Temporal Variation of Ecological Factors Affecting Bird Species Richness in Urban and Peri-Urban Forests in a Changing Environment: A Case Study from Milan (Northern Italy)



Abstract
:1. Introduction
2. Materials and Methods
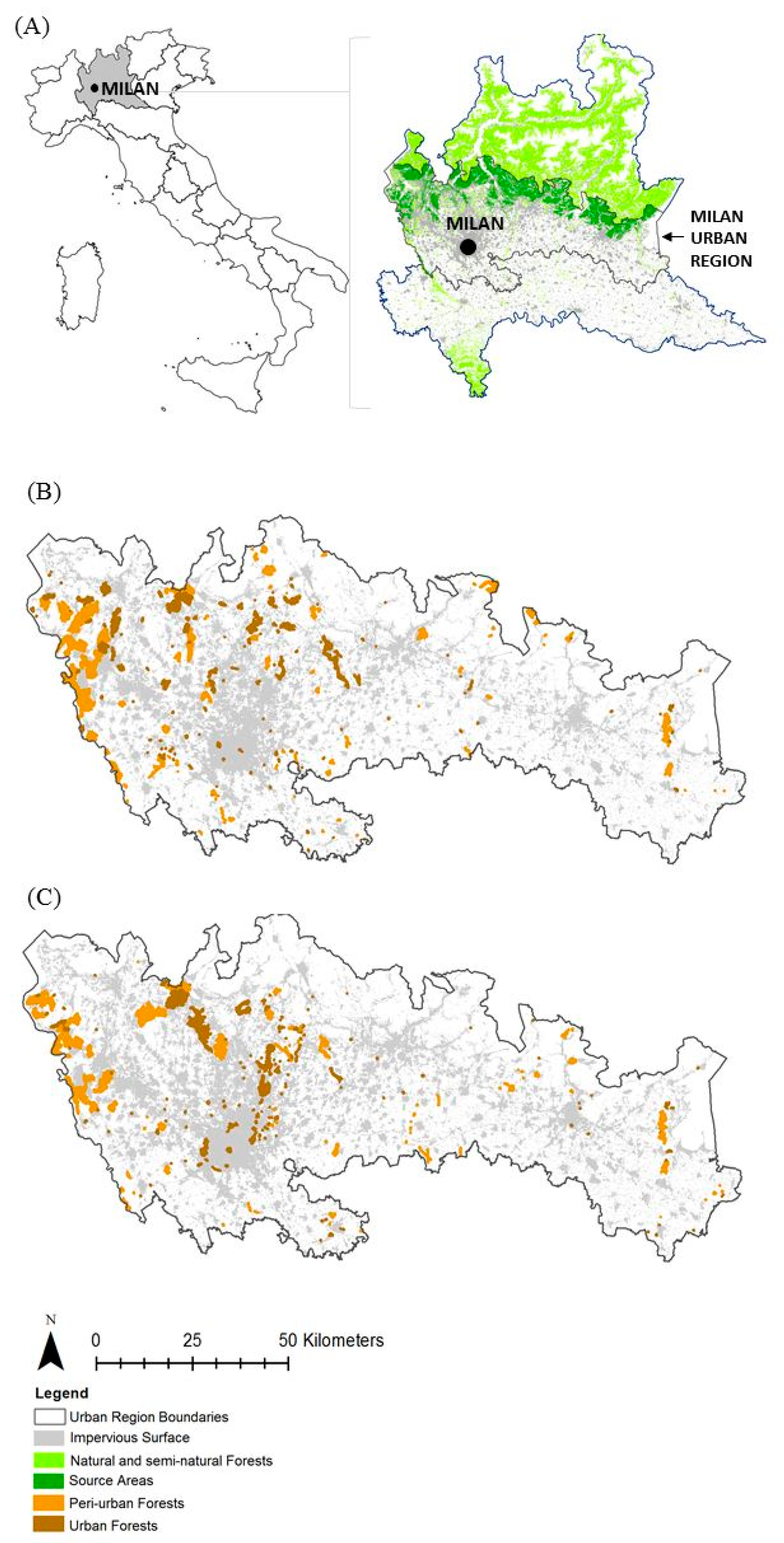
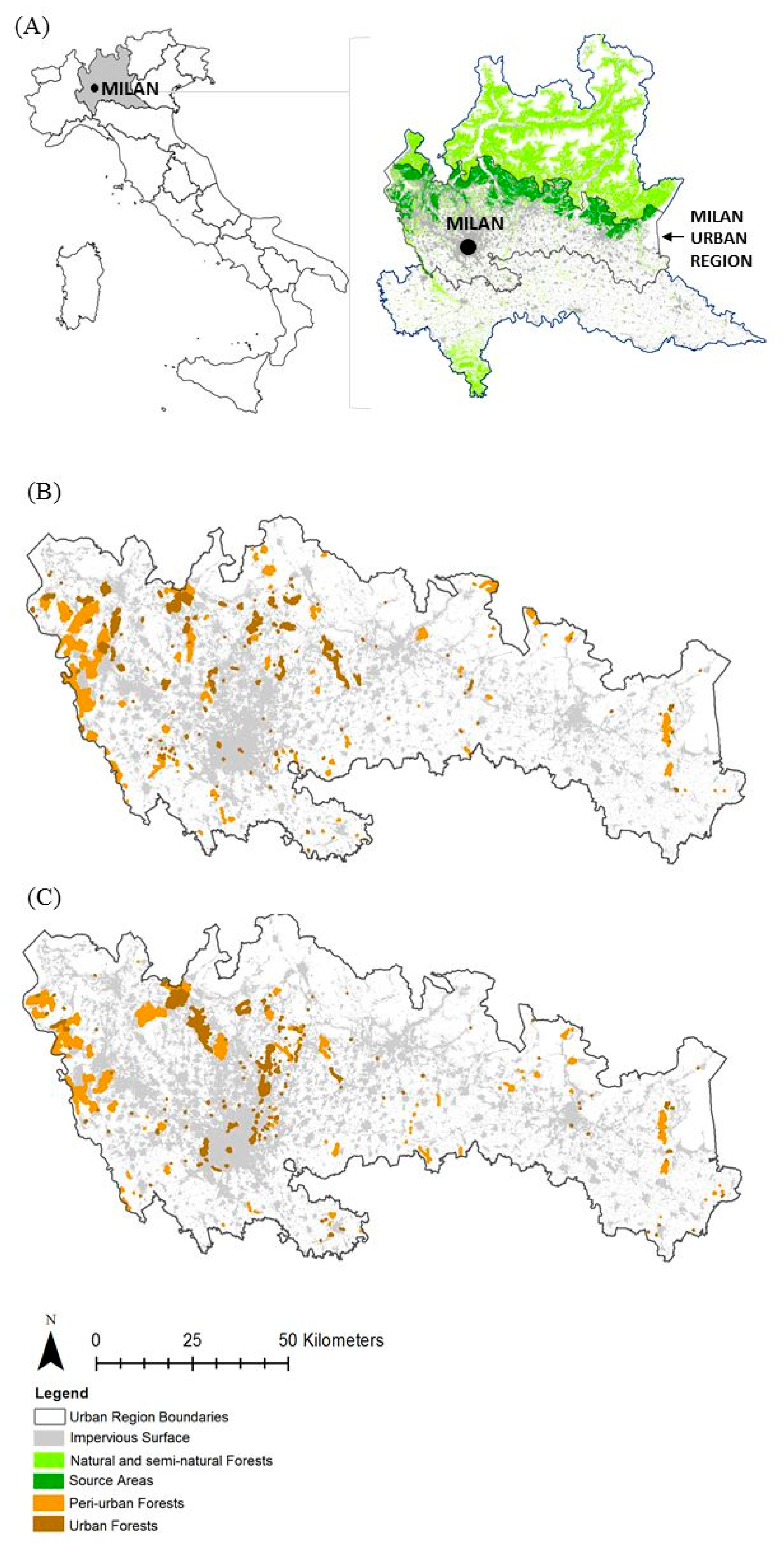
2.1. Study Area
2.2. Bird Data
2.3. Landscape Data
2.4. Statistical Analysis
3. Results
3.1. Forest Bird Richness
3.2. Models
4. Discussion
4.1. Factors Affecting Bird Species Richness in UPUFs
4.2. Temporal Changes of the Effect of UPUFs Characteristics on Bird Richness
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Haase, D.; Frantzeskaki, N.; Elmqvist, T. Ecosystem services in urban landscapes: Practical applications and governance implications. Ambio 2014, 43, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Wratten, S.; Sandhu, H.; Cullen, R.; Costanza, R. Ecosystem Services in Agricultural and Urban Landscapes; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Haines-Young, R.; Potschin, M. The links between biodiversity, ecosystem services and human well-being. In Ecosystem Ecology: A New Synthesis; Raffaelli, D.G., Frid, C.L., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 110–139. [Google Scholar]
- Lafortezza, R.; Coomes, D.A.; Kapos, V.; Ewers, R.M. Assessing the impacts of fragmentation on plant communities in New Zealand: Scaling from survey plots to landscapes. Glob. Ecol. Biogeogr. 2010, 19, 741–754. [Google Scholar] [CrossRef]
- Scolozzi, R.; Morri, E.; Santolini, R. Delphi-based change assessment in ecosystem service values to support strategic spatial planning in Italian landscapes. Ecol. Indic. 2012, 21, 134–144. [Google Scholar] [CrossRef]
- Geneletti, D. Using spatial indicators and value functions to assess ecosystem fragmentation caused by linear infrastructures. Int. J. Appl. Earth Obs. Geoinf. 2004, 5, 1–15. [Google Scholar] [CrossRef]
- Lafortezza, R.; Brown, R.D. A framework for landscape ecological design of new patches in the rural landscape. Environ. Manag. 2004, 34, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Hladnik, D.; Pirnat, J. Urban forestry-Linking naturalness and amenity: The case of Ljubljana, Slovenia. Urban For. Urban Green. 2011, 10, 105–112. [Google Scholar] [CrossRef]
- Sanesi, G.; Gallis, C.; Kasperidus, H.D. Urban forests and their ecosystem services in relation to human health. In Forests, Trees and Human Health; Nilsson, K., Sangster, M., Konijnendijk, C.C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 23–40. [Google Scholar]
- Angold, P.G.; Sadler, J.P.; Hill, M.O.; Pullin, A.; Rushton, S.; Austin, K.; Small, E.; Wood, B.; Wadsworth, R.; Sanderson, R.; et al. Biodiversity in urban habitat patches. Sci. Total Environ. 2006, 360, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Park, C.R.; Lee, W.S. Relationship between species composition and area in breeding birds of urban woods in Seoul, Korea. Landsc. Urban Plan. 2000, 51, 29–36. [Google Scholar] [CrossRef]
- Mörtberg, U.; Wallentinus, H.G. Red-listed forest bird species in an urban environment assessment of green space corridors. Landsc. Urban Plan. 2000, 50, 215–226. [Google Scholar] [CrossRef]
- Sandström, U.G.; Angelstam, P.; Mikusiski, G. Ecological diversity of birds in relation to the structure of urban green space. Landsc. Urban Plan. 2006, 77, 39–53. [Google Scholar] [CrossRef]
- Bolger, D.T.; Scott, T.A.; Rotenberry, J.T. Use of corridor-like landscape structures by bird and small mammal species. Biol. Conserv. 2001, 102, 213–224. [Google Scholar] [CrossRef]
- Elmqvist, T.; Setälä, H.; Handel, S.N.; Van Der Ploeg, S.; Aronson, J.; Blignaut, J.N.; Gómez-Baggethun, E.; Nowak, D.J.; Kronenberg, J.; De Groot, R. Benefits of restoring ecosystem services in urban areas. Curr. Opin. Environ. Sustain. 2015, 14, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Strohbach, M.; Haase, D.; Kabisch, N. Birds and the city: Urban biodiversity, land use, and socioeconomics. Ecol. Soc. 2009, 14, 31. [Google Scholar] [CrossRef]
- Mcdonald, R.I.; Forman, R.T.; Kareiva, P.; Neugarten, R.; Salzer, D.; Fisher, J. Urban effects, distance, and protected areas in an urbanizing world. Landsc. Urban Plan. 2009, 93, 63–75. [Google Scholar] [CrossRef]
- Donnelly, R.; Marzluff, J.M. Relative importance of habitat quantity, structure, and spatial pattern to birds in urbanizing environments. Urban Ecosyst. 2006, 9, 99–117. [Google Scholar] [CrossRef]
- Sekercioglu, C.H. Increasing awareness of avian ecological function. Trends Ecol. Evol. 2006, 21, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Forman, R.T. Urban Regions: Ecology and Planning beyond the City; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Canedoli, C.; Crocco, F.; Comolli, R.; Padoa-Schioppa, E. Landscape fragmentation and urban sprawl in the urban region of Milan. Landsc. Res. 2017, 1–20. [Google Scholar] [CrossRef]
- Bani, L.; Massimino, D.; Orioli, V.; Bottoni, L.; Massa, R. Assessment of population trends of common breeding birds in Lombardy, Northern Italy, 1992–2007. Ethol. Ecol. Evol. 2009, 21, 27–44. [Google Scholar] [CrossRef]
- Blondel, J.; Ferry, C.; Frochor, B. Point counts with unlimited distance. Stud. Avian Biol. 1981, 6, 414–420. [Google Scholar]
- Fornasari, L.; Bani, L.; de Carli, E.; Massa, R. Optimum design in monitoring common birds and their habitat. Gibier Faune Sauvag. 1998, 15, 309–322. [Google Scholar]
- Ente Regionale per i Servizi all’Agricoltura e alle Foreste (ERSAF). Destinazione d’Uso dei Suoli Agricoli e Forestali (DUSAF 1.1); Ente Regionale per i Servizi all’Agricoltura e alle Foreste della Lombardia: Milano, Italy, 2002. [Google Scholar]
- Ente Regionale per i Servizi all’Agricoltura e alle Foreste (ERSAF). Destinazione d’Uso dei Suoli Agricoli e Forestali (DUSAF 4); Ente Regionale per i Servizi all’Agricoltura e alle Foreste della Lombardia: Milano, Italy, 2014. [Google Scholar]
- Dondina, O.; Orioli, V.; D’Occhio, P.; Luppi, M.; Bani, L. How does forest species specialization affect the application of the island biogeography theory in fragmented landscapes? J. Biogeogr. 2017, 44, 1041–1052. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute (ESRI). ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Massimino, D.; Orioli, V.; Pizzardi, F.; Massa, R.; Bani, L. Usefulness of coarse grain data on forest management to improve bird abundance models. Ital. J. Zool. 2010, 77, 71–80. [Google Scholar] [CrossRef]
- Cameron, A.C.; Trivedi, P.K. Regression-based Tests for Overdispersion in the Poisson Model. J. Econom. 1990, 46, 347–364. [Google Scholar] [CrossRef]
- Kleiber, C.; Zeileis, A. Applied Econometrics with R; Springer: New York, NY, USA, 2008; Available online: http://CRAN.R-project.org/package=AER (accessed on 1 July 2017).
- Naimi, B. usdm: Uncertainty Analysis for Species Distribution Models. 2015 R package Version 1.1-15. Available online: https://CRAN.R-project.org/package=usdm (accessed on 1 July 2017).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science+Business Media: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Cliff, A.D.; Ord, J.K. Spatial Processes: Models and Applications; Pion Ltd.: London, UK, 1981. [Google Scholar]
- Bivand, R.; Altman, M.; Anselin, L.; Assunção, R.; Berke, O.; Bernat, A.; Blanchet, G.; Blankmeyer, E.; Carvalho, M.; Christensen, B.; et al. Package Spdep: Spatial Dependence—Weighting Schemes, Statistics and mOdels. R Package Version 0.5-88. 2015. Available online: www.cran.r-project.org (accessed on 1 July 2017).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 1 July 2017).
- Blair, R. The effects of urban sprawl on birds at multiple levels of biological organization. Ecol. Soc. 2004, 9, 2. [Google Scholar] [CrossRef]
- Fernandez-Juricic, E.; Jokimäki, J. A habitat island approach to conserving birds in urban landscapes: Case studies from southern and northern Europe. Biodivers. Conserv. 2001, 10, 2023–2043. [Google Scholar] [CrossRef]
- Faeth, S.H.; Kane, T.C. Urban biogeography. Oecologia 1978, 32, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Basham, R.; Law, B.; Banks, P. Microbats in a ‘leafy’ urban landscape are they persisting, and what factors influence their presence? Austral Ecol. 2010, 36, 663–678. [Google Scholar] [CrossRef]
- Shanahan, D.F.; Miller, C.; Possingham, H.P.; Fuller, R.A. The influence of patch area and connectivity on avian communities in urban revegetation. Biol. Conserv. 2011, 144, 722–729. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Arrhenius, O. Species and area. J. Ecol. 1921, 9, 95–99. [Google Scholar] [CrossRef]
- Drinnan, I.N. The search for fragmentation thresholds in a Southern Sydney Suburb. Biol. Conserv. 2005, 124, 339–349. [Google Scholar] [CrossRef]
- Conole, L.E.; Kirkpatrick, J.B. Functional and spatial differentiation of urban bird assemblages at the landscape scale. Landsc. Urban Plan. 2011, 100, 11–23. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Hanski, I. Metapopulation dynamics. Nature 1998, 396, 41. [Google Scholar] [CrossRef]
- Dondina, O.; Orioli, V.; Massimino, D.; Pinoli, G.; Bani, L. A method to evaluate the combined effect of tree species composition and woodland structure on indicator birds. Ecol. Indic. 2015, 55, 44–51. [Google Scholar] [CrossRef]
- Saura, S.; Bodin, Ö.; Fortin, M.J. Stepping stones are crucial for species’ long-distance dispersal and range expansion through habitat networks. J. Appl. Ecol. 2014, 51, 171–182. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Adams, L.W.; Dove, L.E. Wildlife Reserves and Corridors in the Urban Environment: A Guide to Ecological Landscape Planning and Resource Conservation; National Institute for Urban Wildlife: Columbia, MD, USA, 1989. [Google Scholar]
- Alberti, M. The effects of urban patterns on ecosystem function. Int. Reg. Sci. Rev. 2005, 28, 168–192. [Google Scholar] [CrossRef]
- Laurance, W.F. Theory meets reality: How habitat fragmentation research has transcended island biogeographic theory. Biol. Conserv. 2008, 141, 1731–1744. [Google Scholar] [CrossRef]
- Benfield, F.K. Once there were greenfields. In Forum for Applied Research and Public Policy; Benfield, F.K., Ed.; University of Tennessee, Energy, Environment and Resources Center: Knoxville, TN, USA, 1999; Volume 14, No. 3; p. 6. [Google Scholar]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Giplin, M.E.; Soulé, M.E. Minimum viable populations: Processes of species extinctions. In Conservation Biology: The Science of Scarcity and Diversity; Soulé, M.E., Ed.; Sinauer Associates: Sunderland, MA, USA, 1986; pp. 19–34. [Google Scholar]
- Crnokrack, P.; Roff, D.A. Inbreeding depression in the wild. Heredity 1999, 83, 260–270. [Google Scholar] [CrossRef]
- Frankham, R.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: New York, NY, USA, 2002. [Google Scholar]
- Andren, H. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A review. Oikos 1994, 355–366. [Google Scholar] [CrossRef]
- Whitney, G.G.; Adams, S.D. Man as a maker of new plant communities. J. Appl. Ecol. 1980, 17, 431–448. [Google Scholar] [CrossRef]
- Godron, M.; Forman, R.T. Landscape modification and changing ecological characteristics. In Disturbance and Ecosystems; Mooney, H.A., Godron, M., Eds.; Springer: New York, NY, USA, 1983; pp. 12–28. [Google Scholar]
- Mooney, H.A.; Gulmon, S.L. The determinants of plant productivity—Natural versus man-modified communities. In Disturbance and Ecosystems; Mooney, H.A., Godron, M., Eds.; Springer: New York, NY, USA, 1983; pp. 146–158. [Google Scholar]
- Rudnicky, J.L.; McDonnell, M.J. Forty-eight years of canopy change in a hardwood-hemlock forest in New York City. Bull. Torrey Bot. Club 1989, 116, 52–64. [Google Scholar] [CrossRef]
- Galvin, K.A.; Reid, R.S.; Behnke, R.H.; Hobbs, N.T. Fragmentation in Semi-Arid and Arid Landscapes. Consequences for Human and Natural Systems; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Sicurella, B.; Orioli, V.; Pinoli, G.; Ambrosini, R.; Bani, L. Effectiveness of the system of protected areas of Lombardy (Northern Italy) in preserving breeding birds. Bird Conserv. Int. 2017, 1–18. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, biodiversity, and conservation: The impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Faeth, S.H.; Bang, C.; Saari, S. Urban biodiversity: Patterns and mechanisms. Ann. N. Y. Acad. Sci. 2011, 1223, 69–81. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Predictors | Estimate | SE | z | Pr(>|z|) | VIF |
|---|---|---|---|---|---|
| (Intercept) | 2.290 | 0.044 | 52.135 | <0.001 | - |
| PA | 0.065 | 0.012 | 3.398 | <0.001 | 2.24 |
| DSA | −0.059 | 0.030 | −1.986 | 0.047 | 2.49 |
| NF | 0.063 | 0.019 | 3.398 | <0.001 | 1.75 |
| PrA | 0.036 | 0.048 | 0.745 | 0.456 | 1.48 |
| URB | −0.151 | 0.041 | −3.656 | <0.001 | 1.33 |
| East | −0.097 | 0.022 | −4.369 | <0.001 | 1.46 |
| North | −0.033 | 0.027 | −1.214 | 0.225 | 2.18 |
| Predictors | Estimate | SE | z | Pr(>|z|) | VIF |
|---|---|---|---|---|---|
| (Intercept) | 2.313 | 0.037 | 61.634 | <0.001 | - |
| PA | 0.093 | 0.012 | 7.968 | <0.001 | 2.06 |
| DSA | −0.116 | 0.025 | −4.662 | <0.001 | 1.89 |
| NF | 0.032 | 0.015 | 2.110 | 0.035 | 1.96 |
| PrA | 0.169 | 0.043 | 3.917 | <0.001 | 1.81 |
| URB | −0.163 | 0.033 | −4.947 | <0.001 | 1.12 |
| East | 0.136 | 0.020 | 6.859 | <0.001 | 1.88 |
| North | −0.038 | 0.024 | −1.523 | 0.128 | 2.07 |
| Predictors | Estimate | SE | z | Pr(>|z|) | VIF |
|---|---|---|---|---|---|
| (Intercept) | 2.068 | 0.044 | 47.5341 | <0.001 | - |
| PA | 0.097 | 0.037 | 2.650 | 0.008 | 2.76 |
| DSA | 0.007 | 0.049 | 0.143 | 0.886 | 2.70 |
| NF | 0.002 | 0.042 | 0.053 | 0.958 | 2.57 |
| PrA | 0.098 | 0.061 | 1.600 | 0.110 | 1.18 |
| East | −0.146 | 0.035 | −4.145 | <0.001 | 1.18 |
| North | 0.069 | 0.048 | 1.441 | 0.150 | 2.86 |
| Predictors | Estimate | SE | z | Pr(>|z|) | VIF |
|---|---|---|---|---|---|
| (Intercept) | 2.077 | 0.038 | 54.765 | <0.001 | - |
| PA | 0.162 | 0.013 | 12.092 | <0.001 | 2.74 |
| DSA | −0.187 | 0.042 | −4.461 | <0.001 | 2.64 |
| NU | −0.104 | 0.022 | −4.646 | <0.001 | 2.95 |
| PrA | 0.150 | 0.054 | 2.787 | 0.005 | 1.42 |
| East | 0.105 | 0.025 | 4.215 | <0.001 | 1.42 |
| North | −0.046 | 0.042 | −1.086 | 0.277 | 2.75 |
| Predictors | Estimate | SE | z | Pr(>|z|) | VIF |
|---|---|---|---|---|---|
| (Intercept) | 2.395 | 0.073 | 32.618 | <0.001 | - |
| PA | 0.093 | 0.016 | 5.654 | <0.001 | 2.35 |
| DSA | −0.100 | 0.040 | −2.443 | 0.015 | 2.57 |
| NF | 0.054 | 0.022 | 2.416 | 0.016 | 1.88 |
| PrA | −0.082 | 0.094 | −0.872 | 0.383 | 2.57 |
| East | −0.083 | 0.038 | −2.179 | 0.029 | 2.73 |
| North | −0.131 | 0.036 | −3.608 | <0.001 | 2.12 |
| Predictors | Estimate | SE | z | Pr(>|z|) | VIF |
|---|---|---|---|---|---|
| (Intercept) | 2.572 | 0.037 | 69.086 | <0.001 | - |
| PA | 0.048 | 0.020 | 2.402 | 0.016 | 1.68 |
| DSA | −0.062 | 0.034 | −1.809 | 0.070 | 1.55 |
| NF | 0.040 | 0.023 | 1.735 | 0.083 | 1.52 |
| PrA | −0.058 | 0.049 | −1.177 | 0.239 | 1.17 |
| North | −0.006 | 0.033 | −0.192 | 0.848 | 1.80 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canedoli, C.; Orioli, V.; Padoa-Schioppa, E.; Bani, L.; Dondina, O. Temporal Variation of Ecological Factors Affecting Bird Species Richness in Urban and Peri-Urban Forests in a Changing Environment: A Case Study from Milan (Northern Italy). Forests 2017, 8, 507. https://doi.org/10.3390/f8120507
Canedoli C, Orioli V, Padoa-Schioppa E, Bani L, Dondina O. Temporal Variation of Ecological Factors Affecting Bird Species Richness in Urban and Peri-Urban Forests in a Changing Environment: A Case Study from Milan (Northern Italy). Forests. 2017; 8(12):507. https://doi.org/10.3390/f8120507
Chicago/Turabian StyleCanedoli, Claudia, Valerio Orioli, Emilio Padoa-Schioppa, Luciano Bani, and Olivia Dondina. 2017. "Temporal Variation of Ecological Factors Affecting Bird Species Richness in Urban and Peri-Urban Forests in a Changing Environment: A Case Study from Milan (Northern Italy)" Forests 8, no. 12: 507. https://doi.org/10.3390/f8120507