Comparisons of Soil Properties, Enzyme Activities and Microbial Communities in Heavy Metal Contaminated Bulk and Rhizosphere Soils of Robinia pseudoacacia L. in the Northern Foot of Qinling Mountain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Soil Properties
2.3. Soil Enzyme Activities
2.4. DNA Extraction
2.5. Quantitative PCR
2.6. RFLP Analysis
2.7. Data Analysis
3. Results
3.1. Soil Properties
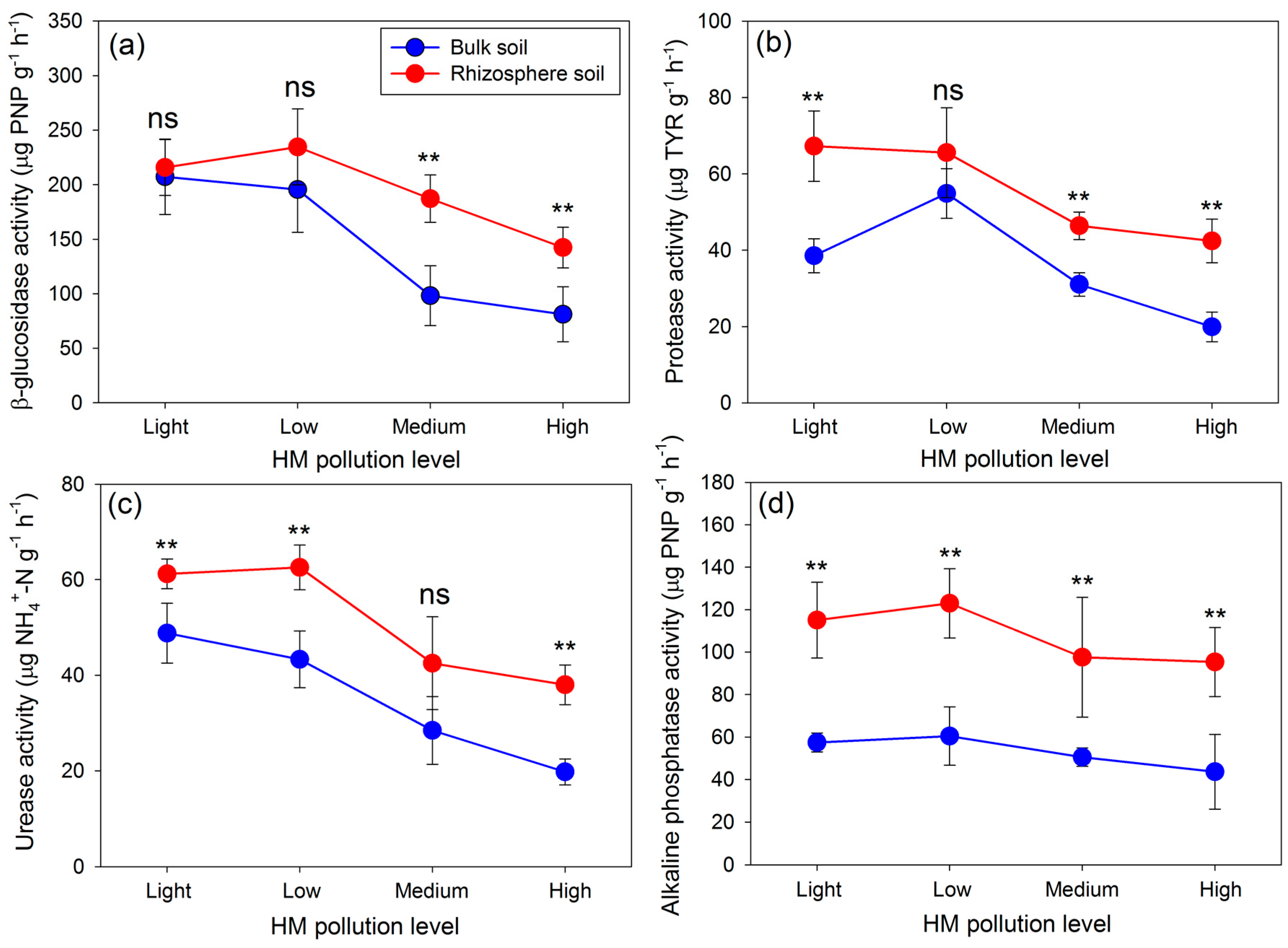
3.2. Soil Enzyme Activities
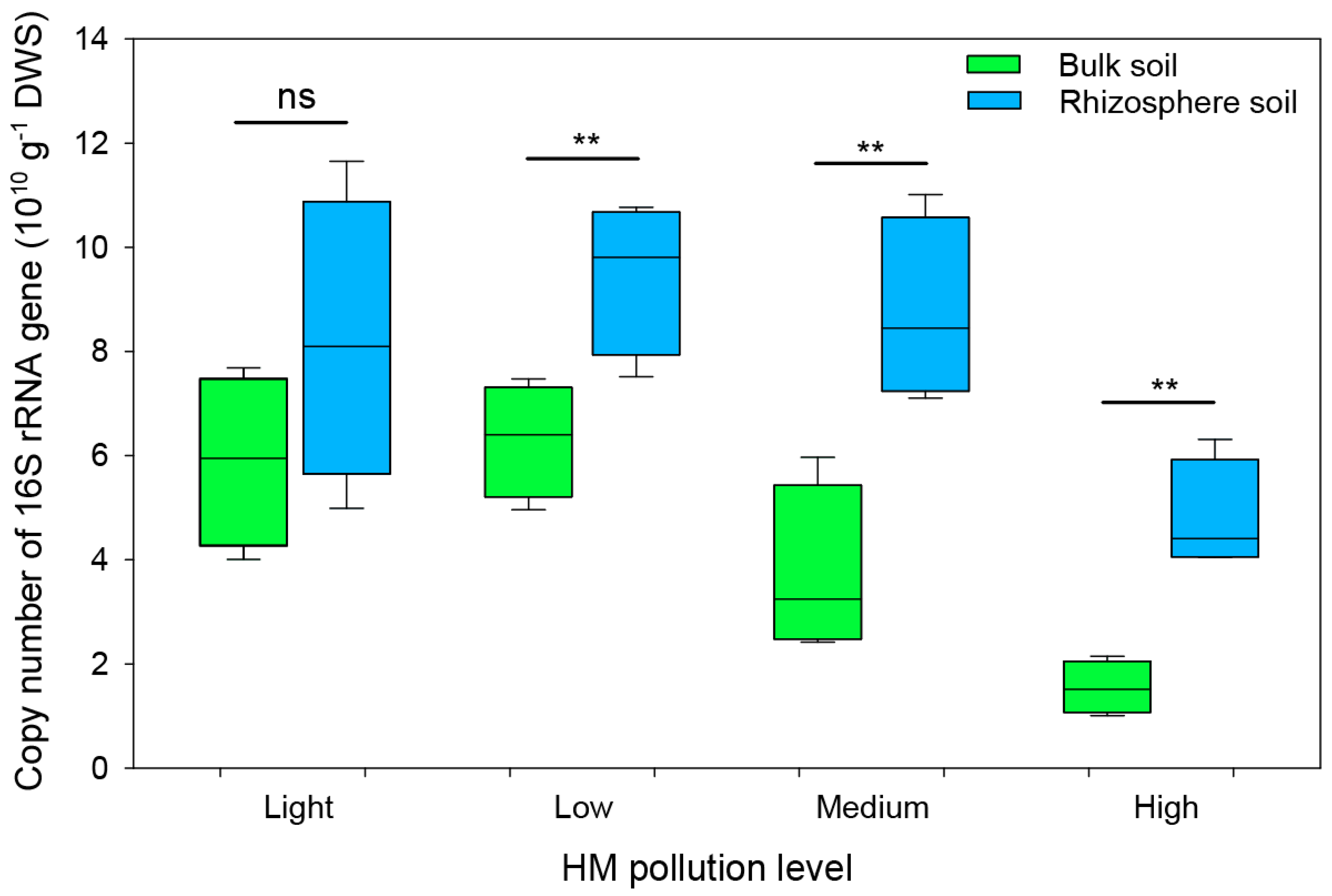
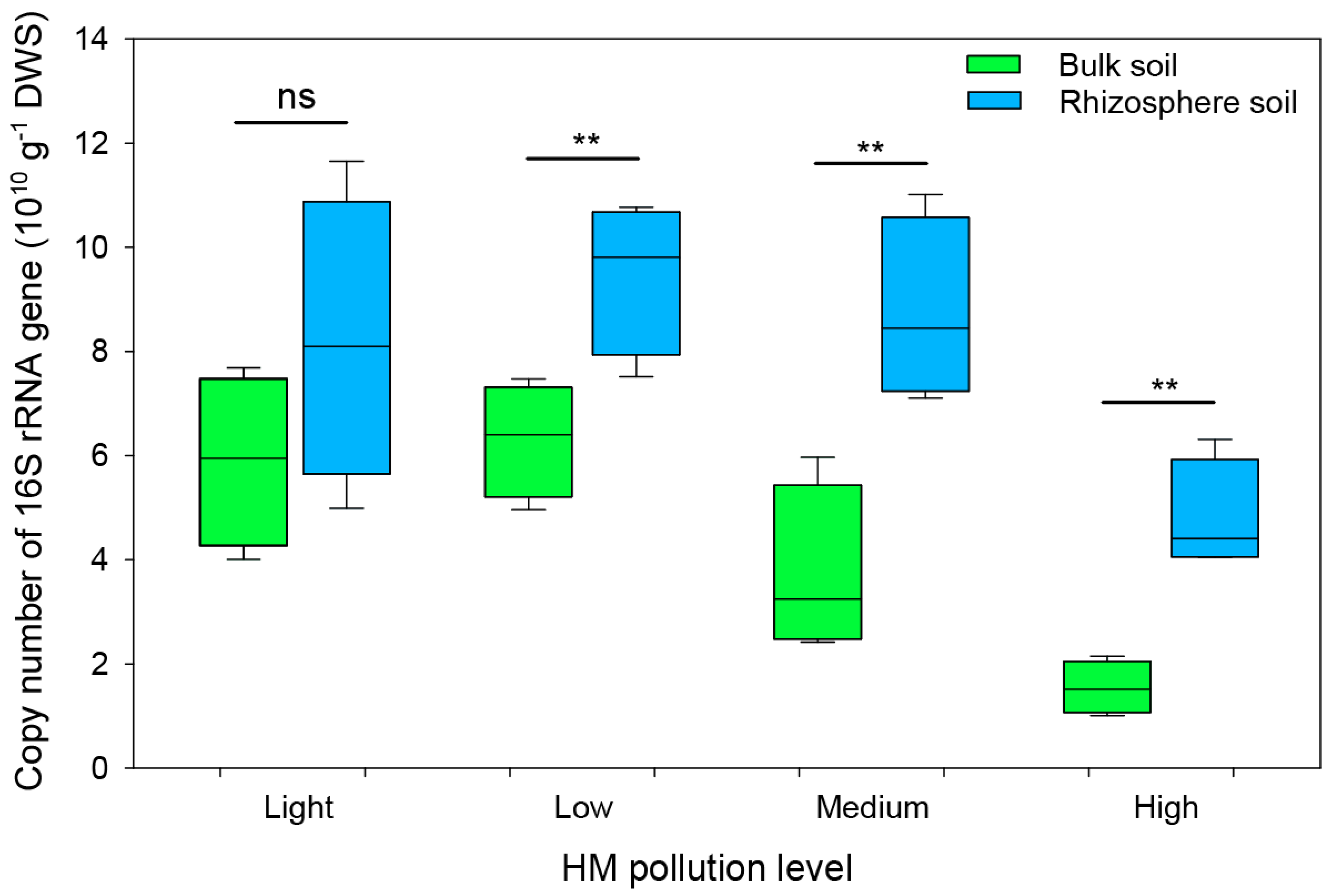
3.3. Soil Bacterial Abundance
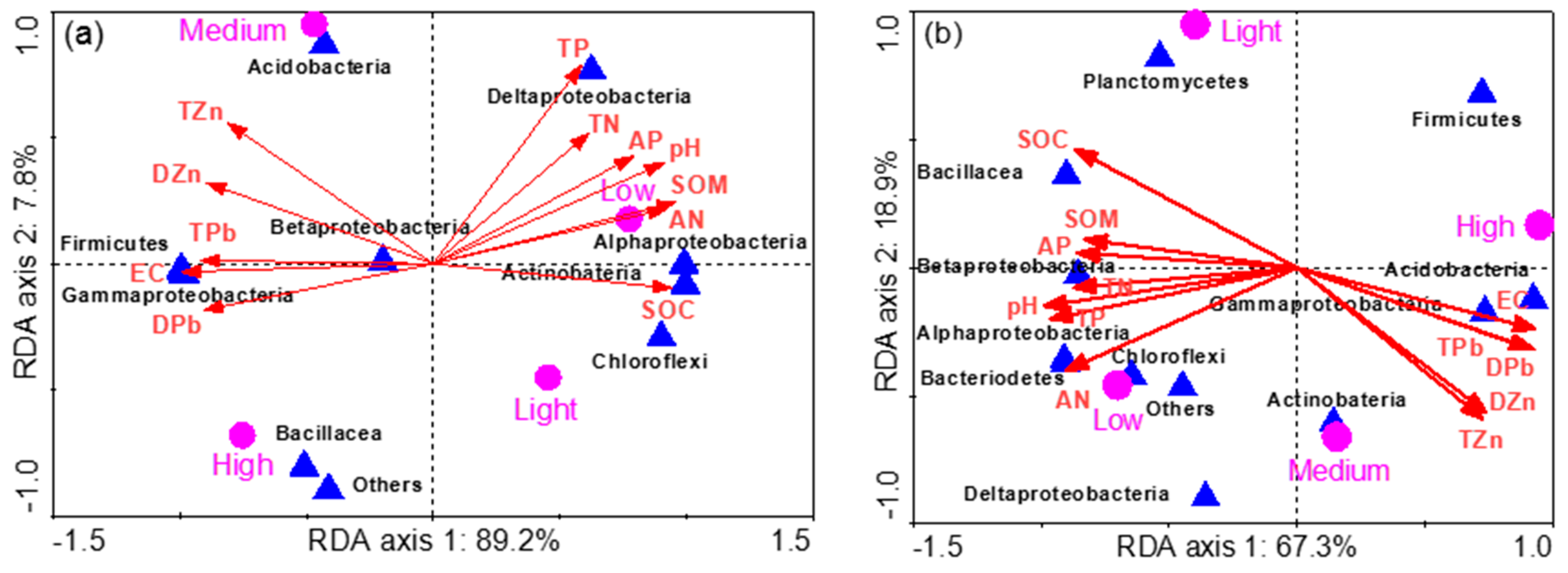
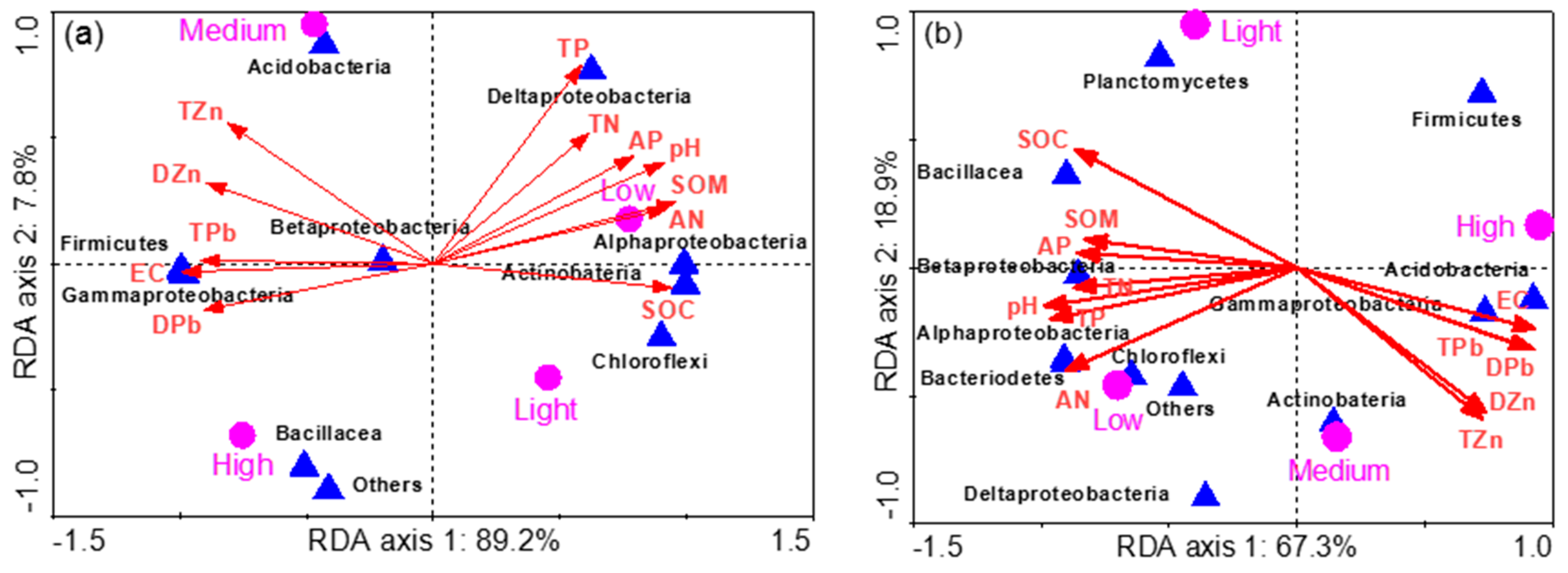
3.4. Phylogenetic Analysis
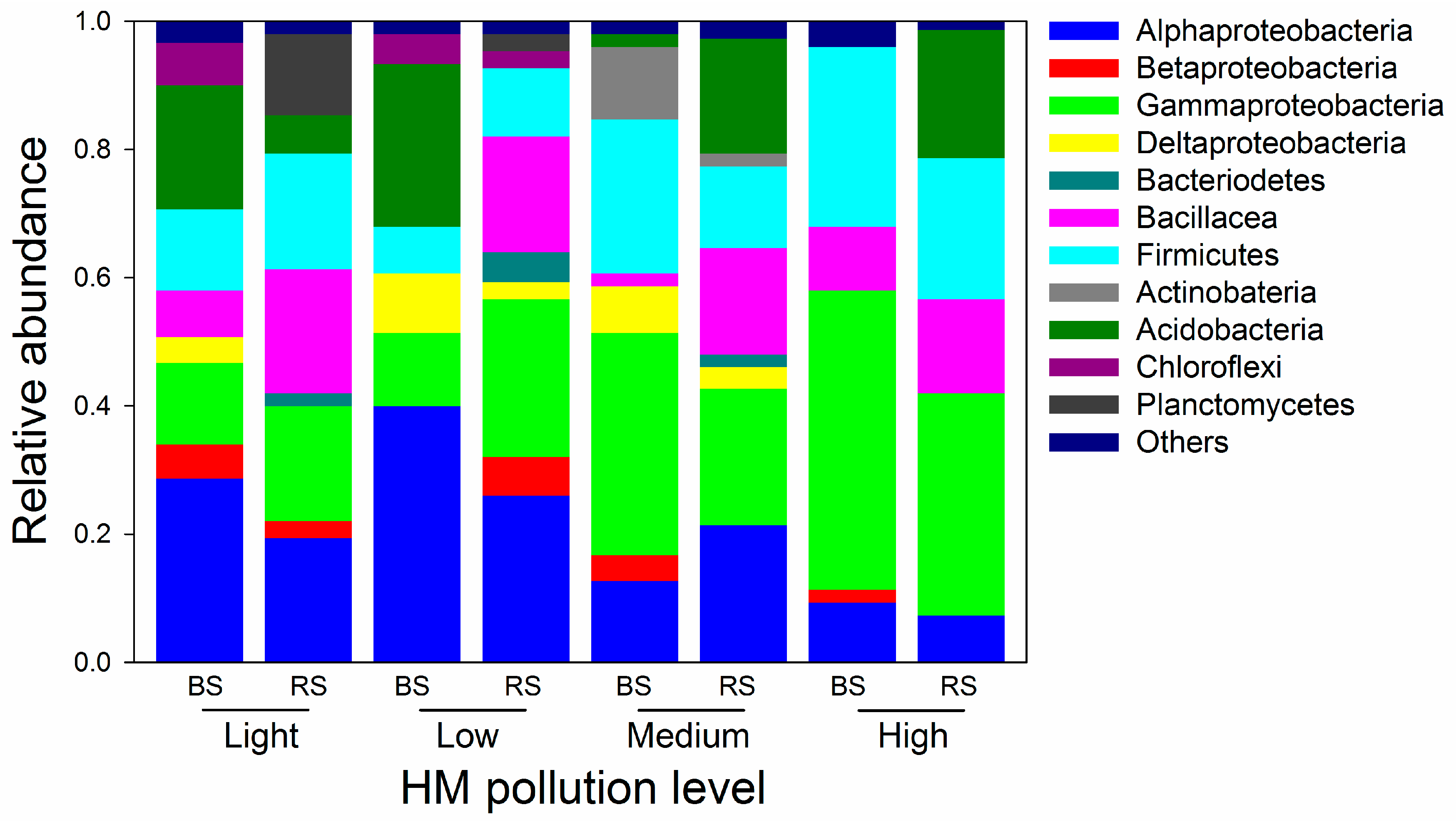
3.5. Bacterial Communities in Rhizosphere Soil
3.6. Bacterial Communities in Bulk Soil
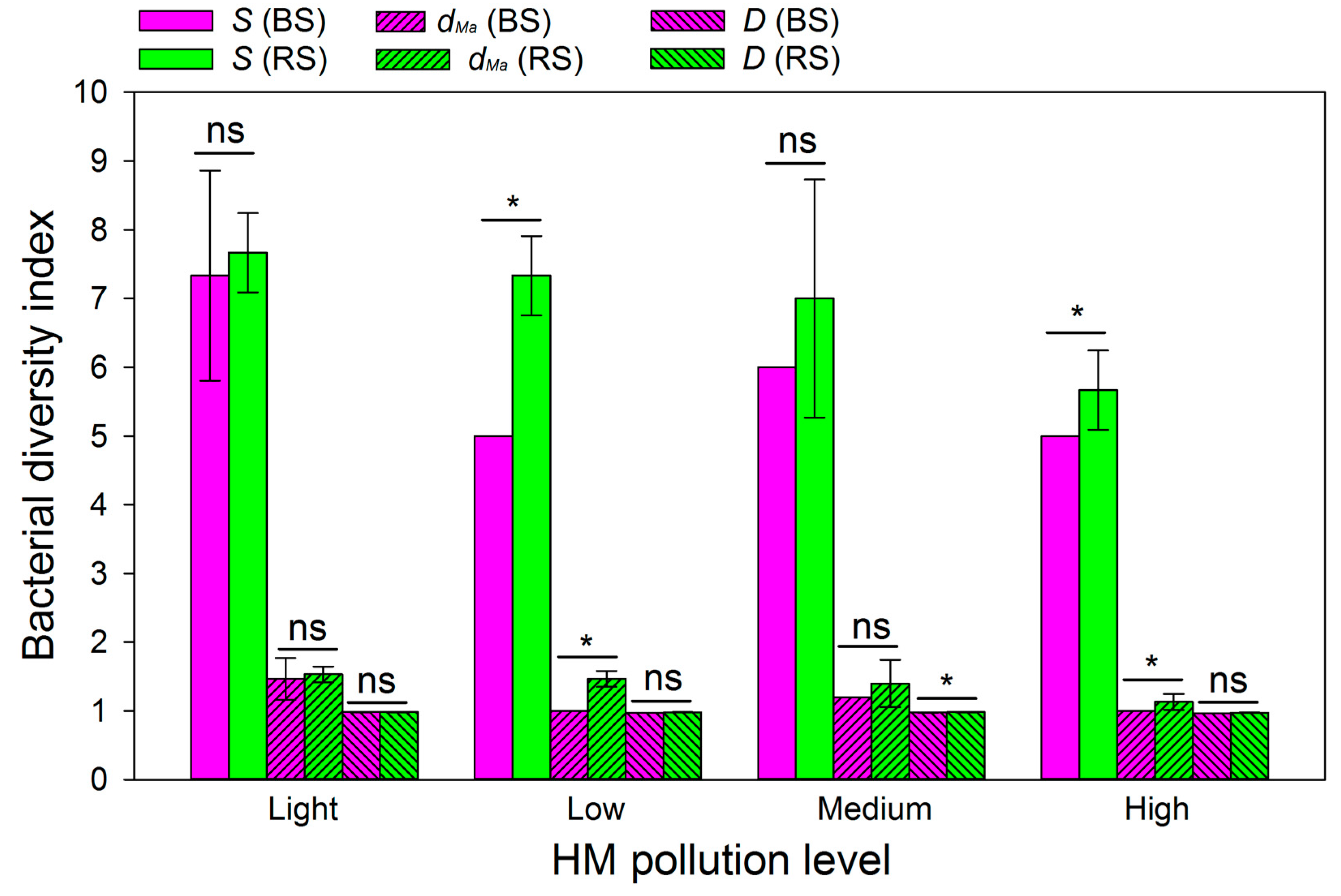
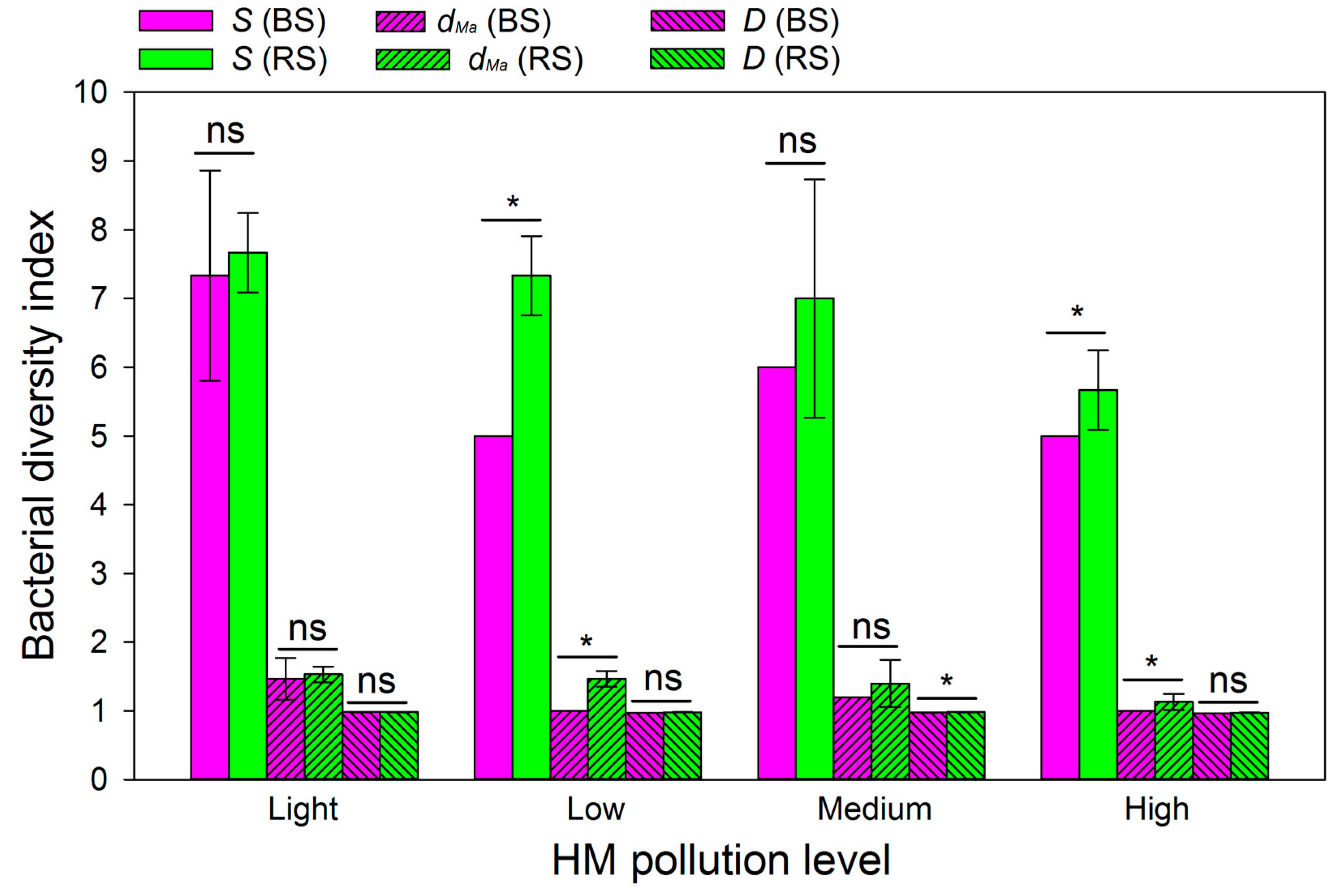
3.7. Bacterial Diversity Index
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arriagada, C.A.; Herrera, M.A.; Ocampo, J.A. Beneficial effect of saprobe and arbuscular mycorrhizal fungi on growth of Eucalyptus globulus co-cultured with Glycine max in soil contaminated with heavy metals. J. Environ. Manag. 2007, 84, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, A.; Carmona, F.F.; Bhargava, M.; Srivastava, S. Approaches for enhanced phytoextraction of heavy metals. J. Environ. Manag. 2012, 105, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Solgi, E.; Esmaili-Sari, A.; Riyahi-Bakhtiari, A.; Hadipour, M. Soil contamination of metals in the three industrial estates, Arak, Iran. Bull. Environ. Contam. Toxcol. 2012, 88, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J. Soil processes and the behaviour of metals. In Heavy Metals in Soils; Alloway, B.J., Ed.; Blackie Academic and Professional: London, UK, 1995; pp. 11–37. [Google Scholar]
- Moreira, H.; Marques, A.P.; Rangel, A.O.; Castro, P.M. Heavy metal accumulation in plant species indigenous to a contaminated Portuguese site: Prospects for phytoremediation. Water Air Soil Pollut. 2011, 221, 377–389. [Google Scholar] [CrossRef]
- Li, Z.; Ma, Z.; van der Kuijp, T.J.; Yuan, Z.; Huang, L. A review of soil heavy metal pollution from mines in China: Pollution and health risk assessment. Sci. Total Environ. 2014, 468, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Liao, R.; Ali, A.; Mahar, A.; Guo, D.; Li, R.; Xining, S.; Awasthi, M.K.; Wang, Q.; Zhang, Z. Spatial distribution and risk assessment of heavy metals in soil near a Pb/Zn smelter in Feng County, China. Ecotoxicol. Environ. Saf. 2017, 139, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Gao, X. Integrated assessment of heavy metal pollution in the surface sediments of the Laizhou Bay and the coastal waters of the Zhangzi Island, China: Comparison among typical marine sediment quality indices. PLoS ONE 2014, 9, e94145. [Google Scholar] [CrossRef] [PubMed]
- Järup, L. Hazards of heavy metal contamination. Br. Med. Bull. 2003, 68, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Burns, R.G. Enzyme activity in soil: Location and a possible role in microbial ecology. Soil Biol. Biochem. 1982, 14, 423–427. [Google Scholar] [CrossRef]
- Das, S.K.; Varma, A. Role of enzymes in maintaining soil health. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer-Verlag: Berlin, Germany, 2011; pp. 25–42. [Google Scholar]
- Xian, Y.; Wang, M.; Chen, W. Quantitative assessment on soil enzyme activities of heavy metal contaminated soils with various soil properties. Chemosphere 2015, 139, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Vig, K.; Megharaj, M.; Sethunathan, N.; Naidu, R. Bioavailability and toxicity of cadmium to microorganisms and their activities in soil: A review. Adv. Environ. Res. 2003, 8, 121–135. [Google Scholar] [CrossRef]
- Brookes, P.C. The use of microbial parameters in monitoring soil pollution by heavy metals. Biol. Fertil. Soils 1995, 19, 269–279. [Google Scholar] [CrossRef]
- Liao, M.; Huang, C.Y. Effect of combined pollution by heavy metals on soil enzymatic activities in areas polluted by tailings from Pb-Zn-Ag mine. J. Environ. Sci. 2005, 17, 637–640. [Google Scholar]
- Giller, K.E.; Witter, E.; Mcgrath, S.P. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem. 1998, 30, 1389–1414. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Tabatabai, M.A. Enzyme activities in a limed agricultural soil. Biol. Fertil. Soils 2000, 31, 85–91. [Google Scholar] [CrossRef]
- Versaw, W.K.; Harrison, M.J. A chloroplast phosphate transporter, PHT2;1, influences allocation of phosphate within the plant and phosphate-starvation responses. Plant Cell 2002, 14, 1751–1766. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.X.; Liu, S.Q.; Zhang, D.W.; Feng, S.D. Effects of cadium, zinc and lead on soil enzyme activities. J. Environ. Sci. 2006, 18, 1135–1141. [Google Scholar] [CrossRef]
- Smalla, K.; Wieland, G.; Buchner, A.; Zock, A.; Parzy, J.; Kaiser, S.; Roskot, N.; Heuer, H.; Berg, G. Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: Plant-dependent enrichment and seasonal shifts revealed. Appl. Environ. Microbiol. 2001, 67, 4742–4751. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Phillips, R.P.; Fahey, T.J. Tree species and mycorrhizal associations influence the magnitude of rhizosphere effects. Ecology 2006, 87, 1302–1313. [Google Scholar] [CrossRef]
- Liu, D.; Fang, S.; Tian, Y.; Chang, S.X. Nitrogen transformations in the rhizosphere of different tree types in a seasonally flooded soil. Plant Soil Environ. 2014, 60, 249–254. [Google Scholar]
- Yang, Y.; Liang, Y.; Ghosh, A.; Song, Y.; Chen, H.; Tang, M. Assessment of arbuscular mycorrhizal fungi status and heavy metal accumulation characteristics of tree species in a lead-zinc mine area: Potential applications for phytoremediation. Environ. Sci. Pollut. Res. 2015, 22, 13179–13193. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.H.; Nan, Z.R.; Su, J.Q.; Wang, N. Heavy metal accumulation by poplar in calcareous soil with various degrees of multi-metal contamination: Implications for phytoextraction and phytostabilization. Environ. Sci. Pollut. Res. 2013, 20, 7194–7203. [Google Scholar] [CrossRef] [PubMed]
- Moser, A.; Rötzer, T.; Pauleit, S.; Pretzsch, H. The urban environment can modify drought stress of small-leaved lime (Tilia cordata Mill.) and black locust (Robinia pseudoacacia L.). Forests 2016, 7, 71. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Tang, M.; Chen, H.; Ban, Y.H.; Zhang, H.H. Microbial community structure in the rhizosphere of Sophora viciifolia grown at a lead and zinc mine of northwest China. Sci. Total Environ. 2012, 435, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.J.; Tian, M.M.; Wu, S.L. Mineral resources exploitation and sustainable development of Fengxian County in Shaanxi Province. Miner. Res. Geol. 2004, 18, 470–475. [Google Scholar]
- Ban, Y.H.; Tang, M.; Chen, H.; Xu, Z.Y.; Zhang, H.H.; Yang, Y.R. The response of dark septate endophytes (DSE) to heavy metals in pure culture. PLoS ONE 2012, 7, e47968. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Song, Y.; Scheller, V.H.; Ghosh, A.; Ban, Y.; Chen, H.; Tang, M. Community structure of arbuscular mycorrhizal fungi associated with Robinia pseudoacacia in uncontaminated and heavy metal contaminated soils. Soil Biol. Biochem. 2015, 86, 146–158. [Google Scholar] [CrossRef]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The combined effects of arbuscular mycorrhizal fungi (AMF) and lead (Pb) stress on Pb accumulation, plant growth parameters, photosynthesis, and antioxidant enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis; Page, A.L., Ed.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1982; pp. 539–579. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Total nitrogen. In Methods of Soil Analysis; Page, A.L., Ed.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice-hall Inc.: Englewood Cliffs, NJ, USA, 1958; pp. 111–133. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of soil analysis. In Chemical and Microbiological Properties; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Ladd, J.N.; Butler, J.H.A. Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol. Biochem. 1972, 4, 19–30. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jackson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ban, Y.; Li, Z.; Chen, H.; Yang, R.; Tang, M. Arbuscular mycorrhizal fungi play a role in protecting roots of Sophora viciifolia Hance. From Pb damage associated with increased phytochelatin synthase gene expression. Environ. Sci. Pollut. Res. 2014, 21, 12671–12683. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Olaniran, A.O.; Balgobind, A.; Pillay, B. Bioavailability of heavy metals in soil: Impact on microbial biodegradation of organic compounds and possible improvement strategies. Int. J. Mol. Sci. 2013, 14, 10197–10228. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, C.; Giraudoux, P.; Cœurdassier, M.; Douay, F.; Raoul, F.; Pruvot, C.; Waterlot, C.; De Vaufleury, A.; Scheifler, R. Spatial distribution of metals in smelter-impacted soils of woody habitats: Influence of landscape and soil properties, and risk for wildlife. Chemosphere 2010, 81, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Bonanno, G.; Lo Giudice, R. Heavy metal bioaccumulation by the organs of Phragmites australis (common reed) and their potential use as contamination indicators. Ecol. Indic. 2010, 10, 639–645. [Google Scholar] [CrossRef]
- Rodriguez, E.; da Conceição Santos, M.; Azevedo, R.; Correia, C.; Moutinho-Pereira, J.; de Oliveira, J.M.P.F.; Dias, M.C. Photosynthesis light-independent reactions are sensitive biomarkers to monitor lead phytotoxicity in a Pb-tolerant Pisum sativum cultivar. Environ. Sci. Pollut. Res. 2015, 22, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Zhou, Q.; Ma, L.Q. Availability and assessment of fixing additives for the in situ remediation of heavy metal contaminated soils: A review. Environ. Monit. Assess. 2006, 116, 513–528. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Wang, S.; Sun, L.N.; Wang, H. Metabolic profiling of root exudates from two ecotypes of Sedum alfredii treated with Pb based on GC-MS. Sci. Rep. 2017, 7, 39878. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, E.; Del Re, A.A.M.; Rao, M.A.; Gianfreda, L. Development and validation of numerical indexes integrating enzyme activities of soils. Soil Biol. Biochem. 2006, 38, 1673–1681. [Google Scholar] [CrossRef]
- De Santiago-Martín, A.; Cheviron, N.; Quintana, J.R.; González, C.; Lafuente, A.L.; Mougin, C. Metal contamination disturbs biochemical and microbial properties of calcareous agricultural soils of the Mediterranean area. Arch. Environ. Contam. Toxicol. 2013, 64, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Garcıa-Gil, J.C.; Plaza, C.; Soler-Rovira, P.; Polo, A. Long-term effects of municipal solid waste compost application on soil enzyme activities and microbial biomass. Soil Biol. Biochem. 2000, 32, 1907–1913. [Google Scholar] [CrossRef]
- Hussein, K.A.; Joo, J.H. Heavy metal resistance of bacteria and its impact on the production of antioxidant enzymes. Afr. J. Microbiol. Res. 2013, 7, 2288–2296. [Google Scholar]
- Xiao, S.; You, H.; You, W.; Liu, J.; Cai, C.; Wu, J.; Ji, Z.; Zhan, S.; Hu, Z.; Zhang, Z.; et al. Rhizosphere and bulk soil enzyme activities in a Nothotsuga longibracteata forest in the Tianbaoyan National Nature Reserve, Fujian Province, China. J. For. Res. 2017, 28, 521–528. [Google Scholar] [CrossRef]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; Zhou, W. Responses of extracellular enzyme activities and microbial community in both the rhizosphere and bulk soil to long-term fertilization practices in a fluvo-aquic soil. Geoderma 2012, 173, 330–338. [Google Scholar] [CrossRef]
- Piotrowska-Seget, Z.; Cycoń, M.; Kozdroj, J. Metal-tolerant bacteria occurring in heavily polluted soil and mine spoil. Appl. Soil Ecol. 2005, 28, 237–246. [Google Scholar] [CrossRef]
- Mendez, M.O.; Neilson, J.W.; Maier, R.M. Characterization of a bacterial community in an abandoned semiarid lead-zinc mine tailing site. Appl. Environ. Microbiol. 2008, 74, 3899–3907. [Google Scholar] [CrossRef] [PubMed]
- Sandaa, R.A.; Torsvik, V.; Enger, Ø. Influence of long-term heavy-metal contamination on microbial communities in soil. Soil Biol. Biochem. 2001, 33, 287–295. [Google Scholar] [CrossRef]
- Gremion, F.; Chatzinotas, A.; Harms, H. Comparative 16S rRNA and 16S rRNA sequence analysis indicates that Actinobacteria might be a dominant part of the metabolically active bacteria in heavy metal-contaminated bulk and rhizosphere soil. Environ. Microbiol. 2003, 5, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhang, J.; Li, Q.; Han, T.; Xie, J.; Hu, Y.; Chai, L. Phylogenetic analysis of bacterial community composition in sediment contaminated with multiple heavy metals from the Xiangjiang River in China. Mar. Pollut. Bull. 2013, 70, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Balderas, C.D.; Cochet, N.; Bert, V.; Tarnaud, E.; Sarde, C.O. 16S rRNA analysis of bacterial communities associated with the hyper accumulator Arabidopsis halleri grown on a Zn and Cd polluted soil. Eur. J. Soil Biol. 2014, 60, 16–23. [Google Scholar] [CrossRef]
- Kuffner, M.; Puschenreiter, M.; Wieshammer, G.; Gorfer, M.; Sessitsch, A. Rhizosphere bacteria affect growth and metal uptake of heavy metal accumulating willows. Plant Soil 2008, 304, 35–44. [Google Scholar] [CrossRef]
- Paterson, E.; Hall, J.M.; Rattray, E.A.S.; Griffiths, B.S.; Ritz, K.; Killham, K. Effect of elevated CO2 on rhizosphere carbon flow and soil microbial processes. Glob. Chang. Biol. 1997, 3, 363–377. [Google Scholar] [CrossRef]
- Khan, S.; Hesham, A.E.L.; Qiao, M.; Rehman, S.; He, J.Z. Effects of Cd and Pb on soil microbial community structure and activities. Environ. Sci. Pollut. Res. 2010, 17, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Ali, S.; Zhang, H.; Ouyang, Y.; Qiu, B.; Wu, F.; Zhang, G. The influence of pH and organic matter content in paddy soil on heavy metal availability and their uptake by rice plants. Environ. Pollut. 2011, 159, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Bending, G.D.; Turner, M.K.; Jones, J.E. Interactions between crop residue and soil organic matter quality and the functional diversity of soil microbial communities. Soil Biol. Biochem. 2002, 34, 1073–1082. [Google Scholar] [CrossRef]
- Kim, J.M.; Roh, A.S.; Choi, S.C.; Kim, E.J.; Choi, M.T.; Ahn, B.K.; Kim, S.K.; Lee, Y.H.; Joa, J.H.; Kang, S.S.; et al. Soil pH and electrical conductivity are key edaphic factors shaping bacterial communities of greenhouse soils in Korea. J. Microbiol. 2016, 54, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Bartram, A.K.; Jiang, X.; Lynch, M.D.; Masella, A.P.; Nicol, G.W.; Dushoff, J.; Neufeld, J.D. Exploring links between pH and bacterial community composition in soils from the Craibstone Experimental Farm. FEMS Microbiol. Ecol. 2014, 87, 403–415. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Soil Type | HM Pollution Level | |||
|---|---|---|---|---|---|
| Low | Light | Medium | High | ||
| pH | BS | 8.47 ± 0.19 ** | 8.62 ± 0.14 ** | 8.35 ± 0.32 ** | 7.99 ± 0.41 * |
| RS | 7.51 ± 0.22 | 7.58 ± 0.33 | 7.47 ± 0.06 | 7.33 ± 0.27 | |
| EC | BS | 0.62 ± 0.05 ns | 0.60 ± 0.05 ns | 0.71 ± 0.02 ** | 0.74 ± 0.04 * |
| RS | 0.69 ± 0.02 | 0.66 ± 0.03 | 0.80 ± 0.03 | 0.82 ± 0.04 | |
| SOM | BS | 11.2 ± 1.78 ns | 12.3 ± 1.51 ns | 8.76 ± 1.46 ** | 5.73 ± 0.75 * |
| RS | 13.7 ± 1.33 | 12.1 ± 1.42 | 12.5 ± 0.65 | 7.90 ± 1.29 | |
| SOC | BS | 13.1 ± 2.41 ns | 12.0 ± 2.00 * | 8.11 ± 0.61 * | 6.75 ± 1.51 ** |
| RS | 15.3 ± 2.58 | 14.8 ± 0.94 | 12.2 ± 2.82 | 12.0 ± 0.98 | |
| TN | BS | 1.03 ± 0.15 ns | 0.95 ± 0.22 * | 0.99 ± 0.11 ns | 0.72 ± 0.04 ns |
| RS | 1.34 ± 0.21 | 1.30 ± 0.15 | 1.30 ± 0.26 | 0.83 ± 0.17 | |
| AN | BS | 14.9 ± 1.74 ns | 14.7 ± 1.94 ** | 12.2 ± 2.78 * | 9.04 ± 1.21 * |
| RS | 16.9 ± 2.35 | 20.3 ± 2.29 | 18.0 ± 1.91 | 11.7 ± 1.55 | |
| TP | BS | 0.66 ± 0.15 ns | 0.84 ± 0.10 ns | 0.78 ± 0.13 * | 0.54 ± 0.10 ns |
| RS | 0.56 ± 0.07 | 0.67 ± 0.13 | 0.52 ± 0.11 | 0.42 ± 0.07 | |
| AP | BS | 9.01 ± 2.13 ** | 8.51 ± 1.48 * | 7.98 ± 1.34 ** | 4.89 ± 1.17 * |
| RS | 13.7 ± 1.03 | 12.3 ± 2.18 | 12.5 ± 1.62 | 7.19 ± 0.73 | |
| TPb | BS | 223 ± 51.5 ns | 922 ± 308 ns | 2405 ± 237 ** | 3396 ± 976 ns |
| RS | 194 ± 44.0 | 751 ± 101 | 1579 ± 268 | 2687 ± 278 | |
| DPb | BS | 10.2 ± 1.83 ns | 46.7 ± 6.39 ns | 152 ± 14.7 ** | 293 ± 70.7 ** |
| RS | 7.82 ± 2.02 | 38.3 ± 10.2 | 73.9 ± 17.0 | 132 ± 15.8 | |
| TZn | BS | 88.4 ± 23.3 ns | 195 ± 6.06 * | 594 ± 116 * | 416 ± 88.8 ns |
| RS | 98.3 ± 23.7 | 147 ± 27.8 | 409 ± 32.8 | 329 ± 77.6 | |
| DZn | BS | 8.94 ± 1.23 ns | 24.9 ± 8.43 ns | 67.5 ± 7.49 ns | 64.8 ± 10.3 ns |
| RS | 12.2 ± 3.90 | 20.4 ± 2.20 | 75.6 ± 4.59 | 60.0 ± 14.0 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Dong, M.; Cao, Y.; Wang, J.; Tang, M.; Ban, Y. Comparisons of Soil Properties, Enzyme Activities and Microbial Communities in Heavy Metal Contaminated Bulk and Rhizosphere Soils of Robinia pseudoacacia L. in the Northern Foot of Qinling Mountain. Forests 2017, 8, 430. https://doi.org/10.3390/f8110430
Yang Y, Dong M, Cao Y, Wang J, Tang M, Ban Y. Comparisons of Soil Properties, Enzyme Activities and Microbial Communities in Heavy Metal Contaminated Bulk and Rhizosphere Soils of Robinia pseudoacacia L. in the Northern Foot of Qinling Mountain. Forests. 2017; 8(11):430. https://doi.org/10.3390/f8110430
Chicago/Turabian StyleYang, Yurong, Miao Dong, Yaping Cao, Jinlong Wang, Ming Tang, and Yihui Ban. 2017. "Comparisons of Soil Properties, Enzyme Activities and Microbial Communities in Heavy Metal Contaminated Bulk and Rhizosphere Soils of Robinia pseudoacacia L. in the Northern Foot of Qinling Mountain" Forests 8, no. 11: 430. https://doi.org/10.3390/f8110430