Growth and Physiology of Senegalia senegal (L.) Britton Seedlings as Influenced by Seed Origin and Salinity and Fertility Treatments

1
Department of Forest Resources and Environmental Conservation, Virginia Polytechnic and State University, Blacksburg, VA 24061, USA
2
Centre National de Recherche Forestière of Institut Sénégalais de Recherche Agricole, Dakar 2312, Senegal
*
Author to whom correspondence should be addressed.
Forests 2017, 8(10), 388; https://doi.org/10.3390/f8100388
Submission received: 2 August 2017
/
Revised: 25 September 2017
/
Accepted: 3 October 2017
/
Published: 11 October 2017
Abstract
:Multipurpose trees such as Senegalia senegal are widespread in arid and semi-arid lands that have natural or induced saline soils and poor soil fertility. Such environmental problems impact growth and have the potential to influence plant physiological adaptations. Identifying superior genotypes better adapted to these environmental stresses will be of great importance for tree selection for reclamation of degraded drylands. The main objective of this study was to examine the growth performance, and physiological and morphological adaptations to salinity, and fertility treatments of different Senegalia senegal families. We used five families (DB16, DB14, K4B19, K17B19, NB1) selected from 60 families of a Senegalia senegal progeny trial in Dahra, Senegal. Seedlings were grown under greenhouse conditions by watering all plants for three weeks and then stopping all watering for three more weeks. In a randomized complete block design, a two-level factorial combination was used for salinity (zero and 183.1 mM NaCl added) and fertility (zero and 100 kg/ha N-P-K added) treatments. A significant family × salt × fertilizer interaction was found for all biomass parameters (leaf dry matter, stem dry matter, root dry matter, and leaf area). The fertilizer application resulted in a significant increase of total biomass of all families, ranging from 63% to 237% for NB1 and K17B19, respectively. In contrast, salt only decreased total biomass of NB1 and K17B19 increased growth. Despite similar net photosynthetic rates before treatment started, fertilizer and salinity induced different effects between families. Prior to drought stress, fertilizer did not affect photosynthesis of DB16, while salt significantly decreased stomatal conductance of all families. DB16 and N1B1, despite significant differences of stomata size and density, significantly decreased transpiration, and thereby increased their intrinsic water use efficiency. Under drought, relative growth rate was significantly decreased. Given that genotype differences were found, these families and salinity and fertilizer treatments need to be explored in field trials.
1. Introduction
In recent years, frequent drought occurrences and anthropogenic pressures on forest resources in arid and semi-arid lands have heavily impacted species composition [1]. The natural vegetation becomes sparse and progressive soil degradation develops, which accelerates desertification and induces salinization [2,3,4]. Drought and salt tolerant species are seeking for restoration of such affected lands. Senegalia senegal is a drought tolerant species, widely used to control desertification in arid and semi-arid lands. It is particularly exploited because it is the source of many products, including gum arabic which is traded on both local and international markets. Recent studies have revealed its potential tolerance to salinity stress [5,6,7]. However, salt tolerance among local Senegalia senegal provenances has not yet been investigated. Salt-affected lands in Senegal are estimated at about 1.7 million hectares [8]. Due to increasing population, it is likely that, in the future, tree crops and fuel wood plantations will increase on degraded lands in general and particularly on salt affected soils.
In arid and semi-arid areas, salt stress is often combined with water limitations in the context of poor soil management. Afforestation/reforestation plans are often proposed solutions for soil improvement of degraded lands. However, plantation failures can occur when inappropriate species are selected. An understanding of plant response to salinity is of great importance for reclamation of salt affected lands. Yet, strategies to improve salt and/or drought tolerance in such ecosystems require an understanding of tree physiological mechanisms that links their growth and adaptation. Growth characteristics and photosynthesis efficiency are traditionally used to select higher quality family or genotypes more adapted to hostile environments. Stomatal response is among one of the most important factors influencing photosynthetic capacity under limiting environmental factors [9]. Different stomatal characteristics may account partially for different stomatal resistance that consequently impact the photosynthetic rate [10]. However, little attention has been given to the leaf physiology of Senegalia senegal, and particularly stomata characteristics. Under water stress, stomatal density is known to impact leaf photosynthesis and water use efficiency [11,12]. Reduced stomatal density can be advantageous under saline stress [13,14]. Drought and salinity stress both induce a reduced photosynthetic rate through stomata and mesophyll restriction [15,16]. Furthermore, selection of adapted genotypes based on leaf traits and water use efficiency to such environments are highly recommended.
This study investigates adaptive features in Senegalia senegal from different families which may allow them to grow better in both saline and dry conditions. The main objective is to examine physiological and morphological changes among Senegalia senegal provenances subjected to salinity and fertility treatments. We tested the hypothesis that salinity tolerance and fertilizer effect on the physiology and biomass accumulation of Senegalia senegal seedlings are influenced by seed origin. We expected that leaf gas exchanges, growth and biomass accumulation will decrease under the effect of salinity whereas the fertilizer may increase those potentials. We are interested in seeking genotypes that perform best in terms of dry matter accumulation and intrinsic water use efficiency under well-watered and then drought conditions, and whether stomatal characteristics explain potential changes among families and environmental conditions.
2. Materials and Methods
2.1. Plant Material
Senegalia senegal families from five randomly selected mother trees (seed origin) were used in this study. Seeds were harvested in January 2015 from each of the 5 different trees in a 20-year-old Senegalia senegal progeny trial in Dahra, Senegal. Ngane21B1, Kidira4B19, and Diamenar27B14 maintained a relatively high gum production over 4 successive years while Kidira17B19 and Diamenar27B16 displayed low gum yields for the same period (Table 1). Each tree label represents a mother tree, and families within the same name and different numbers belong to the same provenance.
2.2. Growth Conditions, Treatment, and Experimental Design
The experiment was conducted from June to October 2015 in a greenhouse where the temperature was maintained above 25 °C and ventilated with box fans when temperatures went above 32 °C. Light was extended using sodium vapor lights to maintain a 16-h photoperiod. Seeds were chemically scarified with concentrated sulfuric acid (98%) in order to perforate the tough seed coat and then rinsed with water to remove the acid before germinating them in shallow flats filled with Promix BX as a growth medium. Once emerged, young germinals were transplanted individually into small tubes (SC10 super cell, 3.8 cm diameter × 21 cm deep, Stuewe and Sons, Tangent, OR, USA) containing Promix and kept well-watered for about six weeks. They were then transplanted into 2.3-liter plastic pots (Treepots, 30.5 cm height and 10.2 cm width, IGC Greenhouse Megastore, Danville, IL, USA) containing either soil substrate (a 2:1 mix of Promix BX and washed sand) or soil substrate with salt added. A factorial combination of salinity and fertilizer treatments was then established.
The salinity treatment consisted of mixing 1070 g of sodium chloride with 100 kg of soil substrate. The salt treatment used was based on an early study which revealed a significant decrease of about 50% on biomass of Senegalia senegal seedlings [17]. The average corresponding electrical conductivity of the saline soil was 7.525 dSm−1, whereas the electrical conductivity of a control soil was 0.31 dSm−1. For the measurement of electrical conductivity, a soil suspension was prepared in distilled water at a ratio of 1:2. The suspension was shaken and allowed to stand overnight [17]. Thereafter, electrical conductivity of the supernatant solution was determined with an electrical conductivity meter (2265FS/2265FSTP, Spectrum Technologies, Inc., Aurora, IL, USA). The fertilization treatment consisted of applying 2.3 g of N-P-K (10-10-10) in each pot to give the equivalent fertilizer supply of 100 kg/ha. The experiment was designed as a randomized complete block with five replications of the 2 by 2 factorial treatments (no salt/no fertilizer (control), salt only, fertilizer only, and salt plus fertilizer) making a total of 100 experimental units.
When treatments started, all plants were kept well-watered for three weeks, and then the drought stress treatment was started where all watering was stopped in all treatments for three more weeks. This procedure was adopted to mimic field conditions (alternation between a wet season and a dry season) under which Senegalia senegal is naturally subjected. The induction of drought in this experiment is not considered as a treatment factor, but a common condition subjected to all experimental units. Prior to drought stress, shallow trays were placed beneath pots that received the salt treatment (the salted and salt-fertilized) in order to recover any salt discharge which occurred when pots were watered. Any recovered water was then poured back into each tree pot. This step was to maintain the same salt concentration in each pot prior drought stress.
2.3. Stomatal Density and Stomatal Size Measurements
Stomatal density and size were assessed at the end of the experiment before seedlings were harvested by using a stereo zoom microscope SMZ 1500 (Nikon Corporation, Chiyoda-ku, Tokyo, Japan) equipped with a digital camera (Nikon digital sight). A colorless nail polish was coated on the abaxial and adaxial leaflet surfaces from the newest fully mature leaf, then leaflets were mounted on glass slides. Images were taken on both leaf faces at 70× magnification.
Stomatal density was expressed as the number of stomata per mm2 and stomatal size reflected the length in micrometers between the junctions of the guard cells for each stoma. Two leaflets per face and per plant were sampled and 2 subsamples of 0.01 mm2 were measured on each leaflet face. The number of stomata for each sample was counted under a photomicroscope system with a computer attachment.
2.4. Leaf Gas Exchanges and Chlorophyll Content Measurements
Measurements of net photosynthetic rate (A), stomatal conductance (Gs), and transpiration (E) of a fully expanded leaf per plant were made using a portable infrared gas analyzer (LI-6400, Lincoln, NE, USA). Environmental conditions in the leaf cuvette were set to 2000 μmols m−2 s−1 photosynthetic photon flux density (PPFD), 398 μmols mol−1 reference CO2 concentration, 25 °C block temperature, and a flow rate of 400 μmols sec−1. Gas exchange was measured weekly from day 0 (when planted into large pots with fertilizer and salt) for the 6 weeks of the experiment. The intrinsic water use efficiency (WUEi) was calculated as the ratio of photosynthetic to transpiration rates. Total chlorophyll content (mg/g) was estimated at the same time in intact leaves using a portable chlorophyll meter CCM-300 (Opti-Sciences Inc., Hudson, NH, USA). Three leaflets per plant were randomly chosen from new fully mature leaves and an average taken for analysis.
2.5. Relative Growth Rate and Biomass
The relative growth rates of height (RGH, cm cm−1day−1) and diameter (RGD, mm mm−1day−1) were determined [18], where RGR = (ln X2 − ln X1)/(t2 − t1), t is the number of days, X is either plant height or diameter, ln X is the natural log of height or diameter.
All plants were harvested at the end of the experiment. We manually washed soil from roots and then separated leaf, stem, and root from each other. Root area from each plant was measured by using WinRHIZO Software (version 2005b Pro, Regent Instruments Inc., Québec, Canada) and an EPSON Expression 1680 scanner (Epson America Inc., Long Beach, CA, USA). The area of leaves for each plant was determined with Photoshop software CS6 (version 13.0.1, Adobe Systems Inc., Mountain View, CA, USA). Different plant parts were then dried at 65 °C to a constant weight before weighing them to determine dry matter.
2.6. Statistical Analysis
Data were analyzed using JMP software (version 12, SAS Institute, Cary, NC, USA). Normality and constant variance were examined across all data. We used a log (natural) transformation of the root/shoot ratio for analysis to meet assumptions of analysis of variance (ANOVA). Also, an outlier identified from specific leaf area dataset was omitted in the analysis. A full factorial model analysis was conducted to test treatments and family effects on all measured variables twice, once under well-watered conditions (prior drought) and then again under water stress conditions (drought). Student’s t-test and Tukey-Kramer Honestly Significance Difference were used for separation of means tests.
3. Results
3.1. Prior Drought Conditions
3.1.1. Leaf Gas Exchanges and Chlorophyll Content
Prior to drought stress, a family × fertilizer, and family × salt interaction was significant for A, Gs, E, and WUEi (Table 2). No interaction effect was found to be significant for total chlorophyll content, which was similar among families at the starting treatment ranging from 264.7 mg m−2 (K17B19) to 330.0 mg m−2 (in K4B19). However, main effects of salt (p < 0.0001) and fertilizer (p < 0.0001) (Table 2) were found on total chlorophyll content. Salt and fertilizer increased chlorophyll from 275.3 to 314.20 mg m−2 and from 281.2 to 309.1 mg m−2, respectively.
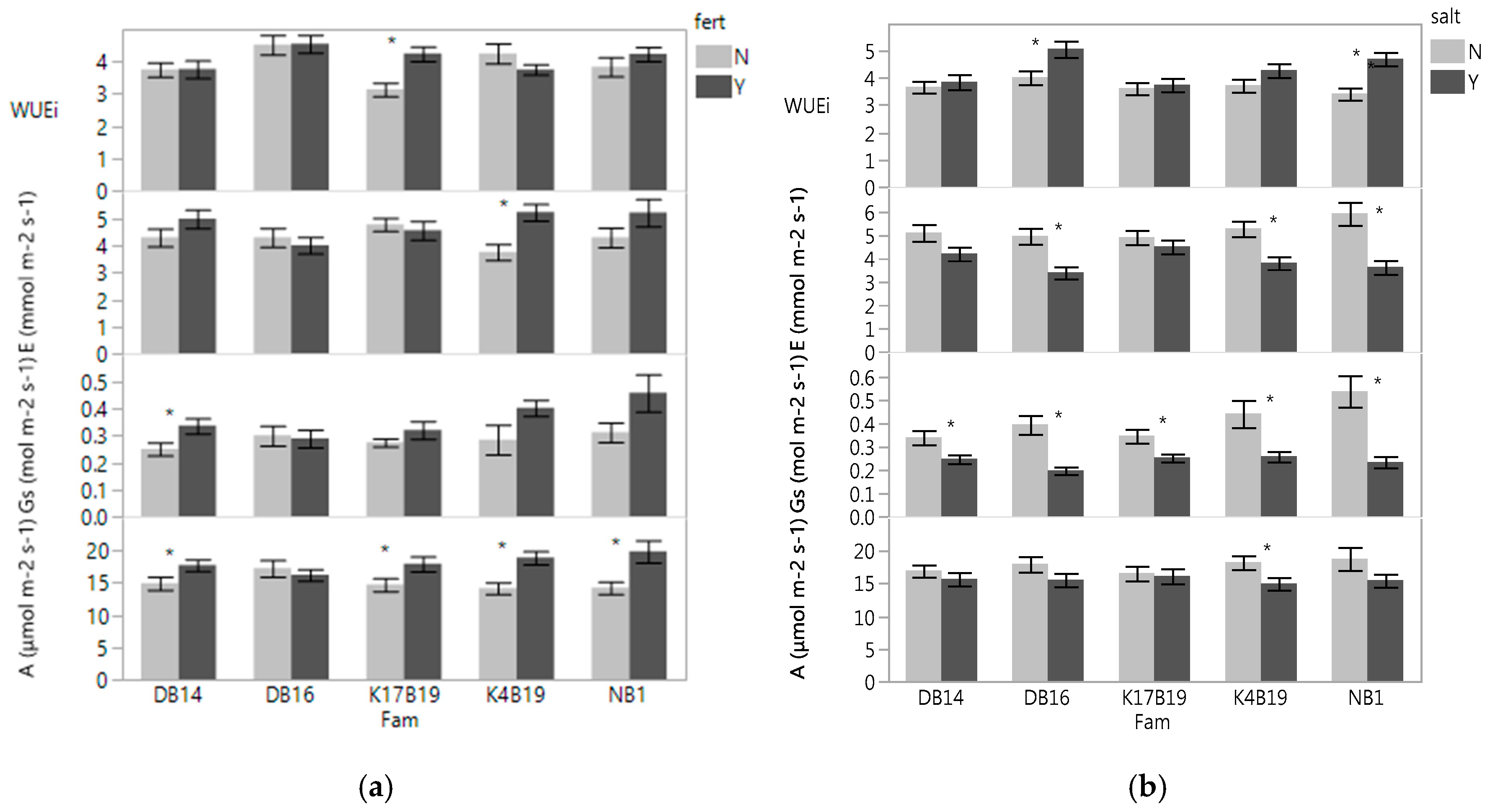
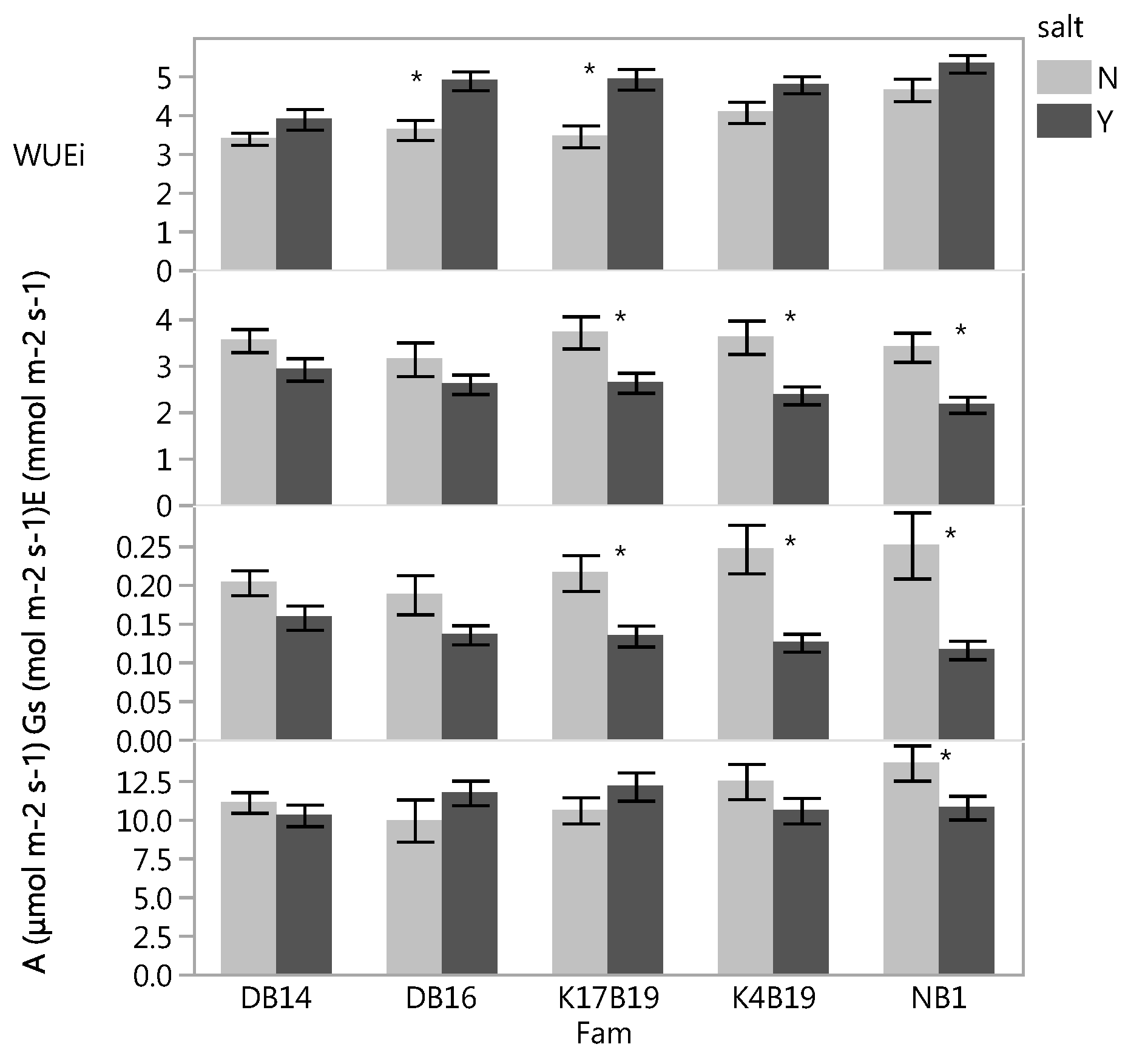
With the exception of DB16, fertilizer significantly increased A (Figure 1a,b). Gs and E were also generally increased with fertilizer, but were only significantly increased in a few families (Figure 1a). Since both A and E increased with fertilizer, the WUEi was generally unaffected by fertilizer. However, KB19 displayed higher WUEi. The salt treatment significantly decreased Gs in all families and E was lower in all families, but only significantly decreased in DB16, K4B19 and NB1. A was lower in the salt treatments in all families, but only significantly decreased in K4B19. WUEi was increased in all families, but only significantly in DB16 and NB1 (Figure 1b).
3.1.2. Relative Growth Rate
3.2. Under Drought Conditions
3.2.1. Stomatal Density and Size
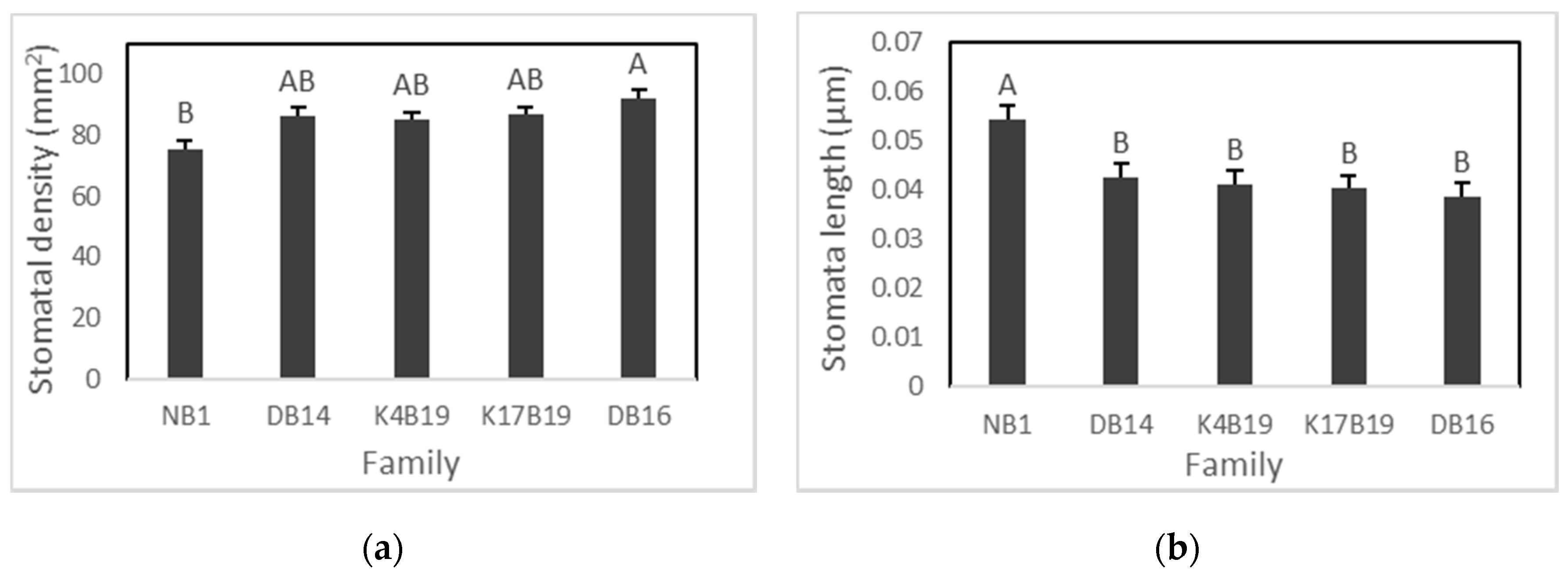
At the end of 3 weeks of drought, we found the presence of stomata on both abaxial and adaxial leaf surfaces and stomatal density and length did not differ significantly between leaf surfaces. Hence, stomatal density was considered to be the average of abaxial and adaxial surfaces. No family by treatment interactions were found to be significant in stomatal density and stomatal size (Table 3). However, salinity significantly increased stomatal density from 82 to 89 stomata per mm2, and a significant effect of family was found on both stomatal length and distribution (Figure 4 and Figure 5a,b).
The Diamenar provenance (DB16) had the highest stomatal density but was only significantly greater than Ngane NB1 (Figure 5a). Ngane displayed significantly larger stomata than all other provenances and significantly lower stomatal density than the Diamenar provenance (DB14 and DB16) (Figure 4 and Figure 5).
3.2.2. Leaf Gas Exchanges
There was a significant salt × family × fertilizer and salt × family interactions for WUEi and A, respectively (Table 3). The magnitude of the effect varied with family (Figure 6). Gas exchange rates were lower after the drought stress and the salt treatment generally still decreased Gs and E, and increased WUEi.
3.2.3. Biomass and Relative Growth Rates
There was a significant family × salt × fertilizer interaction for all biomass parameters (Table 4). K17b19 was the only family which increased biomass in response to the salt treatment. Total weight, leaf area, and root weight in this source were increased by 51%, 17%, and 47%, respectively. Ng21B1 growth was significantly decreased by the salt (Table 4). No other family had growth changed by the salt treatment. Growth was generally increased (more than 100%) in all families in response to added fertilizer (Table 4). Salt generally had a negative impact on this fertilizer growth enhancement.
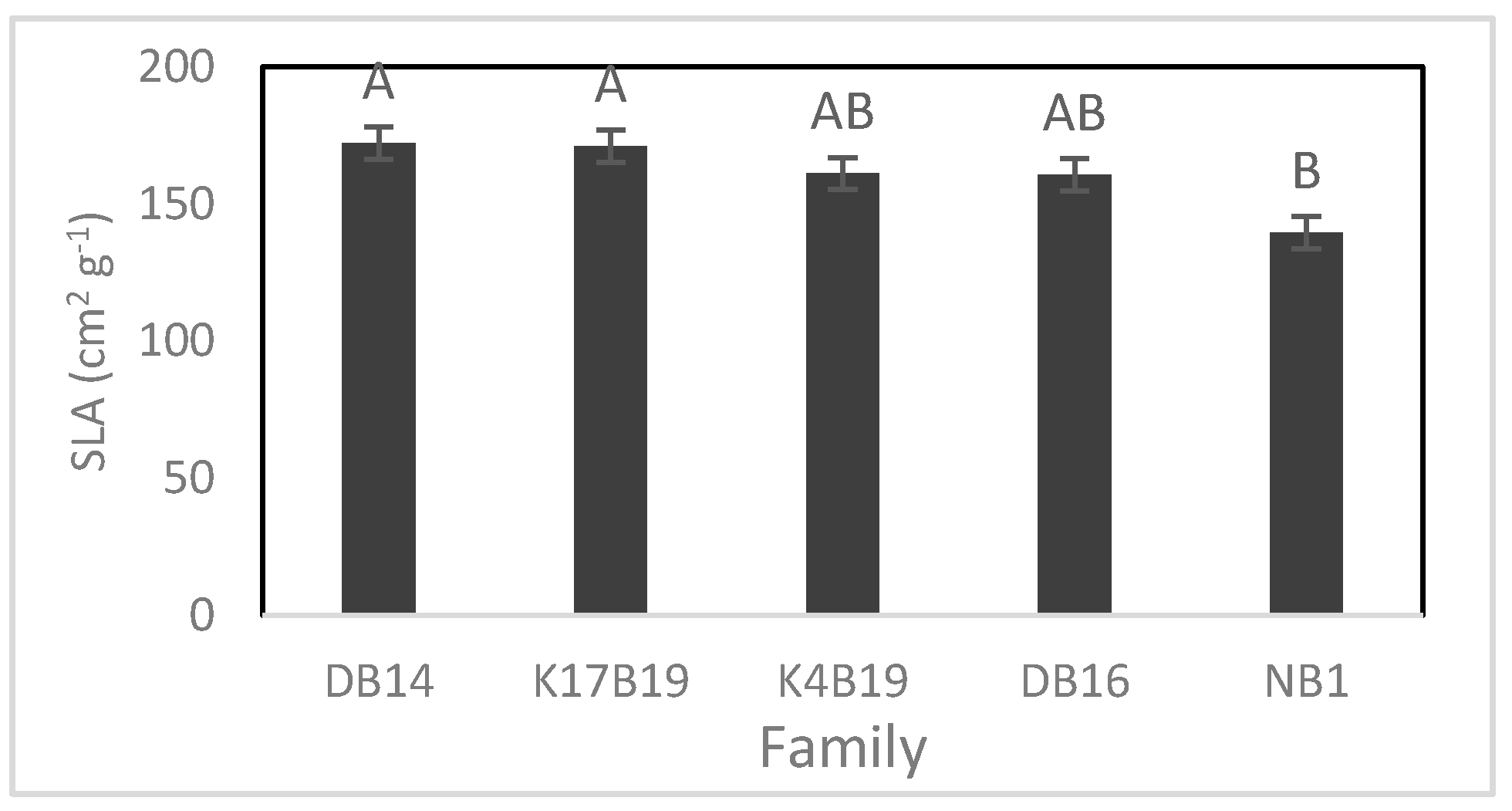
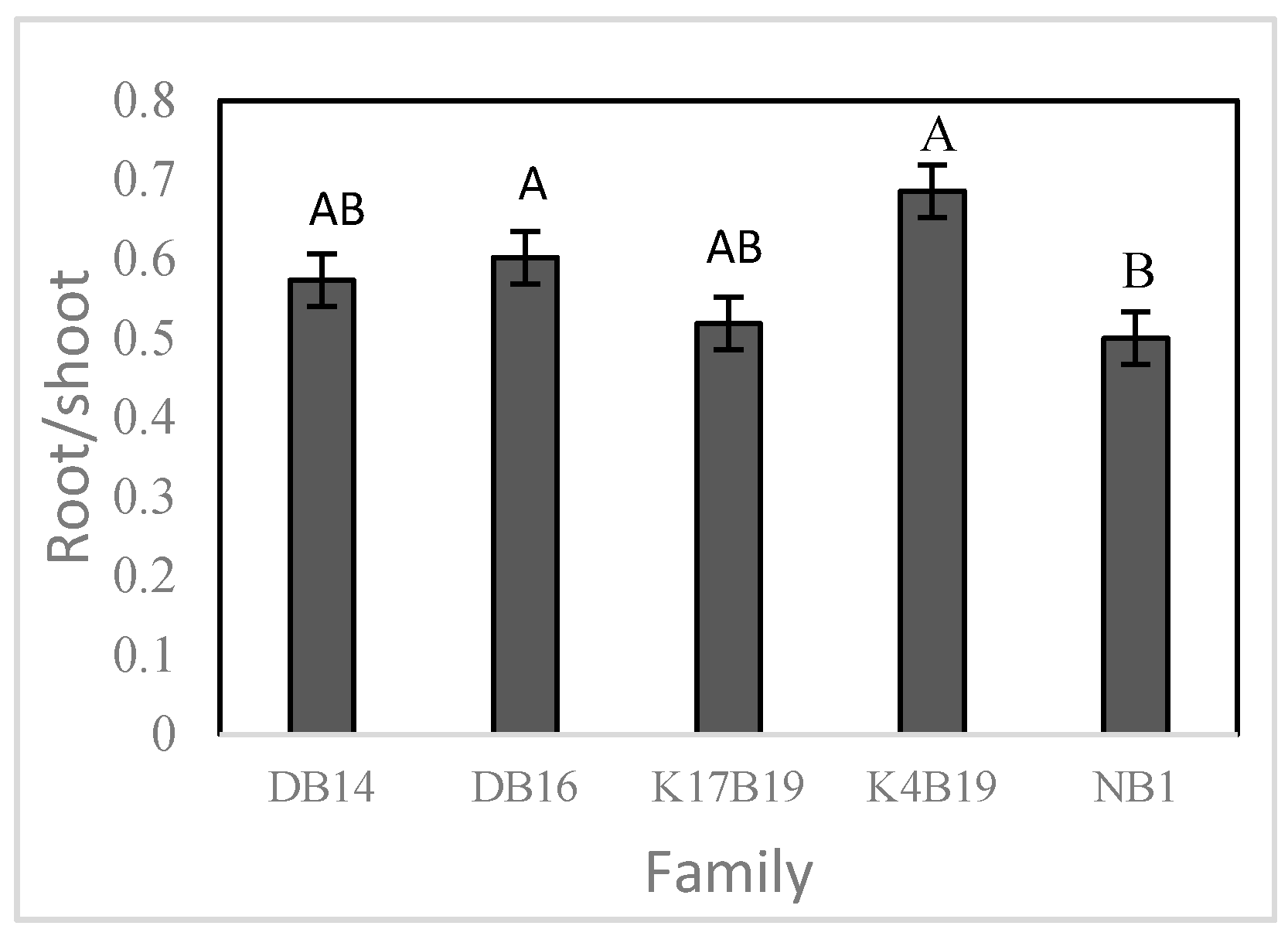
The main effects of family and salt treatments had a significant influence on specific leaf area (SLA) and root/shoot ratio. NB1 had the lowest SLA and root/shoot ratio while the other families did not differ significantly (Figure 7 and Figure 8).
Both SLA and root/shoot ratio were significantly reduced by the salt, decreasing from 169.7 to 152.2 cm2 g−1 and 0.61 to 0.47 in controls versus salt treated. The root/shoot ratio also decreased from 0.66 to 0.43 in controls versus fertilized plants.
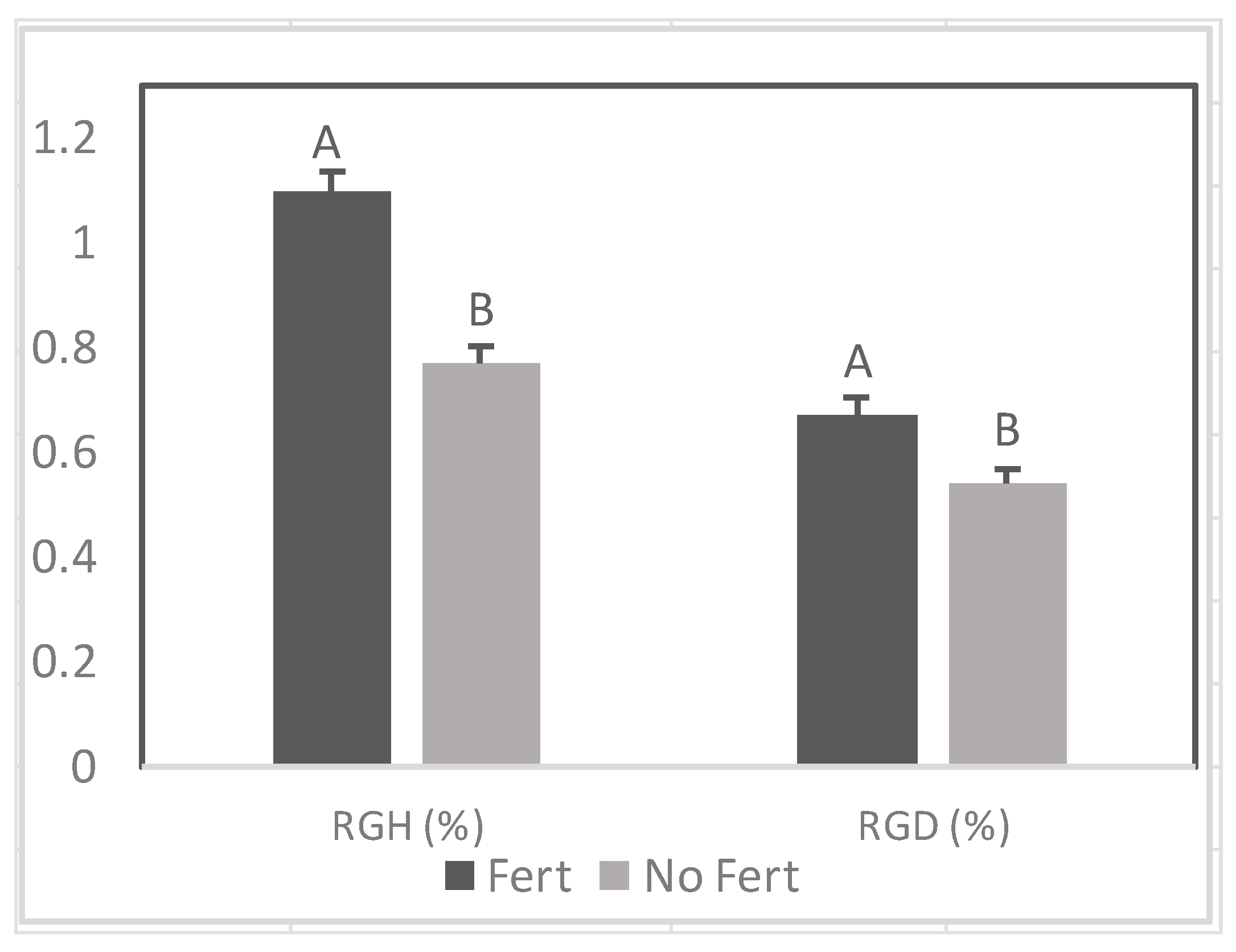
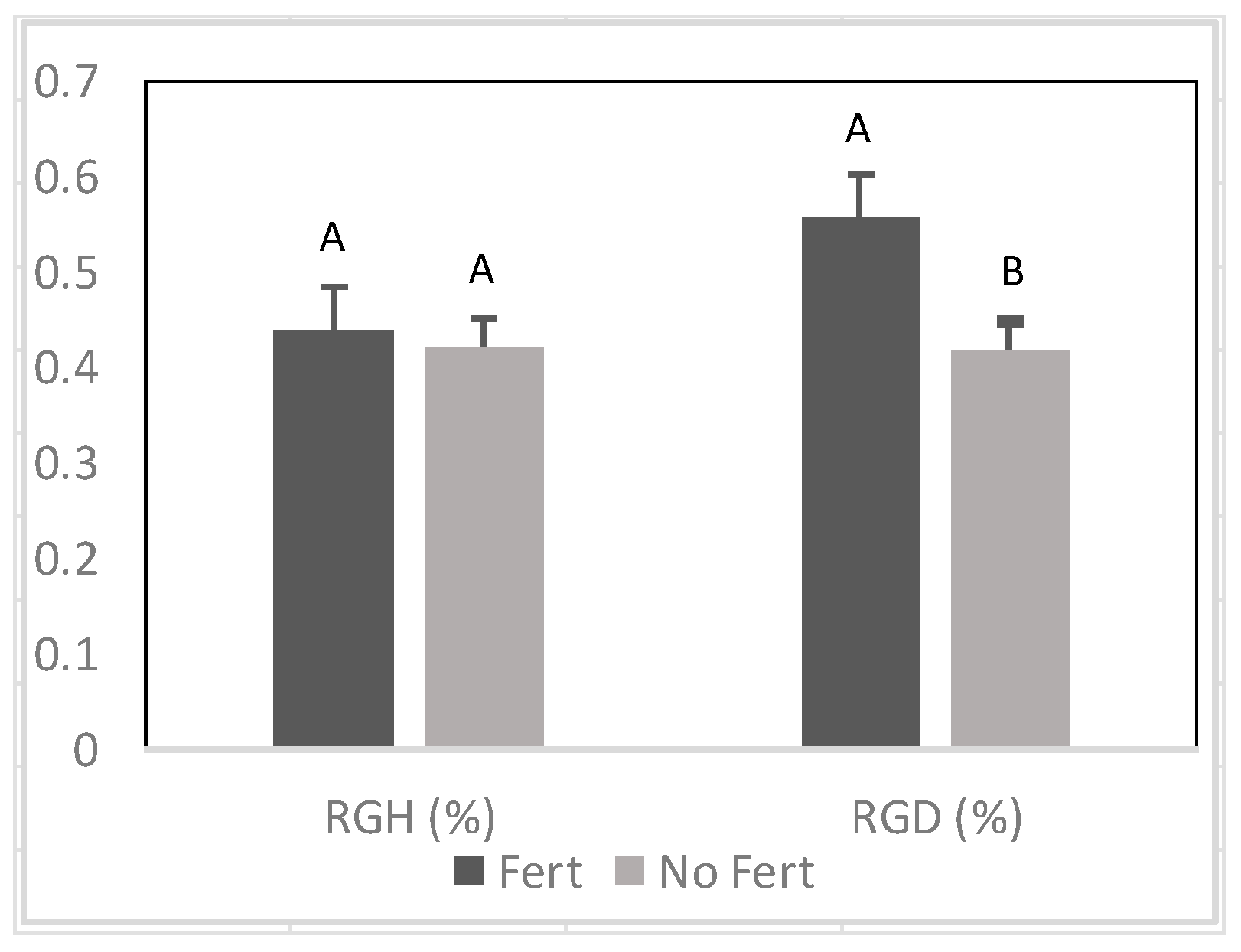
There was a positive effect of fertilizer application on RGD prior to and during the drought stress period (Figure 3 and Figure 9). In contrast, fertilizer did not increase RGH .during drought (Figure 9).
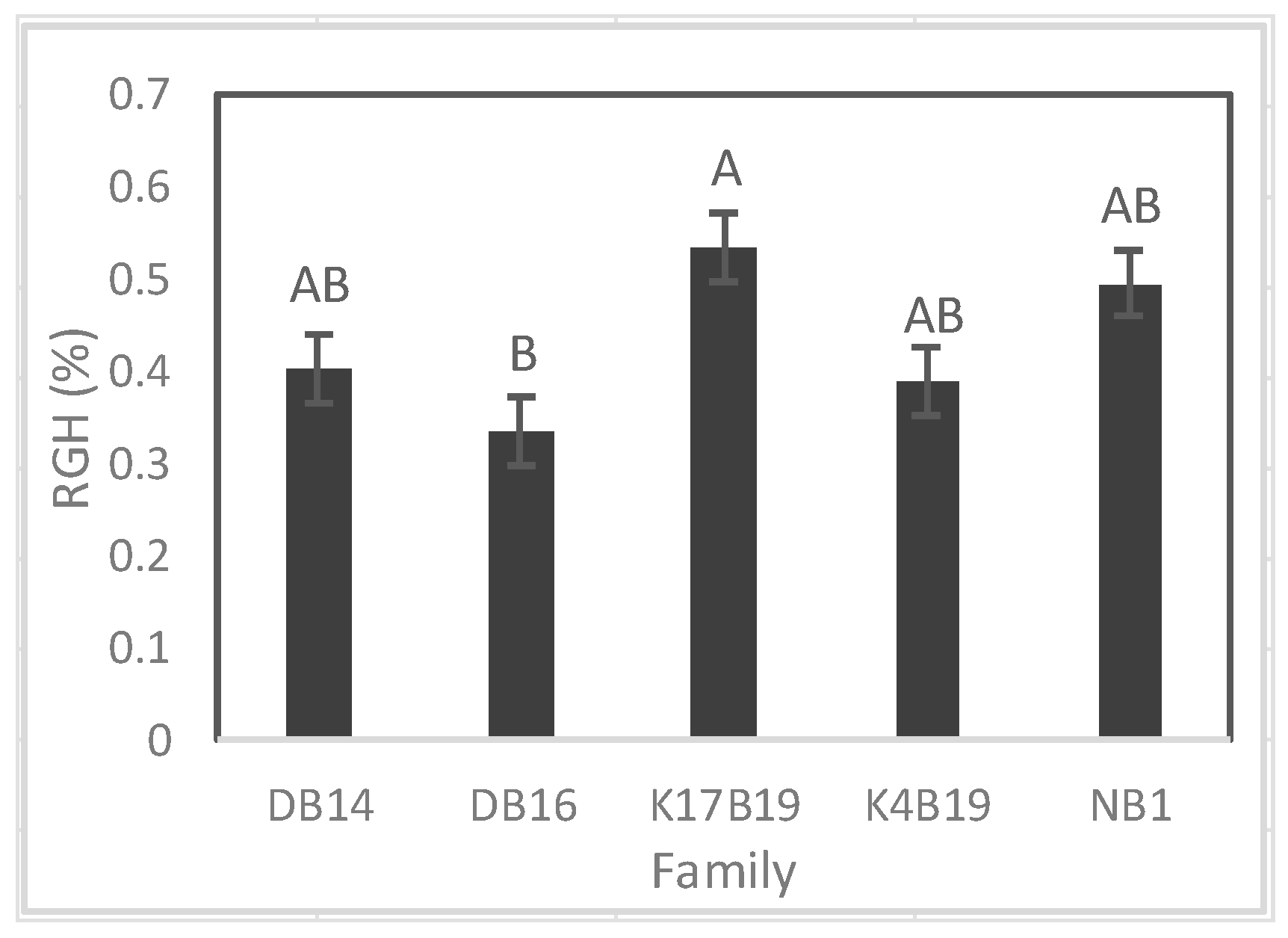
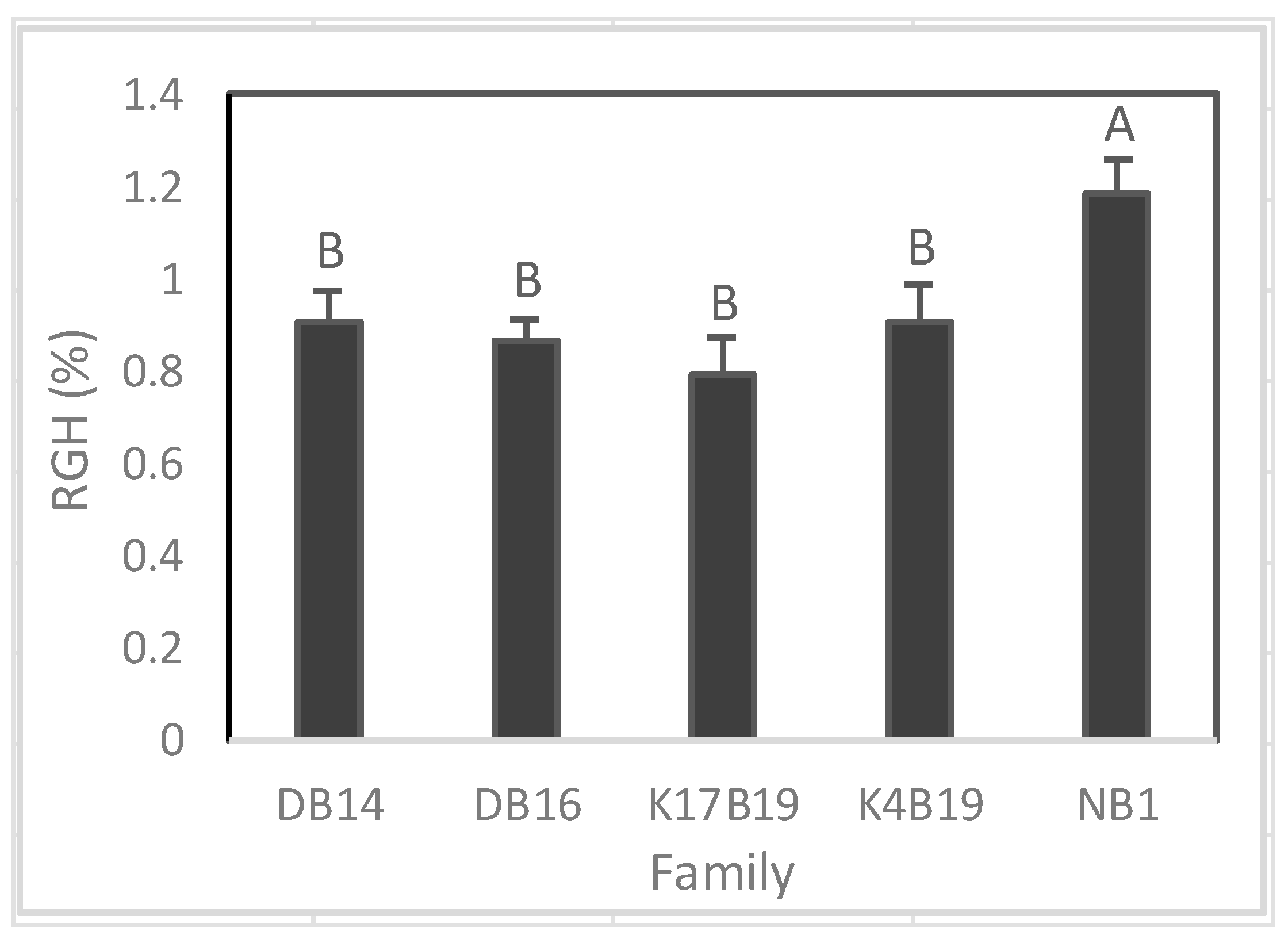
A significant difference among families was found on RGH (Figure 10), whereas RGD was similar between families. Prior to the drought, Ng21B1 had significantly greater RGH than all other families. During drought, RGH was greatly reduced and K17B19 displayed the highest RGH, but it was only significantly higher than DB16 (Figure 10).
4. Discussion
This study was the first attempt to look for growth performance and physiological adaptation under salt, and fertility conditions among different Senegalia senegal families collected in Senegal. Results of this study confirmed that photosynthetic capacities of Senegalia senegal seedlings are similar among families in non-limiting water conditions [19]. However, significant differences in stomatal density and size, SLA, root/shoot ratio, and RGH may be indicative of different responses to environmental stress among the tested families. The significant salinity by family interaction on dry matter accumulation found in this study implies that some Senegalia senegal families may perform better on salt affected drylands than on non-salty drylands [20]. NB1 had significantly reduced growth with salt addition, whereas K17B19 grew better in the salt treatment. An earlier study showed a significant decrease of Senegalia senegal shoot and root dry weights in response to the same salinity level used in our study [21]. However, they used a mixture of seeds collected from one coastal area and did not examine families, so it is possible they utilized a susceptible source such as our NB1.
The significant decrease of total biomass observed in NB1 reveals its higher sensitivity to salt stress than the other genotypes tested in this study. Despite originating from more saline areas, NB1 tended to be more sensitive to salinity, a finding similar to that which was reported in an earlier study which revealed that topodemes from areas subjected to salinity are more salt tolerant than those from non-salted areas [22]. In contrast, salt tolerance in Senegalia tortilis might be attributed to genetic factors that may be site independent [23]. Senegalia senegal is known as a salt tolerant species [5,6,7] and this is confirmed for most of our sources. Further, results on relative growth rate revealed a positive effect of salinity on RGH, which is significantly increased by 21% under drought (p = 0.0435). Our results suggest potential salt tolerance improvement of Senegalia senegal through genotype selection. Genetic variation for salt tolerance has been reported for many other species [23,24,25,26,27]. Surprisingly, K17B19 grew better in saline conditions, increasing total biomass by 51%, compared to non-saline conditions, even though the increase was not significant. Increased growth of plants under saline soils may be due to specific genes that control cell growth and leaf function [26]. K17B19, K4B19, DB14, and DB16 may have inherent ability to control water loss and improve ionic balance to their favor, therefore maintaining growth and biomass under salinity [28]. Recently, a sodium hydrogen antiporter gene (NHX1) has been identified in the Senegalia senegal tonoplast cells, which through an overexpression may confer capacity to tolerate salinity stress [5,29].
Unexpectedly, stomatal density and total chlorophyll content were significantly increased by salinity. Past studies found salinity to negatively affect stomatal density and chlorophyll content [14,23,30,31,32,33]. The presence of salt increased WUEi and decreased SLA (Figure 1b and Figure 7). Reduced SLA (thicker leaves) usually have more mesophyll cells with a higher density of chlorophyll and proteins per unit leaf area, hence a higher photosynthetic capacity than thinner leaves. Our results confirmed the negative association between WUEi and SLA reported by [34], who suggested lower SLA as a selection criterion for enhanced WUEi. Our study revealed that Senegalia senegal response to salt depended on genotypes and whether or not drought was induced. The increased WUEi in DB16 and NB1 in response to salt when water was not a limiting factor may be attributed to the significant decrease of transpiration relative to the assimilation rate through stomata closure (Figure 1b). The same response was observed on K17B19 under drought conditions. The increased WUEi likely is due to high photosynthesis, or low transpiration, or both [35,36]. However, under drought stress, the higher WUEi in response to salt without a significant change in stomata conductance observed in DB16 might be influenced by other traits such as mesophyll conductance. Besides stomatal control, inherent differences in mesophyll CO2 fixation capacity may influence the rate of photosynthesis and accordingly WUEi [37,38]. It is reported that enhancing mesophyll conductance/stomata conductance ratio can improve WUEi [37,39]. Moreover, notable effects of salinity and drought stress on mesophyll conductance have also been reported in Vitis vinifera [40]. Stomatal conductance was decreased in saline conditions by 39% prior to drought and by 37% during drought, averaged across all families. Still, influence of stomatal closure on A inhibition and regulation of stomatal conductance are strongly determined by both mesophyll and stomatal resistance to CO2 diffusion [41].
As expected, the fertilizer significantly increased the total biomass and biomass partition of all families. A positive effect of nutrient supply on Senegalia senegal growth has been revealed in an earlier study [42]. The significant increase in total biomass was likely due, in part, to the significant increase in the net photosynthetic rate under fertilizer conditions in most families (Figure 1a). However, the increased biomass partitioning was higher in leaves and stems than in roots, inducing a lower root/shoot ratio (Figure 8). It is reported that, in young seedlings, nutrient addition favored shoot growth more than root growth [38,43,44]. Our results confirm that fertilizer supply influences not only plant dry matter, but also biomass partitioning.
Trends in growth observed in seedling plants, showing lower RGH for DB16, were similar to what was found in their parent trees grown in the field which displayed lower height for DB16 (396 cm) compared to other genotypes. In other families, height varied from 510 cm for K4B19 to 590 cm for K17B19 [19]. We assume that height and RGH are highly heritable traits among Senegalia senegal genotypes. According to Dvorak et al. [45], heritability of growth traits for some forest trees does not change significantly between early and mature growth stages. With the exception of NB1, all sources (and in particular K17B19) used in this study would be suitable for testing on salt impacted soils. However, it would be important to test these findings in field conditions with more salt and fertilizer levels. Also, gum yield for such potential adapted genotypes would be important to take into account in the future to assess expected profit from growing Senegalia senegal in marginal and harsh dry lands.
5. Conclusions
This study testing the physiological and morphological adaptation of Senegalia senegal seedling under salinity and fertility conditions provided valuable information for improving Senegalia senegal tolerance to harsh environments through genotype selection. Our results revealed significant differences among Senegalia senegal genotypes in their leaf morphology. Large stomata size with low density was observed in NB1, which tended to be more sensitive to salinity despite lower specific leaf area. Photosynthetic capacities and growth rate performances showed significant interactions between families and salinity or fertility treatments. Diamenar and Kidira provenances in general and particularly the K17B19 family might be suitable for afforestation of salt affected drylands. All genotypes have notably increased biomass under the fertilizer effect. Our hypothesis that at least one family would be better adapted to salinity or fertilizer effect could be supported. However, despite the importance of intraspecific variation in changing environmental conditions, more research is needed to test findings in field conditions and to determine salt and fertilizer thresholds for optimized growth of Senegalia senegal in arid lands.
Acknowledgments
This work is made possible, in part, by the generous support of the American people through the United States Agency for International Development (USAID) as part of Feed the Future, the U.S. Government’s global hunger and food security initiative, under the terms of Contract No. AID-685-A-00-10-00194-00 (USAID/Education and Research in Agriculture). The contents are the responsibility of the author(s) and do not necessarily reflect the views of USAID or the United States Government. We would like to thank Audrey Zink-Sharp, professor and associate department head of sustainable biomaterials, Virginia Polytechnic Institute and state university, USA for the material provided and assistance on stomata measurements. Our gratitude to Momar Wade, research assistant at Centre National de Recherche Forestière of Institut Sénégalais de Recherche Agricole (CNRF/ISRA) for his help on harvesting Senegalia senegal seeds. We thank John Petersen, a laboratory specialist of the department Forest Resources and Environmental Conservation (FREC), Virginia Tech for his support along the greenhouse experiment.
Author Contributions
J.R.S. and M.S.S. conceived and designed the experiments; M.S.S. performed the experiment; J.R.S., J.S. and M.S.S. analyzed data and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gonzalez, P. Desertification and a Shift of Forest Species in the West African Sahel. Clim. Res. 2001, 17, 217–228. [Google Scholar] [CrossRef]
- Green Economy Sector Study on Water Resources in Senegal. Available online: https://www.unep.org/greeneconomy/sites/unep.org.greeneconomy/files/publications/ge_water_senegal_final.pdf (accessed on 10 October 2017).
- Bradley, D.; Grainger, A. Social resilience as a controlling influence on desertification in Senegal. Land Degrad. Dev. 2004, 15, 451–470. [Google Scholar] [CrossRef]
- Rapport Exploratoire sur L’Économie Verte au Sénégal. Available online: http://www.cepod.gouv.sn/sites/default/files/L'%C3%A9conomie%20verte%20%20rapport%20provisoire.pdf (accessed on 10 October 2017).
- Fall, D.; Bakhoum, N.; Fall, F.; Diouf, F.; Ly, M.O.; Diouf, M.; Gully, D.; Hocher, V.; Diouf, D. Germination, Growth and Physiological Responses of Senegalia senegal (L.) Britton, Vachellia seyal (Delile) P. Hurter and Prosopis Juliflora (Swartz) DC to Salinity Stress in Greenhouse Conditions. Afr. J. Biotechnol. 2016, 15, 2017–2027. [Google Scholar]
- Pandey, S.; Zhang, W.; Assmann, S.M. Roles of ion channels and transporters in guard cell signal transduction. FEBS Lett. 2007, 581, 2325–2336. [Google Scholar] [CrossRef] [PubMed]
- Sambou, A.; Ndour, B.; Cheng, S.; Senghor, E. Ligneous species tolerance in acid sulphated and saline soils of sine saloum: Case of rural community of Djilass and Loul Secene. J. Sustain. Dev. 2010, 3, 174. [Google Scholar] [CrossRef]
- Sonneveld, B.G.J.S.; Keyzer, M.A.; Zikhali, P.; Merbis, M. National Land Degradation Assessment Senegal and Review of Global Socio-Economic Parameters in the LADA Data Base–Land Degradation Assessment (LADA) Project Paper; Centre for World Food Studies: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Aroca, R. Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Masle, J.; Gilmore, A.; Farquhar, G. The ERECTA Gene Regulates Plant Transpiration Efficiency in Arabidopsis. Nature 2005, 436, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.Q.; Liu, Y.F.; Liu, P.; Lei, G.; He, S.; Ma, B.; Zhang, W.; Zhang, J.; Chen, S. Receptor-like kinase OsSIK1 improves drought and salt stress tolerance in rice (Oryza sativa) plants. Plant J. 2010, 62, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Reginato, M.A.; Reinoso, H.; Llanes, A.S.; Luna, M.V. Stomatal abundance and distribution in Prosopis strombulifera plants growing under different iso-osmotic salt treatments. Am. J. Plant Sci. 2013, 4, 41195. [Google Scholar] [CrossRef]
- Chaves, M.M.; Costa, J.M.; Saibo, N.J.M. Recent Advances in Photosynthesis under Drought and Salinity. Elsevier Sci. Technol. 2011, 57, 49–104. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Hardikar, S.A.; Pandey, A.N. Growth, Water Status and Nutrient Accumulation of Seedlings of Acacia senegal (L.) Willd. in Response to Soil Salinity. An. Biol. 2008, 30, 17–28. [Google Scholar]
- Blackman, V.H. The compound interest law and plant growth. Ann. Bot. 1919, 33, 353–360. [Google Scholar] [CrossRef]
- Sarr, M.S. Morphological and Physiological Responses of Senegalia senegal (L.) Britton Provenances to Drought, Salinity, and Fertility. Ph.D. Thesis, Virginia Tech, Blacksburg, VA, USA, 2017; p. 121. [Google Scholar]
- Flowers, T.J.; Yeo, A.R. Ion Relations of Plants Under Drought and Salinity. Aust. J. Plant Physiol. 1986, 13. [Google Scholar] [CrossRef]
- Briggs, D. Genecological Studies of Salt Tolerance in Groundsel (Senecio vulgaris L.) with Particular Reference to Roadside Habitats. New Phytol. 1978, 81, 381–389. [Google Scholar] [CrossRef]
- Muturi, G.M. Provenance Variation of Salt Tolerance and Seedling Nutrition in Acacia tortilis (Forsk.) Hayne. Ph.D. Thesis, University of Nairobi, Nairobi, Kenya, 1993. [Google Scholar]
- Cha-um, S.; Somsueb, S.; Samphumphuang, T.; Kirdmanee, C. Salt tolerant screening in eucalypt genotypes (Eucalyptus spp.) using photosynthetic abilities, proline accumulation, and growth characteristics as effective indices. In Vitro Cell. Dev. Biol. Plant 2013, 49, 611–619. [Google Scholar] [CrossRef]
- Fung, L.E.; Wang, S.S.; Altman, A.; Hutterman, A. Effect of NaCl on Growth, Photosynthesis, Ion and Water Relations of Four Poplar Genotypes. For. Ecol. Manag. 1998, 107, 135–146. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of Salt Tolerance in Nonhalophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Munns, R. Genes and Salt Tolerance: Bringing them Together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Ghoulam, C.A.F.; Fares, K. Effects of Salt Stress on Growth, Inorganic Ions and Proline Accumulation in Relation to Osmotic Adjustment in Five Sugar Beet Cultivars. Environ. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]
- Yadav, R.S.; Sehgal, D.; Vadez, V. Using genetic mapping and genomics approaches in understanding and improving drought tolerance in pearl millet. J. Exp. Bot. 2011, 62, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates, Incorporated: Sunderland, MA, USA, 2015. [Google Scholar]
- Barbieri, G.; Vallone, S.; Orsini, F.; Paradiso, R.; De Pascale, S.; Negre-Zakharov, F.; Maggio, A. Stomatal Density and Metabolic Determinants Mediate Salt Stress Adaptation and Water Use Efficiency in Basil (Ocimum basilicum L.). J. Plant Physiol. 2012, 169, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1995. [Google Scholar]
- Yang, H.; Wang, G. Leaf stomatal densities and distribution in Triticum aestivum under drought and CO2 enrichment. Acta Ecol. Sinica 2000, 25, 312–316. [Google Scholar]
- Zhang, H.; Li, J.; Yoo, J.H.; Yoo, C.C.; Cho, S.H.; Koh, H.J.; Seo, H.S.; Paek, N.C. Rice Chlorina-1 and Chlorina-9 Encode ChlD and ChlI Subunits of mg Chelatase, a Key Enzyme for Chlorophyll Synthesis and Chloroplast Development. Plant Mol. Biol. 2006, 62, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.N.; Udaykumar, M.; Farquhar, G.D.; Talwar, H.S.; Prasad, T.G. Variation in carbon isotope discrimination and its relationship to specific leaf area and ribulose-1, 5-bisphosphate carboxylase content in groundnut genotypes. Funct. Plant Biol. 1995, 22, 545–551. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Hall, A.E.; Farquhar, G.D. Stable Isotopes and Plant Carbon/Water Relations; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Li, C. Carbon isotope composition, water-use efficiency and biomass productivity of Eucalyptus microtheca populations under different water supplies. Plant Soil 1999, 214, 165–171. [Google Scholar] [CrossRef]
- Flexas, J.; Niinemets, U.; Galle, A.; Barbour, M.; Centritto, M.; Diaz-Espejo, A.; Douthe, C.; Galmes, J.; Ribas-Carbo, M.; Rodrigues, P.L.; et al. Diffusional Conductances to CO2 as a Target for Increasing Photosynthesis and Photosynthetic Water Use Efficiency. Photosynth. Res. 2013, 117, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Pallardy, S.G. Physiology of Woody Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2008. [Google Scholar]
- Flexas, J.; Diaz-Espejo, A.; Conesa, M.A.; Coopman, R.E.; Douthe, C.; Gago, J.; Galle, A.; Galmes, J.; Medrano, H.; Ribas-Carbo, M.; et al. Mesophyll Conductance to CO2 and Rubisco as Targets for Improving Intrinsic Water Use Efficiency in C3 Plants. Plant Cell Environ. 2016, 39, 965–982. [Google Scholar] [CrossRef] [PubMed]
- Ferrio, J.P.; Pou, A.; Florez-Sarasa, I.; Gessler, A.; Kodama, N.; Flexas, J.; Ribas-Carbo, M. The Péclet Effect on Leaf Water Enrichment Correlates with Leaf Hydraulic Conductance and Mesophyll Conductance for CO2. Plant Cell Environ. 2012, 35, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, E.T.; Orcutt, D.M.; Hale, M.G. The Physiology of Plants under Stress; Wiley: New York, NY, USA, 1996. [Google Scholar]
- Daldoum, D.M.A.; Hammad, G.H. Performance of Acacia senegal (L.) Wild Seedlings Growth under some Tree Manures and NPK Fertilizers in Nursery Site. J. Environ. Sci. 2015, 31, 303–311. [Google Scholar] [CrossRef]
- Fernández, M.; Novillo, C.; Pardos, J.A. Effects of Water and Nutrient Availability in Pinus pinaster Ait. Open Pollinated Families at an Early Age: Growth, Gas Exchange and Water Relations. New For. 2006, 31, 321–342. [Google Scholar] [CrossRef]
- Stovall, J.P.; Fox, T.R.; Seiler, J.R. Short Term Changes in Biomass Partitioning of Two Full Sib Clones of Pinus taeda L. Under Differing Fertilizer Regimes over 4 months. Trees 2012, 26, 951–961. [Google Scholar] [CrossRef]
- Dvorak, W.S.; Uruena, H.; Moreno, L.A.; Goforth, J. Provenance and family variation in Sterculia apetala in Colombia. For. Ecol. Manag. 1998, 111, 127–135. [Google Scholar] [CrossRef]
Figure 1.
The effect of fertilizer (a) and salt (b) treatments on net photosynthetic rate (A), stomatal conductance (Gs), transpiration (E), and intrinsic water use efficiency (WUEi) of Senegalia senegal families. Asterisk marks on bars mean significant effect of fertilizer (a) and salt (b) at p < 0.05.
Figure 1.
The effect of fertilizer (a) and salt (b) treatments on net photosynthetic rate (A), stomatal conductance (Gs), transpiration (E), and intrinsic water use efficiency (WUEi) of Senegalia senegal families. Asterisk marks on bars mean significant effect of fertilizer (a) and salt (b) at p < 0.05.
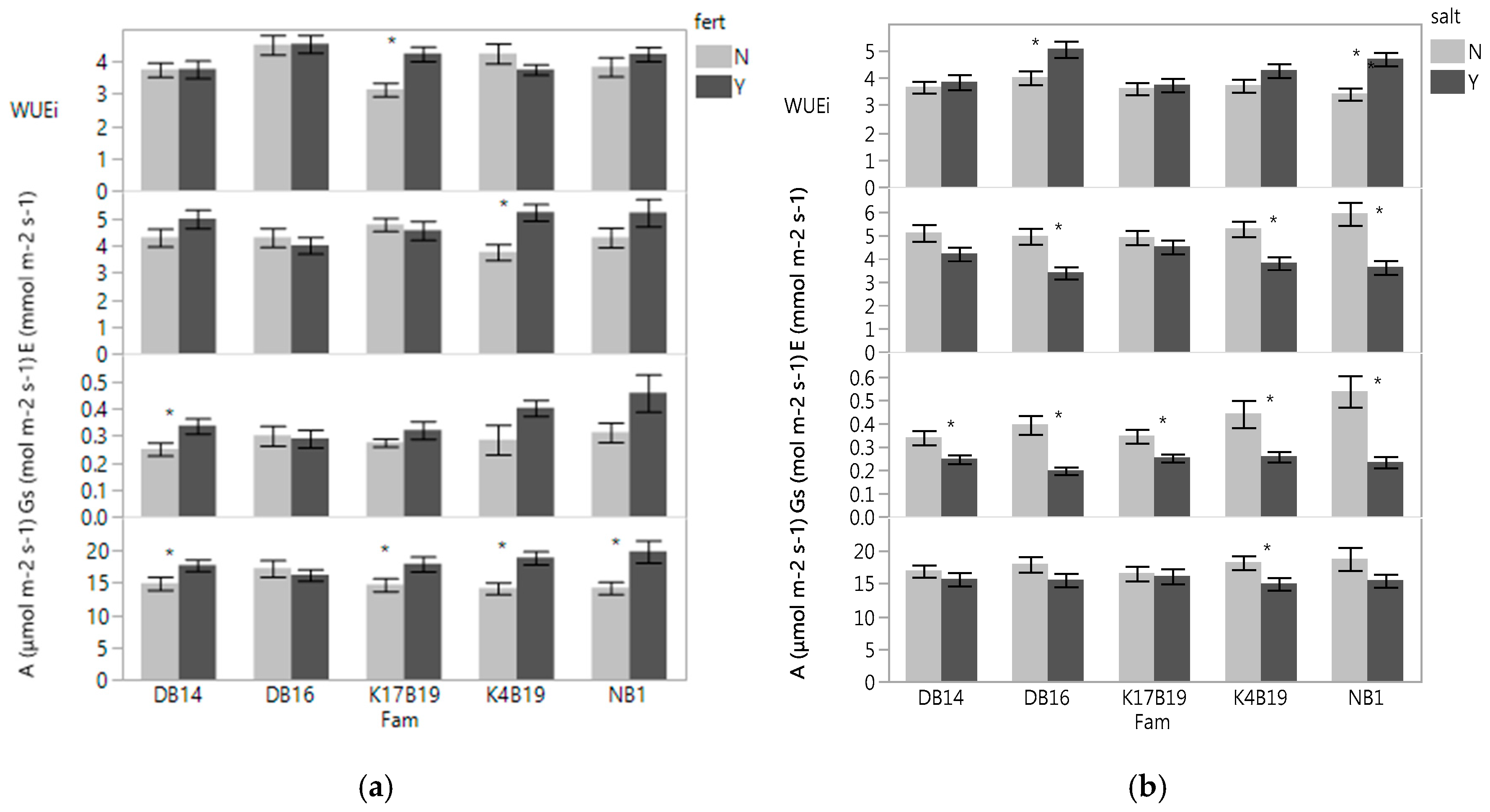
Figure 2.
Variation of relative growth rate of height (RGH) among Senegalia senegal families. Different letters on bars mean significant differences among families at p < 0.05.
Figure 2.
Variation of relative growth rate of height (RGH) among Senegalia senegal families. Different letters on bars mean significant differences among families at p < 0.05.
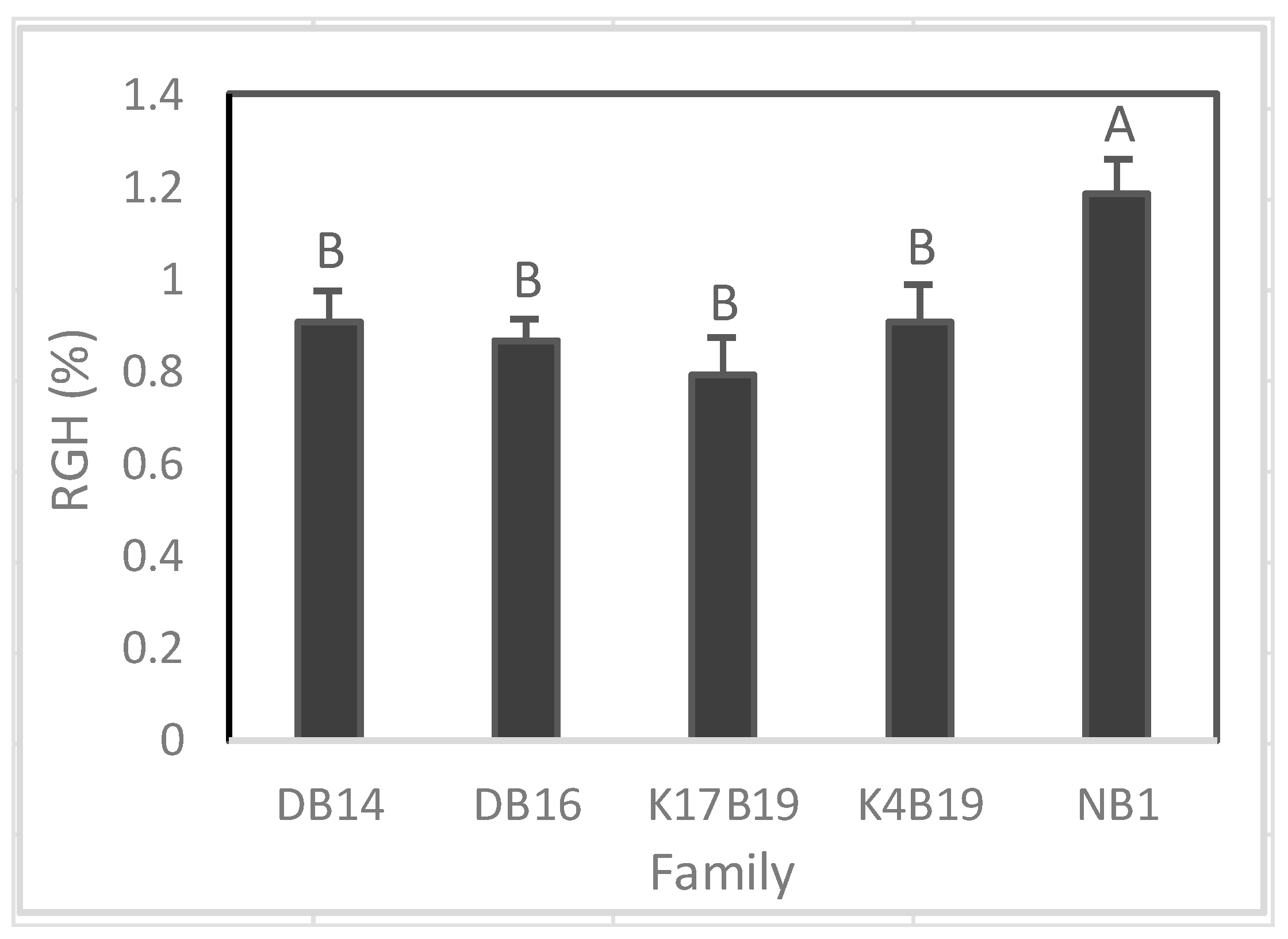
Figure 3.
Effect of fertilizer (Fert) on relative growth rate of height (RGH) and diameter (RGD) among Senegalia senegal seedlings. Different letters on bars mean significant differences between fertilized (Fert) and no fertilized (No Fert) at p < 0.05.
Figure 3.
Effect of fertilizer (Fert) on relative growth rate of height (RGH) and diameter (RGD) among Senegalia senegal seedlings. Different letters on bars mean significant differences between fertilized (Fert) and no fertilized (No Fert) at p < 0.05.
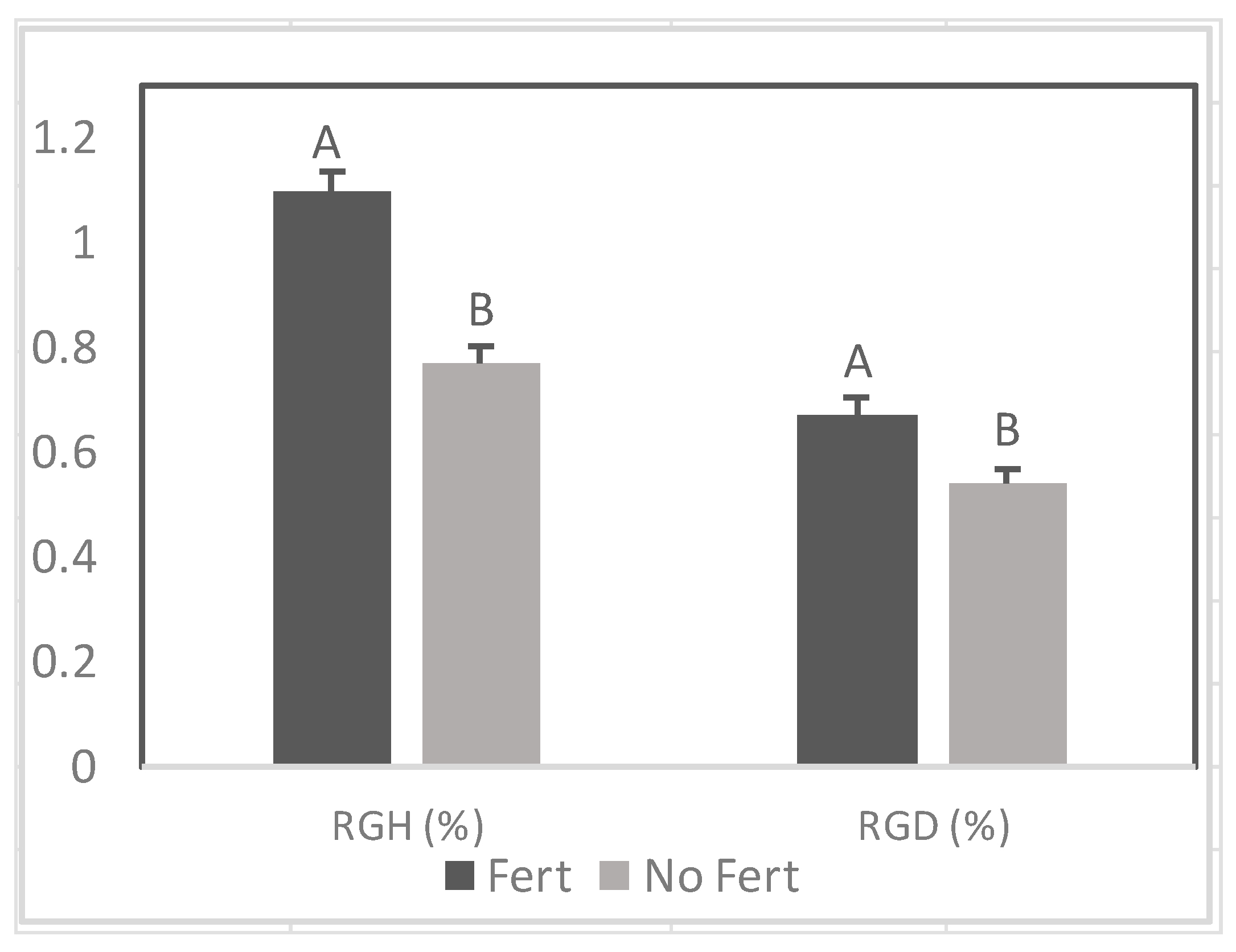
Figure 4.
Stomata on the abaxial and adaxial leaf surfaces of 12-week-old Senegalia senegal seedlings. NB1 (a), K4B19 (b), K17B19 (c), DB14 (d), DB16 (e).
Figure 4.
Stomata on the abaxial and adaxial leaf surfaces of 12-week-old Senegalia senegal seedlings. NB1 (a), K4B19 (b), K17B19 (c), DB14 (d), DB16 (e).

Figure 5.
Variation of stomatal density (a) and size (b) among Senegalia senegal families. Different letters on bars mean significant differences among families at p < 0.05.
Figure 5.
Variation of stomatal density (a) and size (b) among Senegalia senegal families. Different letters on bars mean significant differences among families at p < 0.05.
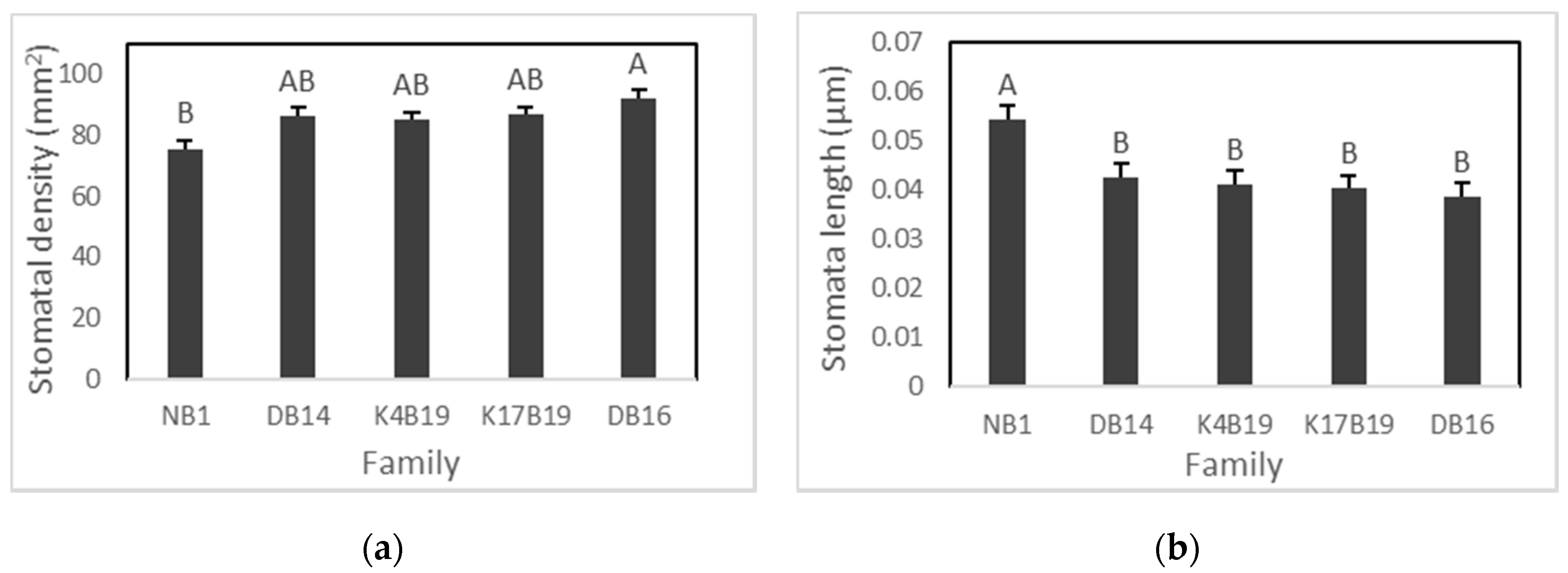
Figure 6.
Effect of salt on net photosynthetic rate (A), stomatal conductance (Gs), transpiration (E), and intrinsic water use efficiency (WUEi) of Senegalia senegal families under drought stress. Asterisk marks on bars mean significant effect of salt at p < 0.05.
Figure 6.
Effect of salt on net photosynthetic rate (A), stomatal conductance (Gs), transpiration (E), and intrinsic water use efficiency (WUEi) of Senegalia senegal families under drought stress. Asterisk marks on bars mean significant effect of salt at p < 0.05.
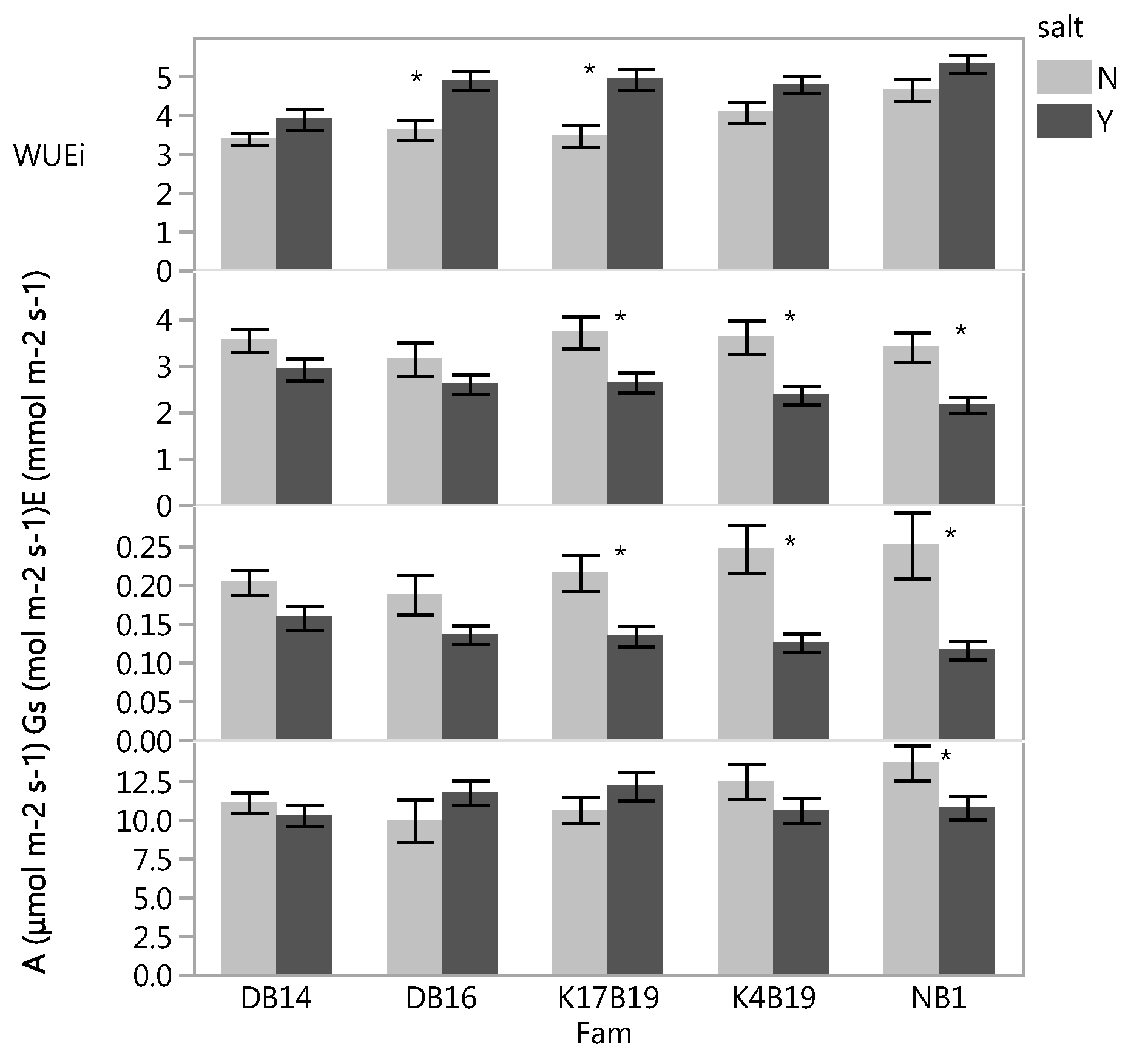
Figure 7.
Variation of specific leaf area (SLA) of Senegalia senegal families. Different letters on bars mean significant differences among families at p < 0.05.
Figure 7.
Variation of specific leaf area (SLA) of Senegalia senegal families. Different letters on bars mean significant differences among families at p < 0.05.
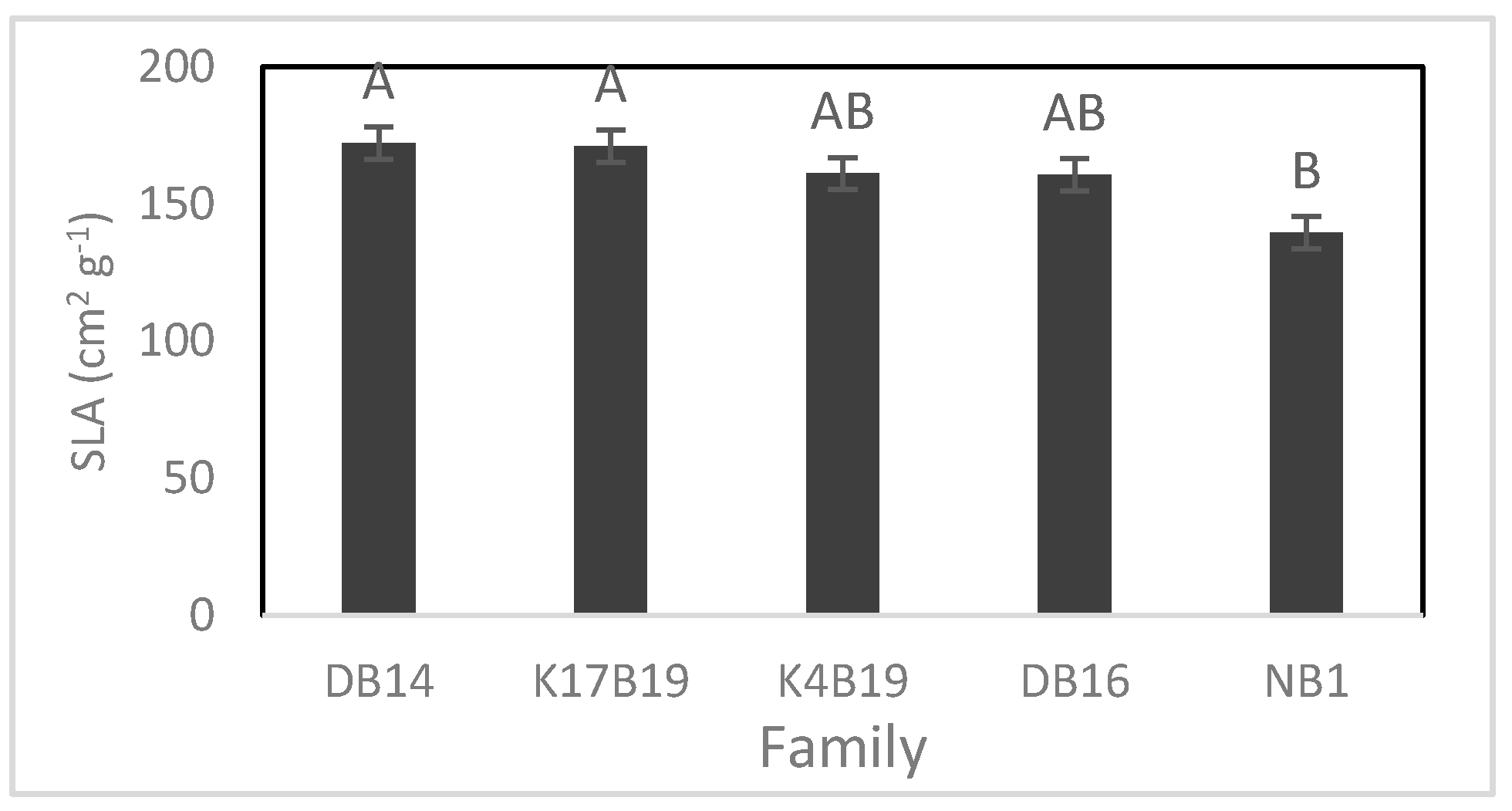
Figure 8.
Variation of root/shoot ratio among families of Senegalia senegal seedlings. Different letters on bars mean significant differences among families at p < 0.05.
Figure 8.
Variation of root/shoot ratio among families of Senegalia senegal seedlings. Different letters on bars mean significant differences among families at p < 0.05.
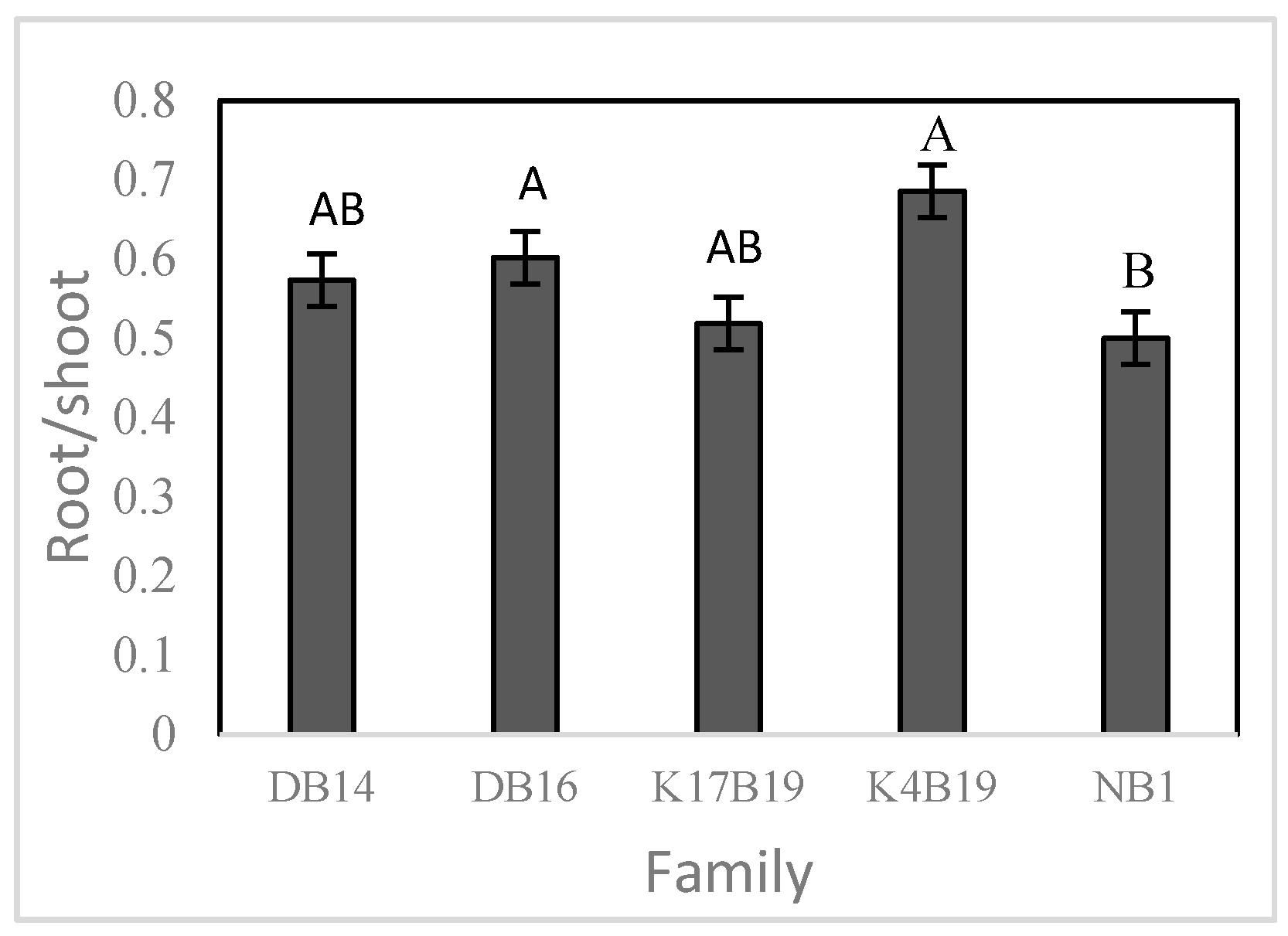
Figure 9.
The effect of fertilizer (Fert: fertilized, No fert: no fertilized) on the relative growth rate of diameter (RGD) and relative growth rate of height (RGH) of Senegalia senegal seedlings during drought stress. Different letters on bars mean significant differences among families at p < 0.05.
Figure 9.
The effect of fertilizer (Fert: fertilized, No fert: no fertilized) on the relative growth rate of diameter (RGD) and relative growth rate of height (RGH) of Senegalia senegal seedlings during drought stress. Different letters on bars mean significant differences among families at p < 0.05.
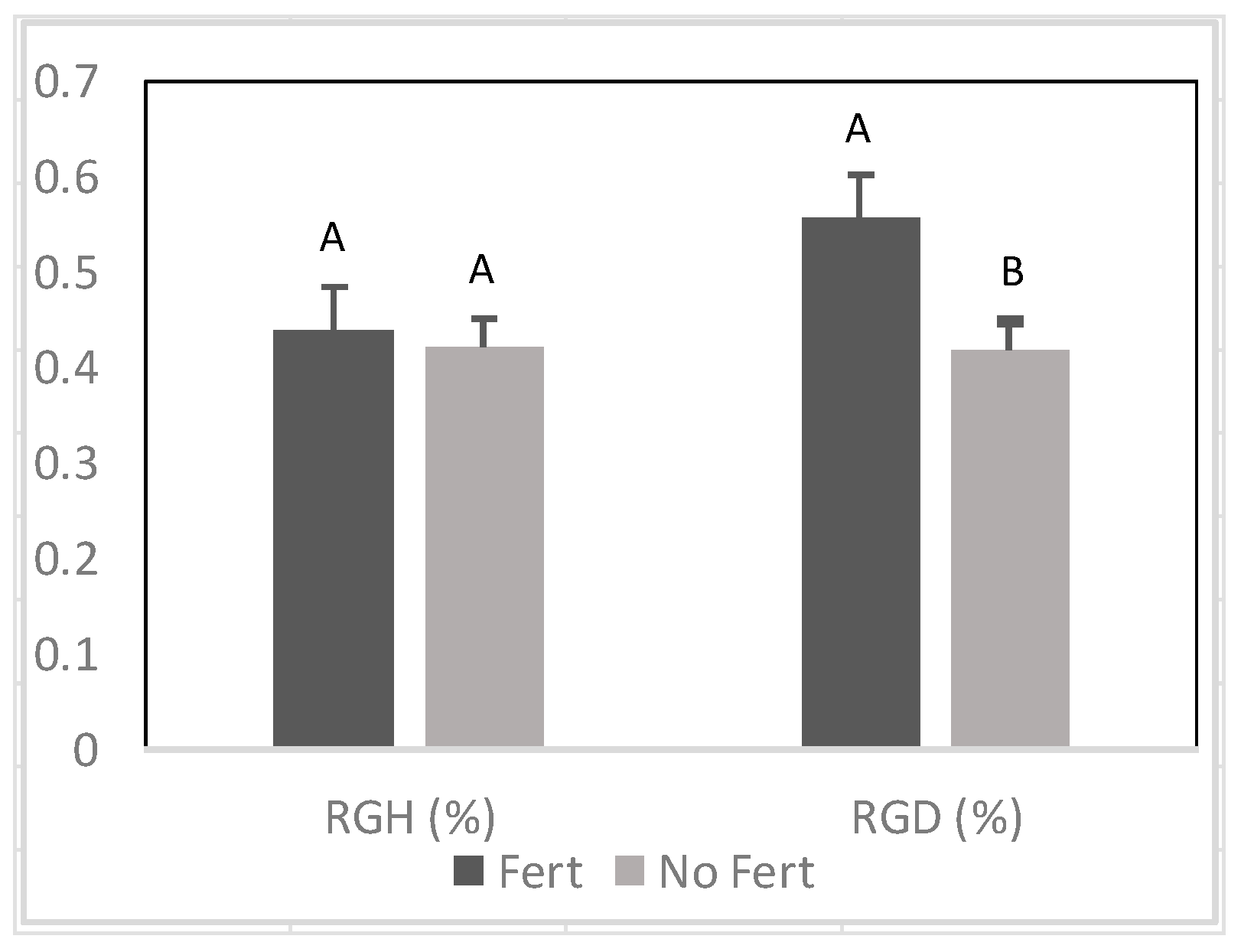
Figure 10.
Variation of relative growth rate of height (RGH) between Senegalia senegal families) and during drought stress. Different letters on bars mean significant differences among families at p < 0.05.
Figure 10.
Variation of relative growth rate of height (RGH) between Senegalia senegal families) and during drought stress. Different letters on bars mean significant differences among families at p < 0.05.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Families and mean annual gum yields (g) by tree of the mother trees from which seeds in this study were harvested. (-) indicates that the tree died.
Table 1.
Families and mean annual gum yields (g) by tree of the mother trees from which seeds in this study were harvested. (-) indicates that the tree died.
| Provenance | Families | Tree Label | 2013 Gum Yield | 2014 Gum Yield | 2015 Gum Yield | 2016 Gum Yield |
|---|---|---|---|---|---|---|
| Diamenar | Diamenar27B16 | DB16 | 29.3 | 9.7 | 0 | - |
| Diamenar | Diamenar27B14 | DB14 | 899.5 | 791.7 | 386.1 | 758.3 |
| Kidira | Kidira4B19 | K4B19 | 555.3 | 551.8 | 583.3 | 990.3 |
| Kidira | Kidira17B19 | K17B19 | 131.3 | 62.1 | 22.2 | 51.7 |
| Ngane | Ngane21B1 | NB1 | 522.2 | 476.4 | 945.2 | 1103.5 |
Table 2.
Analysis of variance (ANOVA) results for the response of different Senegalia senegal families to salinity and fertility for leaf gas exchanges (net photosynthetic rate (A), stomatal conductance (Gs), transpiration (E), and intrinsic water use efficiency (WUEi)), relative growth of height (RGH) and diameter (RGD), and chlorophyll content.
Table 2.
Analysis of variance (ANOVA) results for the response of different Senegalia senegal families to salinity and fertility for leaf gas exchanges (net photosynthetic rate (A), stomatal conductance (Gs), transpiration (E), and intrinsic water use efficiency (WUEi)), relative growth of height (RGH) and diameter (RGD), and chlorophyll content.
| Sources of Variation | df | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| RGH | RGD | A | Gs | E | WUEi | Chlorophyll Content | ||
| Salinity | 1 | 0.5807 | 0.2407 | 0.0025 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Fertility | 1 | <0.0001 | 0.0023 | <0.0001 | 0.0008 | 0.0133 | 0.1899 | <0.0001 |
| Family | 4 | 0.0004 | 0.3738 | 0.9632 | 0.0375 | 0.3877 | 0.0054 | 0.0922 |
| Family × salinity | 4 | 0.6415 | 0.7013 | 0.622 | 0.0188 | 0.0494 | 0.079 | 0.3469 |
| Family × fertility | 4 | 0.7818 | 0.1669 | 0.0302 | 0.1636 | 0.0265 | 0.0246 | 0.4621 |
| Family × salinity × fertility | 4 | 0.1288 | 0.6976 | 0.8522 | 0.5166 | 0.9227 | 0.8606 | 0.5800 |
Table 3.
Analysis of variance (ANOVA )results for the response of different Senegalia senegal families to salinity and fertility for stomatal density and size, total biomass, specific leaf area (SLA), root/shoot, leaf gas exchanges (net photosynthetic rate (A), stomatal conductance (Gs), transpiration (E), and intrinsic water use efficiency (WUEi)), relative growth of height (RGH) and diameter (RGD), and chlorophyll content following a drought period.
Table 3.
Analysis of variance (ANOVA )results for the response of different Senegalia senegal families to salinity and fertility for stomatal density and size, total biomass, specific leaf area (SLA), root/shoot, leaf gas exchanges (net photosynthetic rate (A), stomatal conductance (Gs), transpiration (E), and intrinsic water use efficiency (WUEi)), relative growth of height (RGH) and diameter (RGD), and chlorophyll content following a drought period.
| Sources of Variation | df | p-Value | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stomatal Density | Stomata Size | Total Biomass | SLA | Root/Shoot | RGH | RGD | A | Gs | E | WUEi | Chlorophyll Content | ||
| Salinity | 1 | 0.0158 | 0.2604 | <0.0001 | 0.0059 | <0.0001 | 0.0435 | 0.6073 | 0.452 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Fertility | 1 | 0.6565 | 0.91 | <0.0001 | 0.6169 | <0.0001 | 0.623 | 0.0012 | 0.2827 | 0.0142 | 0.0057 | <0.0001 | <0.0001 |
| Family | 4 | 0.0091 | <0.0001 | <0.0001 | 0.0159 | 0.023 | 0.0421 | 0.0019 | 0.4944 | 0.7874 | 0.3642 | <0.0001 | 0.2614 |
| Family × Salinity | 4 | 0.2459 | 0.5518 | 0.0634 | 0.9215 | 0.2997 | 0.2108 | 0.4326 | 0.0438 | 0.1426 | 0.5021 | 0.233 | 0.6956 |
| Family × fertility | 4 | 0.7811 | 0.0733 | 0.6723 | 0.2896 | 0.1454 | 0.1914 | 0.5758 | 0.9961 | 0.4089 | 0.514 | 0.2803 | 0.5889 |
| Family × Salinity × fertility | 4 | 0.5933 | 0.6437 | 0.0002 | 0.2343 | 0.7375 | 0.0925 | 0.1375 | 0.4873 | 0.1361 | 0.3306 | 0.0410 | 0.7770 |
Table 4.
The effect of salt and fertilizer treatments on growth parameters of Senegalia Senegal families.
Table 4.
The effect of salt and fertilizer treatments on growth parameters of Senegalia Senegal families.
| Variable | Treatment | Families | ||||
|---|---|---|---|---|---|---|
| DB14 | DB16 | K4B19 | K17B19 | NB1 | ||
| Leaf (g) | Control | 1.4 ± 0.47Aa | 1.7 ± 0.28Aab | 2.3 ± 0.61ABab | 1.5 ± 0.17Aa | 3.7 ± 0.40Bb |
| Salt | 1.4 ± 0.24Aa | 1.6 ± 0.12Aa | 1.7 ± 0.18Aa | 2.1 ± 0.16Aab | 1.6 ± 0.17Aa | |
| Fert | 4.2 ± 0.11Ab | 4.9 ± 0.45Ac | 4.2 ± 0.33Abc | 4.8 ± 0.41Ac | 5.3 ± 0.30Ab | |
| Salt × fert | 2.9 ± 0.09BCab | 2.3 ± 0.21Aab | 3.7 ± 0.36ABbc | 3.3 ± 0.30ABCbc | 4.2 ± 0.32Cb | |
| Stem (g) | Control | 1.3 ± 0.40Aa | 1.9 ± 0.34Aa | 1.7 ± 0.47Aa | 1.2 ± 0.16Aa | 2.7 ± 0.23Aab |
| Salt | 1.3 ± 0.23Aa | 2.0 ± 0.11Ba | 1.8 ± 0.14ABa | 2.1 ± 0.15Bab | 1.5 ± 0.17ABa | |
| Fert | 3.9 ± 0.24Ab | 5.3 ± 0.65Ac | 4.8 ± 0.62Ab | 5.0 ± 0.52Ac | 5.2 ± 0.25Ac | |
| Salt × fert | 3.0 ± 0.17ABab | 2.5 ± 0.21Aab | 4.0 ± 0.35Bb | 3.3 ± 0.31ABbc | 4.2 ± 0.35Bbc | |
| Root (g) | Control | 2.0 ± 0.35ABab | 2.7 ± 0.17ABab | 2.9 ± 0.45ABa | 1.7 ± 0.25Aa | 3.3 ± 0.29Bab |
| Salt | 1.5 ± 0.30Aa | 2.4 ± 0.22ABab | 2.6 ± 0.38Ba | 2.5 ± 0.25ABab | 1.7 ± 0.07ABa | |
| Fert | 3.9 ± 0.36Ac | 5.4 ± 0.40Ac | 5.2 ± 0.34Ac | 5.1 ± 0.68Ac | 5.2 ± 0.20Ac | |
| Salt × fert | 1.8 ± 0.19Aab | 1.7 ± 0.15Aa | 3.3 ± 0.14Bab | 2.0 ± 0.22Aab | 3.6 ± 0.41Babc | |
| Leaf area (cm2) | Control | 242.2 ± 111.3Aa | 279.9 ± 57.1Aa | 366.9 ± 106.9Aa | 295.1 ± 52.9Aa | 551.5 ± 61.6Aab |
| Salt | 263.2 ± 58.7Aa | 237.7 ± 22.4Aa | 380.6 ± 90.6Aa | 346.0 ± 72.1Aab | 226.2 ± 50.5Aa | |
| Fert | 891.6 ± 80.6Ac | 909.9 ± 113.8Ac | 686.8 ± 111.3Aab | 818.0 ± 134.8Ac | 756.1 ± 106.1Ab | |
| Salt × fert | 435.1 ± 74.0Aab | 323.9 ± 38.4Aab | 560.8 ± 118.0Aab | 522.5 ± 49.2Aabc | 546.6 ± 92.7Aab | |
| Total dry matter (g) | Control | 4.6 ± 1.16Aa | 6.2 ± 0.75ABa | 6.8 ± 1.41ABab | 4.4 ± 0.49Aa | 9.7 ± 0.76Bb |
| Salt | 4.2 ± 0.71Aa | 6.0 ± 0.22ABa | 6.2 ± 0.63ABa | 6.7 ± 0.53Bab | 4.8 ± 0.38Aa | |
| Fert | 12.0 ± 0.57Ab | 15.6 ± 1.39Ab | 14.1 ± 0.90Acd | 14.9 ± 1.51Ac | 15.7 ± 0.43Ac | |
| Salt × fert | 7.8 ± 0.30ABab | 6.5 ± 0.45Aa | 11.0 ± 0.77BCbc | 8.6 ± 0.80ABb | 12.0 ± 0.99Cbc | |
For each variable, values in each column (between treatment) with different small letters are significantly different (p < 0.05 Tukey test). Values in each line (between families) with different capital letters are significantly different (p < 0.05 Tukey test). Values represent the mean of five replicates ± standard error.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sarr, M.S.; Seiler, J.R.; Sullivan, J. Growth and Physiology of Senegalia senegal (L.) Britton Seedlings as Influenced by Seed Origin and Salinity and Fertility Treatments. Forests 2017, 8, 388. https://doi.org/10.3390/f8100388
AMA Style
Sarr MS, Seiler JR, Sullivan J. Growth and Physiology of Senegalia senegal (L.) Britton Seedlings as Influenced by Seed Origin and Salinity and Fertility Treatments. Forests. 2017; 8(10):388. https://doi.org/10.3390/f8100388
Chicago/Turabian StyleSarr, Mame Sokhna, John R. Seiler, and Jay Sullivan. 2017. "Growth and Physiology of Senegalia senegal (L.) Britton Seedlings as Influenced by Seed Origin and Salinity and Fertility Treatments" Forests 8, no. 10: 388. https://doi.org/10.3390/f8100388
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.