Compatible System for Predicting Total and Merchantable Stem Volume over and under Bark, Branch Volume and Whole-Tree Volume of Pine Species

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data
2.2. Compatible System Fitting
2.3. Model Fitting
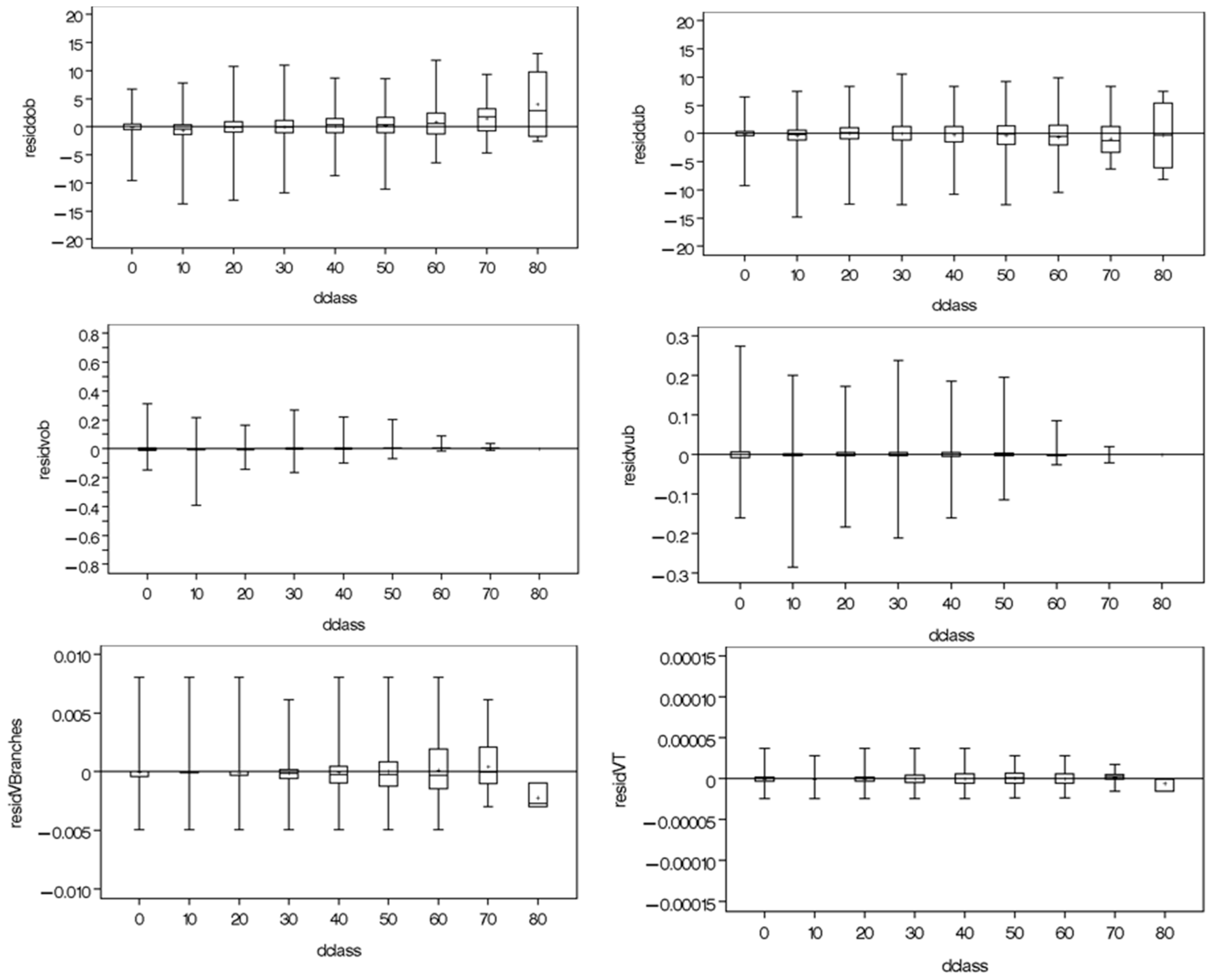
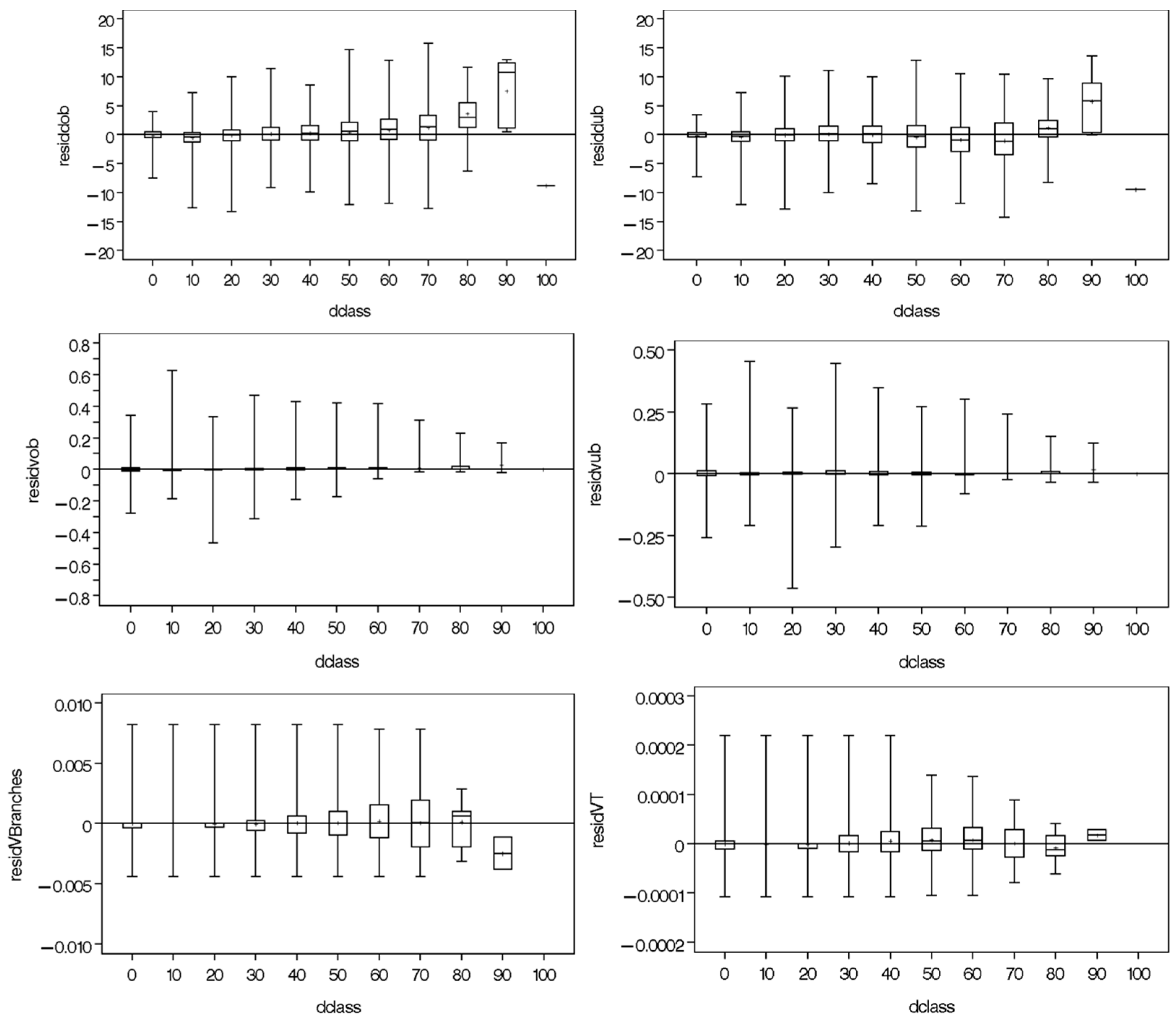
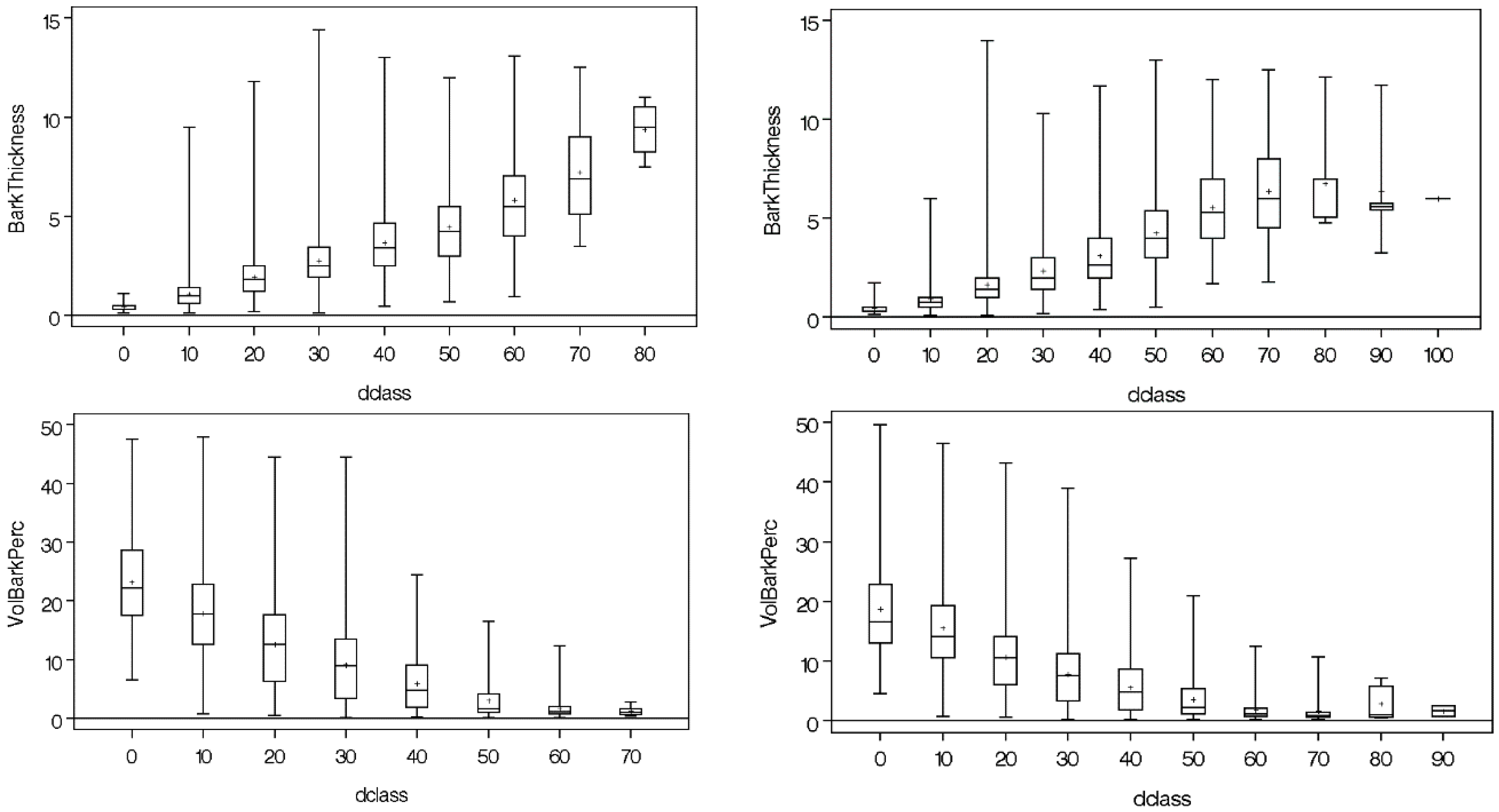
2.4. Autocorrelation and Heteroscedasticity
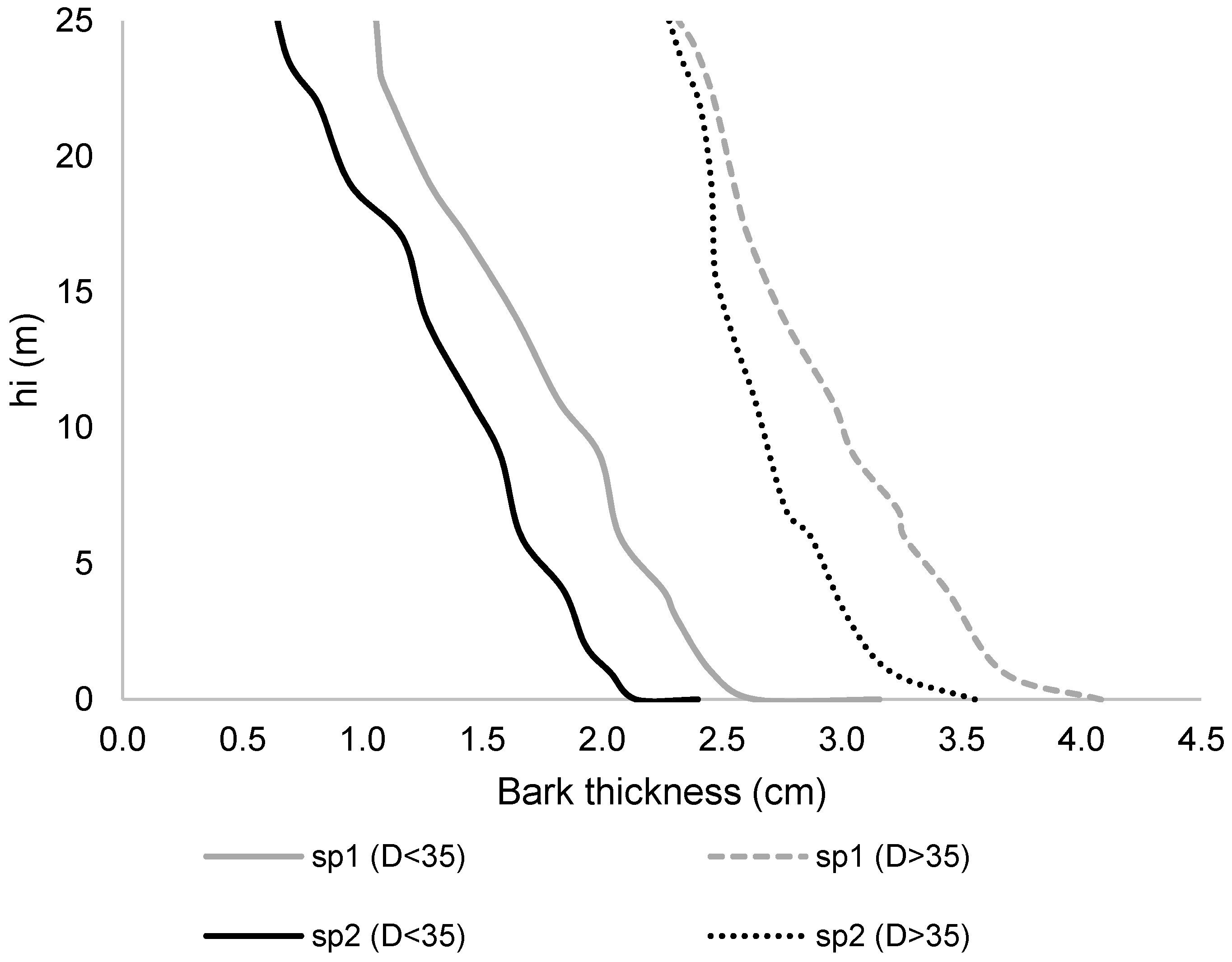
2.5. Effect of Crown Variables on Stem Form Variation
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ozcelik, R. Comparison of formulae for estimating tree bole volumes of Pinus sylvestris. Scand. J. For. Res. 2008, 23, 412–418. [Google Scholar] [CrossRef]
- MacFarlane, D. Predicting branch to bole volume scaling relationships from varying centroids of tree bole volume. Can. J. For. Res. 2010, 40, 2278–2289. [Google Scholar] [CrossRef]
- Planck, N.; MacFarlane, D. Modelling vertical allocation of tree stem and branch volume for hardwoods. Forestry 2014, 87, 459–469. [Google Scholar] [CrossRef]
- Flewelling, J. Compatible Taper Algorithms for California Hardwoods. In Proceedings of the Seventh Annual Forest Inventory and Analysis Symposium, Portland, ME, USA, 3–6 October 2005; McRoberts, R.E., Reams, G.A., Van Deusen, P.C., McWilliams, W.H., Eds.; Gen. Tech. Rep. WO-77. U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2007; pp. 265–267. [Google Scholar]
- MacFarlane, D. Allometric scaling of large branch volume in hardwood trees in Michigan, USA: Implications for aboveground forest carbon stock inventories. For. Sci. 2011, 57, 451–459. [Google Scholar]
- Zakrzewski, W. Estimating wood volume of the stem and branches of sugar maple (Acer saccharum marsh) using a stem profile model with implicit height. For. Sci. 2011, 57, 117–133. [Google Scholar]
- MacFarlane, D.; Weiskittel, A. A new method for capturing stem taper variation for trees of diverse morphological types. Can. J. For. Res. 2016, 46, 804–815. [Google Scholar] [CrossRef]
- Egnell, G.; Valinger, E. Survival, Growth, and Growth Allocation of Planted Scots Pine Trees after Different Levels of Biomass Removal in Clear-Felling. For. Ecol. Manag. 2003, 177, 65–74. [Google Scholar] [CrossRef]
- Merino, A.; Balboa, M.; Rodríguez-Soalleiro, R.; Álvarez-González, J. Nutrient exports under different harvesting regimes in fast-growing forest plantations in southern Europe. For. Ecol. Manag. 2005, 207, 325–339. [Google Scholar] [CrossRef]
- Pérez, S.; Renedo, C.; Ortiz, A.; Manana, M. Energy potential of waste from 10 forest species in the North of Spain (Cantabria). Bioresour. Technol. 2008, 99, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cruzado, C.; Rodríguez-Soalleiro, R. Improvement in accuracy of aboveground biomass estimation in Eucalyptus nitens plantations: Effect of bole sampling intensity and explanatory variables. For. Ecol. Manag. 2011, 261, 16–28. [Google Scholar] [CrossRef]
- Pérez-Cruzado, C.; Merino, A.; Rodríguez-Soalleiro, R. A management tool for estimating bioenergy production and carbon sequestration in Eucalyptus globulus and Eucalyptus nitens grown as short rotation woody crops in north-west Spain. Biomass Bioenergy 2011, 35, 2839–2851. [Google Scholar] [CrossRef]
- Vega-Nieva, D.; Valero, E.; Picos, J.; Jiménez, E. Modeling the above and belowground biomass of planted and coppiced Eucalytpus globulus stands in NW Spain. Ann. For. Sci. 2015, 72, 967–980. [Google Scholar] [CrossRef]
- Gasol, C.; Martínez, S.; Rigola, M.; Rieradeval, J.; Antón, A.; Carrasco, J. Feasibility assessment of poplar bioenergy systems in the Southern Europe. Renew. Sustain. Energy Rev. 2008, 13, 801–812. [Google Scholar] [CrossRef]
- Veláquez-Martí, B.; Estornell, J.; Lopez-Cortés, I.; Martí-Gavilá, I. Calculation of biomass volume of citrus trees from an adapted dendrometry. Biosyst. Eng. 2012, 112, 285–292. [Google Scholar] [CrossRef]
- Vega-Nieva, D.; Ortiz, L.; García-Maraver, A. Physical transformation processes of forest biomass: Chipping, bundling, drying and control of particle size and granulometric distribution. In Biomass Pelletization: Standards and Production; Garcia-Maraver, A., Ed.; Wit Press: Ashurst Lodge, UK, 2014; ISBN 978-1-78466-062-8. [Google Scholar]
- Tavankar, F.; Eynollahi, Y. Estimating of Volume and Weight of Logging Residue from Selection Cutting Under Different Ground Slopes in Hyrcanian Forest. Scholars Acad. J. Biosci. 2014, 2, 172–176. [Google Scholar]
- Biging, G. Taper equation for second-growth mixed conifers of northern California. For. Sci. 1984, 30, 1103–1117. [Google Scholar]
- Rojo, A.; Perales, X.; Sanchez-Rodriguez, F.; Alvarez-Gonzalez, J.; Gadow, K. Stem taper functions for maritime pine (Pinus pinaster Ait) in Galicia (northwestern Spain). Eur. J. For. Res. 2005, 25, 177–186. [Google Scholar] [CrossRef]
- Brooks, J.; Jiang, L.; Ozcelik, R. Compatible stem volume and taper equations for Brutian pine, Cedar of Lebanon, and Cilicica fir in Turkey. For. Ecol. Manag. 2008, 256, 147–151. [Google Scholar] [CrossRef]
- Crecente-Campo, F.; Rojo, A.; Diéguez-Aranda, U. A merchantable volume system for Pinus sylvestris L. in the major mountains ranges of Spain. Ann. For. Sci. 2009, 66, 808. [Google Scholar] [CrossRef]
- Nunes, L.; Tomé, J.; Tomé, M. A system for compatible prediction of total and merchantable volumes allowing for different definitions of tree volume. Can. J. For. Res. 2010, 40, 747–760. [Google Scholar] [CrossRef]
- Arias-Rodil, M.; Diéguez-Aranda, U.; Rodríguez Puerta, F.; López-Sánchez, C.; Canga Líbano, E.; Cámara Obregón, A. Modelling and localizing a stem taper function for Pinus radiata in Spain. Can. J. For. Res. 2015, 45, 647–658. [Google Scholar] [CrossRef]
- Newnham, R. Variable-form taper functions for four Alberta tree species. Can. J. For. Res. 1992, 22, 210–223. [Google Scholar] [CrossRef]
- Muhairwe, C.; Le May, V.; Kozak, A. Effects of adding tree, stand and site variables to Kozak’s variable-exponent taper equation. Can. J. For. Res. 1994, 24, 252–269. [Google Scholar] [CrossRef]
- Kozak, A. A variable-exponent taper equation. Can. J. For. Res. 1988, 18, 1363–1368. [Google Scholar] [CrossRef]
- Kozak, A. My last words on taper equations. For. Chron. 2004, 80, 507–515. [Google Scholar] [CrossRef]
- Bi, H. Trigonometric variable-form taper equations for Australian eucalyptus. For. Sci. 2000, 46, 397–409. [Google Scholar]
- Sharma, M.; Zhang, S. Variable-exponent taper equation for jack pine, black spruce, and balsam fir in eastern Canada. For. Ecol. Manag. 2004, 198, 39–53. [Google Scholar] [CrossRef]
- Max, T.; Burkhart, H. Segmented polynomial regression applied to taper equations. For. Sci. 1976, 22, 283–289. [Google Scholar]
- Demaerschalk, P.; Kozak, A. The Whole-bole system: A conditioned dual-equation system for precise prediction of tree profiles. Can. J. For. Res. 1977, 7, 488–497. [Google Scholar] [CrossRef]
- Byrne, J.; Reed, D. Complex compatible taper and volume estimation systems for red and loblolly pine. For. Sci. 1986, 32, 423–443. [Google Scholar]
- Fang, Z.B.; Borders, E.; Bailey, L. Compatible volume-taper models for loblolly and slash pine based on a system with segmented stem form factors. For. Sci. 2000, 46, 1–12. [Google Scholar]
- Mäkelä, A. Derivation of stem taper from the pipe theory in a carbon balance framework. Tree Physiol. 2002, 22, 891–905. [Google Scholar] [CrossRef] [PubMed]
- Valentine, H.; Mäkelä, A. Bridging process-based and empirical approaches to modelling tree growth. Tree Physiol. 2005, 25, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Valentine, H.; Mäkelä, A.; Green, E.; Amateis, R.; Mäkinen, H.; Duceyet, M. Models relating stem growth to crown length dynamics: Application to loblolly pine and Norway spruce. Trees 2012, 26, 469–478. [Google Scholar] [CrossRef]
- Demaerschalk, J.; Demaerschalk, J.P. Converting volume equations to compatible taper equations. For. Sci. 1972, 18, 241–245. [Google Scholar]
- Clutter, J. Development of taper functions from variable top merchantable volume equations. For. Sci. 1980, 6, 117–120. [Google Scholar]
- Diéguez-Aranda, U.; Castedo-Dorado, F.; Álvarez-González, J.; Rojo, A. Compatible taper function for Scots pine plantations in northwestern Spain. Can. J. For. Res. 2006, 36, 1190–1205. [Google Scholar] [CrossRef]
- Barrio-Anta, M.; Dieguez-Aranda, U.; Castedo-Dorado, F.; Álvarez-Gonzalez, J.; Gadow, K. Merchantable volume system for pedunculate oak in northwestern Spain. Ann. For. Sci. 2007, 64, 511–520. [Google Scholar] [CrossRef]
- Sharma, M.; Oderwald, R.; Amateis, R. A consistent system of equations for tree and stand volume. For. Ecol. Manag. 2002, 165, 183–191. [Google Scholar] [CrossRef]
- Demaerschalk, J. Integrated systems for the estimation of tree taper and volume. Can. J. For. Res. 1973, 3, 90–94. [Google Scholar] [CrossRef]
- Goulding, C.; Murray, J. Polynomial taper equations that are compatible with tree volume equations. N. Z. J. For. Sci. 1976, 5, 313–322. [Google Scholar]
- Li, R.; Weiskittel, A. Comparison of model forms for estimating stem taper and volume in the primary conifer species of the North American Acadian Region. Ann. For. Sci. 2010, 67, 302. [Google Scholar] [CrossRef]
- Antonio, N.; Tomé, M.; Tomé, J.; Soares, P.; Fontes, L. Effect of tree, stand, and site variables on the allometry of Eucalyptus globulus tree biomass. Can. J. For. Res. 2007, 37, 895–906. [Google Scholar] [CrossRef]
- Van Deusen, P.; Roesch, F. Sampling a tree for total volume, biomass, and carbon. J. For. 2011, 109, 131–135. [Google Scholar]
- MacFarlane, D.; Kuyah, S.; Mulia, R.; Dietz, J.; Muthuri, C.; Van Noordwijk, M. Evaluating a non-destructive method for calibrating tree biomass equations derived from tree branching architecture. Trees 2014, 28, 807–817. [Google Scholar] [CrossRef]
- MacFarlane, D. A generalized tree component biomass model derived from principles of variable allometry. For. Ecol. Manag. 2015, 354, 43–55. [Google Scholar] [CrossRef]
- Gómez-García, E.; Biging, G.; García-Villabrille, J.; Crecente-Campo, F.; Castedo-Dorado, F.; Rojo-Alboreca, A. Cumulative continuous predictions for bole and aboveground woody biomass in Eucalyptus globulus plantations in northwestern Spain. Biomass Bioenergy 2015, 77, 155–164. [Google Scholar] [CrossRef]
- Larson, P.R. Stem form development of forest trees. For. Sci. 1963, 9, 1–15. [Google Scholar]
- Kershaw, A.; Maguire, A. Influence of vertical foliage structure on the distribution of stem cross-sectional area increment in western hemlock and balsam fir. For. Sci. 2000, 46, 86–94. [Google Scholar]
- Leites, L.; Robinson, A. Improving taper equations of loblolly pine with crown dimension in a mixed-effects modelling framework. For. Sci. 2004, 50, 204–212. [Google Scholar]
- Adu-Bredu, S.; Foua Tape Bi, A.; Bouillet, J.P.; Kouamé Mé, M.; Yamoah Kyei, S.; Saint André, L. An explicit stem profile model for forked and un-forked teak (Tectona grandis) trees in West Africa. For. Ecol. Manag. 2008, 255, 2189–2203. [Google Scholar] [CrossRef]
- Valentine, H.; Gregoire, T. A switching model of bole taper. Can. J. For. Res. 2001, 31, 1400–1409. [Google Scholar] [CrossRef]
- Navar, J.; Domínguez, P.; Contreras, J.; Estrada, M. Ajuste de siete modelos de ahusamiento a los perfiles fustales de Pinus hartwegii Lindl. del Noroeste de México. Agrociencia 1997, 31, 73–81. [Google Scholar]
- Corral-Rivas, J.; Diéguez-Aranda, U.; Corral-Rivas, S.; Castedo-Dorado, F. A merchantable volume system for major pine species in El Salto, Durango (Mexico). For. Ecol. Manag. 2007, 238, 118–129. [Google Scholar] [CrossRef]
- Pompa-García, M.; Corral-Rivas, J.; Hernández-Díaz, J.; Álvarez-González, J. A system for calculating the merchantable volume of oak trees in the northwest of the state of Chihuahua, Mexico. J. For. Res. 2009, 20, 293–300. [Google Scholar] [CrossRef]
- Quiñonez-Barraza, G.; De los Santos-Posadas, H.; Álvarez-González, J.; Velázquez-Martínez, A. Compatible taper and merchantable volume system for major pine species in Durango, Mexico. Agrociencia 2014, 48, 553–567. [Google Scholar]
- Silva-Flores, R.; Pérez-Verdín, G.; Wehenkel, C. Patterns of Tree Species Diversity in Relation to Climatic Factors on the Sierra Madre Occidental, Mexico. PLoS ONE 2014, 8, e105034. [Google Scholar]
- Wehenkel, C.; Simental-Rodriguez, L.; Silva-Flores, R.; Hernández-Diaz, C.; López-Sánchez, C.; Antunez, P. Discrimination of 59 seed stands of various Mexican pine species based on 44 dendrometric, climatic, edaphic and genetic traits. Forstarchiv 2015, 86, 194–201. [Google Scholar]
- Abbot, P.; Lowore, J.; Werren, M. Models for the estimation of single tree volume in four Miombo woodland types. For. Ecol. Manag. 1997, 97, 25–37. [Google Scholar] [CrossRef]
- Harvey, A. Estimating regression models with multiplicative heterocedasticity. Econometrica 1976, 44, 461–465. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/ETS User’s Guide, 9.2; SAS Institute Inc.: Cary, NC, USA, 2008. [Google Scholar]
- Crecente-Campo, F.; Tomé, M.; Soares, P.; Diéguez-Aranda, U. A generalized nonlinear mixed-effects height-diameter model for Eucalyptus globulus L. in northwestern Spain. For. Ecol. Manag. 2010, 258, 943–952. [Google Scholar] [CrossRef]
- González-García, M.; Hevia, A.; Majada, J.; Barrio-Anta, M. Above-ground biomass estimation at tree and stand level for short rotation plantations of Eucalyptus nitens (Deane & Maiden) Maiden in Northwest Spain. Biomass Bioenergy 2013, 54, 147–157. [Google Scholar]
- Jiménez, E.; Vega, J.; Fernández-Alonso, J.; Vega-Nieva, D.; Álvarez-González, J.; Ruiz-González, A. Allometric equations for estimating canopy fuel load and distribution of pole-size maritime pine trees in five Iberian provenances. Can. J. For. Res. 2013, 43, 149–158. [Google Scholar] [CrossRef]
- Muñoz, F.; Rubilar, R.; Espinosa, M.; Cancino, J.; Toro, J.; Herrera, M. The effect of pruning and thinning on above ground aerial biomass of Eucalyptus nitens (Deane & Maiden) Maiden. For. Ecol. Manag. 2008, 255, 365–373. [Google Scholar]
- Weiskittel, A.; Maguire, D. Branch surface area and its vertical distribution in coastal Douglas-fir. Trees Struct. Funct. 2006, 20, 657–667. [Google Scholar] [CrossRef]
- Fontes, L.; Landsberg, J.; Tomé, J.; Tomé, M.; Pacheco, C.; Soares, P. Calibration and testing of a generalized process-based model for use in Portuguese eucalyptus plantations. Can. J. For. Res. 2006, 36, 3209–3221. [Google Scholar] [CrossRef]
- Castedo-Dorado, F.; Diéguez-Aranda, U.; Álvarez-González, J. A growth model for Pinus radiata D. Don stands in northwestern Spain. Ann. For. Sci. 2007, 64, 453–465. [Google Scholar] [CrossRef]
- Garber, S.; Maguire, D. Modeling stem taper of three Central Oregon species using nonlinear mixed effects models and autoregressive error structure. For. Ecol. Manag. 2003, 179, 507–522. [Google Scholar] [CrossRef]
- Burkhart, H.; Walton, S. Incorporating crown ratio into taper equations for loblolly pine trees. For. Sci. 1985, 31, 478–484. [Google Scholar]
- Walters, D.K.; Hann, D.W. Taper Equations for Six Major Conifer Species in Southwest Oregon; Research Bulletin 56; Forest Research Laboratory, Oregon State University: Corvallis, OR, USA, 1986. [Google Scholar]
- Kozak, A.; Yang, R. Equations for estimating bark volume and thickness of commercial trees in British Columbia. For. Chron. 1981, 57, 112–115. [Google Scholar] [CrossRef]
- Malone, T.; Liang, J. A Bark Thickness Model for White Spruce in Alaska Northern Forests. Int. J. For. Res. 2009, 2009, 876965. [Google Scholar] [CrossRef]
- Assmann, E. The Principles of Forest Yield Study; Pergamon Press: Oxford, UK, 1970; 506p. [Google Scholar]
- Marshall, H.; Murphy, G.; Lachenbruch, B. Effects of bark thickness estimates on optimal log merchandising. For. Prod. J. 2006, 56, 87–93. [Google Scholar]
- Fernandes, P.; Vega, J.; Jiménez, E.; Rigolot, E. Fire resistance of European pines. For. Ecol. Manag. 2008, 256, 246–255. [Google Scholar] [CrossRef]
- Wehenkel, C.; Cruz-Cobos, F.; Carrillo, A.; Luján-Soto, J. Estimating bark volumes for 16 native tree species on the Sierra Madre Occidental, Mexico. Scand. J. For. Res. 2012, 27, 578–585. [Google Scholar] [CrossRef]
- Contreras-Aviña, J.; Návar-Cháidez, J. Ecuaciones aditivas para estimar componentes de volumen para Pinus teocote Schl. de Durango, México. Rev. Cienc. For. Méx. 2002, 27, 67–81. [Google Scholar]
- Sims, R.; Venturi, P. All-year-round harvesting of short rotation coppice eucalyptus compared with the delivered costs of biomass from more conventional short season harvesting systems. Biomass Bioenergy 2004, 26, 27–37. [Google Scholar] [CrossRef]
- Murphy, G.; Pilkerton, S. Seasonal Impacts of Bark Loss on Simulated Payloads, Bark Delivery, and Transport Costs for Freshly Harvested Logs. For. Prod. J. 2011, 61, 71–76. [Google Scholar] [CrossRef]
- Jordan, L.; Souter, R.; Parresol, B.; Daniels, R. Application of the algebraic difference approach for developing self referencing specific gravity and biomass equations. For. Sci. 2006, 52, 81–92. [Google Scholar]
- Okai, R.; Frimpong-Mensah, K.; Yeboah, D. Characterization of moisture content and specific gravity of branchwood and stemwood of Aningeria robusta and Terminalia ivorensis. Eur. J. Wood Wood Prod. 2003, 61, 155–158. [Google Scholar] [CrossRef]
- Swenson, N.; Enquist, B. The relationship between stem and branch wood specific gravity and the ability of each measure to predict leaf area. Am. J. Bot. 2008, 95, 516–519. [Google Scholar] [CrossRef] [PubMed]
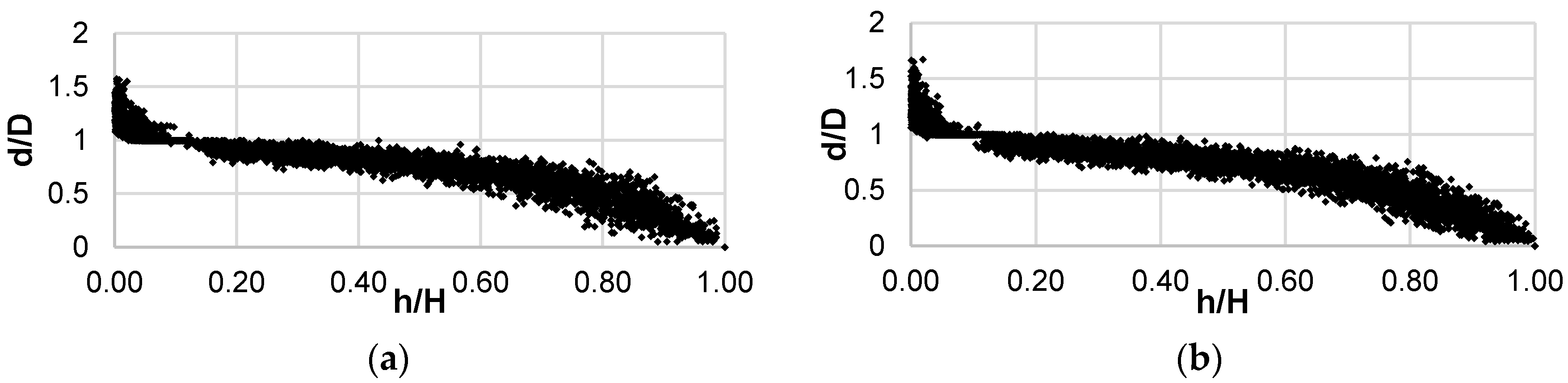

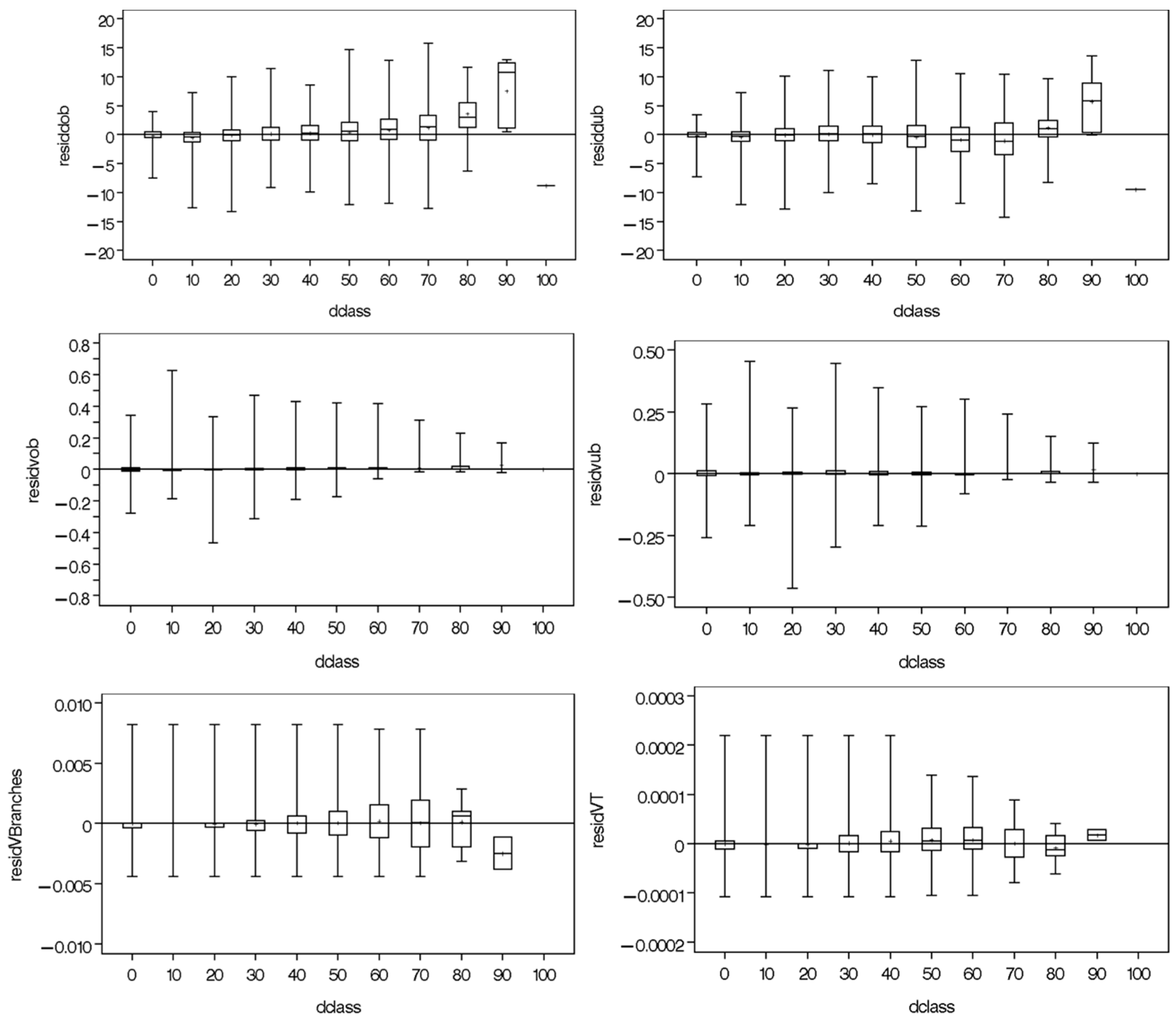

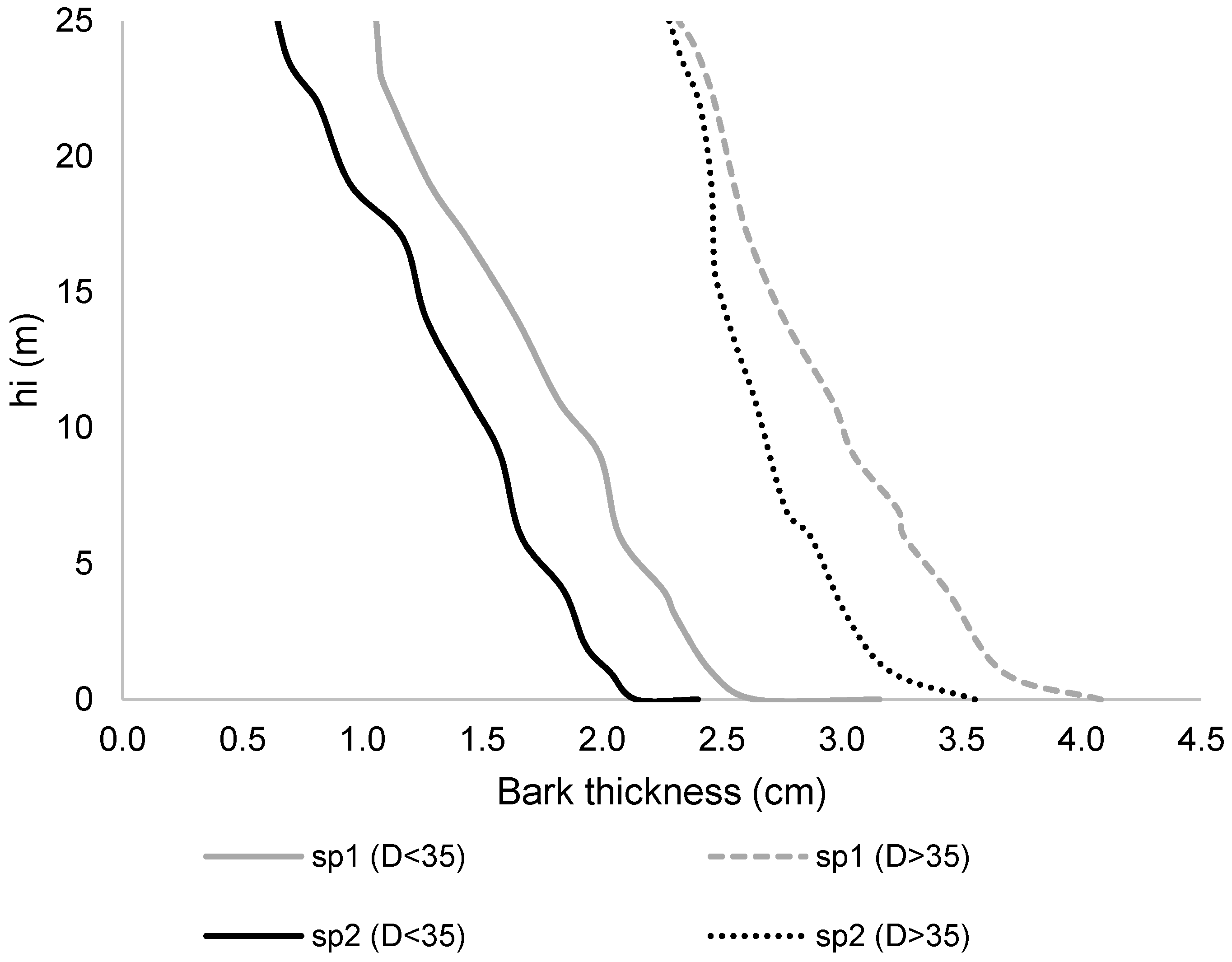

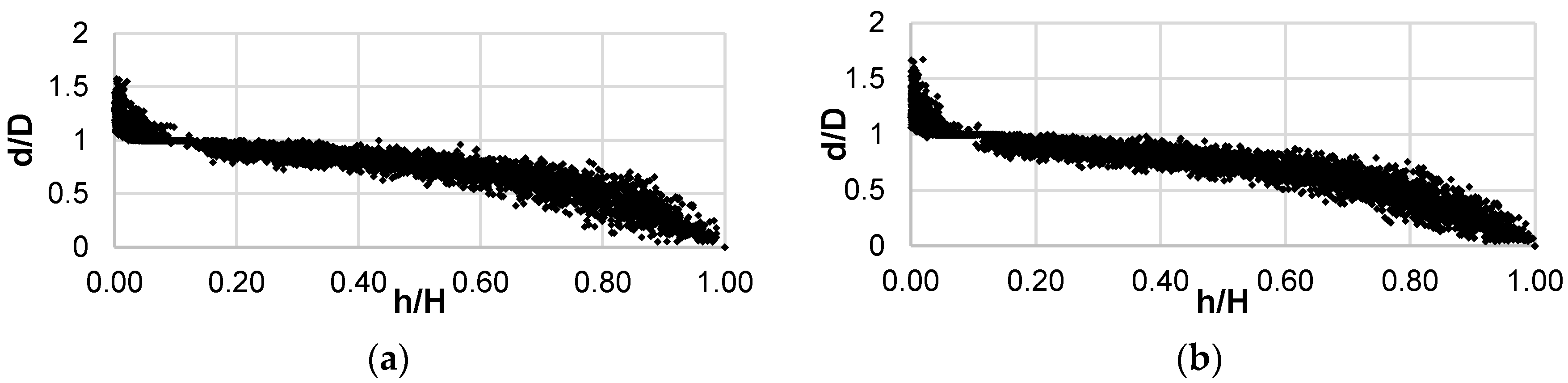{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Variable | N of Observations | Mean | Standard Deviation | Maximum | Minimum |
|---|---|---|---|---|---|---|
| Pinus cooperi | dob | 6090 | 27.9 | 15.5 | 84.0 | 0.0 |
| dub | 6090 | 25.4 | 14.1 | 75 | 0.0 | |
| D | 595 | 45.12 | 12.51 | 84.00 | 10.65 | |
| H | 595 | 19.15 | 4.74 | 33.23 | 6.14 | |
| vob | 595 | 1.29 | 0.89 | 5.55 | 0.02 | |
| vub | 595 | 1.10 | 0.78 | 5.05 | 0.02 | |
| Vbranches | 595 | 0.10 | 0.11 | 0.89 | 0.00 | |
| Vtotal | 595 | 1.39 | 0.96 | 5.74 | 0.02 | |
| Pinus duragensis | dob | 7724 | 28.0 | 15.9 | 88.0 | 0.0 |
| dub | 7724 | 25.6 | 14.5 | 82.25 | 0.0 | |
| D | 700 | 46.35 | 13.34 | 88.00 | 11.40 | |
| H | 700 | 20.50 | 4.99 | 35.42 | 7.43 | |
| vob | 700 | 1.42 | 1.04 | 5.72 | 0.03 | |
| vub | 700 | 1.23 | 0.92 | 4.87 | 0.02 | |
| Vbranches | 700 | 0.10 | 0.11 | 0.59 | 0.00 | |
| Vtotal | 700 | 1.52 | 1.12 | 6.31 | 0.04 |
| Equation | Variable | Species | RMSE | R2 | Durbin_Watson |
|---|---|---|---|---|---|
| (1) | dob | Pinus cooperi | 2.0868 | 0.9817 | 1.96 |
| Pinus durangensis | 2.2057 | 0.9802 | 1.98 | ||
| (2) | vob | Pinus cooperi | 0.0242 | 0.9989 | 2.03 |
| Pinus durangensis | 0.0309 | 0.9989 | 2.01 | ||
| (4) | dub | Pinus cooperi | 2.1444 | 0.9767 | 1.93 |
| Pinus durangensis | 2.0931 | 0.9789 | 1.97 | ||
| (5) | vub | Pinus cooperi | 0.0242 | 0.9985 | 2.04 |
| Pinus durangensis | 0.0303 | 0.9986 | 2.04 | ||
| (7) | Vbranches | Pinus cooperi | 0.0653 | 0.6411 | -- |
| Pinus durangensis | 0.0524 | 0.7012 | -- | ||
| (8) | Vtotal | Pinus cooperi | 0.1555 | 0.9771 | -- |
| Pinus durangensis | 0.1482 | 0.9832 | -- |
| Parameter | Equations | Pinus cooperi | Pinus durangensis | ||
|---|---|---|---|---|---|
| Estimate | Std. Error | Estimate | Std. Error | ||
| a0 | (1), (2), (3), (8) | 6.33 × 10−5 | 3.77 × 10−7 | 6.47 × 10−5 | 2.26 × 10−7 |
| a1 | 1.8946 | 0.0017 | 1.9326 | 0.0007 | |
| a2 | 1.0057 | 0.0019 | 0.9410 | 0.0013 | |
| b1 | (1), (2), (4), (5) | 6.79 × 10−6 | 5.38 × 10−8 | 5.31 × 10−6 | 4.50 × 10−8 |
| b2 | 4.35 × 10−5 | 6.67 × 10−8 | 4.23 × 10−5 | 4.26 × 10−8 | |
| b3 | 3.10 × 10−5 | 1.40 × 10−7 | 2.96 × 10−5 | 1.08 × 10−7 | |
| p1 | 0.0461 | 0.0003 | 0.0230 | 0.0002 | |
| p2 | 0.7133 | 0.0022 | 0.7353 | 0.0015 | |
| e0 | (4), (5), (6) | 3.49 × 10−5 | 2.68 × 10−7 | 4.73 × 10−5 | 2.24 × 10−7 |
| e1 | 1.9564 | 0.0023 | 1.9572 | 0.0012 | |
| e2 | 1.0686 | 0.0025 | 0.9652 | 0.0016 | |
| f0 | (7), (8) | 1.64 × 10−6 | 1.41 × 10−7 | 2.74 × 10−6 | 1.61 × 10−7 |
| f1 | 3.1004 | 0.0215 | 2.8844 | 0.0151 | |
| f2 | 0.2861 | 0.0047 | 0.1211 | 0.0043 | |
| Parameter | Pinus cooperi | Pinus durangensis | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean | Standard Deviation | Max. | Min. | Mean | Standard Deviation | Max. | Min. | |
| b1 | 9.1 × 10−6 | 6.1 × 10−6 | 6.0 × 10−4 | 3.5 × 10−7 | 8.6 × 10−6 | 5.2 × 10−6 | 4.4 × 10−5 | 2.8 × 10−7 |
| b2 | 4.6 × 10−5 | 6.4 × 10−6 | 7.5 × 10−5 | 3.0 × 10−5 | 4.4 × 10−5 | 5.4 × 10−5 | 7.5 × 10−5 | 3.3 × 10−5 |
| b3 | 3.2 × 10−5 | 7.8 × 10−6 | 5.5 × 10−5 | 7.3 × 10−6 | 2.9 × 10−5 | 8.9 × 10−6 | 6.9 × 10−5 | 2.4 × 10−6 |
| p1 | 0.0422 | 0.0139 | 0.0804 | 0.0073 | 0.0349 | 0.0129 | 0.0770 | 0.0009 |
| p2 | 0.5784 | 0.2025 | 0.9004 | 0.0163 | 0.6238 | 0.2143 | 0.9891 | 0.0158 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corral-Rivas, J.J.; Vega-Nieva, D.J.; Rodríguez-Soalleiro, R.; López-Sánchez, C.A.; Wehenkel, C.; Vargas-Larreta, B.; Álvarez-González, J.G.; Ruiz-González, A.D. Compatible System for Predicting Total and Merchantable Stem Volume over and under Bark, Branch Volume and Whole-Tree Volume of Pine Species. Forests 2017, 8, 417. https://doi.org/10.3390/f8110417
Corral-Rivas JJ, Vega-Nieva DJ, Rodríguez-Soalleiro R, López-Sánchez CA, Wehenkel C, Vargas-Larreta B, Álvarez-González JG, Ruiz-González AD. Compatible System for Predicting Total and Merchantable Stem Volume over and under Bark, Branch Volume and Whole-Tree Volume of Pine Species. Forests. 2017; 8(11):417. https://doi.org/10.3390/f8110417
Chicago/Turabian StyleCorral-Rivas, José Javier, Daniel Jose Vega-Nieva, Roque Rodríguez-Soalleiro, Carlos Antonio López-Sánchez, Christian Wehenkel, Benedicto Vargas-Larreta, Juan Gabriel Álvarez-González, and Ana Daría Ruiz-González. 2017. "Compatible System for Predicting Total and Merchantable Stem Volume over and under Bark, Branch Volume and Whole-Tree Volume of Pine Species" Forests 8, no. 11: 417. https://doi.org/10.3390/f8110417