Physical Conditions Regulate the Fungal to Bacterial Ratios of a Tropical Suspended Soil
by
 ,
,
Julian Donald
1,*, 
Sam Bonnett
1,
Michael Cutler
2,
Noreen Majalap
3,
Pete Maxfield
1 and
M. D. Farnon Ellwood
1 1
Centre for Research in Biosciences, University of the West of England, Bristol BS16 1QY, UK
2
The Eden Project, Bodelva, Par, Cornwall PL24 2SG, UK
3
Phytochemistry Unit, Forest Research Centre, Jalan Sepilok, Sepilok, 90715 Sandakan, Sabah, Malaysia
*
Author to whom correspondence should be addressed.
Forests 2017, 8(12), 474; https://doi.org/10.3390/f8120474
Submission received: 20 August 2017
/
Revised: 3 November 2017
/
Accepted: 28 November 2017
/
Published: 2 December 2017
(This article belongs to the Special Issue The Role of Fungi in Tropical Forest Systems)
Abstract
:As a source of ‘suspended soils’, epiphytes contribute large amounts of organic matter to the canopy of tropical rain forests. Microbes associated with epiphytes are responsible for much of the nutrient cycling taking place in rain forest canopies. However, soils suspended far above the ground in living organisms differ from soil on the forest floor, and traditional predictors of soil microbial community composition and functioning (nutrient availability and the activity of soil organisms) are likely to be less important. We conducted an experiment in the rain forest biome at the Eden Project in the U.K. to explore how biotic and abiotic conditions determine microbial community composition and functioning in a suspended soil. To simulate their natural epiphytic lifestyle, bird’s nest ferns (Asplenium nidus) were placed on a custom-built canopy platform suspended 8 m above the ground. Ammonium nitrate and earthworm treatments were applied to ferns in a factorial design. Extracellular enzyme activity and Phospholipid Fatty Acid (PLFA) profiles were determined at zero, three and six months. We observed no significant differences in either enzyme activity or PLFA profiles between any of the treatments. Instead, we observed decreases in β-glucosidase and N-acetyl-glucosaminidase activity, and an increase in phenol oxidase activity across all treatments and controls over time. An increase in the relative abundance of fungi during the experiment meant that the microbial communities in the Eden Project ferns after six months were comparable with ferns sampled from primary tropical rain forest in Borneo.
1. Introduction
The canopy of tropical rain forests provides a unique habitat for a range of organisms in spite of characteristic extremes in climate and nutrient availability [1,2,3,4]. Suspended soils, the accumulation of organic matter supporting microorganisms, invertebrates and plants, are important components of the canopy ecosystem [5]. The inhabitants of canopy soils depend on the tight cycling of limited resources, facilitated by a decomposer community adapted to the extremes in climate that characterise canopy habitats [6,7,8]. In terrestrial soils, microorganisms are known to drive decomposition [9], yet little is known about the abundance of microorganisms or their role in canopy soils [10,11,12,13]. Recent advances in canopy access have improved our understanding of tropical rain forests but information on canopy microbial communities is lacking [14,15]. The composition and functioning of soil microbial communities is known to be influenced by multiple factors, namely nutrient inputs to the soil [16,17,18,19]; the activity of the associated invertebrate community, notably earthworms [20,21,22]; soil temperature, humidity and pH [23,24,25,26]. Given the unique conditions under which canopy soils form, the relative importance of these factors is likely to be markedly different from other soils.
Soil fungi and bacteria drive decomposition by producing extracellular enzymes in response to their nutrient demands [27]. These enzymes convert polymeric compounds contained within organic matter into smaller molecules that can be taken up by organisms [28]. Hydrolytic enzymes such as β-glucosidase and N-acetyl-glucosaminidase generally break down labile carbon stored in cellulose and chitin, whereas oxidative enzymes such as phenol oxidase decompose recalcitrant lignin [29,30]. Suspended soils are high in organic matter and thus organically-bound nitrogen, but it is not known whether canopy microorganisms can access this form of nitrogen [31]. However, forest canopies are also experiencing increasing deposition of inorganic nitrogen from the atmosphere [32]. This inorganic nitrogen has been shown to modify soil microbial activity in certain tropical regions and contribute significantly to epiphyte nitrogen balances [7,33]. It is thought that the addition of inorganic nitrogen from the atmosphere stimulates decomposition in suspended soils that are nutrient limited by providing microorganisms with the resources required to increase extracellular enzyme production [27]. Another factor known to influence extracellular enzyme production is the presence of organisms such as earthworms, which increase the availability of nutrients to microbes through soil mixing or bioturbation [22]. However, the unique nature of suspended soils associated with epiphytes could mean that these traditional predictors of soil community composition and activity are not as important as other predictors in the canopy.
The epiphytic bird’s nest fern (Asplenium spp.) [34] depends on its associated canopy soil for survival. In the context of natural microcosm studies [35], the fern represents an ideal model system for exploring patterns and processes associated with canopy soil microbial communities. The bird’s nest fern’s ubiquity, its discrete nature, and its resilience to experimental manipulation have facilitated initial studies of nutrient cycling between the canopy and the forest floor [36]. Their role as a harbour of high invertebrate abundance and diversity (including earthworms) [37,38], has been used to elucidate complex species assembly rules [39,40,41]. Bird’s nest ferns thus provide an ideal vehicle to assess the influence of nutrient balances and invertebrate activity on microbially-mediated decomposition in suspended soils.
This study examines the potential of bird’s nest fern soil to support microbial communities whose activities are adapted to the canopy environment. We performed an experiment using epiphytic ferns in the tropical biome of the Eden Project in Cornwall, U.K. [42] to test the relative importance of traditional predictors of soil microbial community composition and functioning. Using the ferns as natural microcosms in a model rain forest provided biologically realistic conditions while giving us more control over the physical environment. This study aims to test the impact of the following on microbial community structure and functioning: (1) the addition of inorganic nitrogen; (2) the activity of earthworms; and (3) suspension within the canopy, to simulate the physical conditions (exposure to the air, regular wetting and drying, throughflow of water) experienced by epiphytic soils. We validated our results by comparing the microbial communities of the Eden Project ferns in a large greenhouse environment in the U.K. with bird’s nest ferns collected from primary tropical rain forest in Borneo.
2. Materials and Methods
2.1. Eden Project Experiments
Data collection took place at the Eden Project in Cornwall, U.K. (50°36’ N, 4°74’ W). A total of 20 Asplenium ferns were procured from Western Wholesale Plants Nursery in Weston-super-Mare, U.K. Soil was removed from the plant roots, and was replaced with organic matter mulch made on site at the Eden Project with a mean pH of 8.4. Mean soil nitrogen concentration in the form of NO3-N was 36.1 mg kg−1, and in the form of NH4-N was 20.3 mg kg−1. Before the experiment, ferns were housed in a nursery for a period of four months under mean daily temperatures of 21.8 ± 0.6 °C, with a relative humidity of 88.3 ± 0.16% [43]. These ferns were kept in solid trays meaning that their soil was enclosed between each daily watering event. Baseline soil samples were taken from each fern at this point (n = 20). Approximately 10% of the total soil volume was removed during each sampling event, with care taken not to disturb the remaining soil body.
Ferns were then transferred from the nursery to the rain forest biome, which experiences controlled levels of temperature and humidity (daily temperatures 25.6 ± 0.03 °C and humidity 93.8 ± 0.05%). The host trees used for the experiment were Hopea odorata, a common species of rain forest tree from the same family (Dipterocarpaceae) as our host trees in Borneo. These trees were originally sourced from Danum Valley, the location of our field site [44].
In order to examine the impact of bioturbation and nutrient inputs in shaping canopy soil microbial communities, the ferns were experimentally manipulated under a factorial design, whereby:
- 5 ferns were inoculated with 5 individuals of the earthworm Eisenia fetida;
- 5 ferns were dosed with an ammonium nitrate solution, representing 365.7 mg N per kg dry soil;
- 5 ferns received 5 earthworms and ammonium nitrate per fern;
- 5 non-manipulated ferns served as controls.
Ten ferns were suspended in two H. odorata trees using platforms installed 8 m from the ground to simulate epiphytic conditions. An irrigation system, incorporated into the design of the platforms [42], delivered 0.5 L of water daily over an interval of five minutes to each of the 20 suspended plants (Figure 1a). Perforated aquatic plant pots containing the ferns allowed for drainage and air drying of the soils over a period of 24 h, simulating the drying and rewetting cycles associated with rain forest canopy epiphytes [45].
Following baseline soil collection, soils were sampled after three months, in September 2015 (n = 20), and after a further three months in December 2015 (n = 20). Of these soil samples, approximately 500 g was kept at ambient temperature, whilst a sub-sample of 10 g was immediately frozen and subsequently freeze-dried. Upon collection of the final samples, the ferns were dissected to confirm the presence of earthworms in all appropriate treatments.
2.2. Collection of Ferns from Danum Valley
The soil associated with 20 bird’s nest ferns was collected from the primary lowland dipterocarp rain forest of the Danum Valley Conservation Area (DVCA), located within the state of Sabah, in Malaysian Borneo. The Danum Valley Field Centre (4°58′ N, 117°42′ E, altitude ~170 m) experiences a wet equatorial climate, not strongly seasonal, with a mean rainfall of 2785 mm per year. Daily temperatures are, on average, 26.7 °C, with mean highs of 31.0 °C and lows of 22.5 °C [44].
Asplenium ferns (Figure 1b) were collected along the existing trail network surrounding the field centre. One hectare of forest, split across five 2000 m2 sites was surveyed, and all ferns present below 8 m height were recorded. Details of estimated leaf rosette diameter allowed for a rough estimation of the age of each fern, and height determined their accessibility for sampling. Of those with a diameter greater than 100 cm, and below 8 m in height, a random subsample of 20 ferns across the five sites was collected using a combination of ladders and single rope access techniques. Ferns were removed from their host tree, with care taken to collect as much soil associated with the roots as possible.
The soil associated with each fern was removed and homogenised using a 1 cm mesh sieve. The soil was put into Ziplock bags before being frozen and transported to the Forest Research Centre at Sepilok where they were placed into an Alpha 1–4 LSC freeze drier (Christ, Osterode am Harz, Germany) for 24 h. The resulting freeze-dried samples were stored at −20 °C before being transported back to the U.K. for analysis.
2.3. Hydrolase Enzyme Activity
For detailed information on soil enzyme methodologies, see [46]. The potential enzyme activity of the experimental fern microbial communities at the Eden Project was tested using fresh soil samples (n = 60). Following their collection at the Eden Project, soils were put in Ziplock bags and transported back to the laboratory where they were stored at room temperature for one week, and were opened regularly to avoid the accumulation of carbon dioxide. Methylumbelliferone (MUF) substrate solutions were prepared for the enzymes β-glucosidase and N-acetyl-glucosaminidase. Preliminary data demonstrated that a concentration of 100 µm saturated the enzyme reaction allowing for an estimate of Vmax, the maximum enzyme capacity of the soil. Each soil solution (1:5 wet soil to deionised water) was pipetted into a 96 well plate, with three wells for each soil sample. One contained a soil blank with deionised water, one with the MUF substrate, and one with the MUF standard. The reaction was left active for one hour before 1 M sodium hydroxide was added to terminate it. The plate was then transferred to a Fluostar Optima Fluorometer plate reader (BMG Labtech, Ortenberg, Germany) to record fluorescence. A mean of three sub-samples was calculated for each sample and the data were converted to give β-glucosidase activity (µmol MUF g−1 hour−1).
2.4. Oxidative Enzyme Activity
Potential phenol oxidase activity was determined by pipetting 0.75 mL of soil solution (1:5 wet soil to deionised water) into each of two Eppendorfs (Merck KGaA, Darmstadt, Germany). Deionised water was added to one, whilst a 10 mM solution of L-3,4-dihydroxyphenylalanine (L-DOPA) was added to the other. These were incubated at room temperature for one hour before being centrifuged. The supernatant was pipetted onto a clear microplate and transferred to a Fluostar Optima Fluorometer plate reader (BMG Labtech, Ortenberg, Germany) to measure absorbance at 460 nm. Phenol oxidase activity was calculated by comparing the L-DOPA solution with that of the blank. A mean of three sub-samples was calculated for each sample, and the data were then converted to give phenol oxidase activity based on the production of 2-carboxy-2,3-dihydroindole-5,6-quinone, referred to in the equation as diqc (µmol dicq g−1 hour−1).
2.5. Phospholipid Fatty Acid Analysis
For detailed information on the Phospholipid Fatty Acid (PLFA) method, see [46]. The freeze-dried soils collected from Danum Valley (n = 20) and the Eden Project (n = 60) were analysed for their phospholipid profiles. Each sample was ground to a fine power, with 500 mg added to 2.8 mL of a 2:0.8 methanol:water solution and homogenised. An amount of 1.35 mL of chloroform was added and the sample was centrifuged. The supernatant solution was re-extracted with Bligh-Dyer solution (2:0.8:1 ratio of methanol:water:chloroform) [47]. The organic and aqueous phases were separated by the addition of water and chloroform before being centrifuged. The bottom (organic) layer was removed and the aqueous layer was re-extracted with chloroform. The chloroform was then evaporated under nitrogen gas and the remaining total lipid extract (TLE) was then stored at −20 °C.
The TLE was further separated by column chromatography using the method described by [48], with the neutral fraction and glycolipids separated to leave the phospholipids. The phospholipids were then derivatised in hydrogen chloride, along with 10 µL of a known C18 alkane (N-octadecane, 99+% Acros Organics, Morris, NJ, USA). The sample was heated at 60 °C in a sealed tube, before the resulting fatty acid methyl esters (FAMEs) were extracted into hexane. This solution was evaporated at 40 °C under nitrogen before the FAMEs were re-dissolved in 30 µL of hexane. An amount of 1 µL of this solution was then analysed using gas chromatography (GC) and gas chromatography-mass spectrometry (GC-MS).
Detailed chromatograms were produced for each sample with the peaks displayed identified using Xcalibur 3.0 (Thermo Scientific, Waltham, MA, USA). These were identified as either bacterial or fungal based on the classification of [49] and fungal:bacterial ratios were calculated for each soil sample. In addition, soils from Danum and the Eden Project were classified using [50], in order to gain a broad indication of the prevalence of different groups of microorganisms (Table 1).
2.6. Statistical Analysis
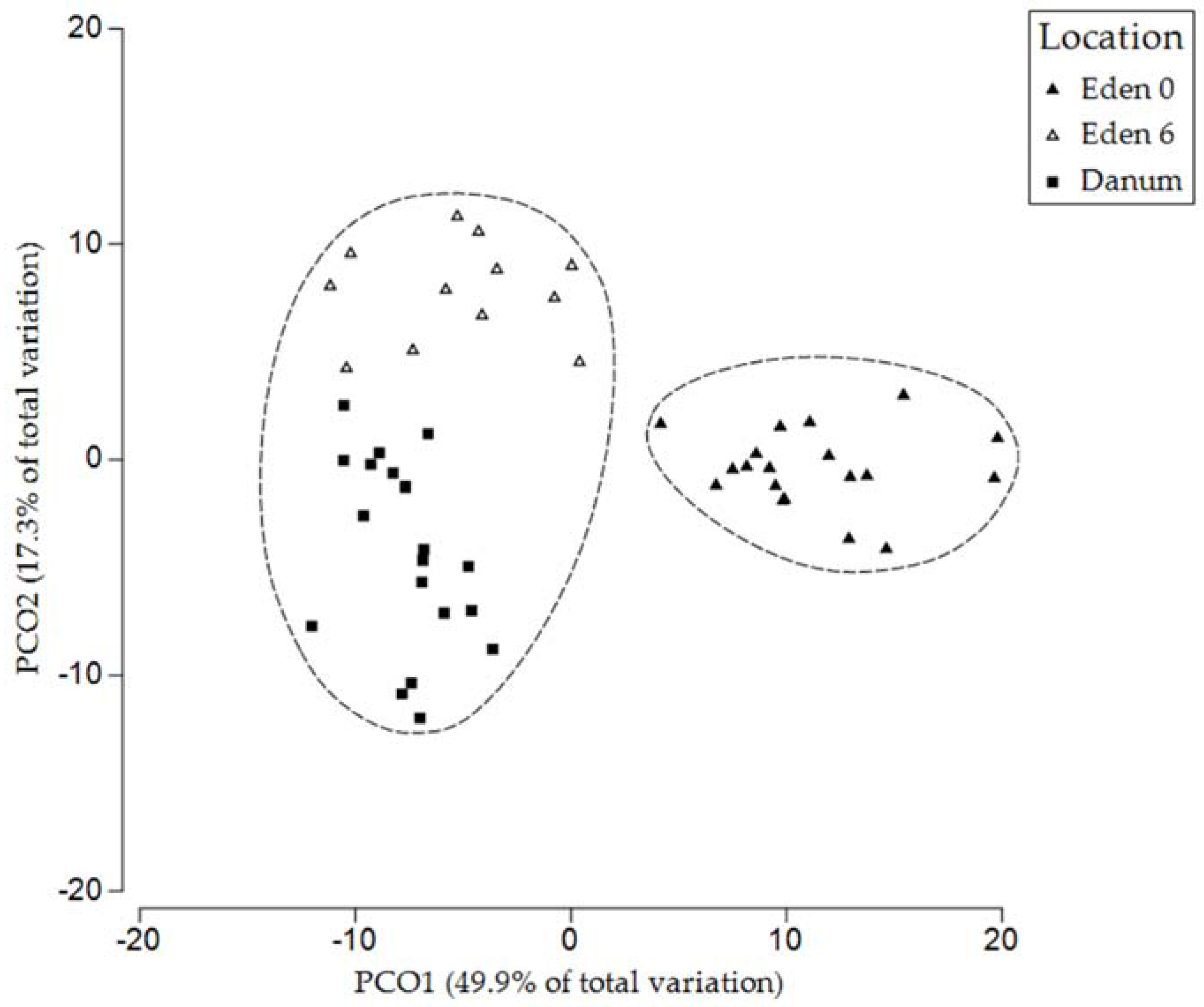
A two-way ANOVA was performed on the Eden Project data, with β-glucosidase activity, N-acetyl-glucosaminidase, phenol oxidase activity, and soil fungal:bacterial ratio as the response variables, and treatment and time as the explanatory variables, including their interaction effects using the R statistical analysis software [51]. The effects of treatment on microbial community composition were tested through multivariate analysis of the PLFA profiles of each fern, using PRIMER version 7.0.13 with PERMANOVA+ [52]. Fern soil microbial community composition was characterised by the relative abundance of PLFAs. The data were square root transformed and Bray–Curtis measures of similarity were calculated across ferns to determine their similarity. The effects of each treatment, and the month of sample collection on microbial community composition, were tested using random permutations of the dataset. A PERMANOVA was used to determine whether these factors exerted significant effects on community composition. We used a Principal Coordinate Ordination (PCO) to compare the microbial communities of ferns from the Eden Project with those from Danum Valley. A cluster analysis based on group average was then used to test for similarity between the three sample groups, namely the Eden Project ferns pre- and post-experimental treatment (0 and 6 months, respectively) and ferns from Danum Valley.
3. Results
3.1. Eden Project Enzyme Activity and Fungal:Bacterial Ratio
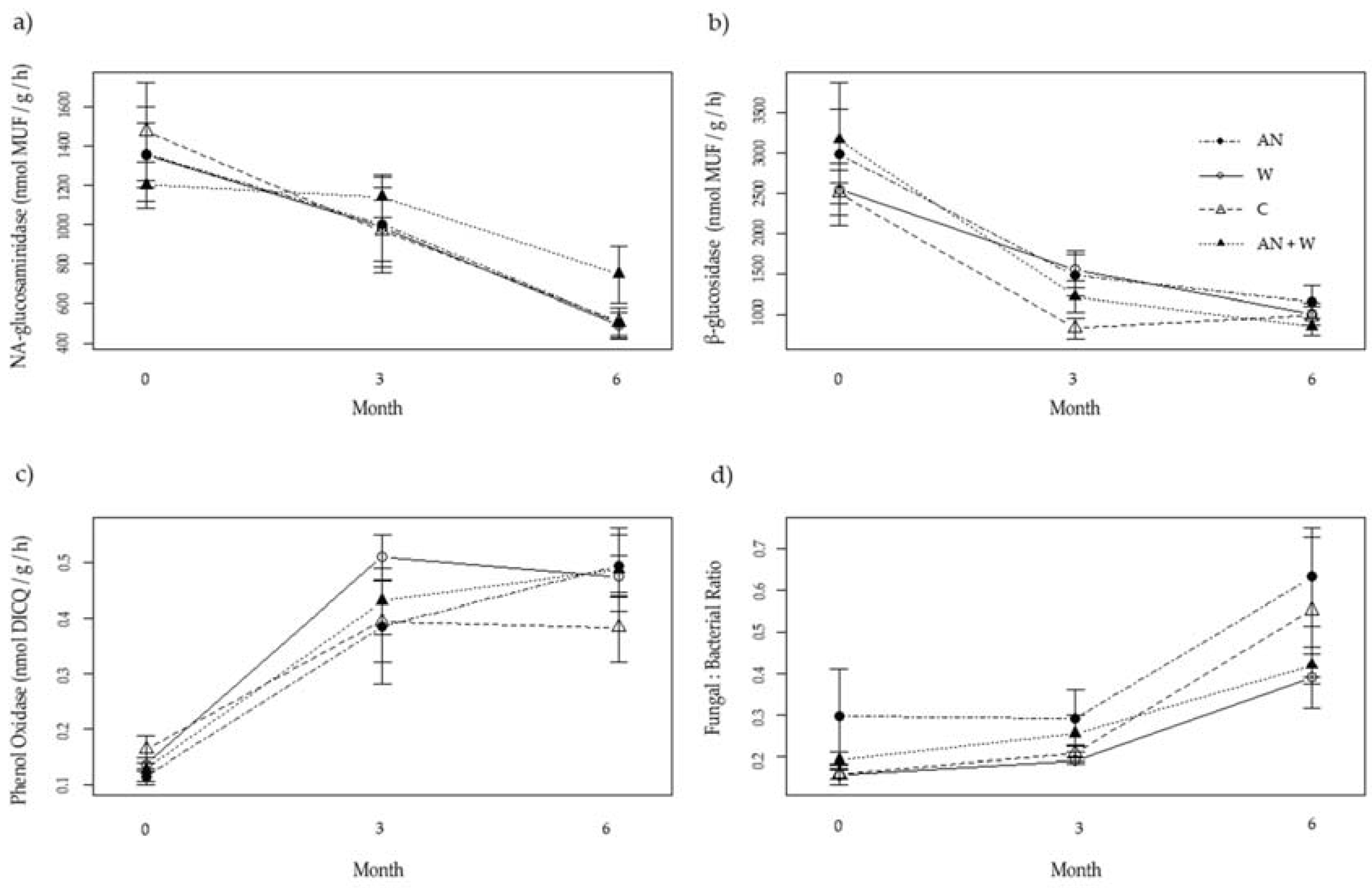
For all three enzymes, the treatments did not result in significant differences in their activity when compared with untreated control ferns after 3 or 6 months. N-acetyl-glucosaminidase and β-glucosidase activity decreased over time, while phenol oxidase activity increased over time. Treatment and its interaction with time had no significant effect across all enzymes (Figure 2; Table 2).
Fungal:bacterial ratios started to increase after 3 months. Treatment and its interaction with time had no significant effect across all sampled enzyme assays and fungal: bacterial ratios (Figure 2; Table 2). Time had a significant effect on PLFA composition (Pseudo-F2,38 = 376.1, p = 0.001), whereas neither treatment (Pseudo-F3,38 = 1.304, p = 0.184), nor the interaction (Pseudo-F6,38 = 0.8892, p = 0.697) were significant.
3.2. Fern Soil Microbial Community Analysis
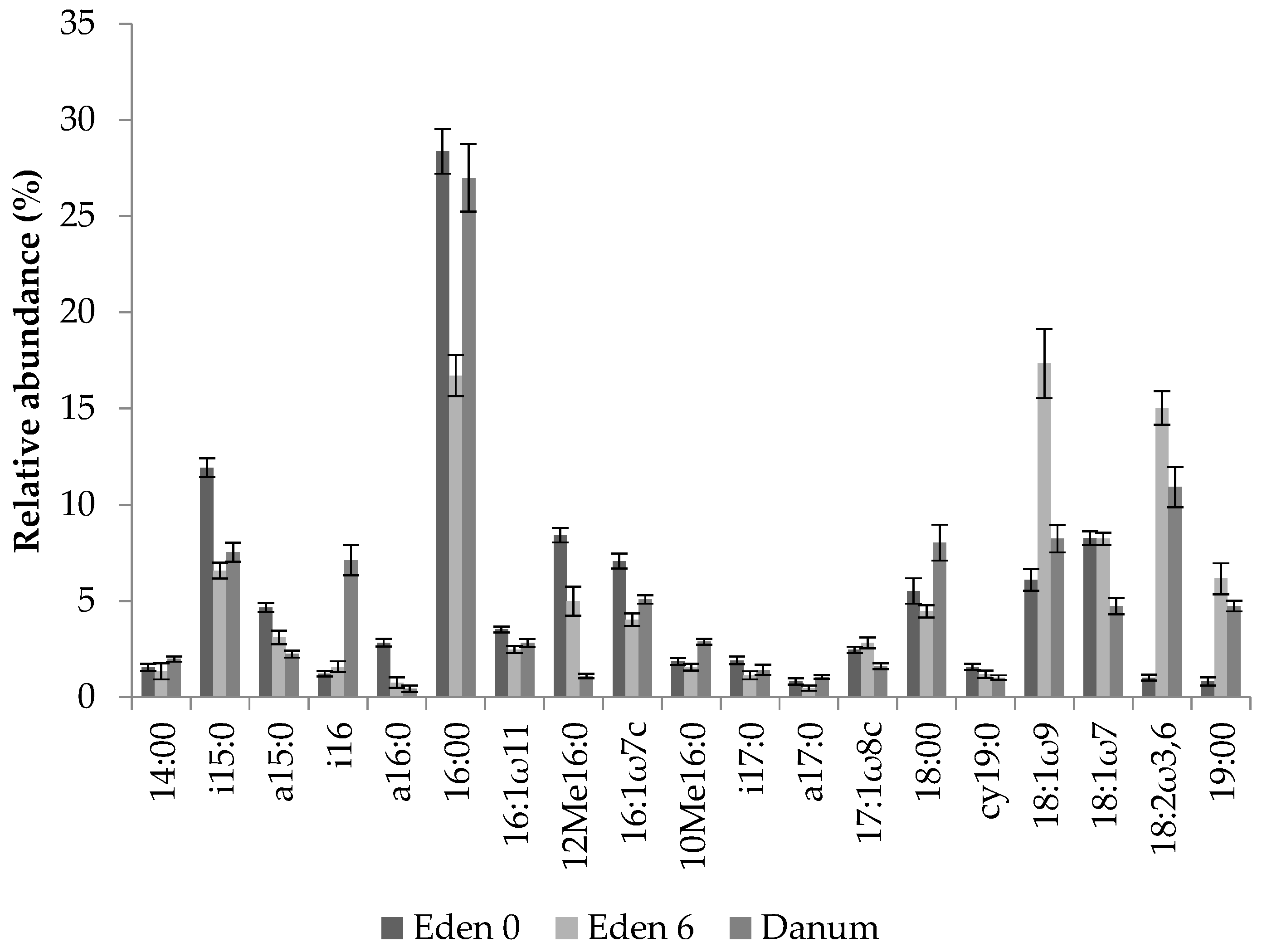
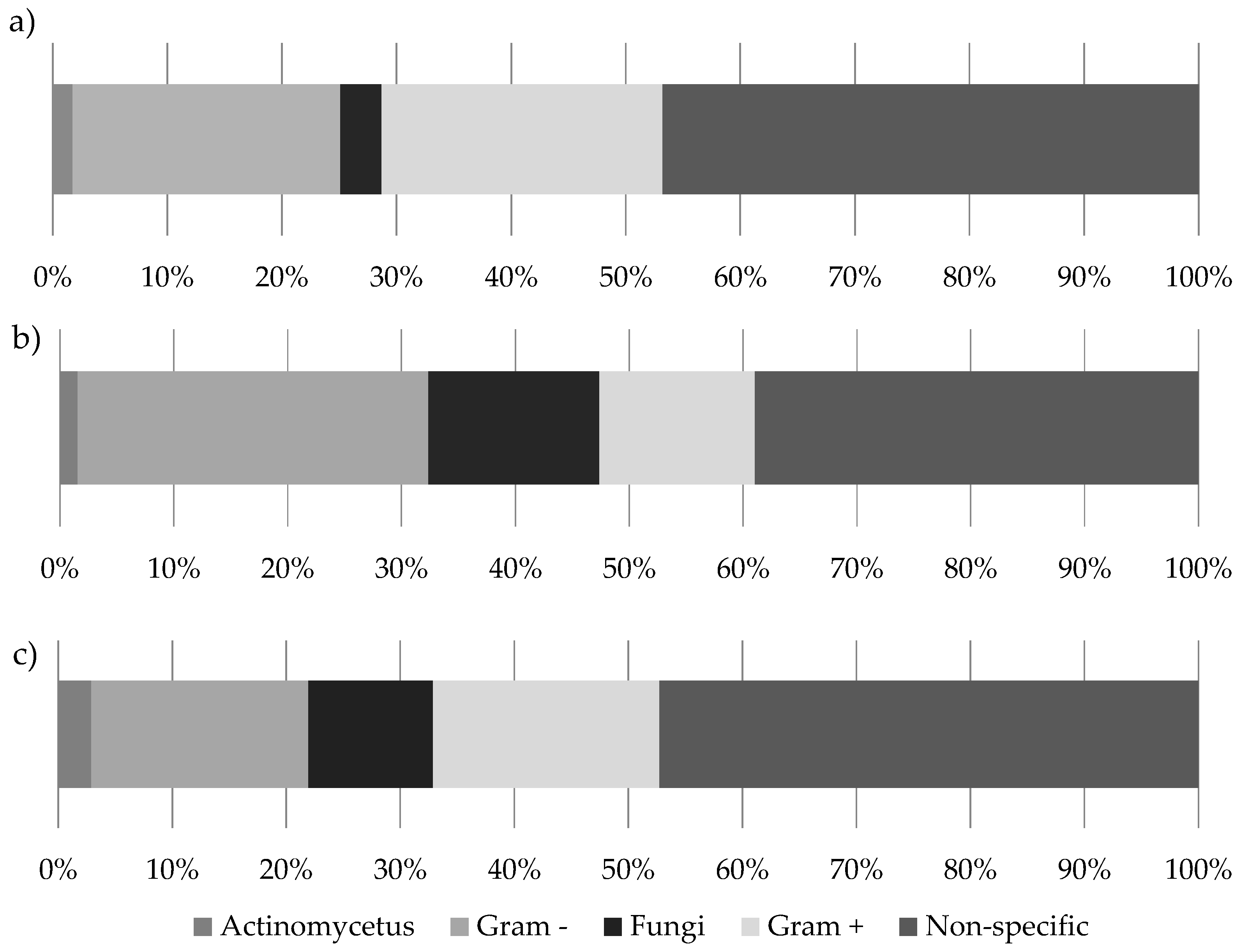
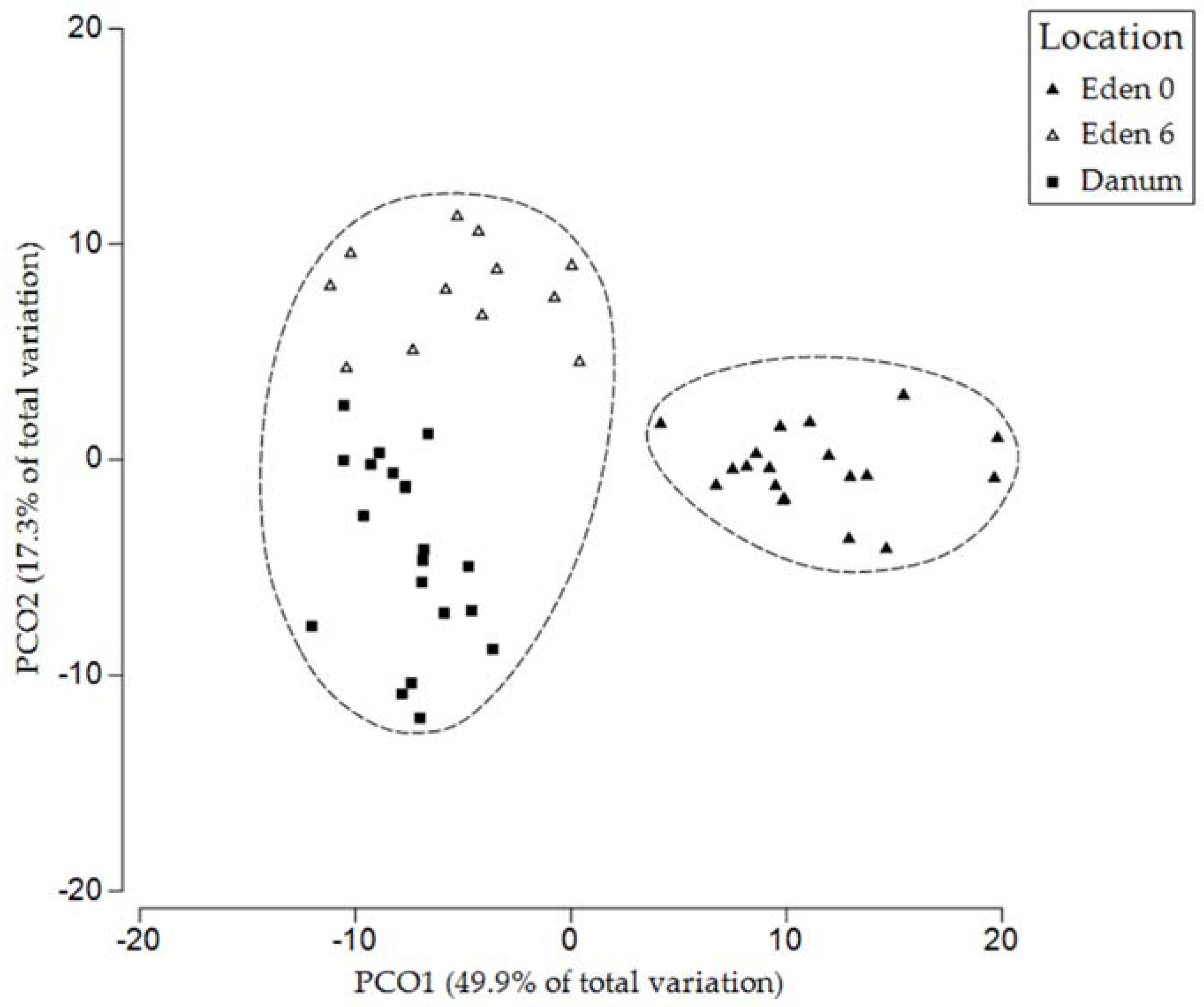
The soils collected from both the Eden Project ferns and those of Danum Valley contained a range of PLFAs of varying abundance. Chromatograms were used to identify the microbial groups present before their relative abundance was calculated (Figure 3). Analysis of PLFAs revealed the presence of Gram-positive and Gram-negative bacteria, actinomycetes and fungi in addition to further peaks, unassigned to specific taxa due to their ambiguous composition [50]. The relative abundance of individual PLFAs varied across collection times at the Eden Project, and also differed from those collected in Danum Valley (Figure 3). However, although the peaks of individual PLFAs differed, analysing entire community profiles revealed a pattern of increasing similarity between the ferns from the Eden Project after 6 months and the ferns from Danum Valley (Figure 4 and Figure 5).
Ferns from the Eden Project at time 0 had a relative abundance of 2% actinomycetes, 23% Gram-negative bacteria, 4% fungi, 24% Gram-positive bacteria, with a further 47% of PLFAs not specifically assigned (Figure 4a). After 6 months, these same ferns contained a microbial community consisting of 1.5% actinomycetes, 30% Gram-negative bacteria, 15% fungi, 14% Gram-positive bacteria, and 39.5% of PLFAs not specifically assigned (Figure 4b). Ferns from Danum Valley had a relative abundance of 3% actinomycetes, 19% Gram-negative bacteria, 11% fungi, 20% Gram-positive bacteria, and 47% of PLFAs not assigned to a specific microbial group (Figure 4c).
4. Discussion
We have shown that soil enzyme activity, used as an indicator of changes in soil microbial communities under simulated epiphytic conditions, did not respond to the addition of inorganic nitrogen or earthworm activity. Enzyme activity did, however, shift significantly in both treatments and in controls during the six-month experiment. Unlike terrestrial soils, the extreme shifts in temperature and moisture experienced by canopy soils will result in a microbial community adapted to the stresses of the canopy environment. Our results suggest that these stresses outweigh the effects of traditional predictors of microbial community dynamics.
Compared with the forest floor, suspended soils associated with epiphytes are nitrogen poor [53], and are likely to depend upon inorganic nitrogen from the atmosphere [54]. It is possible that the fern soil microorganisms at the Eden Project were not nitrogen limited, which would explain the lack of a response to the addition of nitrogen consistent with observations of other tropical soils [55]. It is also possible that the addition of ammonium nitrate failed to affect the microbial communities because our irrigation system delivered enough water (0.5 L per day) to flush the nitrogen straight through the fern soil. This amount of water passing through the fern soil is not unlike the amounts of rainfall observed in Danum Valley [44] and regular throughfall is a feature of bird’s nest ferns. The passage of water through Asplenium ferns has been shown to leach nutrients from their soils in tropical rain forests, with throughfall having higher total conductivity, and significantly more nitrate and potassium [36]. It seems reasonable to assume that the microbial communities of suspended soils must be adapted to conditions of constant leaching and low levels of nitrogen. The existing microbial communities associated with these ferns are unlikely to be determined by nutrient limitation alone. Finally, although an initial increase in microbial activity may have occurred following the addition of nitrogen, it was not possible to detect the legacy of this increase in microbial activity after three months.
Earthworms have for a long time been known to modify soil structure, increasing the aeration and pH of soils, conditions known to influence microbial community structure [22]. Earthworms are also known to colonise canopy soils along with a suite of other invertebrates [37,38], contributing to the decomposition of organic matter. Our study found no effect of earthworms on soil microbial community composition or activity. Traditional microcosm studies using an equivalent number of earthworms for the volume of soil analysed reported changes in soil microbial biomass and nutrient mineralisation [56,57]. The density of earthworms in the Eden Project ferns was much lower than that found in bird’s nest ferns in their natural environment [37,38], but if the worms caused significant changes in microbial community structure we are confident that this would have been apparent. In tropical rain forests, invertebrate decomposers and other animals play significant roles in modifying microbial composition and activity. We chose to focus only on earthworms, as these are well known decomposers, but in our study it seems that other factors outweighed any effect that earthworm activity may have had on microbial community composition and enzyme activity.
Our results challenge the traditional predictors of microbial community structure—predictors that were derived from the characteristics of terrestrial soils. This study shows that changes in microbial community composition in suspended soils are likely the result of the physical conditions under which epiphytic soils form. Over time, the legacy effect of being stored in relatively cool nursery conditions disappeared in the face of water loss and daily desiccation caused by the movement of air around the ferns and their associated soil. The functioning of all fern soils shifted over time, regardless of treatment. Hydrolytic enzyme activity (N-acetyl-glucosaminidase and β-glucosidase) decreased, whereas oxidative enzyme activity (phenol oxidase) increased. Changes in the expression of these extracellular enzymes could be interpreted as a change in the nutrient demands of the microbial communities. It could also, however, be the result of a shift in the structure of the microbial community itself.
One of the advantages of PLFA analysis is that, unlike genetic analysis, only the most recent microbial communities are detected [58]. This makes PLFA profiles useful for the detection of changes in microbial structure through time. Relative abundance of PLFAs and fungal:bacterial ratios confirmed relative increases in the biomass of fungi between three and six months. Further analysis of PLFA concentrations, rather than just relative abundances, would strengthen the measurement of shifts in the microbial biomass and related enzyme activity. Analysis of all microorganisms revealed no significant differences between treatments. However, it was striking to note that a relative increase in fungi brought the overall microbial composition of the Eden Project ferns after six months in line with ferns collected from their natural environment at Danum Valley. This increase in fungi is likely due to their ability to transfer water through networks of hyphae, thus allowing them to survive regular wetting and drying [59]. In temperate forests, fungi contribute more to decomposition in the canopy than on the forest floor [60]. In tropical forests, where the physical environment is more extreme, fungi are likely to be more important. Our study supports this notion, with relative increases in fungi as ferns were transferred from the cooler nursery conditions to hotter and drier epiphytic conditions. A range of epiphytes, including members of the genus Asplenium, have been shown to be physiologically resistant to desiccation [61]. Epiphytic plants are adapted to life in the harsh conditions of rain forest canopies. Our results suggest that the microbial communities associated with epiphytic plants may be more sensitive to physical conditions than was previously thought.
5. Conclusions
Contrary to the predictions of traditional soil science, the microorganisms of suspended soils respond to physical conditions such as exposure to the air, regular wetting and drying, and throughflow of water. As a result of their ability to resist desiccation, fungi, compared with other groups of microorganisms, are likely to play major roles in the ecological functioning of tropical rain forest canopies.
Acknowledgments
We wish to thank the South East Asia Rainforest Research Partnership (SEARRP) for assistance with fieldwork in Borneo. The Forest Research Centre of the Sabah Forestry Department assisted with the processing of samples. The Sabah Biodiversity Council and the Danum Valley Management Committee granted access to the research site. The Science Team at the Eden Project facilitated our tropical biome experiments, Paul Bowdler and Kevin Sudlow assisted with GC analysis. Josie Phillips provided helpful comments on the manuscript. This work was supported by NERC grant number BRIS/84/1015 and a UWE Bristol PhD scholarship.
Author Contributions
J.D., M.D.F.E. conceived and designed the research; J.D. performed the experiments; J.D. analysed the data; P.M., S.B., N.M. contributed reagents, materials and tools for microbial analysis; M.C. designed the irrigation equipment and provided climate data; J.D., M.D.F.E. wrote the manuscript. All authors contributed to manuscript revision.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ozanne, C.M.; Anhuf, D.; Boulter, S.L.; Keller, M.; Kitching, R.L.; Korner, C.; Meinzer, F.C.; Mitchell, A.W.; Nakashizuka, T.; Dias, P.L.; et al. Biodiversity Meets the Atmosphere: A Global View of Forest Canopies. Science 2003, 301, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Bohlman, S.A.; Matelson, T.J.; Nadkarni, N.M. Moisture and Temperature Patterns of Canopy Humus and Forest Floor Soil of a Montane Cloud Forest, Costa Rica. Biotropica 1995, 27, 13–19. [Google Scholar] [CrossRef]
- Nadkarni, N.M. Factors Affecting the Initiation and Growth of Aboveground Adventitious Roots in a Tropical Cloud Forest Tree: An Experimental Approach. Oecologia 1994, 100, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Hietz, P.; Wanek, W.; Wania, R.; Nadkarni, N.M. Nitrogen-15 Natural Abundance in a Montane Cloud Forest Canopy as an Indicator of Nitrogen Cycling and Epiphyte Nutrition. Oecologia 2002, 131, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, N.M.; Schaefer, D.; Matelson, T.J.; Solano, R. Comparison of Arboreal and Terrestrial Soil Characteristics in a Lower Montane Forest, Monteverde, Costa Rica. Pedobiologia 2002, 46, 24–33. [Google Scholar] [CrossRef]
- Wanek, W.; Arndt, S.K.; Huber, W.; Popp, M. Nitrogen Nutrition during Ontogeny of Hemiepiphytic Clusia Species. Funct. Plant Biol. 2002, 29, 733–740. [Google Scholar] [CrossRef]
- Clark, K.L.; Nadkarni, N.M.; Gholz, H.L. Retention of Inorganic Nitrogen by Epiphytic Bryophytes in a Tropical Montane Forest. Biotropica 2005, 37, 328–336. [Google Scholar] [CrossRef]
- Cardelús, C.L. Litter Decomposition within the Canopy and Forest Floor of Three Tree Species in a Tropical Lowland Rain Forest, Costa Rica. Biotropica 2010, 42, 300–308. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Vance, E.D.; Nadkarni, N.M. Microbial Biomass and Activity in Canopy Organic Matter and the Forest Floor of a Tropical Cloud Forest. Soil Biol. Biochem. 1990, 22, 677–684. [Google Scholar] [CrossRef]
- Lambais, M.R.; Crowley, D.E.; Cury, J.C.; Bull, R.C.; Rodrigues, R.R. Bacterial Diversity in Tree Canopies of the Atlantic Forest. Science 2006, 312, 1917. [Google Scholar] [CrossRef] [PubMed]
- Pittl, E.; Innerebner, G.; Wanek, W.; Insam, H. Microbial Communities of Arboreal and Ground Soils in the Esquinas Rainforest, Costa Rica. Plant Soil 2010, 329, 65–74. [Google Scholar] [CrossRef]
- Lambais, M.R.; Lucheta, A.R.; Crowley, D.E. Bacterial Community Assemblages Associated with the Phyllosphere, Dermosphere, and Rhizosphere of Tree Species of the Atlantic Forest are Host Taxon Dependent. Microb. Ecol. 2014, 68, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Lowman, M.D.; Schowalter, T.D. Plant Science in Forest Canopies—The First 30 Years of Advances and Challenges (1980–2010). New Phytol. 2012, 194, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Kitching, R.L.; Cao, M.; Creedy, T.J.; Fayle, T.M.; Freiberg, M.; Hewitt, C.; Itioka, T.; Koh, L.P.; Ma, K. Forests and their Canopies: Achievements and Horizons in Canopy Science. Trends Ecol. Evol. 2017, 32, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S.E.; Vitousek, P.M. Nutrient Limitation of Decomposition in Hawaiian Forests. Ecology 2000, 81, 1867–1877. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Extracellular Enzyme Activities and Carbon Chemistry as Drivers of Tropical Plant Litter Decomposition. Biotropica 2004, 36, 285–296. [Google Scholar] [CrossRef]
- Waldrop, M.; Firestone, M. Altered Utilization Patterns of Young and Old Soil C by Microorganisms Caused by Temperature Shifts and N Additions. Biogeochemistry 2004, 67, 235–248. [Google Scholar] [CrossRef]
- Cusack, D.; Chou, W.; Yang, W.; Harmon, M.; Silver, W.; Lidet Team. Controls on Long-Term Root and Leaf Litter Decomposition in Neotropical Forests. Glob. Chang. Biol. 2009, 15, 1339–1355. [Google Scholar] [CrossRef]
- Scullion, J.; Malik, A. Earthworm Activity Affecting Organic Matter, Aggregation and Microbial Activity in Soils Restored After Opencast Mining for Coal. Soil Biol. Biochem. 2000, 32, 119–126. [Google Scholar] [CrossRef]
- Frouz, J.; Prach, K.; Pižl, V.; Háněl, L.; Starý, J.; Tajovský, K.; Materna, J.; Balík, V.; Kalčík, J.; Řehounková, K. Interactions between Soil Development, Vegetation and Soil Fauna during Spontaneous Succession in Post Mining Sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Blouin, M.; Hodson, M.E.; Delgado, E.A.; Baker, G.; Brussaard, L.; Butt, K.R.; Dai, J.; Dendooven, L.; Pérès, G.; Tondoh, J. A Review of Earthworm Impact on Soil Function and Ecosystem Services. Eur. J. Soil Sci. 2013, 64, 161–182. [Google Scholar] [CrossRef] [Green Version]
- Birkhofer, K.; Schöning, I.; Alt, F.; Herold, N.; Klarner, B.; Maraun, M.; Marhan, S.; Oelmann, Y.; Wubet, T.; Yurkov, A. General Relationships between Abiotic Soil Properties and Soil Biota Across Spatial Scales and Different Land-use Types. PLoS ONE 2012, 7, e43292. [Google Scholar] [CrossRef] [PubMed]
- Swallow, M.; Quideau, S.; MacKenzie, M.; Kishchuk, B. Microbial Community Structure and Function: The Effect of Silvicultural Burning and Topographic Variability in Northern Alberta. Soil Biol. Biochem. 2009, 41, 770–777. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol Oxidase, Peroxidase and Organic Matter Dynamics of Soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Hendershot, J.N.; Read, Q.D.; Henning, J.A.; Sanders, N.J.; Classen, A.T. Consistently Inconsistent Drivers of Patterns of Microbial Diversity and Abundance at Macroecological Scales. Ecology 2017, 98, 1757–1763. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.D.; Vitousek, P.M. Responses of Extracellular Enzymes to Simple and Complex Nutrient Inputs. Soil Biol. Biochem. 2005, 37, 937–944. [Google Scholar] [CrossRef]
- Sinsabaugh, R.; Moorhead, D.; Linkins, A. The Enzymic Basis of Plant Litter Decomposition: Emergence of an Ecological Process. Appl. Soil Ecol. 1994, 1, 97–111. [Google Scholar] [CrossRef]
- Paterson, E.; Osler, G.; Dawson, L.A.; Gebbing, T.; Sim, A.; Ord, B. Labile and Recalcitrant Plant Fractions are Utilised by Distinct Microbial Communities in Soil: Independent of the Presence of Roots and Mycorrhizal Fungi. Soil Biol. Biochem. 2008, 40, 1103–1113. [Google Scholar] [CrossRef]
- Kramer, S.; Dibbern, D.; Moll, J.; Huenninghaus, M.; Koller, R.; Krueger, D.; Marhan, S.; Urich, T.; Wubet, T.; Bonkowski, M. Resource Partitioning between Bacteria, Fungi, and Protists in the Detritusphere of an Agricultural Soil. Front. Microbiol. 2016, 7, 1524. [Google Scholar] [CrossRef] [PubMed]
- Charteris, A.; Knowles, T.; Michaelides, K.; Evershed, R. Compound-specific Amino Acid 15N Stable Isotope Probing of Nitrogen Assimilation by the Soil Microbial Biomass using Gas Chromatography/Combustion/Isotope Ratio Mass Spectrometry. Rapid Commun. Mass Spectrom. 2016, 30, 1846–1856. [Google Scholar] [PubMed]
- Matson, P.; Lohse, K.A.; Hall, S.J. The Globalization of Nitrogen Deposition: Consequences for Terrestrial Ecosystems. AMBIO J. Hum. Environ. 2002, 31, 113–119. [Google Scholar] [CrossRef]
- Mo, J.; Zhang, W.; Zhu, W.; Gundersen, P.; Fang, Y.; Li, D.; Wang, H. Nitrogen Addition Reduces Soil Respiration in a Mature Tropical Forest in Southern China. Glob. Chang. Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Yatabe, Y.; Shinohara, W.; Matsumoto, S.; Murakami, N. Patterns of Hybrid Formation among Cryptic Species of Bird-nest Fern, Asplenium nidus Complex (Aspleniaceae), in West Malesia. Bot. J. Linn. Soc. 2009, 160, 42–63. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Kolasa, J.; Bengtsson, J.; Gonzalez, A.; Lawler, S.P.; Miller, T.E.; Munguia, P.; Romanuk, T.; Schneider, D.C.; Trzcinski, M.K. Are Natural Microcosms Useful Model Systems for Ecology? Trends Ecol. Evol. 2004, 19, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Turner, E.C.; Snaddon, J.L.; Johnson, H.R.; Foster, W.A. The Impact of Bird’s Nest Ferns on Stemflow Nutrient Concentration in a Primary Rain Forest, Sabah, Malaysia. J. Trop. Ecol. 2007, 23, 721–724. [Google Scholar] [CrossRef]
- Ellwood, M.D.F.; Jones, D.T.; Foster, W.A. Canopy Ferns in Lowland Dipterocarp Forest Support a Prolific Abundance of Ants, Termites, and Other Invertebrates. Biotropica 2002, 34, 575–583. [Google Scholar] [CrossRef]
- Ellwood, M.D.F.; Foster, W.A. Doubling the Estimate of Invertebrate Biomass in a Rainforest Canopy. Nature 2004, 429, 549–551. [Google Scholar] [CrossRef] [PubMed]
- Ellwood, M.D.F.; Manica, A.; Foster, W.A. Stochastic and Deterministic Processes Jointly Structure Tropical Arthropod Communities. Ecol. Lett. 2009, 12, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Fayle, T.M.; Eggleton, P.; Manica, A.; Yusah, K.M.; Foster, W.A. Experimentally Testing and Assessing the Predictive Power of Species Assembly Rules for Tropical Canopy Ants. Ecol. Lett. 2015, 18, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Ellwood, M.D.F.; Blüthgen, N.; Fayle, T.M.; Foster, W.A.; Menzel, F. Competition can lead to unexpected patterns in tropical ant communities. Acta Oecol. 2016, 75, 24–34. [Google Scholar] [CrossRef]
- Donald, J.; Maxfield, P.; Murray, D.; Ellwood, M.D.F. How Tropical Epiphytes at the Eden Project Contribute to Rainforest Canopy Science. Sibbaldia 2017, 14, 55–68. [Google Scholar]
- Warmington, R.; Eden Project, Cornwall, UK. Personal communication, 2017.
- Marsh, C.W.; Greer, A.G. Forest Land-use in Sabah, Malaysia: An Introduction to Danum Valley. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1992, 335, 331–339. [Google Scholar] [CrossRef]
- Scheffers, B.R.; Evans, T.A.; Williams, S.E.; Edwards, D.P. Microhabitats in the Tropics Buffer Temperature in a Globally Coherent Manner. Biol. Lett. 2014, 10, 20140819. [Google Scholar] [CrossRef] [PubMed]
- Donald, J.; Bonnett, S.; Maxfield, P.; Ellwood, M.D.F. The Relative Importance of Invertebrate and Microbial Decomposition in a Rainforest Restoration Project. Restor. Ecol. 2017. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Physiol. Pharmacol 1959, 37, 911–917. [Google Scholar]
- Dickson, L.; Bull, I.D.; Gates, P.J.; Evershed, R.P. A Simple Modification of a Silicic Acid Lipid Fractionation Protocol to Eliminate Free Fatty Acids from Glycolipid and Phospholipid Fractions. J. Microbiol. Methods 2009, 78, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Frostegård, Å.; Bååth, E. The use of Phospholipid Fatty Acid Analysis to Estimate Bacterial and Fungal Biomass in Soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Rubino, M.; Dungait, J.; Evershed, R.; Bertolini, T.; De Angelis, P.; D’Onofrio, A.; Lagomarsino, A.; Lubritto, C.; Merola, A.; Terrasi, F. Carbon Input Belowground is the Major C Flux Contributing to Leaf Litter Mass Loss: Evidences from a 13 C Labelled-Leaf Litter Experiment. Soil Biol. Biochem. 2010, 42, 1009–1016. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2015. [Google Scholar]
- Wanek, W.; Zotz, G. Are Vascular Epiphytes Nitrogen Or Phosphorus Limited? A Study of Plant 15N Fractionation and Foliar N: P Stoichiometry with the Tank Bromeliad Vriesea sanguinolenta. New Phytol. 2011, 192, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.E.; Rundel, P.W.; Cardelús, C.L. The Influence of Life Form on Carbon and Nitrogen Relationships in Tropical Rainforest Ferns. Oecologia 2007, 153, 225. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.A.; McDowell, W.H.; Townsend, A.R.; Vitousek, P.M. The globalization of N deposition: Ecosystem consequences in tropical environments. In New Perspectives on Nitrogen Cycling in the Temperate and Tropical Americas; Townsend, A.R., Ed.; Springer: Dordrecht, The Netherlands, 1999; pp. 67–83. [Google Scholar]
- Araujo, Y.; Luizão, F.J.; Barros, E. Effect of Earthworm Addition on Soil Nitrogen Availability, Microbial Biomass and Litter Decomposition in Mesocosms. Biol. Fertil. Soils 2004, 39, 146–152. [Google Scholar] [CrossRef]
- Bohlen, P.J.; Edwards, C.A. Earthworm Effects on N Dynamics and Soil Respiration in Microcosms Receiving Organic and Inorganic Nutrients. Soil Biol. Biochem. 1995, 27, 341–348. [Google Scholar] [CrossRef]
- Zelles, L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: A review. Biol Fertil. Soils 1999, 29, 111–129. [Google Scholar] [CrossRef]
- Guhr, A.; Marzini, C.; Borken, W.; Poll, C.; Matzner, E. Effect of Water Redistribution by Two Distinct Saprotrophic Fungi on Carbon Mineralization and Nitrogen Translocation in Dry Soil. Soil Biol. Biochem. 2016, 103, 380–387. [Google Scholar] [CrossRef]
- Rousk, J.; Nadkarni, N.M. Growth measurements of saprotrophic fungi and bacteria reveal differences between canopy and forest floor soils. Soil Biol. Biochem. 2009, 41, 862–865. [Google Scholar] [CrossRef]
- Watkins, J.E.; Mack, M.C.; Sinclair, T.R.; Mulkey, S.S. Ecological and Evolutionary Consequences of Desiccation Tolerance in Tropical Fern Gametophytes. New Phytol. 2007, 176, 708–717. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Asplenium bird’s nest ferns in (a) the rain forest biome at the Eden Project, Cornwall, U.K.; (b) Danum Valley, Sabah, Malaysian Borneo.
Figure 1.
Asplenium bird’s nest ferns in (a) the rain forest biome at the Eden Project, Cornwall, U.K.; (b) Danum Valley, Sabah, Malaysian Borneo.

Figure 2.
(a) N-acetyl-glucosaminidase activity, (b) β-glucosidase activity, (c) phenol oxidase activity, (d) fungal:bacterial ratio of bird’s nest ferns at 0, 3, 6 months (Mean ± s.e., n = 5). Treatments are indicated in the legend as follows: AN = ammonium nitrate; W = worms; C = control; AN + W = ammonium nitrate plus worms.
Figure 2.
(a) N-acetyl-glucosaminidase activity, (b) β-glucosidase activity, (c) phenol oxidase activity, (d) fungal:bacterial ratio of bird’s nest ferns at 0, 3, 6 months (Mean ± s.e., n = 5). Treatments are indicated in the legend as follows: AN = ammonium nitrate; W = worms; C = control; AN + W = ammonium nitrate plus worms.

Figure 3.
Relative abundance of fern soil PLFAs from experimental bird’s nest ferns at the Eden Project prior to suspension and after 6 months under epiphytic conditions, and from natural specimens from Danum Valley (all means ± s.e., n = 20).
Figure 3.
Relative abundance of fern soil PLFAs from experimental bird’s nest ferns at the Eden Project prior to suspension and after 6 months under epiphytic conditions, and from natural specimens from Danum Valley (all means ± s.e., n = 20).
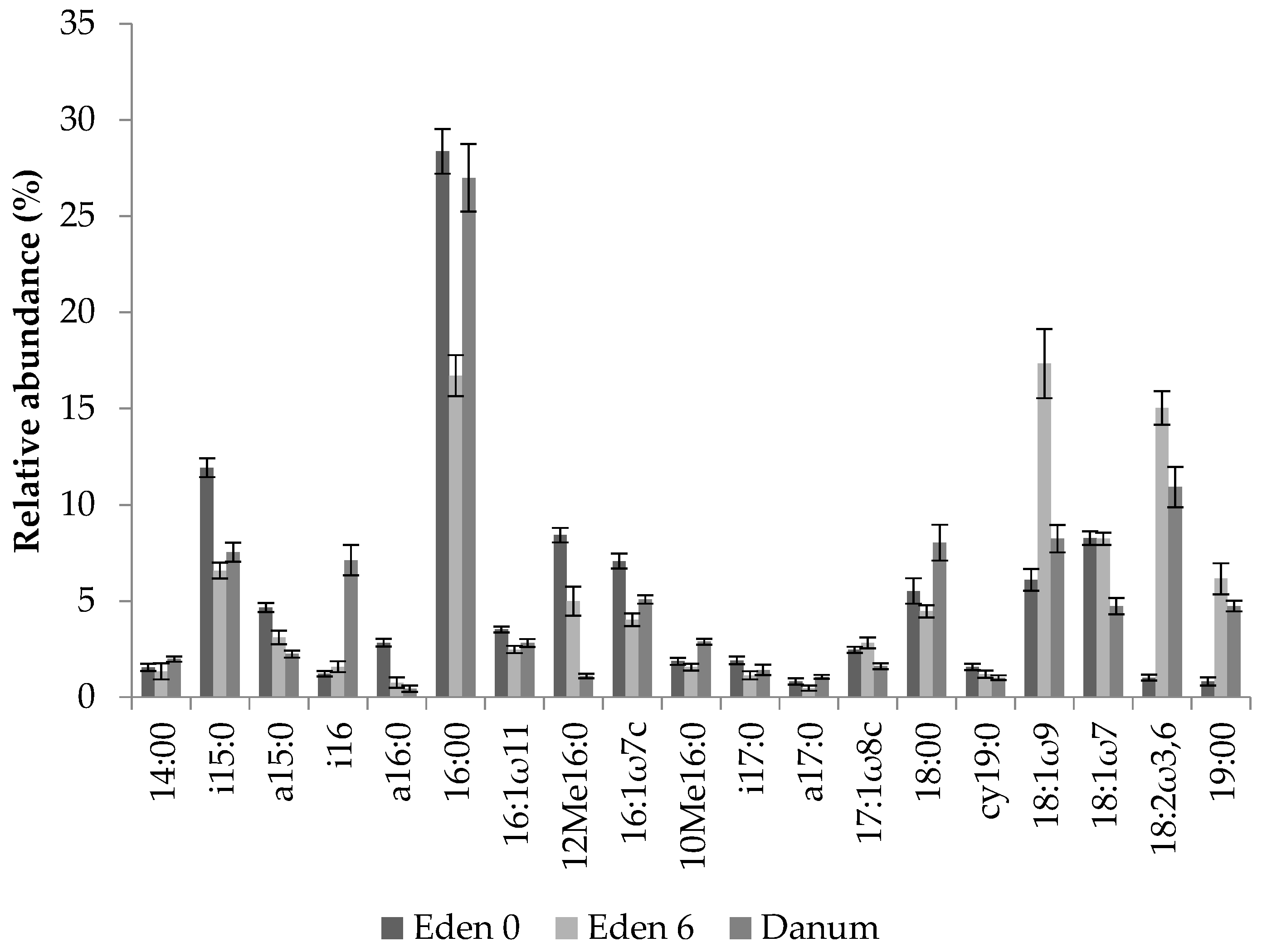
Figure 4.
The microbial community composition of (a) 20 bird’s nest ferns from the Eden Project prior to experimental manipulation; (b) 20 bird’s nest ferns after 6 months in the rain forest biome; (c) 20 natural bird’s nest ferns from Danum Valley.
Figure 4.
The microbial community composition of (a) 20 bird’s nest ferns from the Eden Project prior to experimental manipulation; (b) 20 bird’s nest ferns after 6 months in the rain forest biome; (c) 20 natural bird’s nest ferns from Danum Valley.
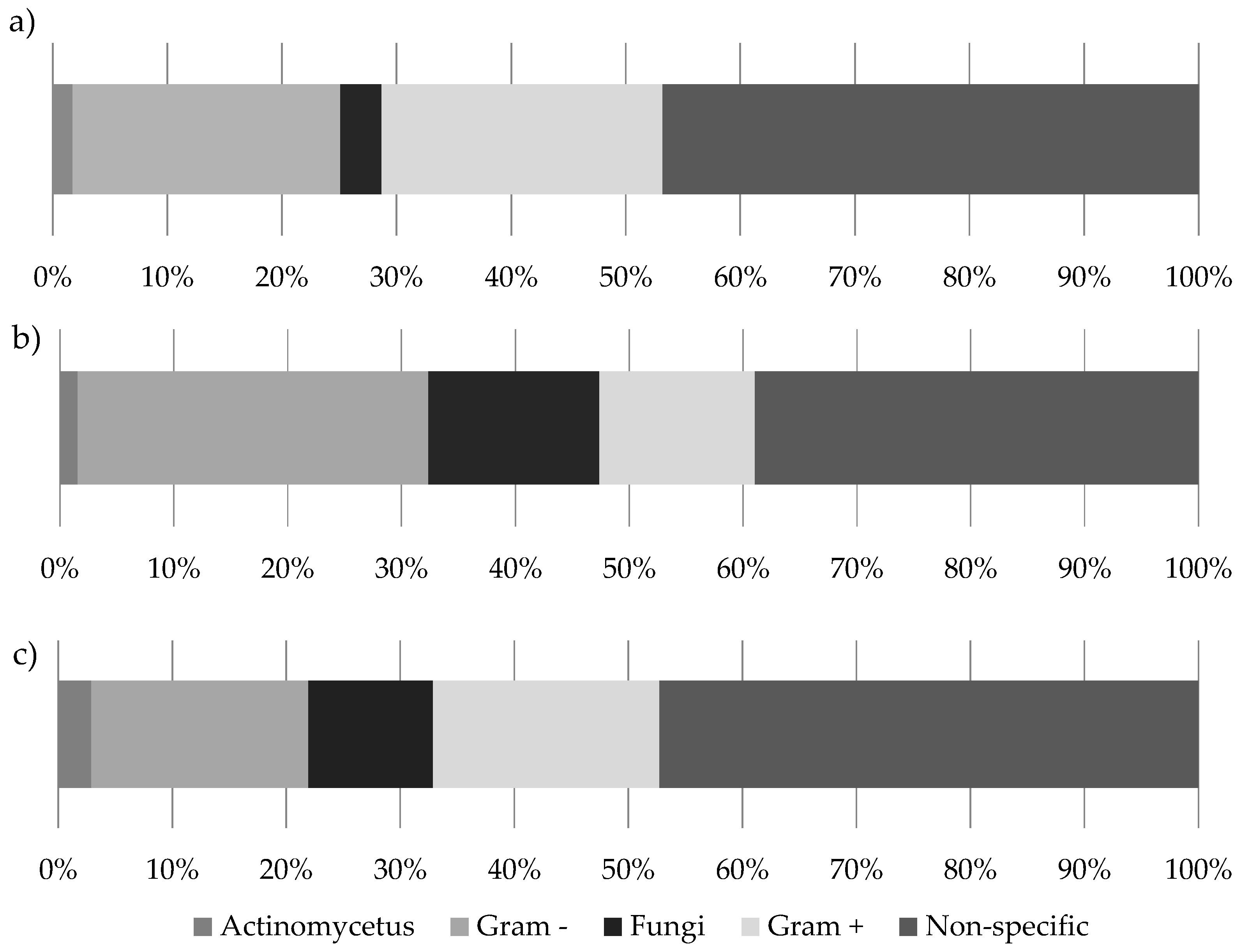
Figure 5.
The PLFA profile of bird’s nest fern soils collected from the Eden Project prior to placement in the canopy, after 6 months in the canopy, and from natural fern soils collected in Danum Valley. Dashed lines enclose points within 80% similarity envelopes.
Figure 5.
The PLFA profile of bird’s nest fern soils collected from the Eden Project prior to placement in the canopy, after 6 months in the canopy, and from natural fern soils collected in Danum Valley. Dashed lines enclose points within 80% similarity envelopes.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Classification of Phospholipid Fatty Acid (PLFA) peaks as specified by [50].
Table 1.
Classification of Phospholipid Fatty Acid (PLFA) peaks as specified by [50].
| Microbial Group | PLFA Peaks |
|---|---|
| Gram + bacteria | i15:0, a15:0, i16:0, a16:0, i17:0, a17:0 |
| Gram − bacteria | 16:1ω7c, cy19:0, 18:1ω9, 18:1ω7 |
| Actinomycetes | 10Me16:0 |
| Fungi | 18:2ω3,6 |
Table 2.
Two-way ANOVA results of time and treatment on enzyme activities and fungal:bacterial ratios in soil associated with bird’s nest ferns.
Table 2.
Two-way ANOVA results of time and treatment on enzyme activities and fungal:bacterial ratios in soil associated with bird’s nest ferns.
| N-a-glucosaminidase | β-glucosidase | Phenol oxidase | Fungi:Bacteria | |
|---|---|---|---|---|
| Time | F1,52 = 42.2 p < 0.001 | F1,49 = 46.0 p < 0.001 | F1,52 = 55.0 p < 0.001 | F1,43 = 29.1 p < 0.001 |
| Treatment | F3,52 = 0.918 p = 0.439 | F3,49 = 0.754 ns p = 0.526 | F3,52 = 0.508 p = 0.678 | F3,43 = 2.30 p = 0.090 |
| Ti x Tr | F3,52 = 0.910 ns p = 0.443 | F3,49 = 0.288 ns p = 0.834 | F3,52 = 0.666 ns p = 0.577 | F3,43 = 0.455 ns p = 0.722 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Donald, J.; Bonnett, S.; Cutler, M.; Majalap, N.; Maxfield, P.; Ellwood, M.D.F. Physical Conditions Regulate the Fungal to Bacterial Ratios of a Tropical Suspended Soil. Forests 2017, 8, 474. https://doi.org/10.3390/f8120474
AMA Style
Donald J, Bonnett S, Cutler M, Majalap N, Maxfield P, Ellwood MDF. Physical Conditions Regulate the Fungal to Bacterial Ratios of a Tropical Suspended Soil. Forests. 2017; 8(12):474. https://doi.org/10.3390/f8120474
Chicago/Turabian StyleDonald, Julian, Sam Bonnett, Michael Cutler, Noreen Majalap, Pete Maxfield, and M. D. Farnon Ellwood. 2017. "Physical Conditions Regulate the Fungal to Bacterial Ratios of a Tropical Suspended Soil" Forests 8, no. 12: 474. https://doi.org/10.3390/f8120474
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.