
Landscape Structure and Mature Forest Biodiversity in Wet Eucalypt Forests: A Spatial Analysis of Timber Production Areas in South-Eastern Australia

Abstract
:1. Introduction
2. Materials and Methods
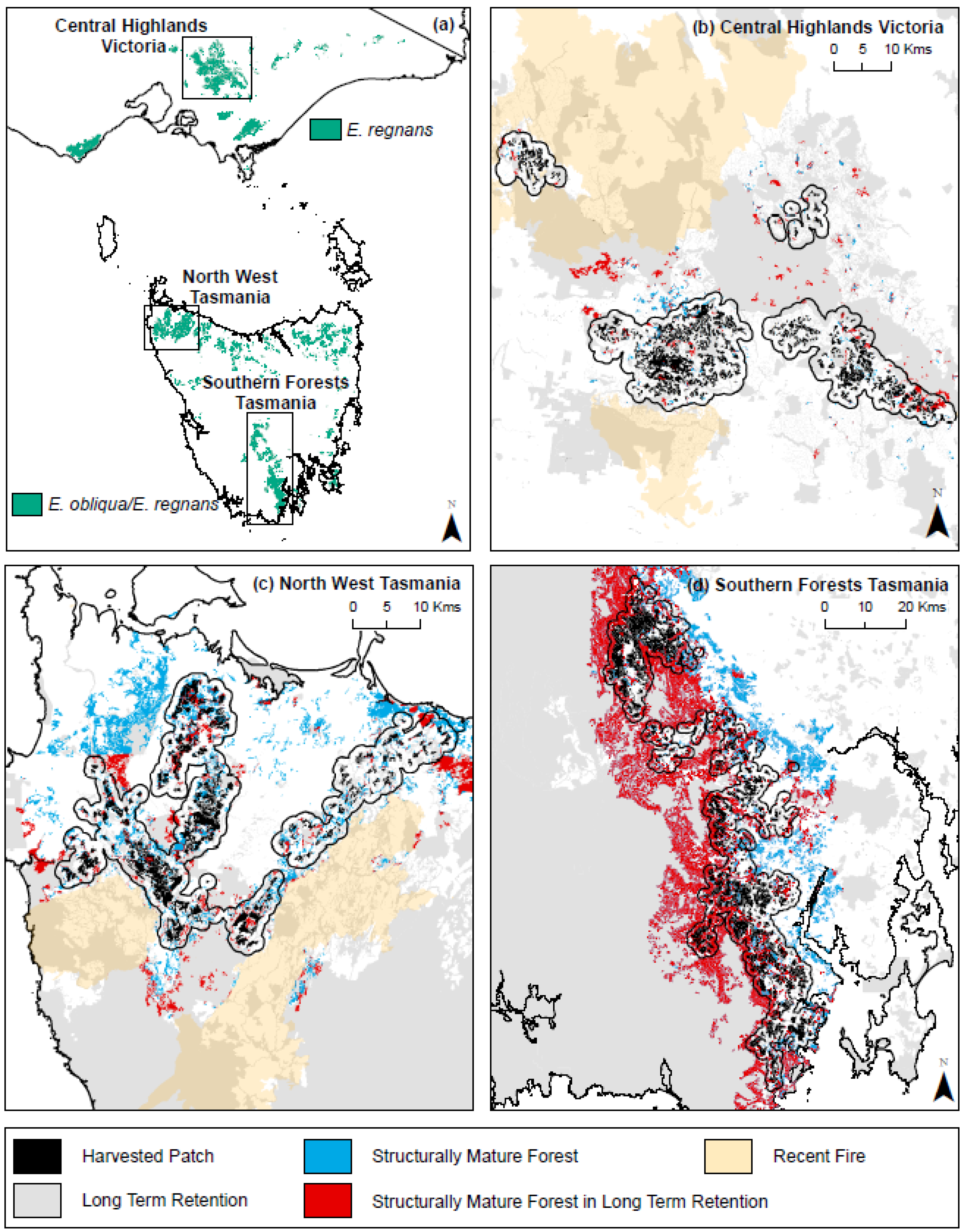
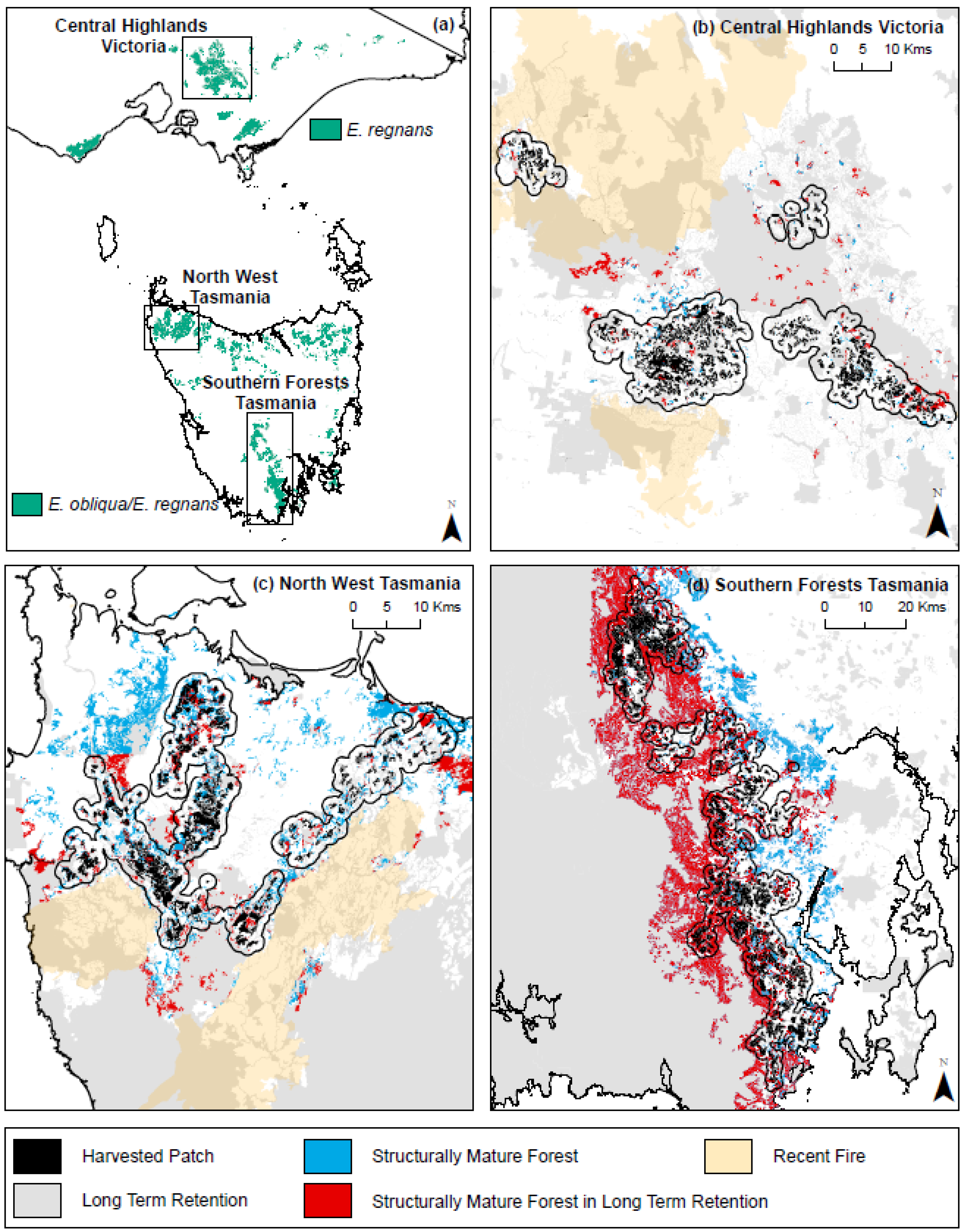
2.1. Forest Landscapes
2.2. Spatial Datasets
2.3. Harvested Patches
2.4. Structurally Mature Forest
2.5. Long-Term Retention
2.6. 1939 Regrowth in Long-Term Retention
2.7. Analysis
2.7.1. Geospatial Metrics
2.7.2. Long-Term Retention
3. Results
3.1. Structural Maturity
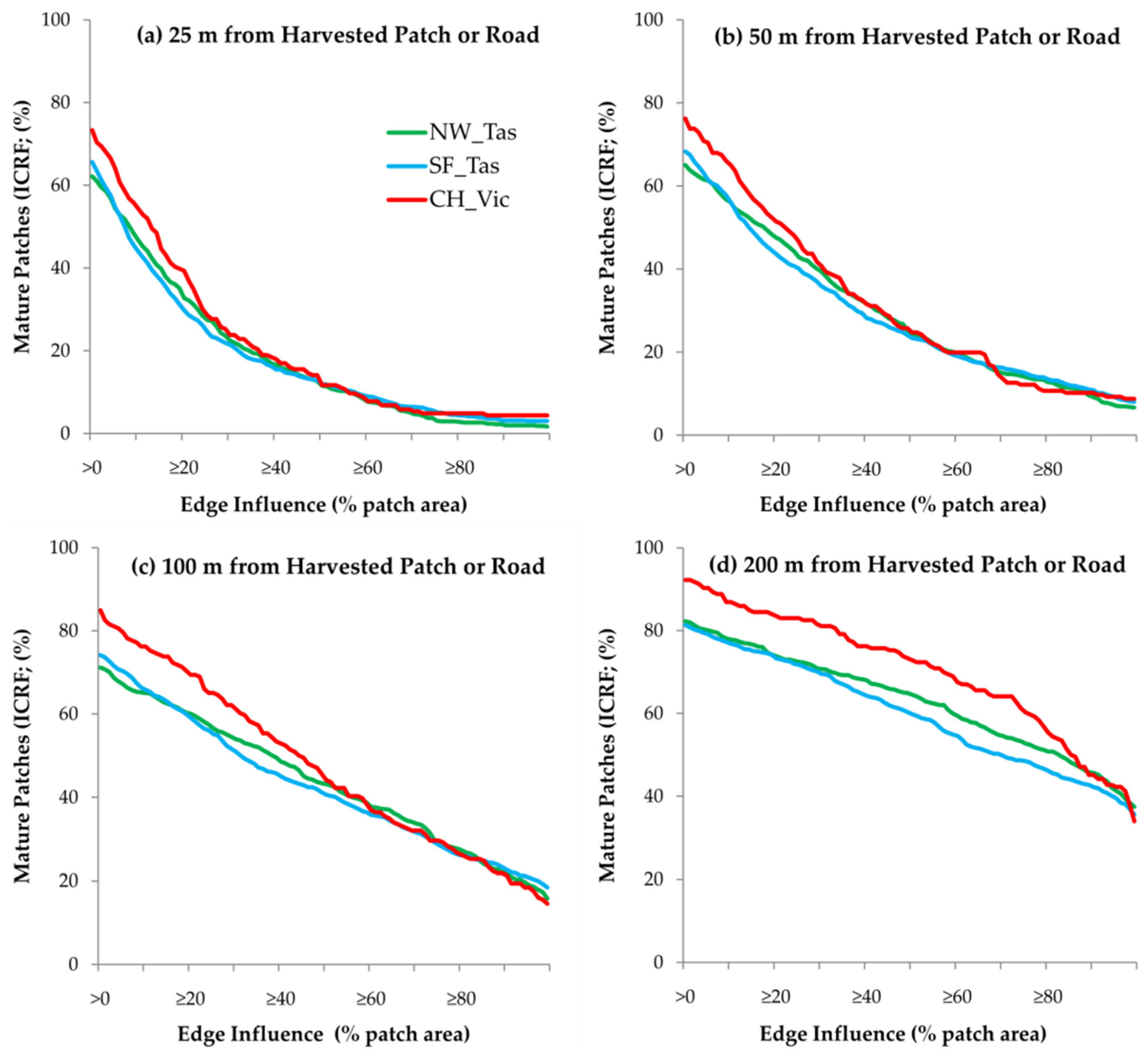
3.2. Edge Influence
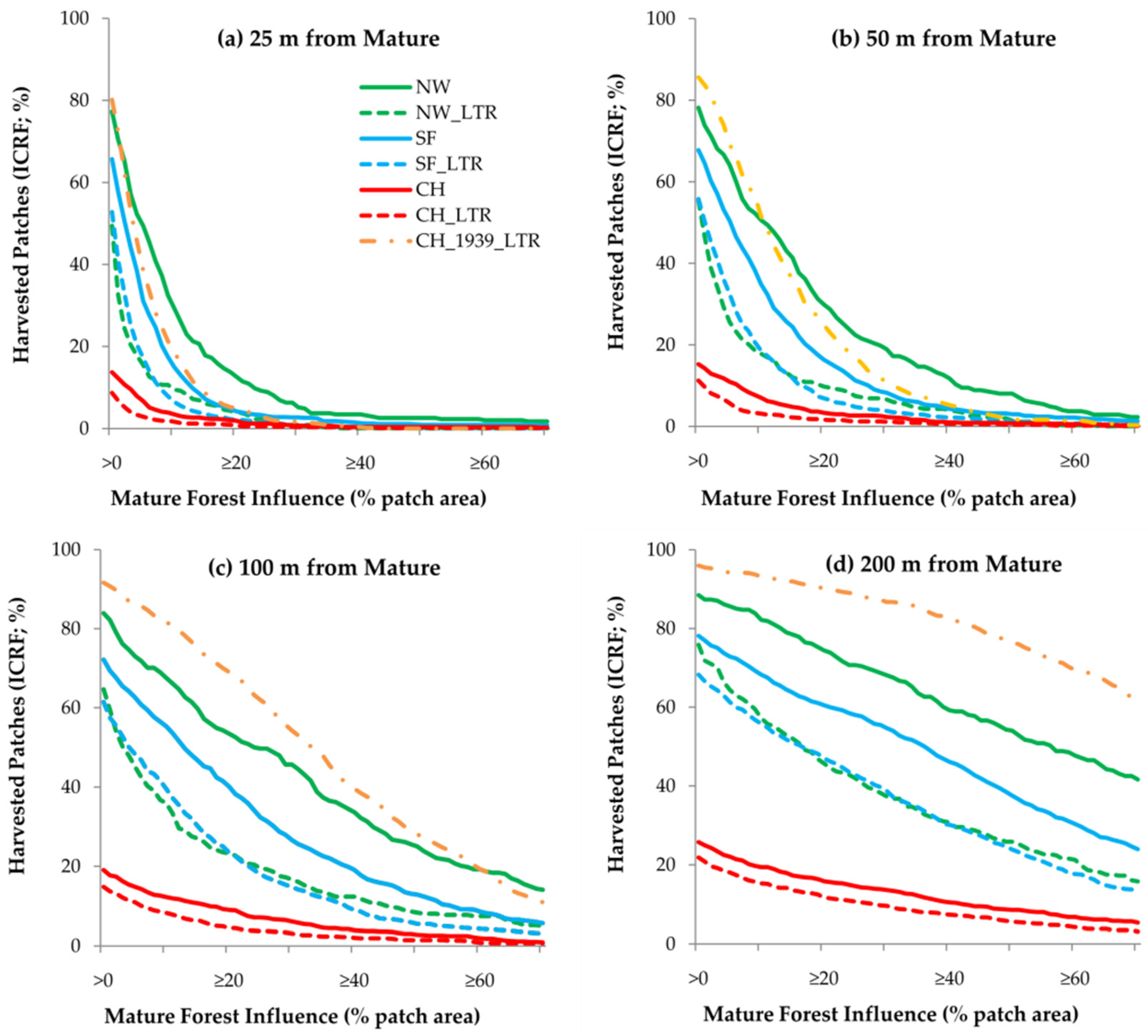
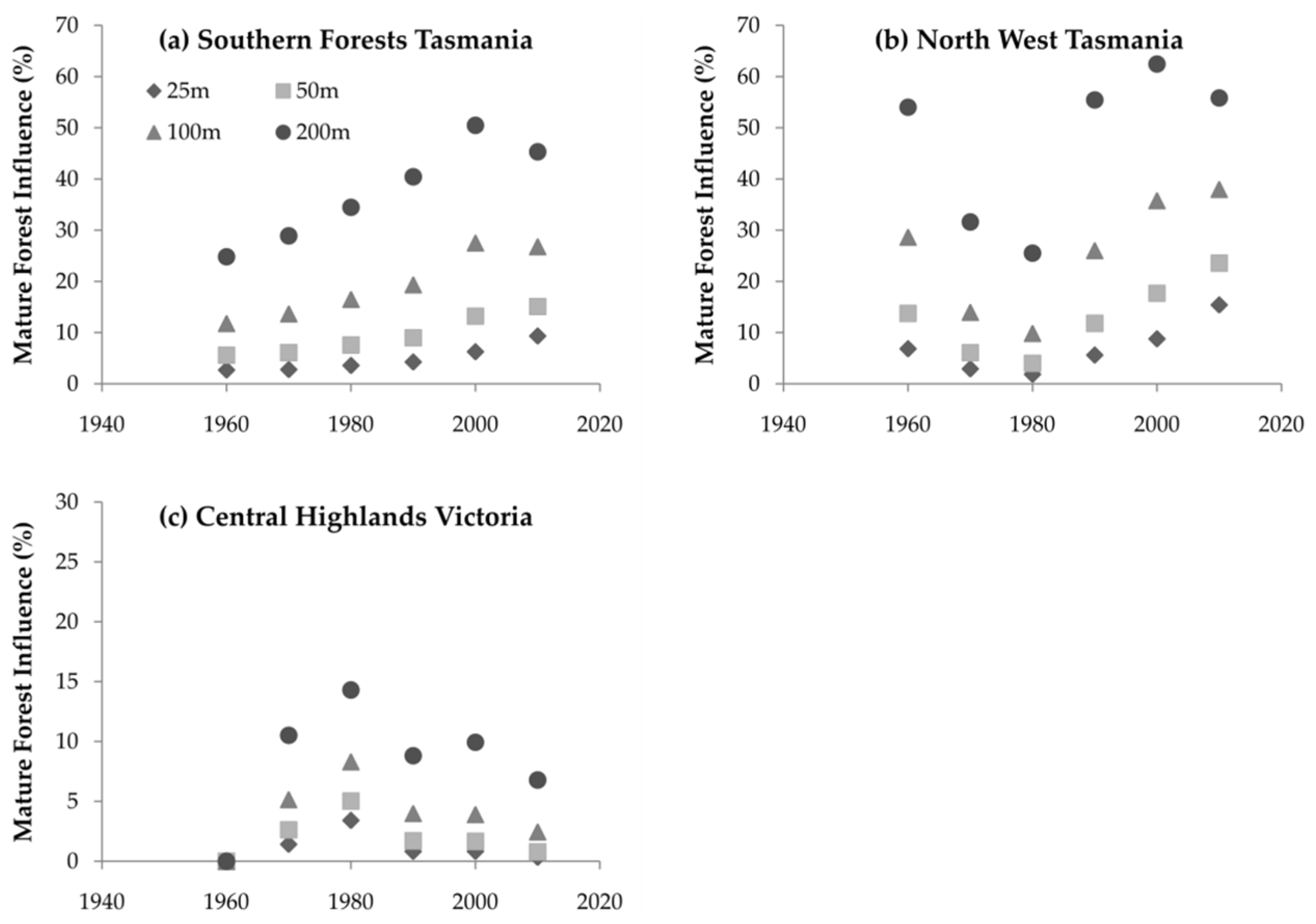
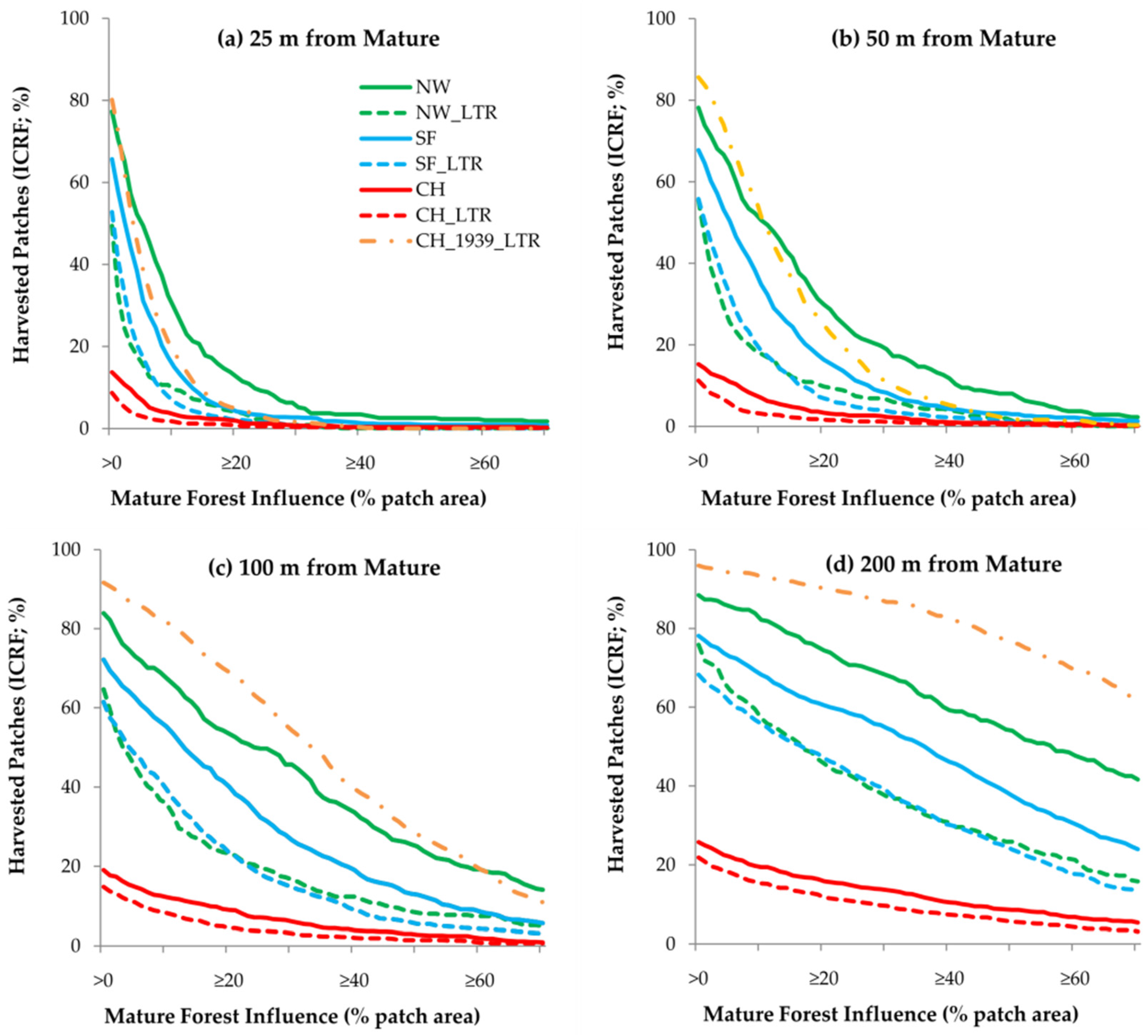
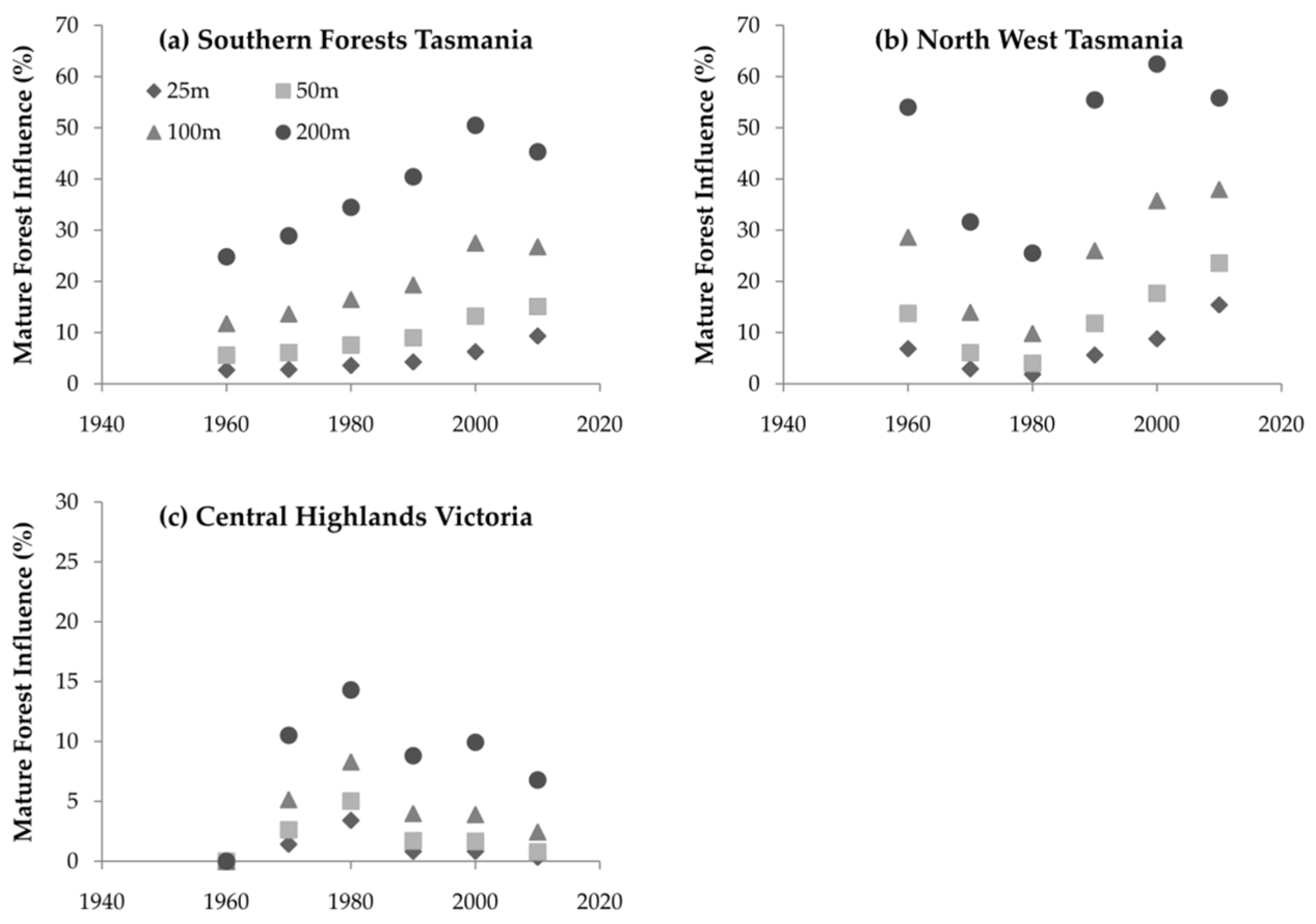
3.3. Mature Forest Influence
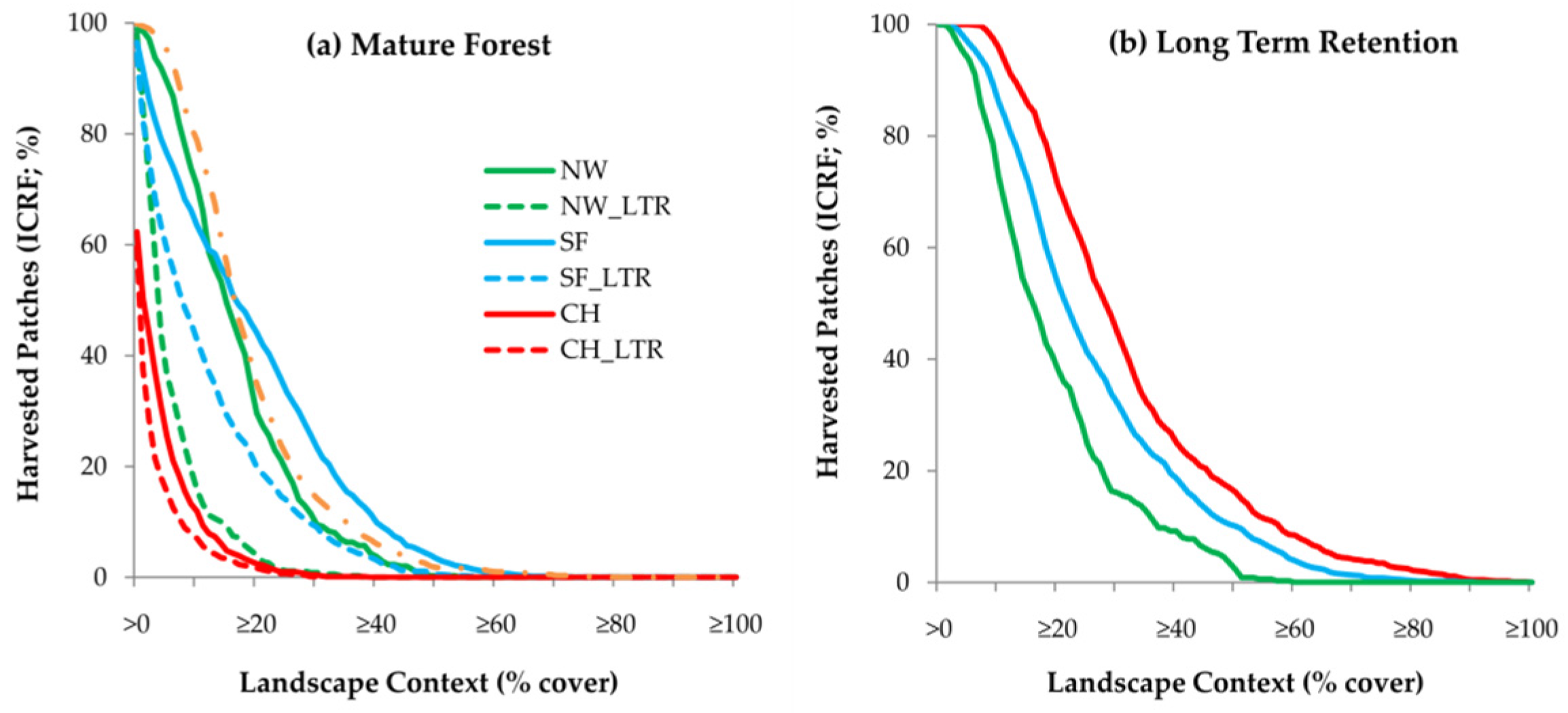
3.4. Landscape Context
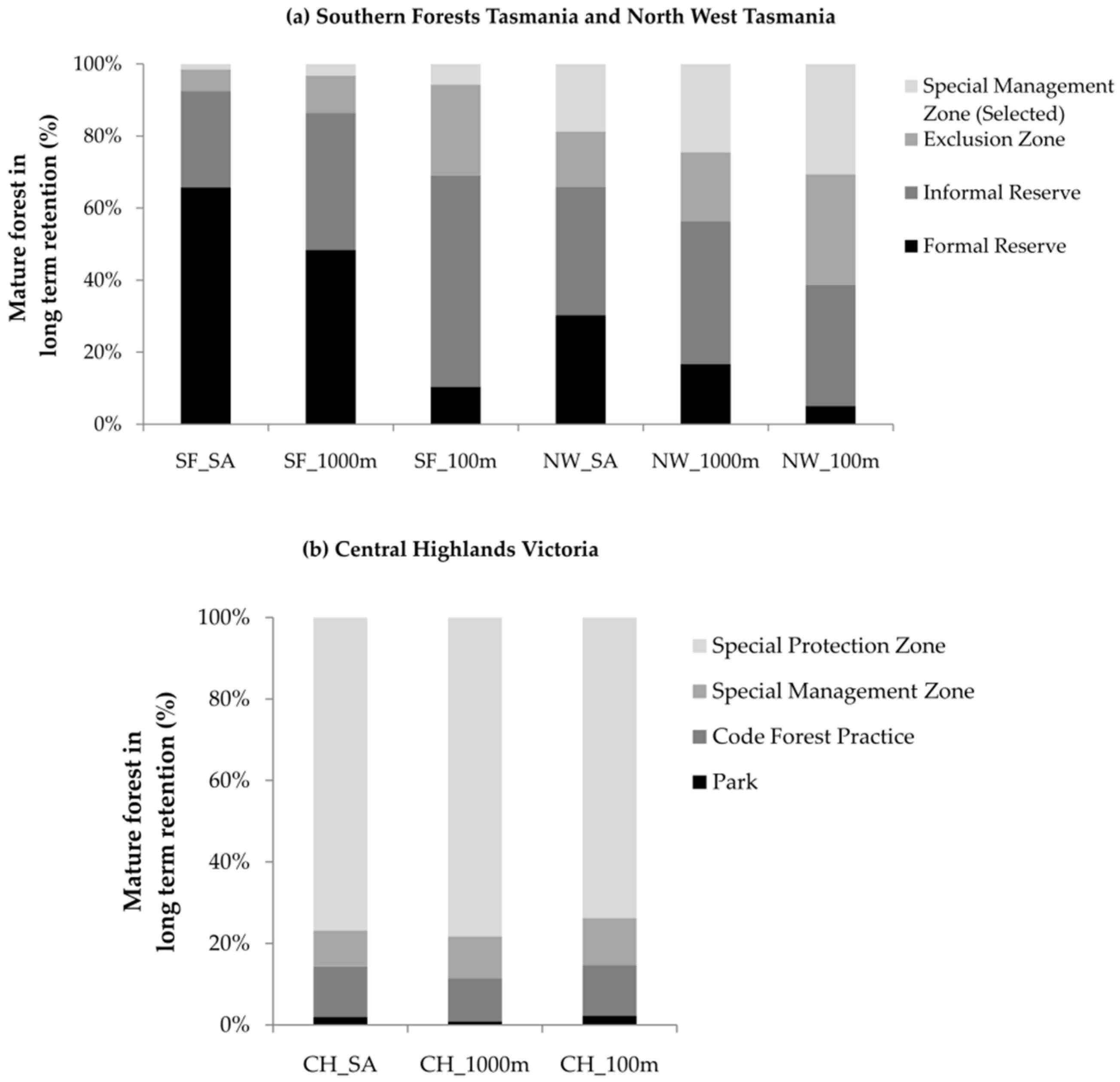
3.5. Long-Term Retention
4. Discussion
4.1. Structural Maturity
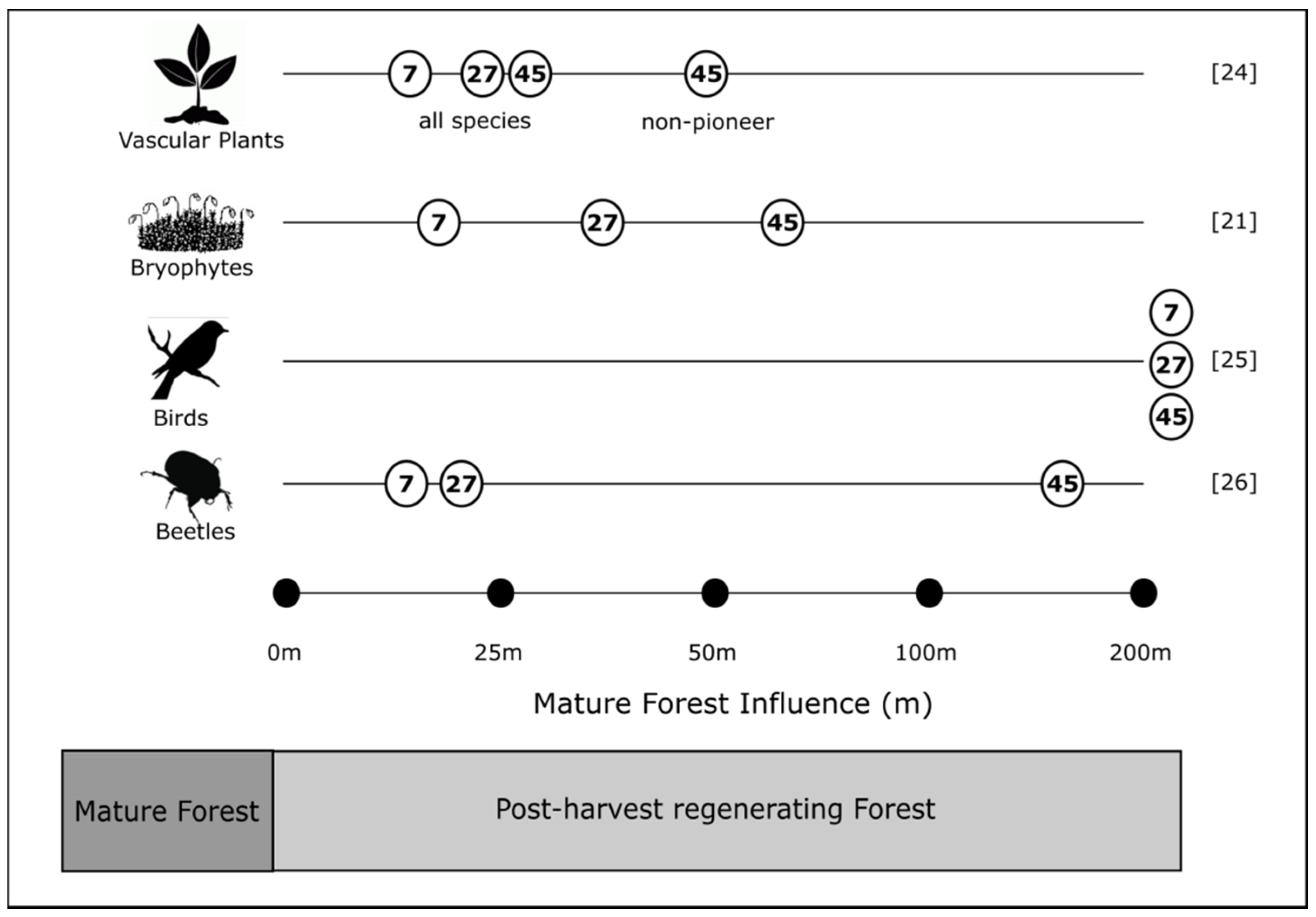
4.2. Edge Influence
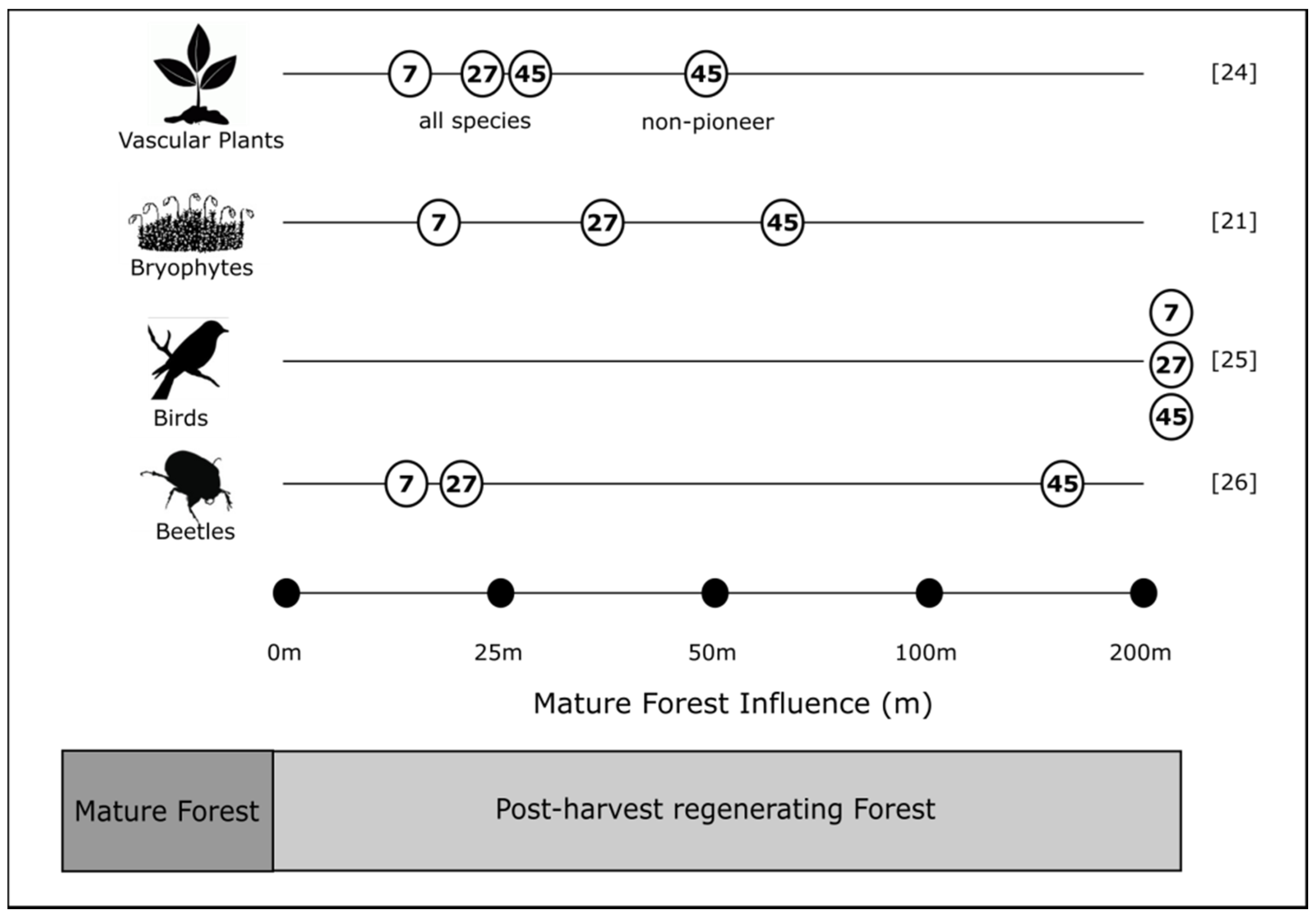
4.3. Mature Forest Influence
4.4. Landscape Context
4.5. Long-Term Retention
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tasmania | Description | Long-Term Retention (Y/N) |
|---|---|---|
| Public Formal | Formal Reserves on Public Land | Y |
| Public Informal | Informal Reserves on Public Land including those established through Forest Practices Code | Y |
| Special Management Zones | Masked Owl | Y |
| Swift Parrot | Y | |
| Long-Term Retention | Y | |
| Stag Beetle | Y | |
| Variable Retention; | Y | |
| Other Special Management Zones # | N | |
| Provisional Coupe Exclusion Zones | Streamside Reserves | Y |
| Wildlife Habitat Clumps | Y | |
| Other Exclusion Zones ^ | N | |
| Private Informal | Informal Private Reserves | N |
| Future Potential Production | (not previously reserved) | N |
| Plantations | Forestry Tasmania Plantations | N |
| Wood Production Zones | Forestry Tasmania Wood Production | N |
| Other Public and Private Land | Other Public and Private Land | N |
| Victoria | Description | Long-Term Retention (Y/N) |
|---|---|---|
| Park | National Parks, State Parks, Regional Parks, Flora and Fauna Reserves | Y |
| Special Management Zone | Managed to conserve specific features, while catering for timber harvesting operations under special conditions | Y |
| Special Protection Zone | Managed for particular conservation values # to complement formal conservation reserve system. Timber harvesting operations generally excluded | Y |
| Code Forest Practice | Exclusion areas where timber harvesting cannot occur due to streamside protection buffers, steep slopes, etc.) | Y |
| General Management Zone | Managed for a range of uses and values, but timber harvesting operations will have a high priority. | N |
| Other Public Land | Other Public Land | N |
| Plantation | Plantation | N |
References
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Howell, C.; Wilson, A.; Davey, S.; Eddington, M. Sustainable forest management reporting in Australia. Ecol. Indic. 2008, 8, 123–130. [Google Scholar] [CrossRef]
- Hunter, M.L. Maintaining Biodiversity in Forest Ecosystems; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Lindenmayer, D.B.; Franklin, J.F. Conserving Forest Biodiversity: A Comprehensive Multiscaled Approach; Island Press: Washington, DC, USA, 2002. [Google Scholar]
- Lindenmayer, D. Interactions between forest resource management and landscape structure. Curr. Landsc. Ecol. Rep. 2016, 1, 10–18. [Google Scholar] [CrossRef]
- Gustafsson, L.; Perhans, K. Biodiversity conservation in Swedish forests: Ways forward for a 30-year-old multi-scaled approach. Ambio 2010, 39, 546–554. [Google Scholar] [CrossRef] [PubMed]
- McGarigal, K.; McComb, W.C. Relationships between landscape structure and breeding birds in the Oregon Coast Range. Ecol. Monogr. 1995, 65, 235–260. [Google Scholar] [CrossRef]
- Jokimäki, J.; Huhta, E. Effects of landscape matrix and habitat structure on a bird community in northern Finland: A multi-scale approach. Ornis Fenn. 1996, 73, 97–113. [Google Scholar]
- Drapeau, P.; Leduc, A.; Giroux, J.-F.; Savard, J.-P.L.; Bergeron, Y.; Vickery, W.L. Landscape-scale disturbances and changes in bird communities of boreal mixed-wood forests. Ecol. Monogr. 2000, 70, 423–444. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.; Jaiteh, M.S.; Esseen, P.A. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Larrivée, M.; Drapeau, P.; Fahrig, L. Edge effects created by wildfire and clear-cutting on boreal forest ground-dwelling spiders. For. Ecol. Manag. 2008, 255, 1434–1445. [Google Scholar] [CrossRef]
- Dupuch, A.; Fortin, D. The extent of edge effects increases during post-harvesting forest succession. Biol. Conserv. 2013, 162, 9–16. [Google Scholar] [CrossRef]
- Chen, J.; Franklin, J.F.; Spies, T.A. Vegetation responses to edge environments in old-growth Douglas-fir forests. Ecol. Appl. 1992, 2, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Mascarúa López, L.E.; Harper, K.A.; Drapeau, P. Edge influence on forest structure in large forest remnants, cutblock separators, and riparian buffers in managed black spruce forests. Ecoscience 2006, 13, 226–233. [Google Scholar] [CrossRef]
- Baker, T.P.; Jordan, G.J.; Dalton, P.J.; Baker, S.C. Impact of distance to mature forest on the recolonisation of bryophytes in a regenerating Tasmanian wet eucalypt forest. Aust. J. Bot. 2014, 61, 633–642. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Peres, C.A.; Dent, D.; Sheil, D.; Lugo, A.E.; Lamb, D.; Stork, N.E.; Miller, S.E. The potential for species conservation in tropical secondary forests. Conserv. Biol. 2009, 23, 1406–1417. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.C.; Spies, T.A.; Wardlaw, T.J.; Balmer, J.; Franklin, J.F.; Jordan, G.J. The harvested side of edges: Effect of retained forests on the re-establishment of biodiversity in adjacent harvested areas. For. Ecol. Manag. 2013, 302, 107–121. [Google Scholar] [CrossRef]
- Pulsford, S.A.; Lindenmayer, D.B.; Driscoll, D.A. A succession of theories: Purging redundancy from disturbance theory. Biol. Rev. 2014, 91, 148–167. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, T.; Grove, S.; Balmer, J.; Hingston, A.; Forster, L.; Schmuki, C.; Read, S. Persistence of Mature Forest Biodiversity in a Production Forest Landscape Managed under a Regional Forest Agreement; Forest and Wood Products Australia: Hobart, Tasmania, Australia, 2012.
- Heithecker, T.D.; Halpern, C.B. Edge-related gradients in microclimate in forest aggregates following structural retention harvests in western Washington. For. Ecol. Manag. 2007, 248, 163–173. [Google Scholar] [CrossRef]
- Baker, T.P.; Jordan, G.J.; Baker, S.C. Microclimatic edge effects in a recently harvested forest: Do remnant forest patches create the same impact as large forest areas? For. Ecol. Manag. 2016, 365, 128–136. [Google Scholar] [CrossRef]
- Tabor, J.; McElhinny, C.; Hickey, J.; Wood, J. Colonisation of clearfelled coupes by rainforest tree species from mature mixed forest edges, Tasmania, Australia. For. Ecol. Manag. 2007, 240, 13–23. [Google Scholar] [CrossRef]
- Matlack, G.R. Vegetation dynamics of the forest edge–trends in space and successional time. J. Ecol. 1994, 82, 113–123. [Google Scholar] [CrossRef]
- Balmer, J. Floristic Response to Landscape Context in Vascular Plant Communities in Eucalyptus obliqua and Eucalyptus regnans Wet Forest, Southern Tasmania. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 2015. [Google Scholar]
- Hingston, A.B.; Jordan, G.J.; Wardlaw, T.J.; Baker, S.C. Bird assemblages in Tasmanian clearcuts are influenced by the age of eucalypt regeneration but not by distance from mature forest. Glob. Ecol. Conserv. 2014, 2, 138–147. [Google Scholar] [CrossRef]
- Fountain-Jones, N.M.; Jordan, G.J.; Baker, T.P.; Balmer, J.M.; Wardlaw, T.; Baker, S.C. Living near the edge: Being close to mature forest increases the rate of succession in beetle communities. Ecol. Appl. 2015, 25, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Mazerolle, M.J.; Villard, M.-A. Patch characteristics and landscape context as predictors of species presence and abundance: A review. Ecoscience 1999, 6, 117–124. [Google Scholar]
- Radford, J.Q.; Bennett, A.F.; Cheers, G.J. Landscape-level thresholds of habitat cover for woodland-dependent birds. Biol. Conserv. 2005, 124, 317–337. [Google Scholar] [CrossRef]
- Betts, M.G.; Forbes, G.J.; Diamond, A.W. Thresholds in songbird occurrence in relation to landscape structure. Conserv. Biol. 2007, 21, 1046–1058. [Google Scholar] [CrossRef] [PubMed]
- Drapeau, P.; Villard, M.A.; Leduc, A.; Hannon, S.J. Natural disturbance regimes as templates for the response of bird species assemblages to contemporary forest management. Divers. Distrib. 2016, 22, 385–399. [Google Scholar] [CrossRef]
- Wood, S.; Bowman, D.; Prior, L.; Lindenmayer, D.; Wardlaw, T.; Robinson, R. Tall eucalypt forests. In Biodiversity and Environmental Change; Lindenmayer, D., Burns, E., Thurgate, N., Lowe, A., Eds.; CSIRO Publishing: Canberra, ACT, Australia, 2014; pp. 531–581. [Google Scholar]
- Lindenmayer, D. Forest Pattern and Ecological Process: A Synthesis of 25 Years of Research; CSIRO Publishing: Melbourne, Australia, 2009. [Google Scholar]
- Lindenmayer, D.; Blair, D.; McBurney, L.; Banks, S. Mountain Ash: Fire, Logging and the Future of Victoria’s Giant Forests; CSIRO Publishing: Melbourne, Australia, 2015. [Google Scholar]
- Baker, S.C.; Read, S.M. Variable retention silviculture in Tasmania’s wet forests: Ecological rationale, adaptive management and synthesis of biodiversity benefits. Aust. For. 2011, 74, 218–232. [Google Scholar] [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Pastur, G.M.; Messier, C. Retention forestry to maintain multifunctional forests: A world perspective. Bioscience 2012, 62, 633–645. [Google Scholar]
- VicForests. Regrowth Retention Harvesting Instruction. Version 2.0.; VicForests: Melbourne, Victoria, Australia, 2016. [Google Scholar]
- Keenan, R.J.; Kimmins, J. The ecological effects of clear-cutting. Environ. Rev. 1993, 1, 121–144. [Google Scholar] [CrossRef]
- Scott, R.E.; Neyland, M.G.; McElwee, D.J.; Baker, S.C. Burning outcomes following aggregated retention harvesting in old-growth wet eucalypt forests. For. Ecol. Manag. 2012, 276, 165–173. [Google Scholar] [CrossRef]
- Forestry Tasmania. Landscape Context Planning System Manual; Forestry Tasmania: Hobart, Tasmania, Australia, 2014. [Google Scholar]
- Department of Environment and Primary Industries (DEPI). Planning Standards for Timber Havesting Operations in Victoria’s State Forests 2014: Appendix 5 to the Management Standards and Procedures for Timber Harvesting Operations in Victoria’s State Forests 2014; Department of Environment and Primary Industries: Melbourne, Victoria, Australia, 2014. [Google Scholar]
- Wood, S.W.; Prior, L.D.; Stephens, H.C.; Bowman, D.M. Macroecology of Australian tall eucalypt forests: Baseline data from a continental-scale permanent plot network. PLoS ONE 2015, 10, e0137811. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.A.; Balmer, J.; Kirkpatrick, J. Stand-replacing wildfires: The incidence of multi-cohort and single-cohort Eucalyptus regnans and E. obliqua forests in southern Tasmania. For. Ecol. Manag. 2009, 258, 366–375. [Google Scholar] [CrossRef]
- Attiwill, P.M. The disturbance of forest ecosystems: The ecological basis for conservative management. For. Ecol. Manag. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Hickey, J.; Wilkinson, G. The development and current implementation of silvicultural pratices in native forests in Tasmania. Aust. For. 1999, 62, 245–254. [Google Scholar] [CrossRef]
- McCarthy, M.A.; Lindenmayer, D.B. Multi-aged mountain ash forest, wildlife conservation and timber harvesting. For. Ecol. Manag. 1998, 104, 43–56. [Google Scholar] [CrossRef]
- Stone, M. Forest-type mapping by photo-interpretation: A multi-purpose base for Tasmania’s forest management. Tasforests 1998, 10, 15–32. [Google Scholar]
- Hamilton, F.; Penny, R.; Black, P.; Cumming, F.; Irvine, M. Victoria’s Statewide Forest Resource Industry—an outline of methods. Aust. For. 1999, 62, 353–359. [Google Scholar] [CrossRef]
- Brookhouse, M.; Brack, C.; McElhinny, C. The distance to structural complement (DiSCo) approach for expressing forest structure described by Aerial Photograph Interpretation data sets. For. Ecol. Manag. 2010, 260, 1230–1240. [Google Scholar] [CrossRef]
- Fox, J.C.; Hamilton, F.; Ades, P.K. Models of tree-level hollow incidence in Victorian State forests. For. Ecol. Manag. 2008, 255, 2846–2857. [Google Scholar] [CrossRef]
- Fox, J.C.; Hamilton, F.; Occhipinti, S. Tree hollow incidence in Victorian state forests. Aust. For. 2009, 72, 39–48. [Google Scholar] [CrossRef]
- Koch, A.; Baker, S. Remotely assessing hollow availability using aerial photographs. Biodivers. Conserv. 2011, 20, 1089–1101. [Google Scholar] [CrossRef]
- Gibbons, P.; Lindenmayer, D. Tree Hollows and Wildlife Conservation in Australia; CSIRO Publishing: Melbourne, Australia, 2002. [Google Scholar]
- Koch, A.J.; Munks, S.A.; Driscoll, D.; Kirkpatrick, J. Does hollow occurrence vary with forest type? A case study in wet and dry Eucalyptus obliqua forest. For. Ecol. Manag. 2008, 255, 3938–3951. [Google Scholar] [CrossRef]
- Yee, M.; Grove, S.; Richardson, A.M.; Mohammed, C. Brown rot in inner heartwood: Why large logs support characteristic saproxylic beetle assemblages of conservation concern. In Insect Biodiversity and Dead Wood: Proceedings of a Symposium for the 22nd International Congress of Entomology; Gen. Tech. Rep. SRS-93; Grove, S.J., Hanula, J.L., Eds.; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 2006; pp. 42–56. [Google Scholar]
- Wardlaw, T.; Grove, S.; Hopkins, A.; Yee, M.; Harrison, K.; Mohammed, C. The uniqueness of habitats in old eucalypts: Contrasting wood-decay fungi and saproxylic beetles of young and old eucalypts. Tasforests 2009, 18, 17–32. [Google Scholar]
- Lindenmayer, D.B.; Blanchard, W.; McBurney, L.; Blair, D.; Banks, S.; Likens, G.E.; Franklin, J.F.; Laurance, W.F.; Stein, J.A.; Gibbons, P. Interacting factors driving a major loss of large trees with cavities in a forest ecosystem. PLoS ONE 2012, 7, e41864. [Google Scholar] [CrossRef] [PubMed]
- Westphalen, G. The Ecology of Edges in Tasmanian Wet Forests Managed for Timber Production. Ph.D. Thesis, University of Tasmania, Hobart, Tasmania, Australia, 2003. [Google Scholar]
- Baker, S.C.; Barmuta, L.A.; McQuillan, P.B.; Richardson, A.M. Estimating edge effects on ground-dwelling beetles at clearfelled non-riparian stand edges in Tasmanian wet eucalypt forest. For. Ecol. Manag. 2007, 239, 92–101. [Google Scholar] [CrossRef]
- Baker, S.C.; Barmuta, L.A.; Richardson, A.M. Response of ground-dwelling beetles across logging coupe edges into streamside reserves. Aust. J. Entomol. 2009, 48, 194–203. [Google Scholar] [CrossRef]
- Spies, T.A.; Duncan, S.L. Old Growth in a New World: A Pacific Northwest Icon Reexamined; Island Press: Washington, DC, USA, 2012. [Google Scholar]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Hickey, J.; Su, W.; Rowe, P.; Brown, M.; Edwards, L. Fire history of the tall wet eucalypt forests of the Warra ecological research site, Tamania. Aust. For. 1999, 62, 66–71. [Google Scholar] [CrossRef]
- Wood, S.W.; Murphy, B.P.; Bowman, D.M. Firescape ecology: How topography determines the contrasting distribution of fire and rainforest in the south-west of the Tasmanian Wilderness World Heritage Area. J. Biogeogr. 2011, 38, 1807–1820. [Google Scholar] [CrossRef]
- Koch, A.; Chuter, A.; Munks, S.A. A Review of Forestry Impacts on Biodiversity and the Effectiveness of "Off-Reserve" Management Actions in Areas Covered by the Tasmanian Forest Practices System; Forest Practices Authority: Hobart, Tasmania, Australia, 2012. [Google Scholar]
- Berg, Å.; Ehnström, B.; Gustafsson, L.; Hallingbäck, T.; Jonsell, M.; Weslien, J. Threatened plant, animal, and fungus species in Swedish forests: Distribution and habitat associations. Conserv. Biol. 1994, 8, 718–731. [Google Scholar] [CrossRef]
- Bunnell, F.L.; Dunsworth, G.B. Forestry and Biodiversity: Learning How to Sustain Biodiversity in Managed Forests; UBC Press: Vancouver, BC, Canada, 2010. [Google Scholar]
- Neyland, M.G.; Jarman, S.J. Early impacts of harvesting and burning disturbances on vegetation communities in the Warra silvicultural systems trial, Tasmania, Australia. Aust. J. Bot. 2012, 59, 701–712. [Google Scholar] [CrossRef]
- Kowal, V.A.; Cartar, R.V. Edge effects of three anthropogenic disturbances on spider communities in Alberta’s boreal forest. J. Insect Conserv. 2012, 16, 613–627. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Mayerhofer, M.S.; Biswas, S.R.; Esseen, P.A.; Hylander, K.; Stewart, K.J.; Mallik, A.U.; Drapeau, P.; Jonsson, B.G. Edge influence on vegetation at natural and anthropogenic edges of boreal forests in Canada and Fennoscandia. J. Ecol. 2015, 103, 550–562. [Google Scholar] [CrossRef]
- Lacasella, F.; Gratton, C.; de Felici, S.; Isaia, M.; Zapparoli, M.; Marta, S.; Sbordoni, V. Asymmetrical responses of forest and “beyond edge” arthropod communities across a forest–grassland ecotone. Biodivers. Conserv. 2015, 24, 447–465. [Google Scholar] [CrossRef]
- Elbakidze, M.; Angelstam, P.; Andersson, K.; Nordberg, M.; Pautov, Y. How does forest certification contribute to boreal biodiversity conservation? Standards and outcomes in Sweden and NW Russia. For. Ecol. Manag. 2011, 262, 1983–1995. [Google Scholar] [CrossRef]
- Baker, S.C.; Halpern, C.B.; Wardlaw, T.J.; Kern, C.; Edgar, G.J.; Thomson, R.J.; Bigley, R.E.; Franklin, J.F.; Gandhi, K.J.; Gustafsson, L.; et al. A cross-continental comparison of plant and beetle responses to retention of forest patches during timber harvest. Ecol. Appl. 2016, 26, 2493–2504. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Beese, W. The retention system: Reconciling variable retention with the principles of silvicultural systems. For. Chron. 2002, 78, 397–403. [Google Scholar] [CrossRef]
- Franklin, J.F.; Berg, D.R.; Thornburgh, D.A.; Tappeiner, J.C. Alternative silvicultural approaches to timber harvesting: Variable retention harvest systems. In Creating a Forestry for the 21st Century: The Science of Ecosystem Management; Kohm, K.A., Franklin, J.F., Eds.; Island Press: Washington, DC, USA, 1997; pp. 111–139. [Google Scholar]
- Lindenmayer, D.; Blanchard, W.; McBurney, L.; Blair, D.; Banks, S.; Driscoll, D.; Smith, A.; Gill, A.M. Fire severity and landscape context effects on arboreal marsupials. Biol. Conserv. 2013, 167, 137–148. [Google Scholar] [CrossRef]
- Todd, C.R.; Lindenmayer, D.B.; Stamation, K.; Acevedo-Cattaneo, S.; Smith, S.; Lumsden, L.F. Assessing reserve effectiveness: Application to a threatened species in a dynamic fire prone forest landscape. Ecol. Model. 2016, 338, 90–100. [Google Scholar] [CrossRef]
- Andren, H. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A review. Oikos 1994, 71, 355–366. [Google Scholar] [CrossRef]
- Angelstam, P.; Dönz-Breuss, M.; Roberge, J.-M. Targets and tools for the maintenance of forest biodiversity: An introduction. Ecol. Bull. 2004, 51, 11–24. [Google Scholar]
- Leonard, S.W.; Bennett, A.F.; Clarke, M.F. Determinants of the occurrence of unburnt forest patches: Potential biotic refuges within a large, intense wildfire in south-eastern Australia. For. Ecol. Manag. 2014, 314, 85–93. [Google Scholar] [CrossRef]
- Mok, H.F.; Arndt, S.K.; Nitschke, C.R. Modelling the potential impact of climate variability and change on species regeneration potential in the temperate forests of south-eastern Australia. Glob. Chang. Biol. 2012, 18, 1053–1072. [Google Scholar] [CrossRef]
- Kouki, J.; Hyvärinen, E.; Lappalainen, H.; Martikainen, P.; Similä, M. Landscape context affects the success of habitat restoration: Large-scale colonization patterns of saproxylic and fire-associated species in boreal forests. Divers. Distrib. 2012, 18, 348–355. [Google Scholar] [CrossRef]
- Tilman, D.; May, R.M.; Lehman, C.L.; Nowak, M.A. Habitat destruction and the extinction debt. Nature 1994, 371, 65–66. [Google Scholar] [CrossRef]
- Commonwealth of Australia. Tasmanian Regional Forest Agreement between the Commonwealth of Australia and the State of Tasmania; Department of Agriculture, Fisheries and Forestry: Canberra, Australia, 1997.
- Commonwealth of Australia. The Central Highlands Regional Forest Agreement between the Commonwealth of Australia and the State of Victoria; Department of Agriculture, Fisheries and Forestry: Canberra, Australia, 1998.
- Slade, C.; Law, B. The other half of the coastal State Forest estate in New South Wales; the value of informal forest reserves for conservation. Aust. Zool. 2016. [Google Scholar] [CrossRef]
- Patry, C.; Kneeshaw, D.; Wyatt, S.; Grenon, F.; Messier, C. Forest ecosystem management in North America: From theory to practice. For. Chron. 2013, 89, 525–537. [Google Scholar] [CrossRef]
- Fox-Hughes, P.; Harris, R.; Lee, G.; Grose, M.; Bindoff, N. Future fire danger climatology for Tasmania, Australia, using a dynamically downscaled regional climate model. Int. J. Wildland Fire 2014, 23, 309–321. [Google Scholar] [CrossRef]
- Orr, S.; Gerrand, A. Management Decision Classification: A system for zoning land managed by Forestry Tasmania. Tasforests 1998, 10, 1–14. [Google Scholar]
- Department Natural Resources and Environment (DNRE). Forest Management Plan for the Central Highlands; DNRE: Melbourne, Victoria, Australia, 1998. [Google Scholar]
| Southern Forests (Tas) | North West (Tas) | Central Highlands (Vic) | |
|---|---|---|---|
| Study Area | |||
| Total area (ha) | 174,228 | 73,060 | 81,238 |
| Harvested Patches | |||
| Harvest year | 1960–2015 | 1960–2015 | 1968–2015 |
| No. patches | 1002 | 348 | 897 |
| Total patch area (ha) | 48,128 | 13,836 | 16,251 |
| Mean patch area (ha) | 48 | 40 | 18 |
| Harvest intensity (% area harvested) | 28% | 19% | 20% |
| Structurally Mature Forest Patches | |||
| Total patch area (ha) | 36,138 | 10,800 | 3194 |
| Proportion of study area (%) | 20.7% | 13.8% | 3.9% |
| No. patches | 997 | 721 | 206 |
| Mean patch area (ha) | 36 | 15 | 15 |
| Irregular crown cover 5%–20% | 46% | 63% | 98% |
| 21%–40% | 29% | 26% | 1% |
| 41%–70% | 22% | 10% | 1% |
| 71%–100% | 3% | 1% | 0% |
| Mature Forest Influence (MFI) | Edge Influence (EI) | Landscape Context (LC) | |
|---|---|---|---|
| Definition | Proportion of harvested patch within a particular distance of a structurally mature patch | Proportion of mature patch within a particular distance of harvested patch or road | Percent cover of mature forest in 1000 m buffer around harvested patch |
| Schematic # | 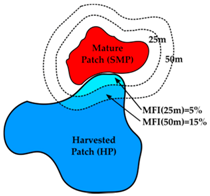 | 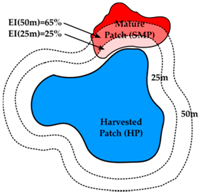 |  |
| Calculations | Proportion of HP within a particular distance (25 m, 50 m, 100 m, 200 m) of (i) SMP; (ii) LTR; (iii) SMP + LTR; or (iv) 1939 + LTR. | Proportion of SMP within a particular distance (25 m, 50 m, 100 m, 200 m) of HP or RD. | Percent cover of (i) SMP; (ii) LTR; (iii) SMP + LTR; or (iv) 1939 + LTR in a 1000 m buffer around perimeter * of HP. |
| Units | % harvested patch area | % mature patch area | % buffer area |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wood, S.W.; Wardlaw, T.J.; Pryde, E.C.; Baker, S.C. Landscape Structure and Mature Forest Biodiversity in Wet Eucalypt Forests: A Spatial Analysis of Timber Production Areas in South-Eastern Australia. Forests 2017, 8, 89. https://doi.org/10.3390/f8030089
Wood SW, Wardlaw TJ, Pryde EC, Baker SC. Landscape Structure and Mature Forest Biodiversity in Wet Eucalypt Forests: A Spatial Analysis of Timber Production Areas in South-Eastern Australia. Forests. 2017; 8(3):89. https://doi.org/10.3390/f8030089
Chicago/Turabian StyleWood, Sam W., Timothy J. Wardlaw, Elizabeth C. Pryde, and Susan C. Baker. 2017. "Landscape Structure and Mature Forest Biodiversity in Wet Eucalypt Forests: A Spatial Analysis of Timber Production Areas in South-Eastern Australia" Forests 8, no. 3: 89. https://doi.org/10.3390/f8030089