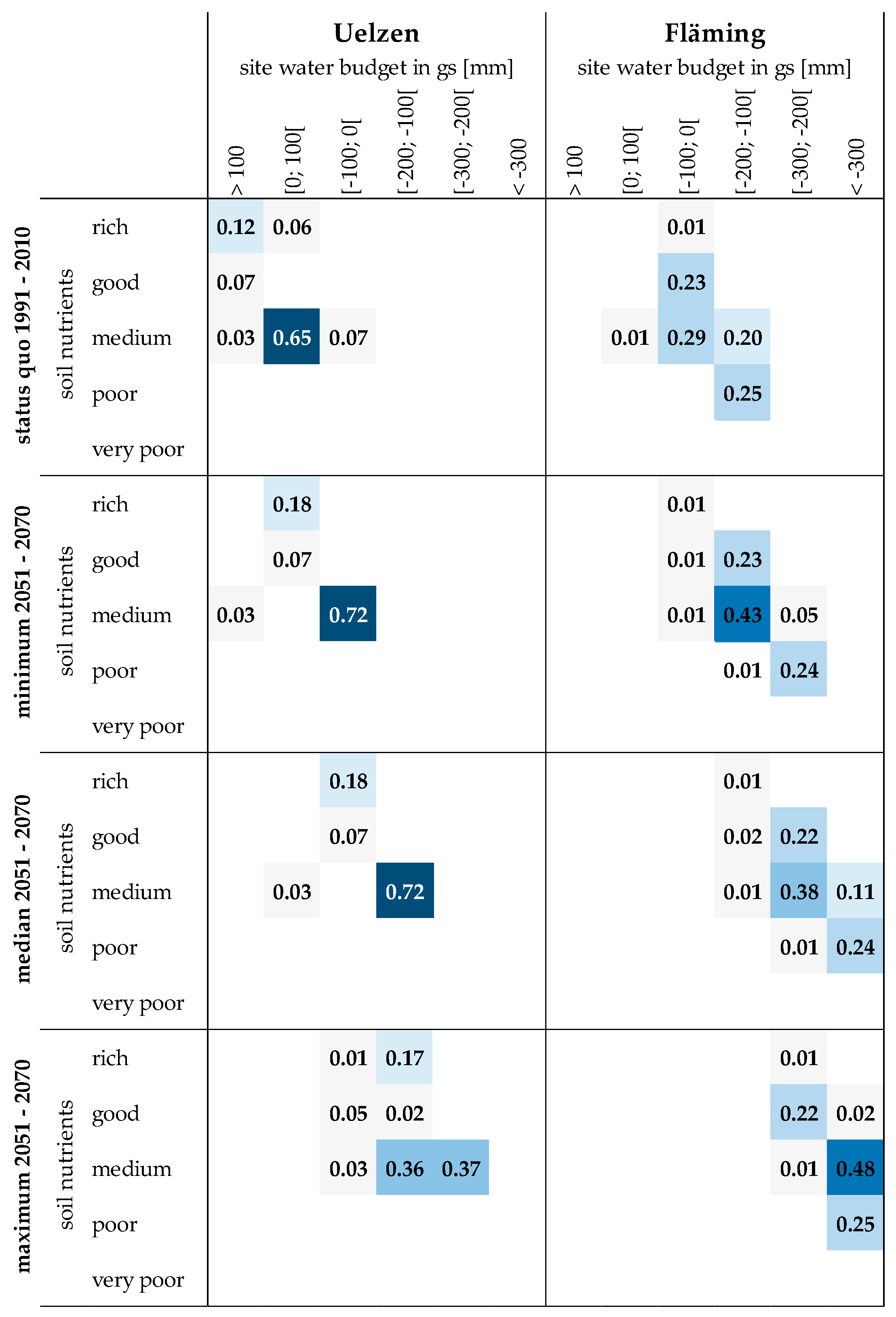{kind=link}
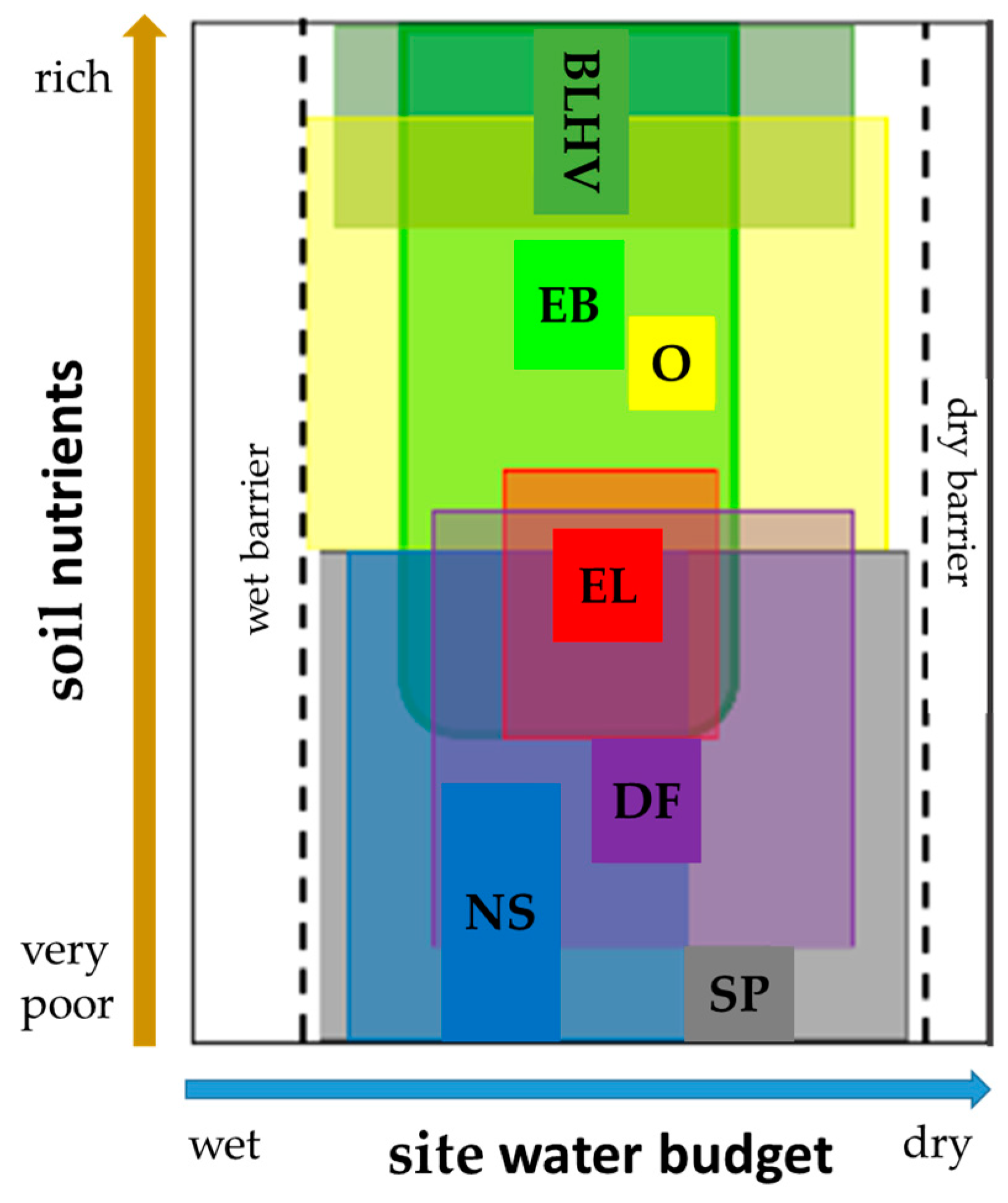{kind=link}
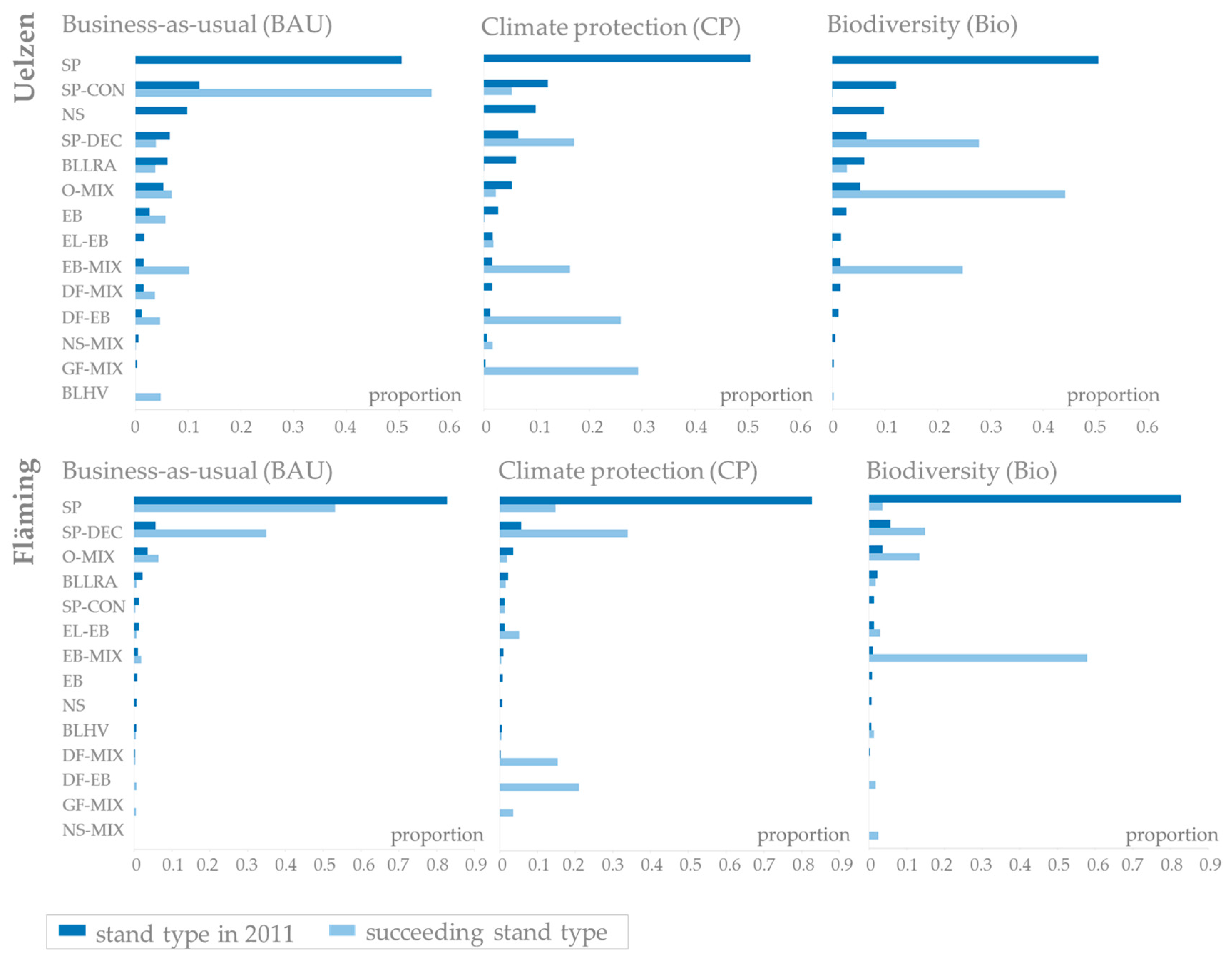{kind=link}
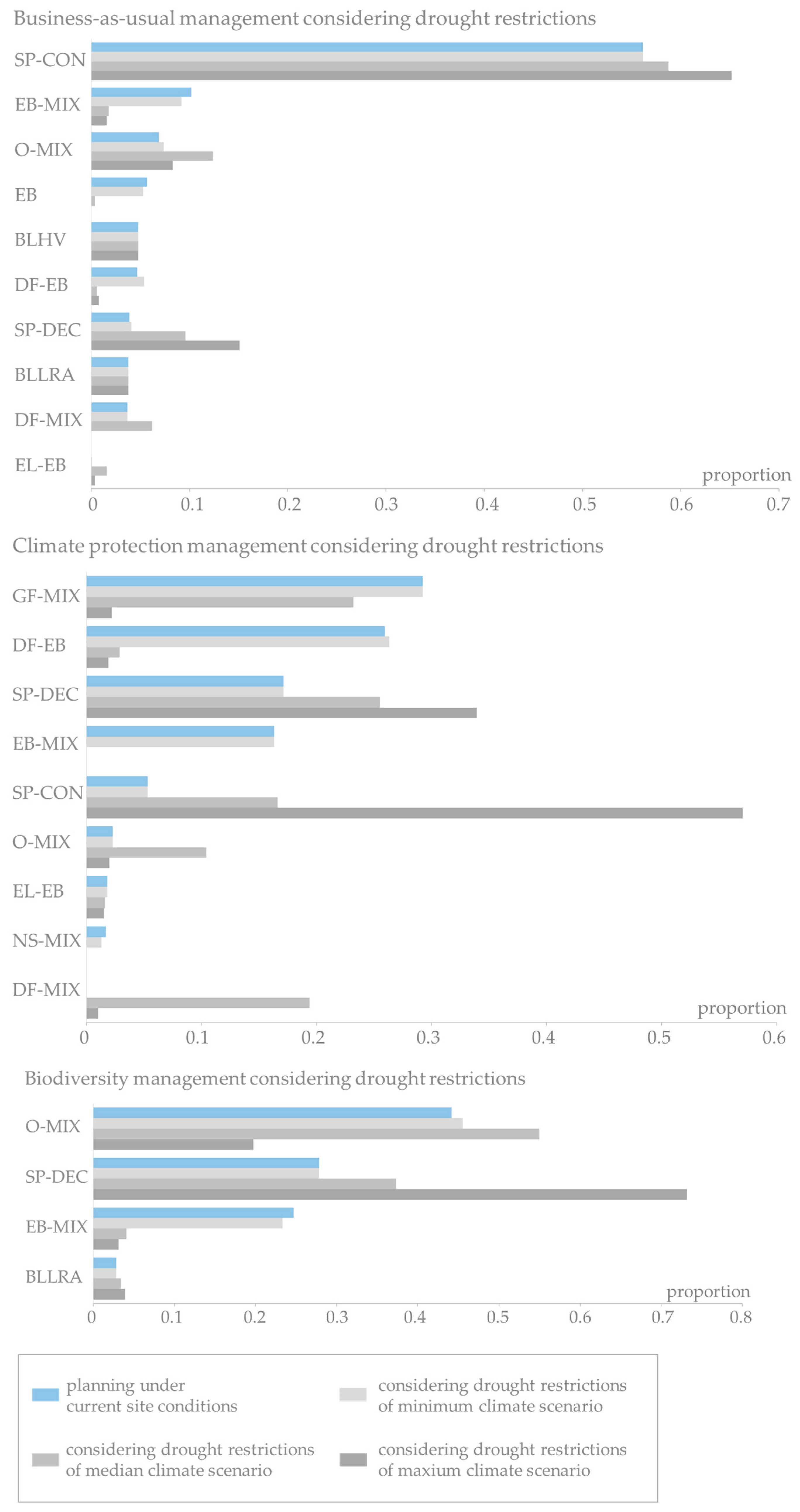{kind=link}
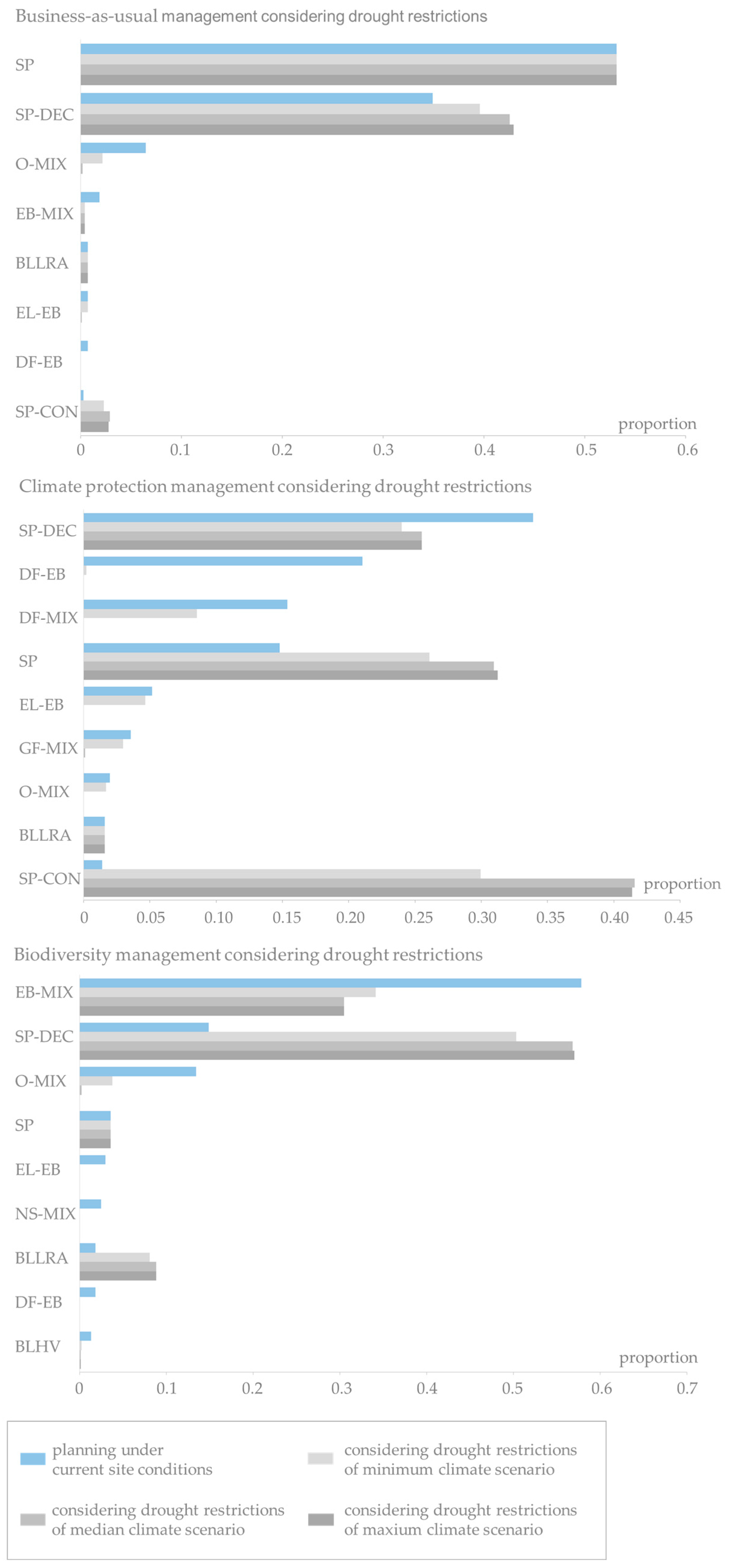{kind=link}
Table 1.
Proportions of species area [%] in Uelzen and Fläming in 2011 according to available inventory data (BLHV = broadleaves of high value; BLLRA = broadleaves of low rotation age).
Table 1.
Proportions of species area [%] in Uelzen and Fläming in 2011 according to available inventory data (BLHV = broadleaves of high value; BLLRA = broadleaves of low rotation age).
| European Beech | Oak | BLHV | BLLRA | Norway Spruce | Scots Pine | Douglas-Fir | European Larch | Grand Fir | |
|---|---|---|---|---|---|---|---|---|---|
| Uelzen | 4.8 | 6.1 | 0.7 | 8.0 | 13.3 | 61.3 | 3.9 | 1.9 | - |
| Fläming | 12.8 | 5.9 | 1.0 | 3.3 | 1.8 | 73.5 | 0.4 | 1.0 | 0.2 |
Table 2.
Site characteristics in Uelzen and Fläming (climate values as averages in the period 1991 to 2010; asm: available soil moisture calculated for 1 m soil depth, annual temp.: annual mean temperature; annual prec.: total annual precipitation; cwbgs: climatic water balance (=precipitation − evapotranspiration for grass) in growing season).
Table 2.
Site characteristics in Uelzen and Fläming (climate values as averages in the period 1991 to 2010; asm: available soil moisture calculated for 1 m soil depth, annual temp.: annual mean temperature; annual prec.: total annual precipitation; cwbgs: climatic water balance (=precipitation − evapotranspiration for grass) in growing season).
| Asm [mm] | Annual Temp. [°C] | Annual Prec. [mm] | cwbgs [mm] | |
|---|---|---|---|---|
| Uelzen | 110 | 9.2 | 727 | −65 |
| Fläming | 123 | 9.8 | 572 | −216 |
Table 3.
Definition of drought vulnerability classes for five major tree species and corresponding planning measures according to [28,53].
| Drought Vulnerability | Site Water Budget in Growing Season [mm] | Planning Measures | |||
|---|---|---|---|---|---|
| Norway Spruce | European Beech | Oak/Douglas-Fir | Scots Pine | ||
| low | >0 | >−25 | >−150 | >−180 | primary species |
| medium | 0 to −80 | −25 to −100 | −150 to −400 | −180 to −450 | admixture |
| high | <−80 | <−100 | <−400 | <−450 | no cultivation |
Table 4.
Abbreviations for stand types used throughout the text.
| SP | Scots pine (Pinus sylvestris L.) |
| SP-CON | Scots pine-conifer mixture |
| SP-DEC | Scots pine-deciduous species mixture |
| NS | Norway spruce (Picea abies L.) |
| NS-MIX | Norway spruce mixture |
| DF-MIX | Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco)-mixture |
| DF-EB | Douglas-fir-European beech mixture |
| GF-MIX | Grand fir (Abies grandis (Douglas ex D. Don) Lindl.)-Douglas-fir mixture |
| EB | European beech (Fagus sylvatica) |
| EL-EB | European larch (Larix decidua Mill.)-European beech mixture |
| EB-MIX | European beech mixture |
| BLLRA | broadleaves of low rotation age |
| BLHV | broadleaves of high value |
| O-MIX | oak (Quercus petraea (Matt.) Liebl. and Quercus robur L.)-mixture |
Table 5.
Adaptation effort score (AES; see Section 2.5 for definition) under the forest management strategies business-as-usual (BAU), climate protection (CP) and biodiversity management (BIO) applying the climate scenarios of minimum, median and maximum run in Uelzen and Fläming. The uncertainty range is the difference between the AES values of the maximum and the minimum climate run.
Table 5.
Adaptation effort score (AES; see Section 2.5 for definition) under the forest management strategies business-as-usual (BAU), climate protection (CP) and biodiversity management (BIO) applying the climate scenarios of minimum, median and maximum run in Uelzen and Fläming. The uncertainty range is the difference between the AES values of the maximum and the minimum climate run.
| Climate Scenario | Uncertainty | ||||
|---|---|---|---|---|---|
| Minimum | Median | Maximum | Range | ||
| Uelzen | BAU | 2.0 | 17.9 | 21.8 | 19.8 |
| CP | 0.3 | 31.5 | 54.8 | 54.5 | |
| BIO | 3.2 | 48.5 | 74.1 | 70.9 | |
| Fläming | BAU | 12.2 | 14.9 | 15.1 | 2.9 |
| CP | 28.2 | 40.4 | 40.6 | 12.4 | |
| BIO | 50.5 | 56.1 | 56.2 | 5.7 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Albert, M.; Nagel, R.-V.; Nuske, R.S.; Sutmöller, J.; Spellmann, H. Tree Species Selection in the Face of Drought Risk—Uncertainty in Forest Planning. Forests 2017, 8, 363. https://doi.org/10.3390/f8100363
AMA Style
Albert M, Nagel R-V, Nuske RS, Sutmöller J, Spellmann H. Tree Species Selection in the Face of Drought Risk—Uncertainty in Forest Planning. Forests. 2017; 8(10):363. https://doi.org/10.3390/f8100363
Chicago/Turabian StyleAlbert, Matthias, Ralf-Volker Nagel, Robert S. Nuske, Johannes Sutmöller, and Hermann Spellmann. 2017. "Tree Species Selection in the Face of Drought Risk—Uncertainty in Forest Planning" Forests 8, no. 10: 363. https://doi.org/10.3390/f8100363
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.