Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding


Thuenen Institute of Forest Genetics, 22927 Grosshansdorf, Germany
*
Author to whom correspondence should be addressed.
Forests 2017, 8(12), 492; https://doi.org/10.3390/f8120492
Submission received: 18 September 2017
/
Revised: 5 December 2017
/
Accepted: 6 December 2017
/
Published: 8 December 2017
(This article belongs to the Special Issue Gene-Based SNP Discovery and Diversity of Forests Trees)
Abstract
:Within the genus Populus, about 30 species are classified into six sections, of which some are cross-compatible. Besides naturally occurring hybrids, huge breeding programs have led to a high number of artificially produced hybrids, for which the determination of genetically involved species by morphological characteristics is often difficult. This necessitates the use of molecular markers for the identification of both maternal as well as paternal species, and in the case of complex hybrids, the genealogy. For this reason, we developed new chloroplast and nuclear markers for the differentiation of up to 19 poplar species, with one to 32 individuals per species regularly used in breeding programs based on already known barcoding, other chloroplast regions, and nuclear genes of interest. We developed methods to identify species by either species-specific nucleotide variations or, when no initial information for the species was given, by using a set of markers either in a procedure of exclusion or in a multiplexed marker set. The developed markers can all be used with low-cost equipment, and some can additionally be applied using a genetic analyzer. We combined these markers in multiplexes for a very fast and easy-to-use application for the identification of poplar species and their hybrids.
1. Introduction
A high growth rate and a broad applicability ranging from wood and paper to energy production has led to the widespread cultivation of poplar (Populus) species in Europe and North America [1]. The approximately 29 species within the genus Populus [2] are classified into six sections, of which some are intersectional and cross-compatible. In particular, hybrids between various Populus species belonging to the same or different sections are commonly used in short rotation plantations (SRC) for biomass production because of their superior growth and advanced resistance traits. For classical breeding, these features of the poplar species are an advantage, and therefore, a lot of artificially and naturally produced interspecies hybrids exist today [3]. However, these long-lasting breeding activities, including repetitive crosses and back-crosses, have caused a lack of background information for many of the commercially available hybrid clones that were originally used. Additionally, due to the extreme variability of Populus hybrids, species identification within the genus Populus using morphological characteristics has sometimes proven to be difficult. However, species identification is necessary for the registration of new clones, yield stability within SRC, and the design of complex breeding strategies. For the identification of clones, highly polymorphic microsatellite markers (SSR: Short Sequence Repeats) are long established, and a broad range of poplar SSR markers are available for this purpose [4,5,6,7]. The transferability of SSR markers to different poplar sections has been examined previously [8]. Besides clone identification, only a few studies report on species identification using single nucleotide polymorphisms (SNPs) in Populus [9,10,11,12,13], all of which focussed on only a narrow range of species. For these reasons, molecular markers for species and hybrid identification within the genus Populus are desirable.
Due to its small genome size, huge genomic resources, and easy-to-use biotechnological handling ability, the genus Populus has been selected as a model species for tree genomics, resulting in the publication of the full genome of western black cottonwood, P. trichocarpa, only a decade ago [14]. Additionally to P. trichocarpa, complete chloroplast (cp) DNA genome sequences are available for P. alba [15], P. balsamifera, P. fremontii [16], P. euphratica [17], P. tremula [18], P. cathayana [19], and P. yunnanensis [20]. In deciduous trees, both chloroplasts and mitochondria are maternally inherited [21], while the nuclear genome is biparentally inherited. The advantages of using cp genome markers are: (a) only one allele exists per cell and organism; (b) organelle DNA is present in many copies per cell; and (c) thus, the DNA can be more easily extracted, even from low-quantity or low-quality samples. These reasons led to the choice of cp DNA markers by the Barcode of Life Consortia for the genetic differentiation of all eukaryotic species [22]. Several cp regions have been tested for their ability as barcodes (e.g., Chase et al. (2005), Lahaye et al. (2008), Hollingsworth et al. (2009) [23,24,25]), leading to the recommendation to use molecular markers based on DNA variations in two chloroplast regions (rbcL and matK) [25].
The application of these two barcoding regions for different poplar species led to the result that these two cp regions are not sufficient for species differentiation within the genus Populus [26,27,28]. Therefore, 40 cp regions have been tested using seven poplar species (P. tremula, P. tremuloides, P. alba, P. trichocarpa, P. maximowiczii, P. nigra, and P. deltoides) in 2012 [26]. From these already designed primers, 17—plus one new primer—have been validated with an additional seven species (P. simonii, P. koreana, P. cathayana, P. szechuanica, P. ussuriensis, P. wilsonii, and P. euphratica) in 2014 [28]. During 2008 and 2018, an interdisciplinary breeding study in Germany used 19 poplar species to produce new high-performance clones (www.fastwood.org). For this reason, the study presented here has been expanded to these 19 poplar species. For additional identification of large insertion–deletion polymorphisms (indels), we only recently used the whole cpDNA genome sequence information of the poplar species mentioned above [18].
By applying cp or mitochondrial marker [13] information, it is possible to identify the maternal partner of a crossing. Nuclear DNA markers are necessary for the determination of the paternal cross parent, and therefore the possible identification of hybrids. Several nuclear regions have been tested in earlier investigations for their suitability to find species-specific SNPs in different poplar species, e.g., Meirmans et al. (2007) [11] investigated seven nuclear regions in five species, and found 12 species-specific SNPs. A further example is the work of Fladung and Buschbom (2009) [12], who also tried seven regions (partly the same as in Meirmans et al. (2007) [11]) to create a preliminary phylogeny of six poplar species. With the purpose of identifying the paternal cross parent, four regions from the four genes GA20ox, TB1, KNOX_int2, and LFY, which were described earlier and considered most suitable for poplar [11,12], have been applied for marker development.
The aim of this study was to develop easy-to-use marker sets for the differentiation of the 19 most widely used poplar species. To this end, we combined newly designed chloroplast and nuclear markers with previously developed chloroplast markers, and evaluated their potential for species differentiation. We present (i) one set of four cp markers to identify the maternal parent; (ii) a second set of six nuclear markers for the identification of the paternal cross parent; and (iii) 15 cp and nuclear (nc) markers that may be used individually, even in low-equipped laboratories. Two of these individually usable markers are also included in the nc marker set. All of them can be used for the unambiguous assignment of a high number of poplar species, and thus represent an important step forward in marker-assisted breeding.
2. Materials and Methods
2.1. Sample Collection
In addition to the sampling sources described by Schroeder et al. (2012) and Schroeder & Fladung (2014) [26,28], colleagues provided the plant material of: 80 individuals of P. balsamifera from Agricultural and Agri-Food Canada, five individuals each of P. adenopoda and the hybrid P. × tomentosa (P. adenopoda × P. alba) from the Agricultural University of Hebei (China), 77 P. fremontii from the Universidad Juárez del Estado de Durango, México, 67 P. grandidentata from the University of Minnesota (Department of Forest Resources), and two specimens of P. lasiocarpa from the Botanical Gardens in Tübingen and Dresden, both in Germany. Overall, this study included 19 poplar species from five sections.
2.2. Tested Chloroplast and Nuclear Regions
From the 40 chloroplast regions earlier described and tested, four intergenic spacers have been chosen (trnH_psbA, psbK_psbI, rpoC2_rpoC1, and trnV_atpE) that contained SNPs for species discrimination and had already been used for the identification of single species, as described in Schroeder et al. (2012) and Schroeder and Fladung (2014) [26,28]. Additionally, promising indels have been found beside SNPs in the same investigations in four chloroplast regions (psbA_matK, rps4_trnL, rps2_rpoC2, and trnC_petN). In a previous paper, we aligned whole plastome sequences of eight poplar species (P. tremula, P. alba, P. trichocarpa, P. balsamifera, P. euphratica, P. cathayana, P. fremontii, and P. yunnanensis) and looked in the first line for long indels [18]. One of the cp regions examined there, psaA_ycf13, was very promising because of a high number of polymorphisms, and has therefore been included as a new region to be tried in the present study. These nine chloroplast regions were tested in a total of 19 poplar species, partly with the already published primers, and partly with newly designed primers, as given in Table 1. Based on the above-mentioned whole plastome alignment [18], a very long indel was identified in the region psbE_petL that was further investigated with up to 18 poplar species in this study.
For the development of markers within the biparentally inherited nuclear genome, the following genes/introns were tested with several primer combinations in all 19 poplar species: the gene Gibberellin acid 20-oxidase (GA20ox), an intron of the gene Class 1 KNOTTED3-like homeobox (KNOX3-int2), the meristem identity gene FLORICAULA/LEAFY (LFY), and the transcription factor TEOSINTE BRANCHED (TB1). In Table 1, only those primer combinations are listed that were successfully validated for species identification.
2.3. DNA Extraction and PCR Amplification
One cm2 of a single leaf was ground to powder in liquid nitrogen. Total DNA was extracted, following a modified alkyltrimethylammonium bromide (ATMAB)) protocol [30]. A standard protocol was used for PCR reactions [26]. The PCR reactions contained 20 to 50 ng template DNA, 50 mM KCl, 20 mM Tris-HCl (pH 8.4), 1.8 mM MgCl2, 200 μM dNTPs, 1 unit Taq polymerase, and 0.4 μM of each primer in a total volume of 15 μL. PCR was carried out in a Biometra Personal Thermocycler (Goettingen, Germany) with a pre-denaturation step at 94 °C for 4 min, followed by 30 cycles of 94 °C for 1 min, a suitable annealing temperature for each primer combination (between 52 °C and 60 °C, Table 1) for 45 s or 1 min, 72 °C for 1 min, and a final elongation at 72 °C for 10 min. PCR amplification products were checked on a 1% agarose gel stained with Roti-Safe GelStain (Carl Roth GmbH & Co. KG, Karlsruhe, Germany).
2.4. Sanger Sequencing and PCR-RFLP
For sequencing, 15 μL of the PCR product was purified either with 5 μL of 4 M lithium chloride or using the “High Pure PCR Product Purification” kit from Roche (Mannheim, Germany). For Sanger sequencing, the service provided by StarSeq (Mainz, Germany) was used. Up to five individuals (as far as available) per species from the additional five species and the above-mentioned nine cp (without psbE_petL) primer combinations were sequenced. The cp region psbE_petL was not sequenced with further individuals, because it was identified using the whole genome alignment, and in these sequences, only the long indel was striking in this region. For the four nuclear regions, up to five individuals from all 19 poplar species were sequenced using the nine primer combinations given in Table 1. Obtained sequences were aligned and screened for the presence of potential species-specific SNPs and indels by using either the software SeqMan 7.1.0 from DNAStar (Madison, WI, USA) or Sequencher 4.9 (Gene Codes Corporation, Ann Arbor, MI, USA). Differentiating sequences (SNPs, indels) were checked for a possible application of restriction enzymes using the software NEBcutter V2.0 from New England BioLabs Inc. (Ipswich, MA, USA). The already sequenced individuals from earlier studies of the cp genome (three to 10 individuals per species for 14 species [26,28]), plus up to 32 individuals (as far as available) of the additional five species, have been used to validate the usability of PCR-RFLPs (Restriction Fragment Length Polymorphisms) for species differentiation (Table 2). For PCR-RFLP application, the amplicons (10 μL) were digested with two units of the respective restriction enzyme in a total volume of 20 μL restriction enzyme buffer, according to the manufacturer’s recommendations (MBI Fermentas, St. Leon-Rot, Germany; New England BioLabs Inc., Ipswich, MA, USA). Aliquots (10 μL to 20 μL) of the digested DNA were separated on 1% to 2% agarose gels, and the resultant bands were visualized using Roti-Safe GelStain (Carl Roth GmbH & Co. KG, Karlsruhe, Germany). A 100 bp ladder obtained from GIBCO BRL (Life Technologies, Martinsried, Germany) was used as a size marker.
3. Results
3.1. Amount of Variations
3.1.1. Variations within the Chloroplast
The primer combinations used for the nine above-mentioned cp regions (without psbE_petL) were validated for the 19 poplar species, and resulted in amplification products for all of the species, which were successfully sequenced for the five newly added species. These new sequences were aligned together with the previous produced sequences [26,28] and checked for SNPs and indels. The number of SNPs and indels varied for the different species, where the highest number of SNPs and indels could be identified for P. tremuloides (Table 2). For 14 species, species-specific SNPs and/or indels could be identified, and for eight of these species, restriction enzymes were also available for application in PCR-RFLPs (Table 2). The number of tested specimens per species varied (Table 2) because of the restricted availability of individuals for some species. This has to be kept in mind for the evaluation of identified possible species-specific variations.
Within the region psbE_petL resulting from the whole cp genome alignment [18], a large indel of 99 bp was identified, where P. tremula was 99 bp shorter than the other species. The indel has been validated with one to three individuals per species for 17 of the 19 species (P. grandidentata and P. fremontii were not included), and with an additional 14 P. tremula specimens simply on an agarose gel. All 14 tested P. tremula specimens revealed the PCR fragment that was 99 bp shorter than the other species.
3.1.2. Variations within the Nuclear Regions
The four nuclear primer combinations (Table 1) could be successfully amplified for all 19 species. Sequencing was performed for the five additional poplar species. An alignment of the sequences of all 19 species was used for the search for SNPs and indels. Overall, we identified between 119 (P. alba) and 17 SNPs (P. cathayana), when considering 14 species where all of the four nuclear regions were sequenced (Table 3). For all except three poplar species, species-specific SNPs and/or indels could be identified, and for 10 species, the identification was possible using restriction enzymes in simple PCR-RFLP analyses (Table 3).
3.1.3. Variations per Base Pair
The amount of nucleotide variations within the chloroplast and the nuclear regions was calculated as the percent variation for each species, and summed over all of the regions to better compare the overall variability between the two genomes. In total, for the chloroplast genome, 5511 bp were sequenced, and for the nuclear regions, 4676 bp were sequenced (Table 4).
The value for the overall variations per 100 base pairs is nearly three times higher in the nuclear regions (17.8%) than in the cp regions (6.5%) analyzed in this study. For the species-specific SNPs, the difference between the variation density in the cp and nuclear genome is lower (Table 4). The highest variability in the nuclear genomic regions was detected in the two aspen P. tremula and P. tremuloides, and the white poplar P. alba, with all three belonging to the same section, Populus. A high number of variations documented for these three species are shared by only these three, and differentiate them from all of the other species. The other two species from the section Populus (P. adenopoda, P. grandidentata) are much less variable, though based on the same number of sequenced individuals.
3.2. Applicability of the Molecular Markers
For the application of the above-described cp and nc genetic markers in species identification, different prerequisites are conceivable: (i) if detailed information about a species is known, and only a test will be performed to determine whether this is the given species or not, then the use of a single SNP or indel is reasonable to prove a given claim (Section 3.2.1); (ii) if no information at all is available, then a procedure of species exclusion should be applied (Section 3.2.2 and Section 3.2.3). Finally, applying these marker sets, reconstruction of the genealogy of long-used (“old”) hybrids is attempted (Section 3.2.4).
3.2.1. Use of Single Markers for Species Identification
In Table 2 and Table 3, species-specific SNPs and indels are listed. Some of these SNPs and indels are already proved in the daily routine for testing the species identity in our laboratory. Thus, in Table 5, an overview of these routinely used markers for species identification is given together with detailed information of fragment sizes after restriction for an easy application on agarose gels.
3.2.2. Cp Indel Marker Set
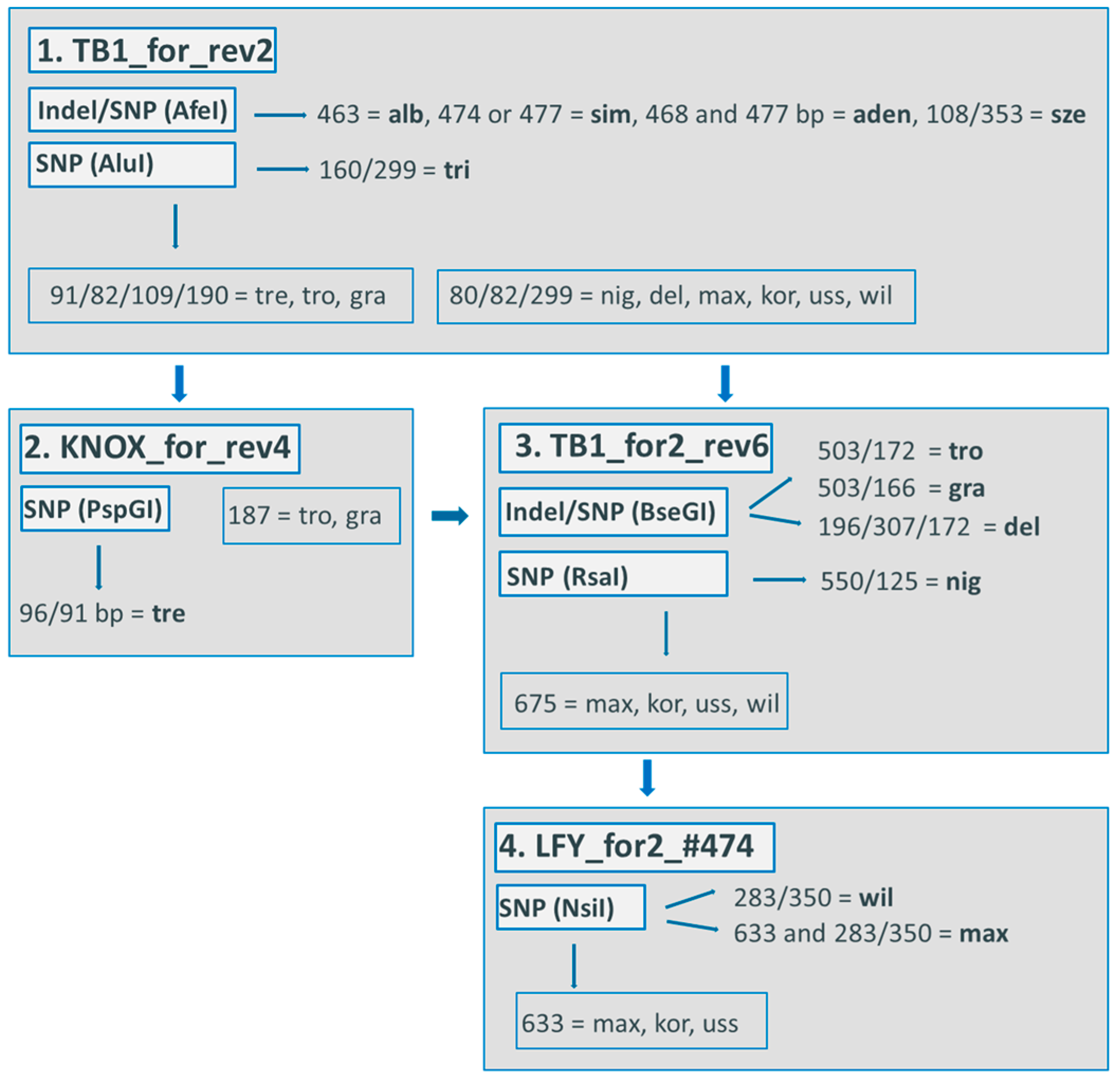
The combination of four indel markers led to the differentiation of up to 16 of the 19 poplar species (Figure 1). Species identification is possible through using a procedure of exclusion. In the first step, the following four species are identified by individual fragment length: P. alba (alb), P. nigra (nig), P. adenopoda (aden), and P. euphratica (eup). The other species are divided into two groups. In the second step, P. tremula (tre) could be excluded, and what was formerly two groups were divided into five groups. Due to this division, the second step led to the identification of nine further species, i.e., P. balsamifera (bal), P. szechuanica (sze), P. wilsonii (wil), P. maximowiczii (max), P. tremuloides (tro), P. trichocarpa (tri), P. fremontii (fre), P. grandidentata (gra), and P. deltoides (del). The last step led to the differentiation of P. simonii (sim) and P. lasiocarpa (las), which showed fragment lengths of 445 bp and 441 bp, respectively. When individuals of both species revealed a 443 bp-long fragment, no differentiation was possible. The remaining three species, P. koreana (kor), P. ussuriensis (uss), and P. cathayana (cath), were not distinguishable by any chloroplast marker.
This method is also usable in laboratories without a genetic analyzer, because the differences in the fragment size can be visualized on polyacrylamide gels.
We also developed a multiplexed marker set for a genetic analyzer (ABI3730 Capillary sequencer, Applied Biosystems, Foster City, CA, USA). To apply this method, two PCR runs, each in a total volume of 15 μL, are necessary because of different annealing temperatures, followed by a multiplexing on a plate for the genetic analyzer (Table 6).
Both PCRs runs were carried out in a Biometra Personal Thermocycler (Goettingen, Germany) with a pre-denaturation step at 94 °C for 4 min, followed by 28 cycles of 94 °C for 45 s, an annealing temperature of 60 °C for PCR 1 and 55 °C for PCR 2 (Table 6) for 45 s, and then 72 °C for 1 min, and a final elongation at 72 °C for 10 min.
3.2.3. Procedure of Exclusion for Nuclear Markers
For the identification of the species with nuclear markers, we used a combination of four regions (four PCRs) with six SNPs and two indels in a procedure of exclusion to determine 12 out of 14 poplar species (Figure 2). In the first step, five species could be identified either by their different length (indel) or by different restriction sites. The remaining species divided into two groups. In the second step, P. tremula (tre) can be identified by a restriction site, and P. tremuloides (tro) and P. grandidentata (gra) remained as one group to be differentiated in the third step. The differentiation of P. deltoides (del) and P. nigra (nig) from the other species of the second remaining group from step one is possible through a direct way to step three, using two different SNPs and the associated restriction enzymes. The last step led to the identification of P. wilsoni (wil) and P. maximowiczii (max) when P. maximowiczii is hetereozygous. Otherwise, P. maximowiczii (max), P. koreana (kor), and P. ussuriensis (uss) are not distinguishable.
Only 14 species are included in this procedure of exclusion. Unfortunately, for the other five species, the necessary sequences are not yet available.
3.2.4. Reconstruction of Genealogy
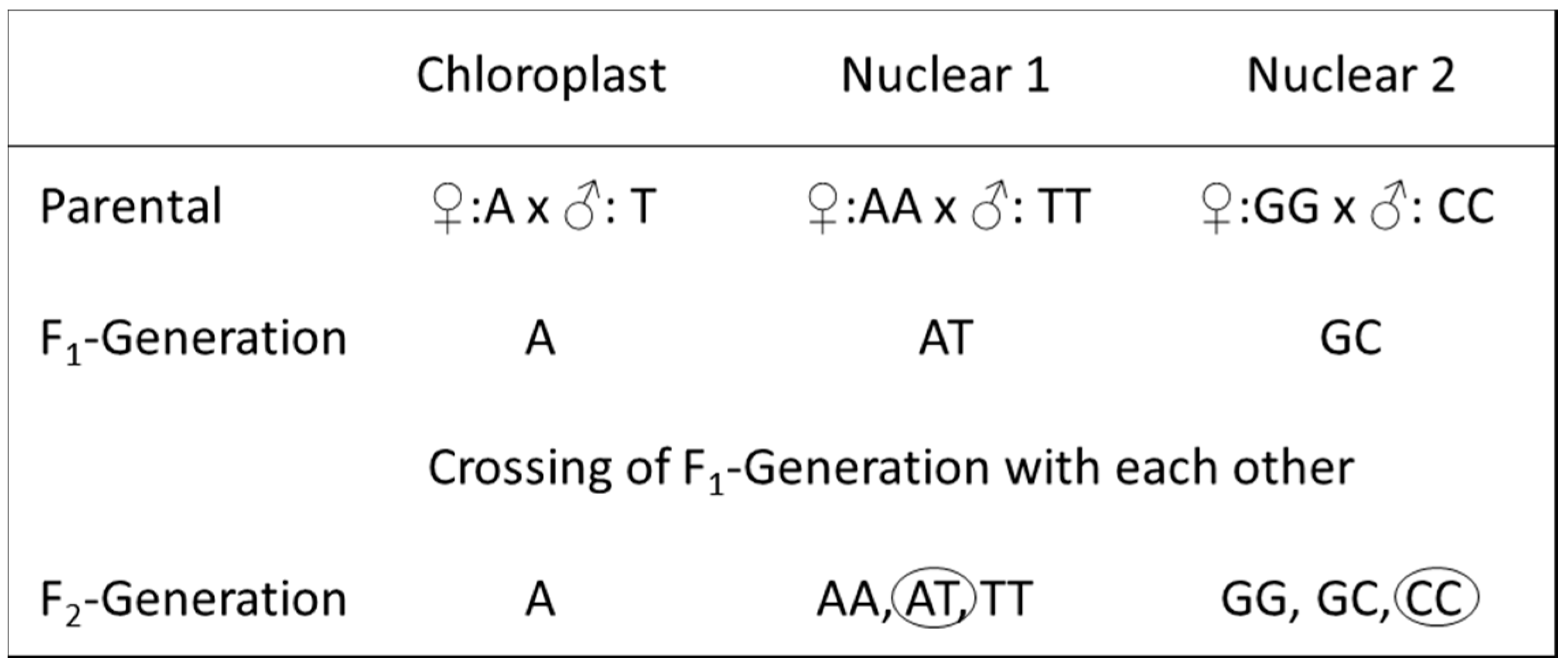
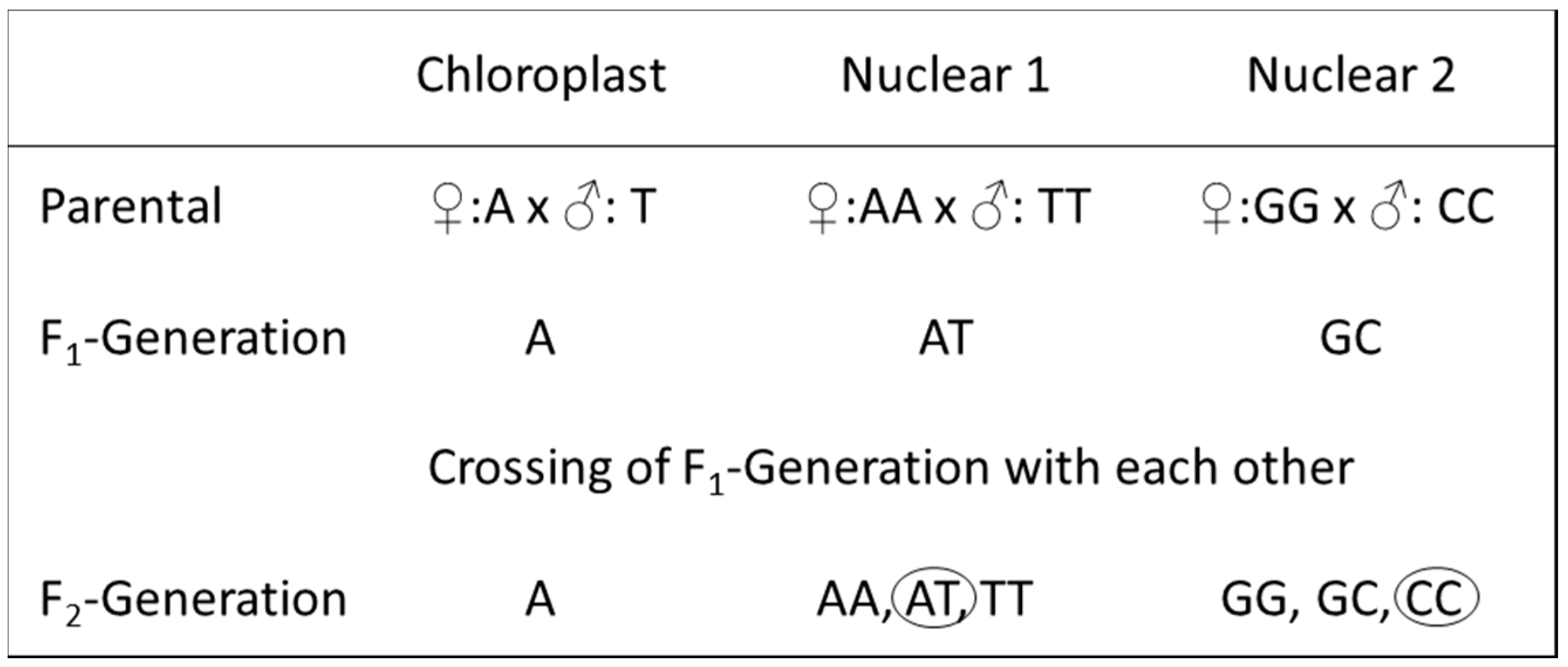
Using markers of the cp genome, the mother and therefore the direction of a crossing can be identified, whereas the molecular markers from the nuclear genomes allow the identification of both crossing partners, but not of the cross direction. For long-used (therefore “old”) poplar hybrids, very often a lack of information becomes evident, e.g., in respect to whether the parents of a used tree are homozygous or already hybrids. To identify the genealogy of such hybrids, a combination of cp and nc markers enables the detection of the preceding generations, thus leading to the present hybrid. Figure 3 gives an example of the combinations of cp and nc markers to reconstruct the genealogy of a hybrid. The cp marker allows the identification of just the mother. Assuming homozygous parents, the descending hybrid would be identifiable by means of heterozygous nuclear markers as being a F1 hybrid. For a deeper look into the genealogy, at least a second nuclear marker is necessary. If one nc marker is heterozygous and a second is homozygous, then it is highly possible that the descending hybrid already stemmed from heterozygous (hybrid) parents; thus, it is at least a F2 generation (Figure 3).
4. Discussion
In this investigation, we combined nine different cp and five nc markers for the identification/differentiation of up to 19 different poplar species (out of the 29 according to Eckenwalder (1996) [2]) belonging to five (out of six) sections within the genus Populus. By application of these 14 genetic markers, we developed protocols (i) to differentiate a poplar species from other species by single SNPs in an unknown sample when at least some reasonable species information is available; (ii) to identify a poplar species by following a species exclusion procedure or by use of a multiplex marker set when no information at all is available; and (iii) to reconstruct the genealogy of long-used (“old”) poplar hybrids.
The identification of poplar species is necessary for breeding activities; nevertheless, most authors concentrate on smaller species complexes, such as Chinese poplars [27,31] or on a smaller number of species to be differentiated [13]. After starting with seven species in 2012 [26] and increasing the number to 14 in 2014 [28], we are now able to identify/differentiate 19 different poplar species. Nevertheless, for five of the investigated species, only one individual was included in the analyses. Thus, the use of the term “species-specific” markers has to be considered provisional for these species. In further investigations, we will aim to increase the number of specimens to prove the usability of the markers for these species.
4.1. Differences between Species
The number of variations varied remarkably between the 19 species, even when the different sample numbers per species—and interestingly, within sections—are taken into account. All of the SNPs and indels were counted as variations that were different from homologous P. trichocarpa sequences. That means that the higher the number of variations, the larger the genetic difference between this species and P. trichocarpa. Looking at the cp genome and comparing only the three species with the highest sample number—P. grandidentata (32), P. tremuloides (21), and P. balsamifera (18)—P. tremuloides has the overall highest number of variations, as well as a remarkable number of species-specific SNPs and indels (Table 2), though it is not the species with the highest number of individuals tested. Whereas P. balsamifera—with its high number of individuals used for validation (18)—is in the group of species with the lowest number of variations, together with: P. fremontii, for which 16 specimens are included in the analysis; P. deltoides, with 14; and P. maximowiczii, with nine tested individuals. These species revealed about only one quarter to one third of the variations of P. tremuloides. As expected, most of these species with low sequence differences to P. trichocarpa belong to the same section as P. trichocarpa. However, two black poplar species (P. deltoides and P. fremontii) also showed low sequence differences to P. trichocarpa. On the other hand, two Asian species (P. simonii with five individuals tested, and P. szechuanica with only one individual sequenced) within the section Tacamahaca revealed a comparable amount of sequence variations to the third black poplar species, P. nigra, and to the other species from the section Populus.
In general, the sequence variations within the cp genome differ within species, genera, and higher systematic taxa. A comprehensive study to test the barcoding regions of the cp genome revealed variations per 100 base pairs between closely related species within different genera and families, from 0.5% for the comparison of two Acorus species, and up to 8.2% difference between two Aethionema species [32]. In our study, the highest difference within the analyzed regions of the cp genome is between P. tremuloides and P. trichcocarpa (0.73%, Table 4).
By analyzing the nuclear genome, the differences between species of the section Populus and P. trichocarpa are far more obvious. Here, we have to keep in mind that for the two species P. adenopoda and P. grandidentata, only the results for two nuclear regions are available. However, when comparing the sequence variations for all five species belonging to the section Populus for only two gene regions (TB1 and KNOX, data not shown), all five species are nearly equally variable. Thus, it is assumed that the evolutionary difference between the species of the section Populus and P. trichocarpa is much higher than between P. trichocarpa and the other sections (e.g., Zhang et al. (2017), Wang et al. (2014) [20,33]). Interestingly, P. maximowiczii and P. deltoides differ much more from P. trichocarpa with regard to the nuclear genome than the cp genome.
A phylogenetic study based on all of the sequence data collected within this study will be published elsewhere later, because this would be beyond the scope of this publication.
4.2. Comparison of Cp and Nc Genomic Regions
As discussed earlier, intergenic spacers have more discrimination power in the genus Populus than the recommended barcoding genes rbcL and matK [26,28]. Two of the intergenic spacers analyzed in this study—trnH_psbA and psbK_psbI—have also been used for the differentiation of poplar species from Western China [27]. In our study, trnH_psbA revealed the highest percent variations summarized over all species (5.2%), followed by trnV_atpE (4.6%), trnC_petN (3.6%), and psbK_psbI (3.2%). In the research study of the Western China species, where six of the species investigated in this study have also been included, psbK_psbI exposed the highest variations (4.22%), but trnH_psbA also showed a high variation (3.66%) compared with our values [27]. In a recent barcoding study, over 40 species from different sections of the genus Quercus have been analyzed using matK, trnH_psbA, and internal transcribed spacer (ITS) sequences [34]. In this study, matK revealed the lowest sequence variation (5.4%), followed by trnH_psbA (8.1%) and the nuclear ITS region (21.6%). Thus, oaks showed overall higher variations than poplars, which is especially true for the nuclear ITS region, whereas the Western Chinese poplars had only 6.54% variation [27]. We did not include the ITS region in our study, but the percent variation per nuclear region is comparable to the poplars from Western China: LEAFY: 9.8%, TB1: 7.7%, GA20ox: 6.7% and KNOX: 4.2%.
Furthermore, in our study, the overall variation per 100 base pairs in the nuclear regions (17.8%) is nearly three times higher than the variation within the cp genome (6.5%), which is comparable to the findings for oaks [34]. The higher overall variation in nuclear genomes can be explained by the different modes of inheritance for cp (uniparental) and nc (biparental) genomes, and the much higher mutation rates in nuclear genomes [35,36].
4.3. Applicability
With the set of different markers proposed in this study, we offer possibilities for a wide range of applications in different equipped laboratories. Single markers for the identification of poplar species can be used with PCR-RFLPs on agarose gels, or as length polymorphisms either on agarose (e.g., for the long indel for the identification of P. tremula) or on polyacrylamide gels for shorter indels (Table 5). Also, the proposed indel marker set can be applied in a laboratory without a genetic analyzer on polyacrylamide gels. Our procedures of exclusion, which are applicable in all laboratories, allow for the identification of up to 16 poplar species. For higher-equipped laboratories, we developed the multiplex kit to be run on a genetic analyzer (e.g., ABI3730 Capillary sequencer, Applied Biosystems, Foster City, CA, USA). Combinations of chloroplast and nuclear markers allow the identification (i) of the crossing direction (maternal part), (ii) of both crossing partners, and therefore (iii) the identification of hybrids, and last but not least (iv) a tracing of the genealogy of single trees of interest.
Many of the poplar species within and also between sections are cross-compatible, and therefore, a high number of natural hybrids exist. This can be most often observed in habitats where two or more poplar species occur sympatric. In Europe, especially the two species of the section Populus—P. alba and P. tremula—naturally hybridize where the distribution areas of both species overlap [37]. This hybrid is called grey poplar (P. × canescens). The preferred cross-direction is P. alba as a female partner with P. tremula as pollen donor, and back-crossing of the hybrids with P. alba [37]. In a recent study, 28 of randomly selected P. × canescens have been investigated, applying both two cp and two nuclear markers. Of these 28 specimens, 26 were revealed to be P. alba × P. tremula, and only two had a cross-direction the other way round of being P. tremula × P. alba (unpublished data). This mating strategy led to a huge number of morphologically different hybrids. Sometimes, hybrids are recognizable as consisting of P. tremula and P. alba, but also, a lot of morphological forms are possible where the parental species are difficult to identify, or the hybrid even just looks like a pure species. The same is true for the introduced hybrid of Eastern cottonwood (P. deltoides) and the European black poplar (P. nigra), which is called P. × canadensis. These hybrids are fertile and cultivated in close vicinity to the relic native Eurasian P. nigra. Furthermore, they are principally able to generate second-generation hybrids (F2) and first generation back-crosses, respectively [38,39]. Also, high variability in leaf morphology was found for these hybrids [40], and thus, species identification just based on morphological characteristics is difficult. These natural hybrids have been used for breeding over decades [3], and are involved in economically used clones. For this reason, the tracing of the genealogy of trees with interesting characteristics for breeders is necessary and possible using the molecular markers presented here.
5. Conclusions
Here, we presented a comprehensive study on the identification of a large number of Populus species with a high number of alternative applications that, to our knowledge, have not yet been performed across such a wide range. By applying combinations of chloroplast and nuclear markers, the identification (i) of the crossing direction (maternal part); (ii) of both crossing partners; and therefore (iii) of hybrids, and last but not least; (iv) tracing the genealogy of single trees of interest, is easily feasible in any lab, no matter how poorly equipped. The sophisticated multiplex marker sets to be used in a genetic analyzer are appropriate when large sample collections have to be analyzed.
Acknowledgments
This project is financially supported by the Federal Ministry of Food, Agriculture and Consumer protection via the “Fachagentur Nachwachsende Rohstoffe e.V.” (FNR). We like to thank the greenhouse staff for harvesting leaves from tall poplar trees, our technical assistants Susanne Bein and Stefanie Palczewski for the laboratory work, and our colleagues Mirko Liesebach and Georg von Wühlisch (both Thuenen-Institute) for information about plants origin. We are grateful to the colleagues from the NW-FVA, further “Fastwood”-partners, as well as Raju Soolanayakanahally (Agriculture and Agri-Food Canada), Minsheng Yang (Agricultural University of Hebei, Baoding, China), Egon Humenberger (University of Minnesota, Department of Forest Resources), and Christian Wehenkel (Instituto de Silvicultura e Industria de la Madera, Universidad Juárez del Estado de Durango, México) providing plant material. We thank the Botanical Gardens Hamburg, Marburg, Tübingen, and Dresden (all Germany) for the supply with poplar reference material. We received no funds for covering the costs to publish in open access.
Author Contributions
M.F. initiated the project; H.S., M.F. and B.K. conceived and designed the experiments; H.S. performed the experiments and coordinated the sample collections; H.S. and M.F. analyzed the data and coordinated the project; H.S. wrote the manuscript and M.F. and B.K. contributed to and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Dickman, D.I.; Stuart, K. The Culture of Poplars in Eastern North America; Department of Forestry, Michigan State University: East Lansing, MI, USA, 1983. [Google Scholar]
- Eckenwalder, J.E. Systematics and evolution of Populus. In Biology of Populus and its Implications for Management and Conservation Part I; Stettler, R.F., Bradshaw, H.D., Jr., Heilman, P.E., Hinckley, T.M., Eds.; NRC Research Press: Ottawa, ON, Canada, 1996; pp. 7–32. [Google Scholar]
- Stettler, R.; Bradshaw, T.; Heilman, P.; Hinckley, T. Biology of Populus and its Implications for Management and Conservation; NRC Research Press: Ottawa, ON, Canada, 1996; p. 542. [Google Scholar]
- Rahman, M.H.; Rajora, O.P. Microsatellite DNA fingerprinting, differentiation, and genetic relationships of clones, cultivars, and varieties of six poplar species from three sections of the genus Populus. Genome 2002, 45, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Rajora, O.P.; Rahman, M.H. Microsatellite DNA and RAPD fingerprinting, identification and genetic relationships of hybrid poplar (Populus × canadensis). Theor. Appl. Genet. 2003, 106, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Orlović, S.; Galović, V.; Zorić, M.; Kovačović, B.; Pilipović, A.; Galić, Z. Evaluation of interspecific DNA variability in poplars using AFLP and SSR markers. Afr. J. Biotechnol. 2009, 8, 5241–5247. [Google Scholar] [CrossRef]
- Schroeder, H.; Fladung, M. SSR and SNP markers for identification of clones, hybrids and species within the genus Populus. Silv. Genet. 2010, 59, 257–263. [Google Scholar] [CrossRef]
- Bruegmann, T.; Fladung, M. Potentials and limitations of the cross-species transfer of nuclear microsatellite marker in six species belonging to three sections of the genus Populus L. Tree Gen. Genom. 2013, 9, 1413–1421. [Google Scholar] [CrossRef]
- Ingvarsson, P.K. Nucleotide polymorphism and linkage disequilibrium within and among natural populations of European aspen (Populus tremula L., Salicaceae). Genetics 2005, 169, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, E.J.; Haughn, G.W.; Ying, C.C.; Otto, S.P.; Zhuang, J.; Cheung, D.; Hamberger, B.; Aboutorabi, F.; Kalynyak, T.; Johnson, L.; et al. Use of Ecotilling as an efficient SNP discovery tool to survey genetic variation in wild populations of Populus trichocarpa. Mol. Ecol. 2006, 15, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Meirmans, P.G.; Lamothe, M.; Périnet, P.; Isabel, N. Species-specific single nucleotide polymorphism markers for detecting hybridization and introgression in poplar. Can. J. Bot. 2007, 85, 1082–1091. [Google Scholar] [CrossRef]
- Fladung, M.; Buschbom, J. Identification of single nucleotide polymorphisms in different Populus species. Trees 2009, 23, 1199–1212. [Google Scholar] [CrossRef]
- Kersten, B.; Voss, M.-M.; Fladung, M. Development of mitochondrial SNP markers to differentiate Populus species. Trees 2015, 29, 575–582. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Okumura, S.; Sawada, S.; Park, Y.W.; Hayashi, T.; Shimamura, M.; Takase, H. Transformation of poplar (Populus alba) plastids and expression of foreign proteins in tree chloroplasts. Transgenic Res. 2006, 15, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.I.; Hefer, C.A.; Kolosova, N.; Douglas, C.J.; Cronk, Q.C.B. Whole plastome sequencing reveals deep plastid divergence and cytonuclear discordance between closely related balsam poplars, Populus balsamifera and P. trichocarpa (Salicaceae). New Phytol. 2014, 204, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.J.; Gao, L.Z. The complete chloroplast genome sequence of desert poplar (Populus euphratica). Mitochondrial DNA Part A 2016, 27, 721–723. [Google Scholar] [CrossRef] [PubMed]
- Kersten, B.; Rampant, P.F.; Mader, M.; Le Paslier, M.-C.; Bounon, R.; Berard, A.; Vettori, C.; Schroeder, H.; Leplé, J.-C.; Fladung, M. Genome sequences of Populus tremula chloroplast and mitochondrium: Implications for holistic poplar breeding. PLoS ONE 2016, 11, e0147209. [Google Scholar] [CrossRef] [PubMed]
- Shang, C.; Du, F.K.; Yin, K.; Zhang, Z. The complete chloroplast genome of Cathay polar: Populus cathayana Rehder. Mitochondrial DNA Part B 2016, 1, 86–87. [Google Scholar] [CrossRef]
- Zhang, L.; Shang, C.; Du, F.K.; Zhao, F.; Ziong, B.; Zhang, Z. Chloroplast phylogenomic analyses maternal relationships among sections in the genus Populus. Biochem. Syst. Ecol. 2017, 70, 132–140. [Google Scholar] [CrossRef]
- Birky, C.W. Uniparental inheritance of mitochondrial and chloroplast genes—Mechanisms and evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 11331–11338. [Google Scholar] [CrossRef] [PubMed]
- Barcode of Life. Available online: http://www.barcodeoflife.org/ (accessed on 16 August 2017).
- Chase, M.W.; Salamin, N.; Wilkinson, M.; Dunwell, J.M.; Kesanakurthi, R.P.; Haidar, N.; Savolainen, V. Land plants and DNA barcodes: Short-term and long-term goals. Philos. Trans. R. Soc. Lond. Ser. B 2005, 360, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Lahaye, R.; Van der Bank, M.; Bogarin, D.; Warner, J.; Pupulin, F.; Gigot, G.; Maurin, O.; Duthoit, S.; Barraclough, T.G.; Savolainen, V. DNA barcoding the floras of biodiversity hotspots. Proc. Natl. Acad. Sci. USA 2008, 105, 2923–2928. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, M.L.; Clark, A.A.; Forrest, L.L.; Richardson, J.; Pennington, R.T.; Long, D.G.; Cowan, R.; Chase, M.W.; Gaudeul, M.; Hollingsworth, P.M. Selecting barcoding loci for plants: Evaluation of seven candidate loci with species-level sampling in three divergent groups of land plants. Mol. Ecol. Resour. 2009, 9, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.; Hoeltken, A.M.; Fladung, M. Differentiation of Populus species using chloroplast SNP-markers—Essential for comprehensible and reliable poplar breeding. Plant Biol. 2012, 14, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Jiang, D.; Shang, H.; Dong, M.; Wang, G.; He, X.; Zhao, C.; Mao, K. Barcoding poplars (Populus L.) from Western China. PLoS ONE 2013, 8, e71710. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.; Fladung, M. Differentiation of Populus species by chloroplast SNP markers for barcoding and breeding approaches. IFOREST 2014, 8, 544–546. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef] [PubMed]
- Dumolin, S.; Demesure, B.; Petit, R.J. Inheritance of chloroplast and mitochondrial genomes in pedunculated oak investigated with efficient PCR method. Theor. Appl. Genet. 1995, 91, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Fan, L.; Milne, R.I.; Zhang, L.; Wang, Y.; Mao, K. Species delimitation and lineage separation history of a species complex of aspen in China. Front. Plant Sci. 2017, 8, 375. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Du, S.; Dayanandan, S.; Wang, D.; Zeng, Y.; Zhang, J. Phylogeny reconstruction and hybrid analysis of Populus (Salicaceae) based on nuclear sequences of multiple single-copy nuclear genes and plastid fragments. PLoS ONE 2014, 9, e103645. [Google Scholar] [CrossRef] [PubMed]
- Simeone, M.C.; Piredda, R.; Papini, A.; Vessella, F.; Schirone, B. Application of plastid and nuclear markers to DNA barcoding of Euro-Mediterranean oaks (Quercus, Fagaceae): Problems, prospects and phylogenetic implications. Bot. J. Linn. Soc. 2013, 172, 478–499. [Google Scholar] [CrossRef]
- Mort, M.E.; Archibald, J.K.; Randle, C.P.; Levsen, N.D.; O’Leary, T.R.; Topalov, K.; Wiegand, C.M.; Crawford, D.J. Inferring phylogeny at low taxonomic levels: Utility of rapidly evolving cpDNA and nuclear ITS loci. Am. J. Bot. 2007, 94, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.J.; Excoffier, L. Gene flow and species delimitation. Trends Ecol. Evol. 2009, 24, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Lexer, C.; Fay, M.F.; Joseph, J.A.; Nica, M.-S.; Heinze, B. Barrier to gene flow between two ecologically divergent Populus species, P. alba (white poplar) and P. tremula (European aspen): The role of ecology and life history in gene introgression. Mol. Ecol. 2005, 14, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, H.; Ceulemans, R.; Davis, J.; Stettler, R. Emerging model systems in plant biology: Poplar (Populus) as a model forest tree. J. Plant Growth Regul. 2000, 19, 306–313. [Google Scholar] [CrossRef]
- Vanden Broeck, A.; Cox, K.; Quataert, P.; Van Bockstaele, E.; Van Slycken, J. Flowering phenology of Populus nigra L., P. nigra cv. italica and P. × canadensis Moench. And the potential for natural hybridisation in Belgium. Silv. Genet. 2003, 52, 280–283. [Google Scholar]
- DeWoody, J.; Trewin, H.; Taylor, G. Genetic and morphological differentiation in Populus nigra L.: Isolation by colonization or isolation by adaptation? Mol. Ecol. 2015, 24, 2641–2655. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Procedure of exclusion for four cp indels: a step-by-step procedure for the use of indels from four cp regions to differentiate up to 16 of the 19 poplar species. For the fourth step, if P. simonii shows a fragment length of 445 bp and P. lasiocarpa shows a fragment length of 441 bp, then the differentiation of these two species is possible. If the individuals of both species revealed a fragment length of 443 bp, then the species were not differentiated. Abbreviations: aden = P. adenopoda, alb = P. alba, bal = P. balsamifera, cath = P. cathayana, del = P. deltoides, eup = P. euphratica, fre = P. fremontii, gra = P. grandidentata, kor = P. koreana, max = P. maximowicziix, nig = P. nigra, sim = P. simonii, sze = P. szechuanica, tre = P. tremula, tri = P. trichocarpa, tro = P. tremuloides, uss = P. ussuriensis, wil = P. wilsonii.
Figure 1.
Procedure of exclusion for four cp indels: a step-by-step procedure for the use of indels from four cp regions to differentiate up to 16 of the 19 poplar species. For the fourth step, if P. simonii shows a fragment length of 445 bp and P. lasiocarpa shows a fragment length of 441 bp, then the differentiation of these two species is possible. If the individuals of both species revealed a fragment length of 443 bp, then the species were not differentiated. Abbreviations: aden = P. adenopoda, alb = P. alba, bal = P. balsamifera, cath = P. cathayana, del = P. deltoides, eup = P. euphratica, fre = P. fremontii, gra = P. grandidentata, kor = P. koreana, max = P. maximowicziix, nig = P. nigra, sim = P. simonii, sze = P. szechuanica, tre = P. tremula, tri = P. trichocarpa, tro = P. tremuloides, uss = P. ussuriensis, wil = P. wilsonii.

Figure 2.
Procedure of exclusion for nuclear regions: a step-by-step procedure for the use of four nuclear regions to differentiate up to 12 of 14 poplar species. For the fourth step, P. maximowiczii could be homozygous, and therefore remain uncut. Then, it either cannot be differentiated from P. koreana and P. ussuriensis, or alternatively, P. maximowiczii can be hetereozygous, in which case the differentiation from the other two species is possible. Abbreviations and explanation for the species: see the legend of Figure 1 and the main text.
Figure 2.
Procedure of exclusion for nuclear regions: a step-by-step procedure for the use of four nuclear regions to differentiate up to 12 of 14 poplar species. For the fourth step, P. maximowiczii could be homozygous, and therefore remain uncut. Then, it either cannot be differentiated from P. koreana and P. ussuriensis, or alternatively, P. maximowiczii can be hetereozygous, in which case the differentiation from the other two species is possible. Abbreviations and explanation for the species: see the legend of Figure 1 and the main text.
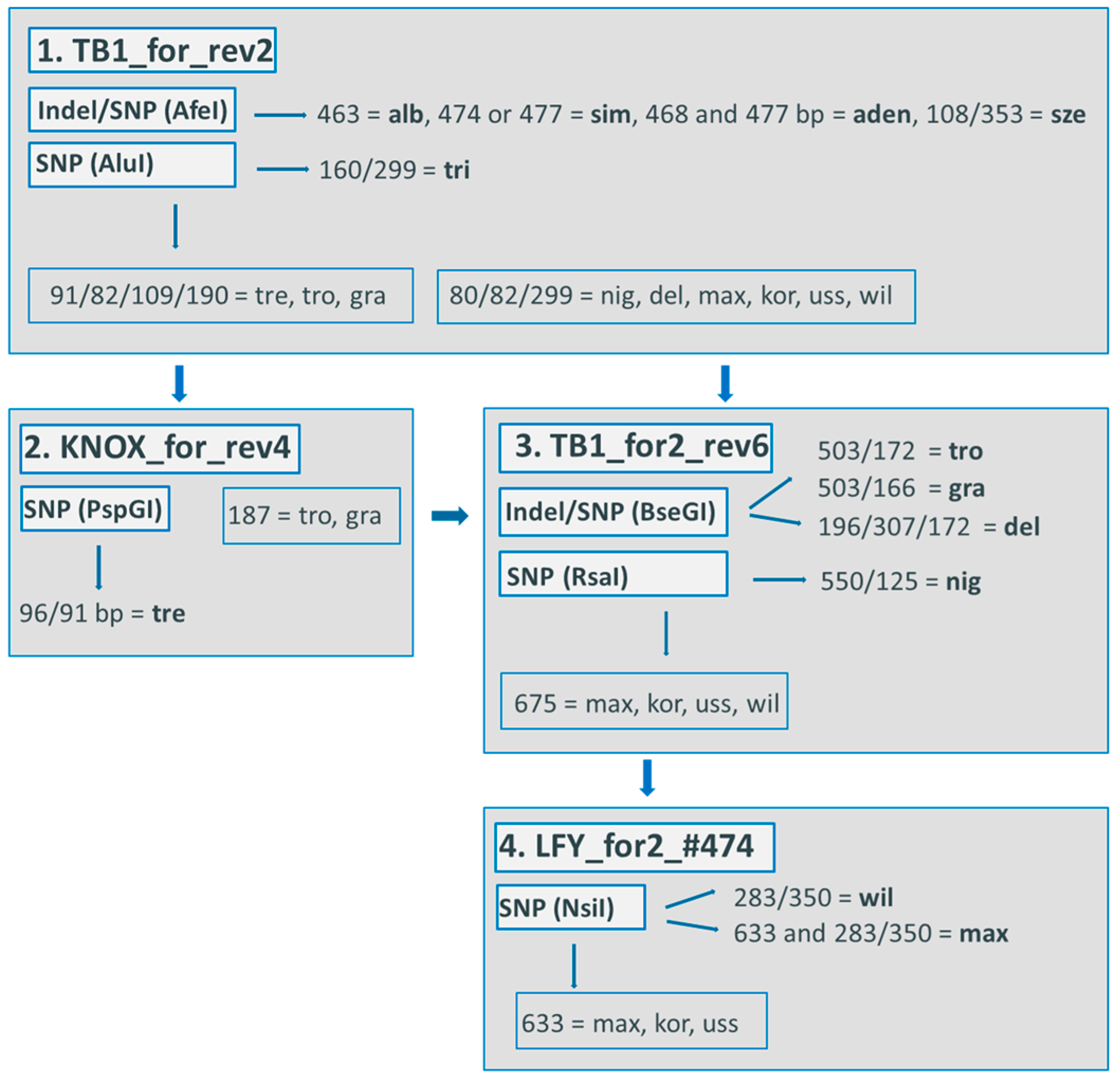
Figure 3.
Scheme of a hypothetical genealogy. Presentation of the genealogy of a hybrid by using two different parental species, both homozygous in two different hypothetical markers. Circles in the row “F2-Generation” show a possible combination of two markers, one heterozygous and one homozygous, allowing the identification of the hybrid as a F2 generation. For details, see main text.
Figure 3.
Scheme of a hypothetical genealogy. Presentation of the genealogy of a hybrid by using two different parental species, both homozygous in two different hypothetical markers. Circles in the row “F2-Generation” show a possible combination of two markers, one heterozygous and one homozygous, allowing the identification of the hybrid as a F2 generation. For details, see main text.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Chloroplast (cp) intergenic spacers and investigated nuclear regions. Overview of the primers and PCR conditions for chloroplast and nuclear regions adopted for species differentiation in poplar. The column ‘length’ gives the fragment length in base pairs for P. trichocarpa. Abbreviations in primer names: f, for = forward; r, rev = reverse. Ta = annealing temperature.
Table 1.
Chloroplast (cp) intergenic spacers and investigated nuclear regions. Overview of the primers and PCR conditions for chloroplast and nuclear regions adopted for species differentiation in poplar. The column ‘length’ gives the fragment length in base pairs for P. trichocarpa. Abbreviations in primer names: f, for = forward; r, rev = reverse. Ta = annealing temperature.
| Region | Primer Name | Sequence (5′-3′) or Reference | Length | Ta (°C) |
|---|---|---|---|---|
| Chloroplast | ||||
| trnH_psbA | trnHf | [29] | ||
| psbAr | [29] | 367 | 58 | |
| psbK_psbI | 4a_f | [26] | ||
| 4b_r | [26] | 434 | 55 | |
| rpoC2_rpoC1 | 8a_f | [26] | ||
| 8b_r | [26] | 533 | 60 | |
| trnV_atpE | 12a_f | [26] | ||
| 12b_r | [26] | 740 | 58 | |
| psbA_matK | 1a_f | [26] | ||
| 1b_r | [26] | 866 | 58 | |
| rps4_trnL | rps4f | GAAACGAGGCCCTCGGTAACGTG | ||
| trnLr | GTCCGTAGCGTCTACCAATTTCG | 821 | 60 | |
| rps2_rpoC2 | 7a_f | [26] | ||
| 7b_r | [26] | 524 | 58 | |
| trnC_petN | trnCf | CCAGTTCAAATCCGGGTGTCGC | ||
| petNr | GCAGCCCAAGCGAGACTTACTAT | 444 | 58 | |
| psaA_ycf13 | psaAf | GTGGTAGTATCAGGTCCTTTAGCT | ||
| ycf13r | GAAGCGCAGAATTGGAAGATC | 782 | 58 | |
| psbE_petL | indel3_f | TTCTTTGAATGAAGTTATACAACAC | ||
| indel3_r | GATACGAAGATAAGCAAAAATACC | 283 | 55 | |
| Nuclear | ||||
| GA20ox | #471_f | ATAGATTGCATCAAAACC | ||
| #743_r | TTAATTACCAGATCTATGGACATAAAT | 1688 | 52 | |
| GA20_for4 | TAATGAGGCTCAACTACTACC | |||
| GA20_rev5 | CTTACCATGAAGGTGTCACC | 202 | 54 | |
| KNOX | KNOX_int2for4 | TGGAGGACTACAATCAAATGAG | ||
| KNOX3_int2rev2 | TGAGATCATTACAGGCGTG | 982 | 52 | |
| KNOX3_int2f | CCAAACAATTAACCAGCAGC | |||
| KNOX3_int2rev4 | GCTACTGAAATGAATTTAATTTGA | 187 | 52 | |
| LEAFY | LFY_for2 | TACTCTTGATGCTCTCTCCCAAG | ||
| LFY_rev2 | CTTGAACTAAATTCCATATATAAG | 263 | 53 | |
| #474_r | TTGCTGTACTGGCTCCTCAGA | 633 | 53 | |
| TB1 | TB1_f | CTAACCATACCATGATTGCA | ||
| TB1_rev2 | TATGGATCTTGCTGTGCCT | 461 | 53 | |
| TB1_rev6 | GGTCATGAGAATTGAAATGGAG | 1369 | 53 | |
| TB1_for2 (combined rev6) | GTTTCAGGCATCAAGCTG | 675 | 53 |
Table 2.
Number of single nucleotide polymorphisms (SNPs) and insertion–deletion polymorphisms (indels) in the cp regions. Overview of the total number of found SNPs and indels, species-specific SNPs and indels, and related restrictions enzymes for the species-specific SNPs for use in PCR-RFLPs (Restriction Fragment Length Polymorphisms) in nine chloroplast regions (without psbE_petL). All of the nucleotide variations that were different from P. trichocarpa were counted as SNPs/indels. The number of individuals per species used for marker validation is also given.
Table 2.
Number of single nucleotide polymorphisms (SNPs) and insertion–deletion polymorphisms (indels) in the cp regions. Overview of the total number of found SNPs and indels, species-specific SNPs and indels, and related restrictions enzymes for the species-specific SNPs for use in PCR-RFLPs (Restriction Fragment Length Polymorphisms) in nine chloroplast regions (without psbE_petL). All of the nucleotide variations that were different from P. trichocarpa were counted as SNPs/indels. The number of individuals per species used for marker validation is also given.
| Species | SNPs/Indels | Species-Specific SNPs/Indels | Restriction Enzymes | [N] Number of Individuals |
|---|---|---|---|---|
| P. alba a | 22/3 | 2/2 | 2 | 5–13 |
| P. tremula a | 28/6 | 9/3 | 1 | 5 |
| P. tremuloides a | 40/9 | 17/7 | 4 | 5–21 |
| P. adenopoda a (P. × tomentosa) | 19/5 | 1/3 | 1 | 5 |
| P. grandidentata a | 26/12 | 4/7 | 3 | 32 |
| P. nigra b | 28/5 | 5/2 | 2 | 5–12 |
| P. deltoides b | 12/3 | 2/1 | 0 | 5–14 |
| P. fremontii b | 14/7 | 3/5 | 1 | 5–16 |
| P. trichocarpa c | - | 4/1 | 1 | 5 |
| P. maximowiczii c | 13/4 | 0/2 | 0 | 3–9 |
| P. cathayana c | 12/3 | 0/0 | 0 | 1 |
| P. koreana c | 12/3 | 0/0 | 0 | 3 |
| P. ussuriensis c | 12/3 | 0/0 | 0 | 1 |
| P. simonii c | 25/2 | 1/0 | 0 | 5 |
| P. szechuanica c | 24/3 | 0/0 | 0 | 1 |
| P. balsamifera c | 11/5 | 1/2 | 0 | 18 |
| P. wilsonii d | 17/2 | 3/2 | 0 | 1 |
| P. lasiocarpa d | 21/3 | 0/0 | 0 | 2 |
| P. euphratica e | 22/7 | 7/4 | 0 | 1 |
a These species belong to the section Populus. b These species belong to the section Aigeiros. c These species belong to the section Tacamahaca. d These species belong to the section Leucoides. e This species belongs to the section Turanga.
Table 3.
Number of SNPs and indels in the nuclear regions analyzed in this study. Overview of the total number of found SNPs and indels, species-specific SNPs and indels, and related restrictions enzymes for the species-specific SNPs for use in PCR-RFLPs in the four nuclear regions. All of the SNPs/indels counted were nucleotide variations that were different from P. trichocarpa. The number of individuals per species used for marker validation is also given.
Table 3.
Number of SNPs and indels in the nuclear regions analyzed in this study. Overview of the total number of found SNPs and indels, species-specific SNPs and indels, and related restrictions enzymes for the species-specific SNPs for use in PCR-RFLPs in the four nuclear regions. All of the SNPs/indels counted were nucleotide variations that were different from P. trichocarpa. The number of individuals per species used for marker validation is also given.
| Species | SNPs/Indels | Species-Specific SNPs/Indels | Restriction Enzymes | [N] Number of Individuals |
|---|---|---|---|---|
| P. alba a | 119/14 | 13/4 | 2 | 6 |
| P. tremula a | 107/12 | 8/0 | 1 | 5 |
| P. tremuloides a | 113/10 | 8/0 | 0 | 4–6 |
| P. adenopoda 2,a (P. × tomentosa) | 35/3 | 1/0 | 0 | 3–5 |
| P. grandidentata 2,a | 36/4 | 3/2 | 1 | 5–32 |
| P. nigra b | 53/12 | 17/1 | 4 | 4–6 |
| P. deltoide s b | 48/11 | 18/0 | 3 | 4–5 |
| P. fremontii 1,b | 23/0 | 6/0 | 0 | 10 |
| P. trichocarpa c | - | 7/0 | 2 | 5–7 |
| P. maximowiczii c | 52/11 | 4/0 | 0 | 4–6 |
| P. cathayana c | 17/0 | 0/0 | 0 | 1 |
| P. koreana c | 36/6 | 1/0 | 0 | 3 |
| P. ussuriensis c | 28/6 | 0/0 | 0 | 1 |
| P. simonii c | 49/10 | 7/3 | 2 | 3 |
| P. szechuanica c | 32/4 | 2/0 | 1 | 1 |
| P. balsamifera c | 27/7 | 1/0 | 0 | 3–18 |
| P. wilsonii d | 35/4 | 3/0 | 1 | 1 |
| P. lasiocarpa 2,d | 9/0 | 1/0 | 0 | 2 |
| P. euphratica 2,e | 14/2 | 6/1 | 2 | 1 |
1 Only data for KNOX included. 2 Only data for KNOX and TB1 included. a These species belong to the section Populus. b These species belong to the section Aigeiros. c These species belong to the section Tacamahaca. d These species belong to the section Leucoides. e This species belongs to the section Turanga.
Table 4.
Calculation of percent variations. Variations per 100 base pairs (% variation) calculated for the chloroplast and nuclear genome regions, as given in Table 1 (without psbE_petL, because it has not been sequenced in all of the species), for the 19 poplar species. cp = chloroplast, nc = nuclear.
Table 4.
Calculation of percent variations. Variations per 100 base pairs (% variation) calculated for the chloroplast and nuclear genome regions, as given in Table 1 (without psbE_petL, because it has not been sequenced in all of the species), for the 19 poplar species. cp = chloroplast, nc = nuclear.
| Species | Overall cp Variations | Species-Specific cp Variations | Overall nc Variations | Species-Specific nc Variations | ||||
|---|---|---|---|---|---|---|---|---|
| SNP | Indel | SNP | Indel | SNP | Indel | SNP | Indel | |
| P. alba a | 0.40 | 0.05 | 0.04 | 0.04 | 2.54 | 0.30 | 0.28 | 0.09 |
| P. tremula a | 0.51 | 0.11 | 0.16 | 0.05 | 2.29 | 0.26 | 0.17 | 0.00 |
| P. tremuloides a | 0.73 | 0.16 | 0.31 | 0.13 | 2.42 | 0.21 | 0.17 | 0.00 |
| P. adenopoda 2,a (P. × tomentosa) | 0.34 | 0.09 | 0.02 | 0.05 | 0.75 | 0.06 | 0.02 | 0.00 |
| P. grandidentata 2,a | 0.47 | 0.22 | 0.07 | 0.13 | 0.77 | 0.09 | 0.06 | 0.04 |
| P. nigra b | 0.51 | 0.09 | 0.09 | 0.04 | 1.13 | 0.26 | 0.36 | 0.02 |
| P. deltoids b | 0.22 | 0.05 | 0.04 | 0.02 | 1.03 | 0.24 | 0.38 | 0.00 |
| P. fremontii 1,b | 0.25 | 0.13 | 0.05 | 0.09 | 0.49 | 0.00 | 0.13 | 0.00 |
| P. trichocarpa c | - | - | 0.07 | 0.02 | - | - | 0.15 | 0.00 |
| P. maximowiczii c | 0.24 | 0.07 | 0.00 | 0.04 | 1.11 | 0.24 | 0.09 | 0.00 |
| P. cathayana c | 0.22 | 0.05 | 0.00 | 0.00 | 0.36 | 0.00 | 0.00 | 0.00 |
| P. koreana c | 0.22 | 0.05 | 0.00 | 0.00 | 0.77 | 0.13 | 0.02 | 0.00 |
| P. ussuriensis c | 0.22 | 0.05 | 0.00 | 0.00 | 0.60 | 0.13 | 0.00 | 0.00 |
| P. simonii c | 0.45 | 0.04 | 0.02 | 0.00 | 1.05 | 0.21 | 0.15 | 0.06 |
| P. szechuanica c | 0.44 | 0.05 | 0.00 | 0.00 | 0.68 | 0.09 | 0.04 | 0.00 |
| P. balsamifera c | 0.20 | 0.09 | 0.02 | 0.04 | 0.58 | 0.15 | 0.02 | 0.00 |
| P. wilsonii d | 0.31 | 0.04 | 0.05 | 0.04 | 0.75 | 0.09 | 0.06 | 0.00 |
| P. lasiocarpa 2,d | 0.38 | 0.05 | 0.00 | 0.00 | 0.19 | 0.00 | 0.02 | 0.00 |
| P. euphratica 2,e | 0.40 | 0.13 | 0.13 | 0.07 | 0.30 | 0.04 | 0.13 | 0.02 |
| Average (species) | 0.36 | 0.09 | 0.06 | 0.04 | 0.99 | 0.14 | 0.12 | 0.01 |
| Sum over regions and species | 6.50 | 1.54 | 1.07 | 0.74 | 17.81 | 2.48 | 2.27 | 0.24 |
1 Only data for KNOX included. 2 Only data for KNOX and TB1 included. a These species belong to the section Populus. b These species belong to the section Aigeiros. c These species belong to the section Tacamahaca. d These species belong to the section Leucoides. e This species belongs to the section Turanga.
Table 5.
Applicable SNP and indel markers. Selection of easy-to-use SNP and indel markers for the identification of 14 poplar species. The column “source” contained the information if the markers are already published or newly designed, and first presented in this study (new).
Table 5.
Applicable SNP and indel markers. Selection of easy-to-use SNP and indel markers for the identification of 14 poplar species. The column “source” contained the information if the markers are already published or newly designed, and first presented in this study (new).
| Species | Region | Marker Type | Restriction Enzyme | Length [bp] | Source |
|---|---|---|---|---|---|
| P. alba | trnH_psbA | cp SNP | Alw61I | 139/228 | [26] |
| P. tremula | rpoC2_rpoC1 psbE_petL | cp SNP indel | MlsI / | 409/124 283 (−99) | [26] new |
| P. tremuloides | trnH_psbA | cp SNP | DraI | 129/238 | [26] |
| P. adenopoda | psbK_psbI | cp SNP | AseI | 144/290 | new |
| P. grandidentata | psbK_psbI | cp SNP + indel | DraI | 264/174 | new |
| P. nigra | trnV_atpE | cp SNP + indel | ApoI | 404/62/260 | new |
| P. deltoides | TB1 | nc SNP | BseGI | 194/481 | new |
| P. fremontii | trnH_psbA | cp SNP | BsmI | 102/265 | new |
| P. trichocarpa | psbK_psbI | cp SNP | TaqI | 78/209/49/43/214 | new |
| P. maximowiczii | psaA_ycf13 | cp indel | / | (ca. 790) + 22 | new |
| P. simonii | GA20ox | nc SNP | CviKI | 8/27/167 | new |
| P. szechuanica | TB1 | nc SNP | AfeI | 108/353 | new |
| P. wilsonii | trnH_psbA | cp indel | / | (367) + 4 | new |
| P. euphratica | KNOX_int2 | nc SNP | BfaI | 64/223 | new |
Table 6.
Multiplex PCR for the amplification of four cp regions to be analyzed on the ABI3730 Genetic Analyzer.
Table 6.
Multiplex PCR for the amplification of four cp regions to be analyzed on the ABI3730 Genetic Analyzer.
| PCR 1 | Concentration | PCR 2 | Concentration |
|---|---|---|---|
| BD PCR-Puffer 10× | 1× | BD PCR-Puffer 10× | 1× |
| MgCl 25 mM | 1.5 mM | MgCl 25 mM | 1.75 mM |
| dNTPs 25 mM | 0.2 mM | dNTPs 25 mM | 0.2 mM |
| rps4_2_VIC 10 μM | 0.18 μM | psbA_2_PET 10 μM | 0.2 μM |
| trnL_2_R 10 μM | 0.18 μM | matK_2_R 10 μM | 0.2 μM |
| rps2-mod_FAM 10 μM | 0.12 μM | Enhancer DMSO | |
| rpoc2_R 10 μM | 0.12 μM | Taq 5 U/μL | 0.6 U |
| trnC_FAM 10 μM | 0.04 μM | DNA 10 ng/μL | 30 ng |
| petN_R 10 μM | 0.04 μM | ||
| Enhancer DMSO | |||
| Taq 5 U/μL | 0.6 U | ||
| DNA 10 ng/μL | 30 ng |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Schroeder, H.; Kersten, B.; Fladung, M. Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding. Forests 2017, 8, 492. https://doi.org/10.3390/f8120492
AMA Style
Schroeder H, Kersten B, Fladung M. Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding. Forests. 2017; 8(12):492. https://doi.org/10.3390/f8120492
Chicago/Turabian StyleSchroeder, Hilke, Birgit Kersten, and Matthias Fladung. 2017. "Development of Multiplexed Marker Sets to Identify the Most Relevant Poplar Species for Breeding" Forests 8, no. 12: 492. https://doi.org/10.3390/f8120492
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.