Spatial Patterns of Canopy Disturbance, Structure, and Species Composition in a Multi-Cohort Hardwood Stand

Abstract
:1. Introduction
2. Study Area and Methods
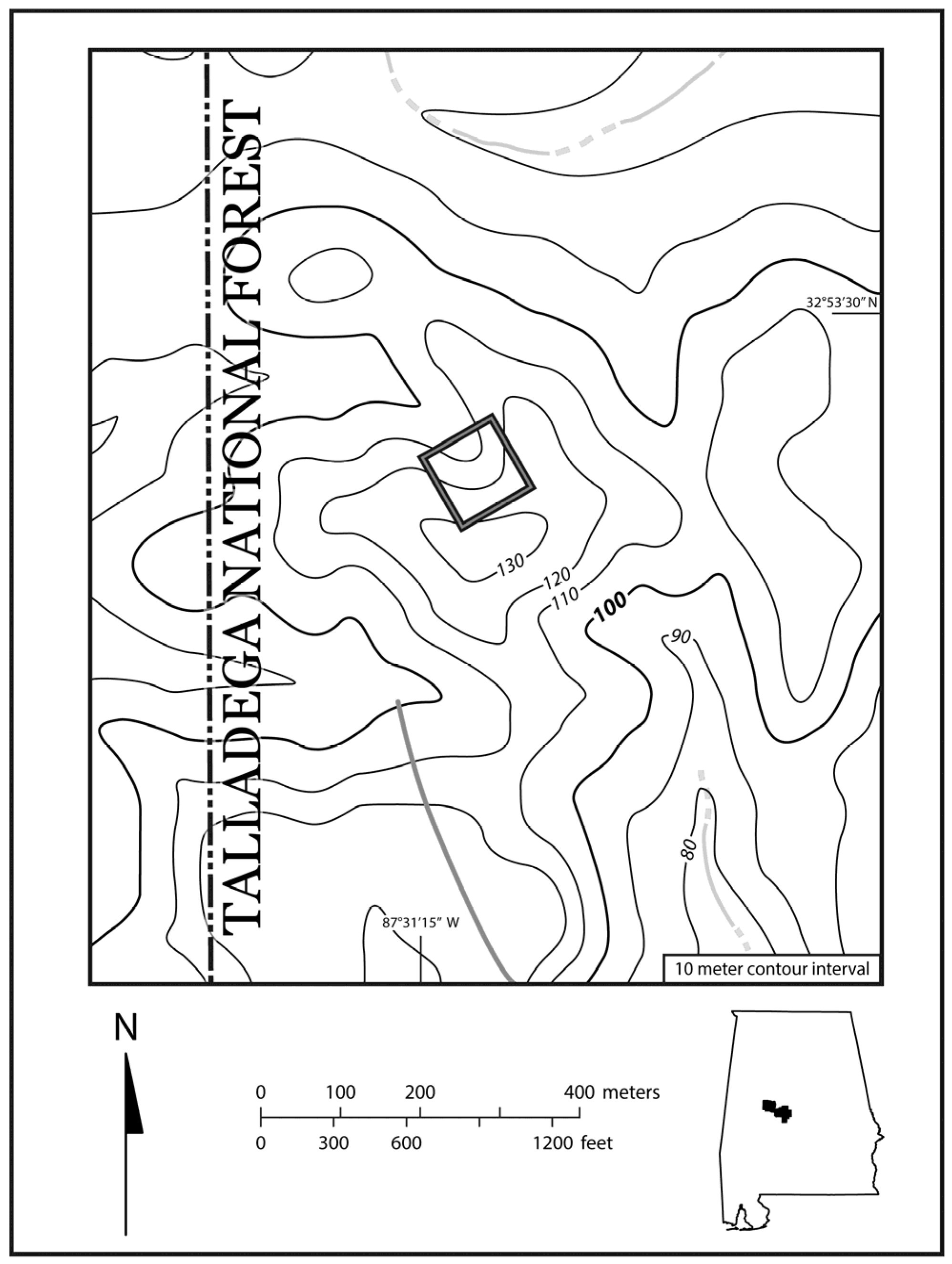
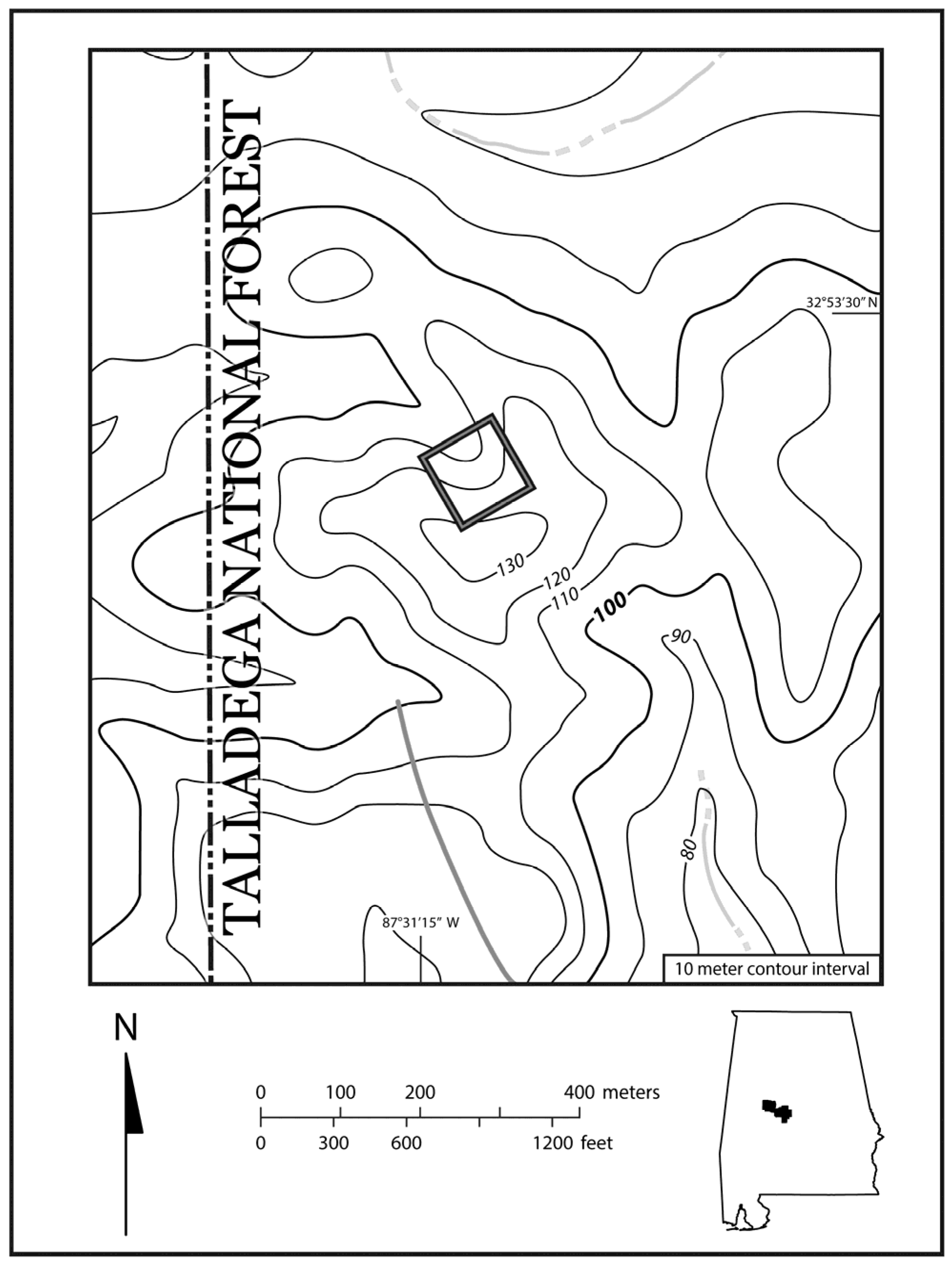
2.1. Study Site
2.2. Field Methods
2.3. Laboratory and Analytical Methods
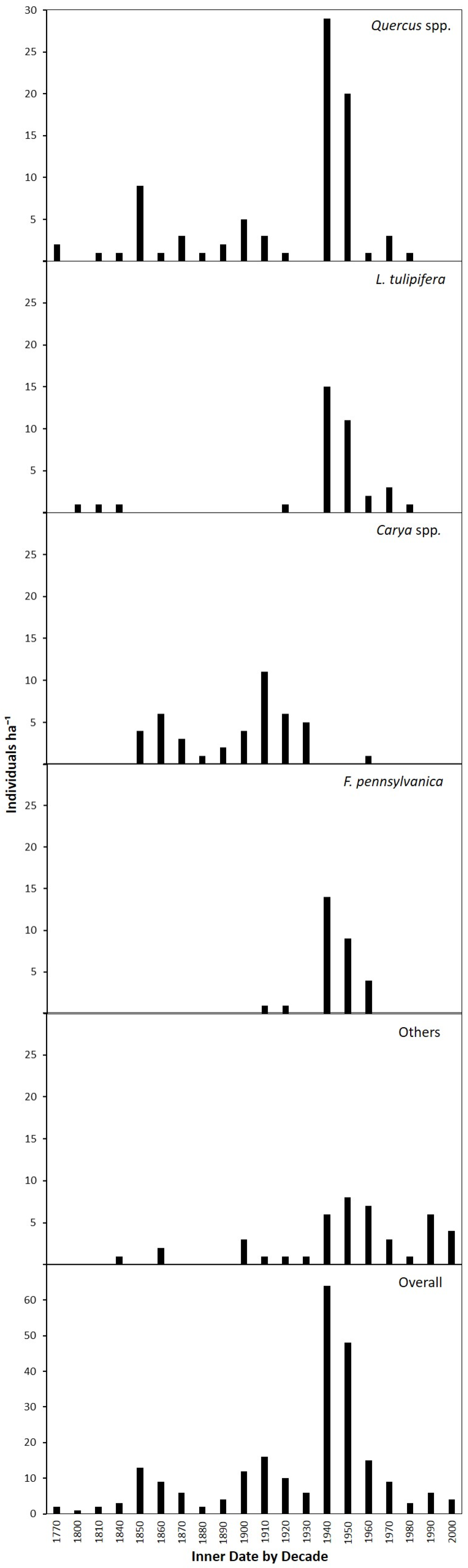
3. Results
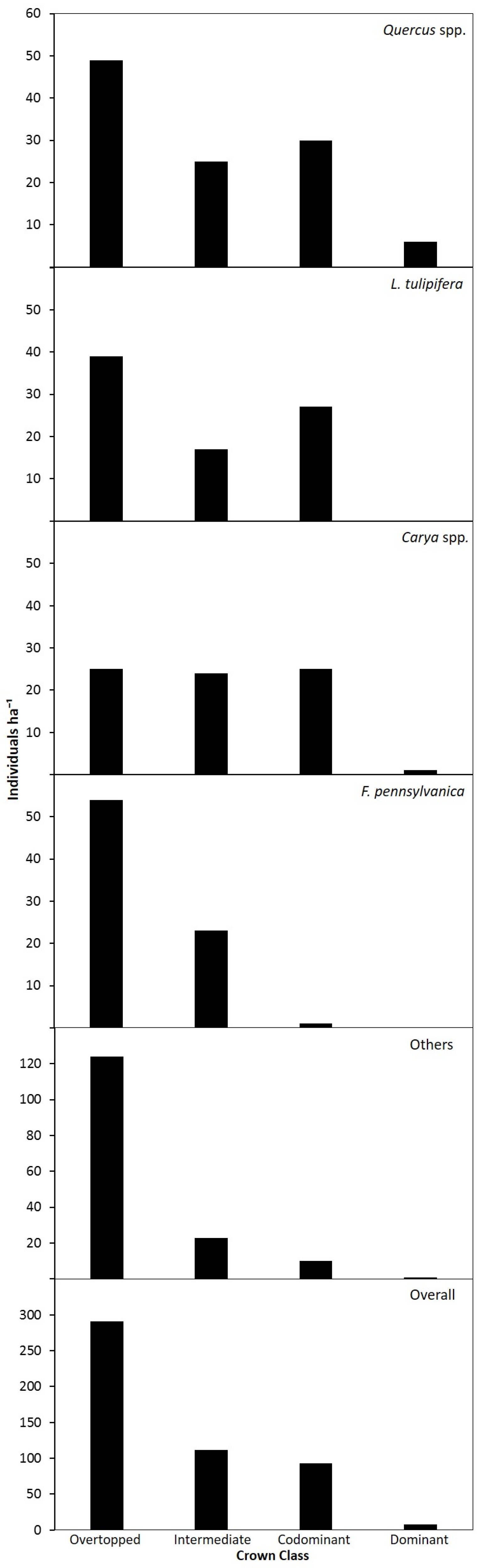
3.1. Species Composition
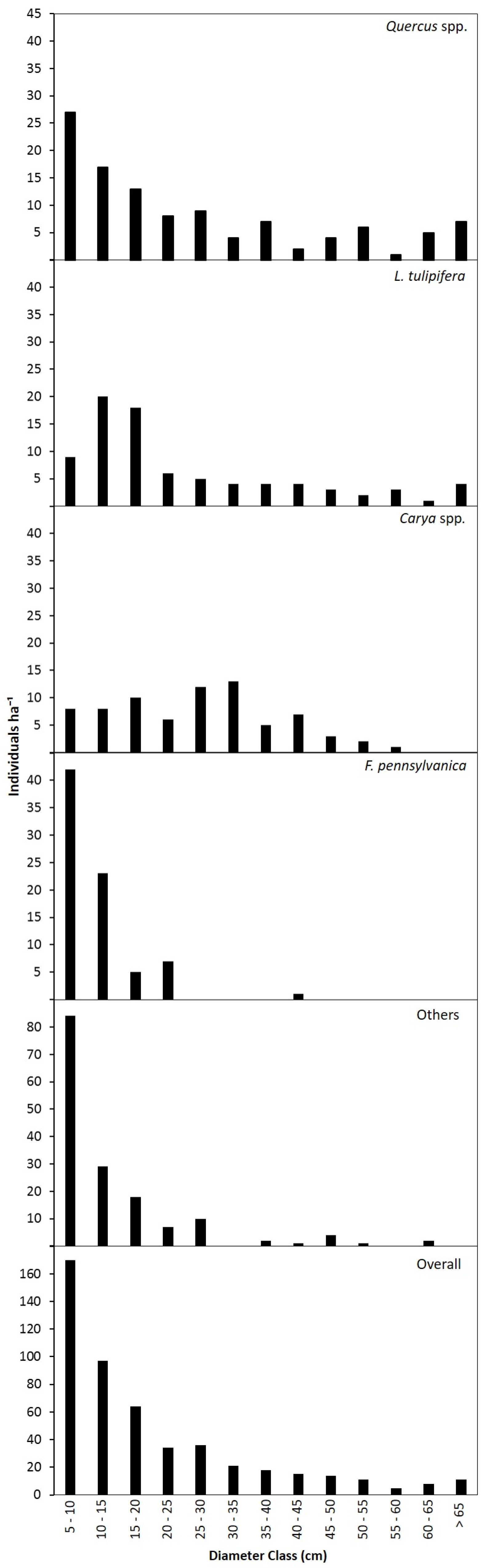
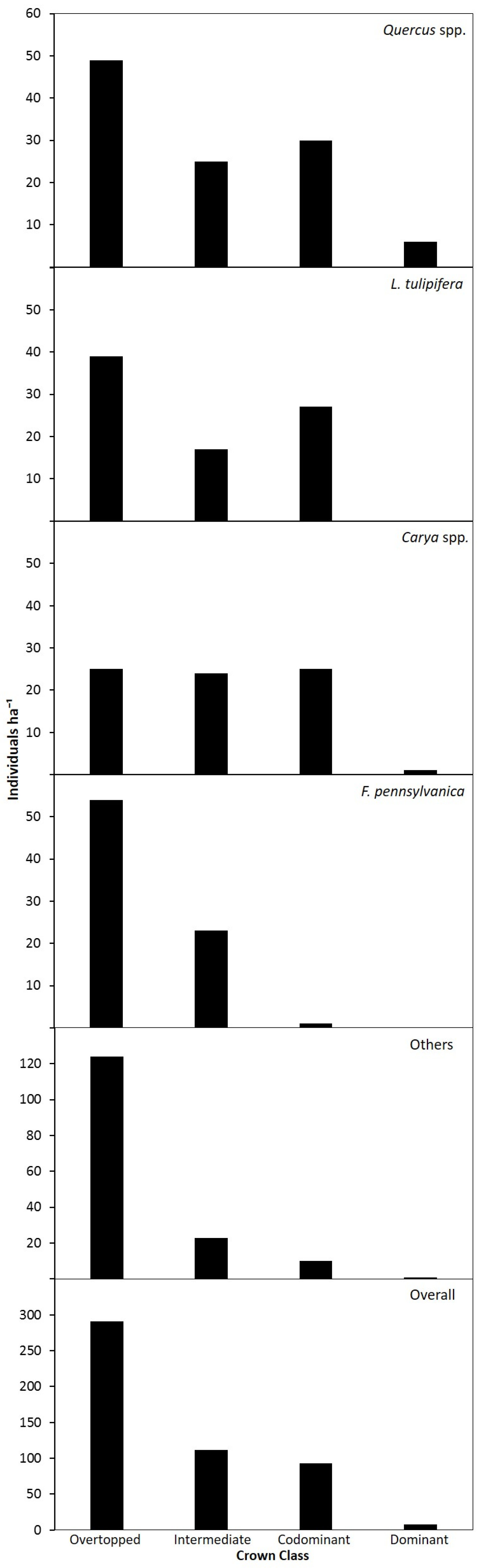
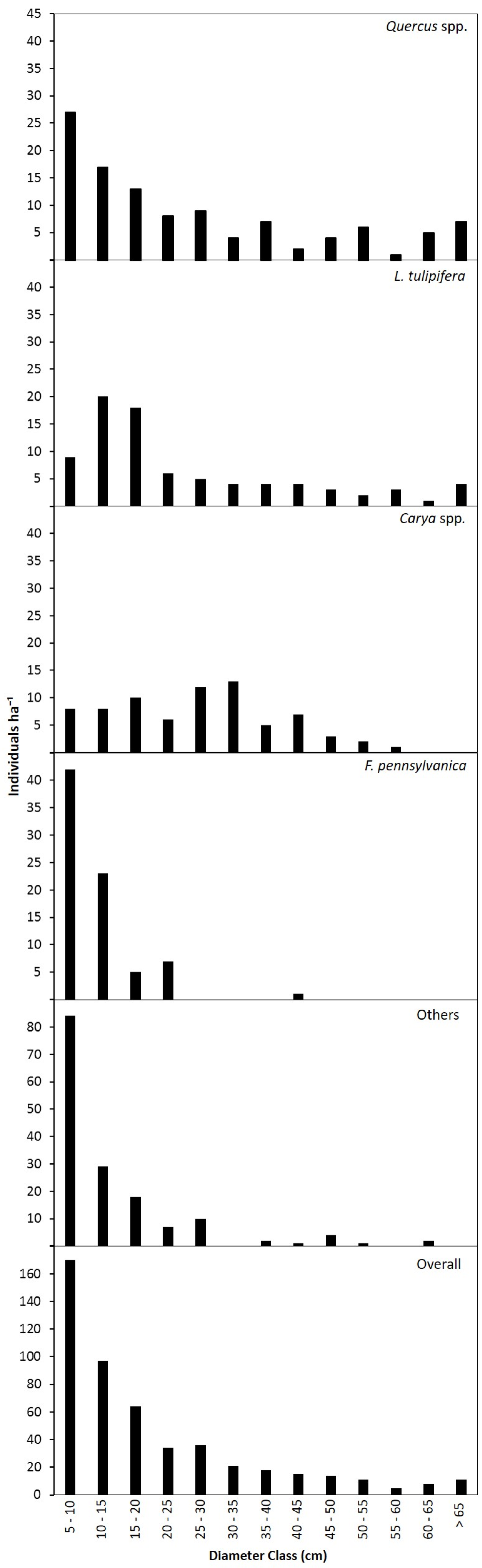
3.2. Canopy and Diameter Structure
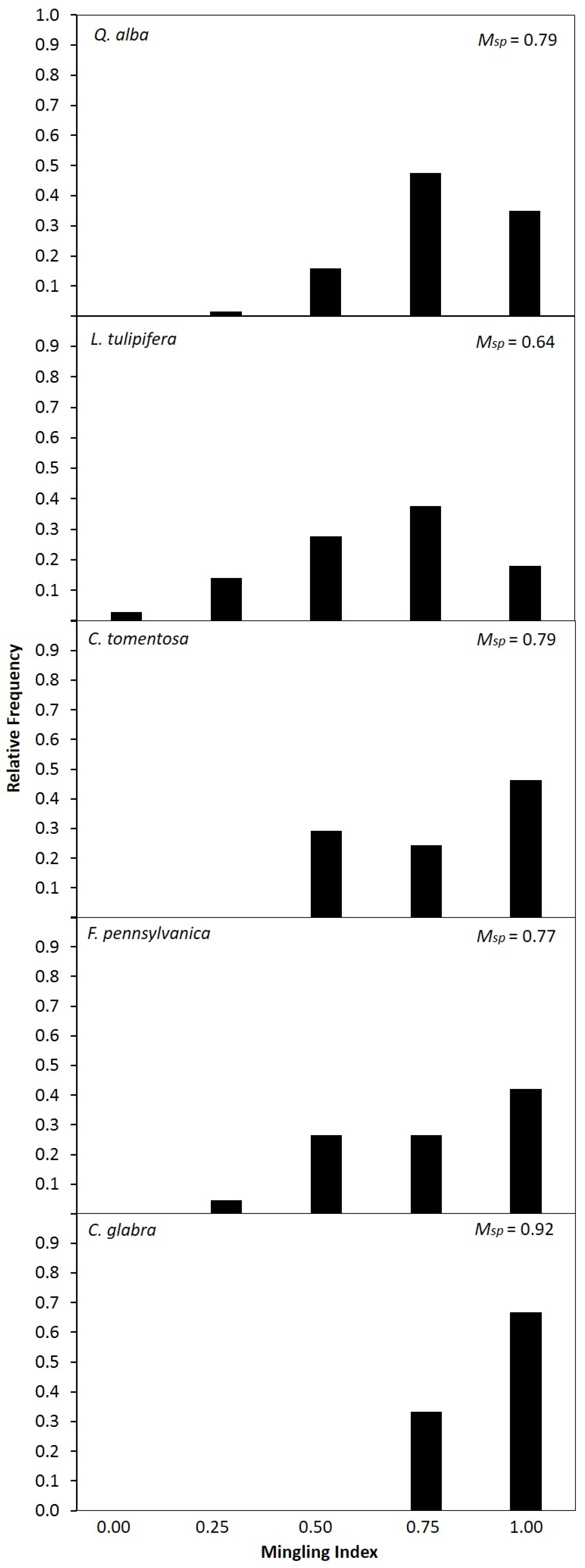
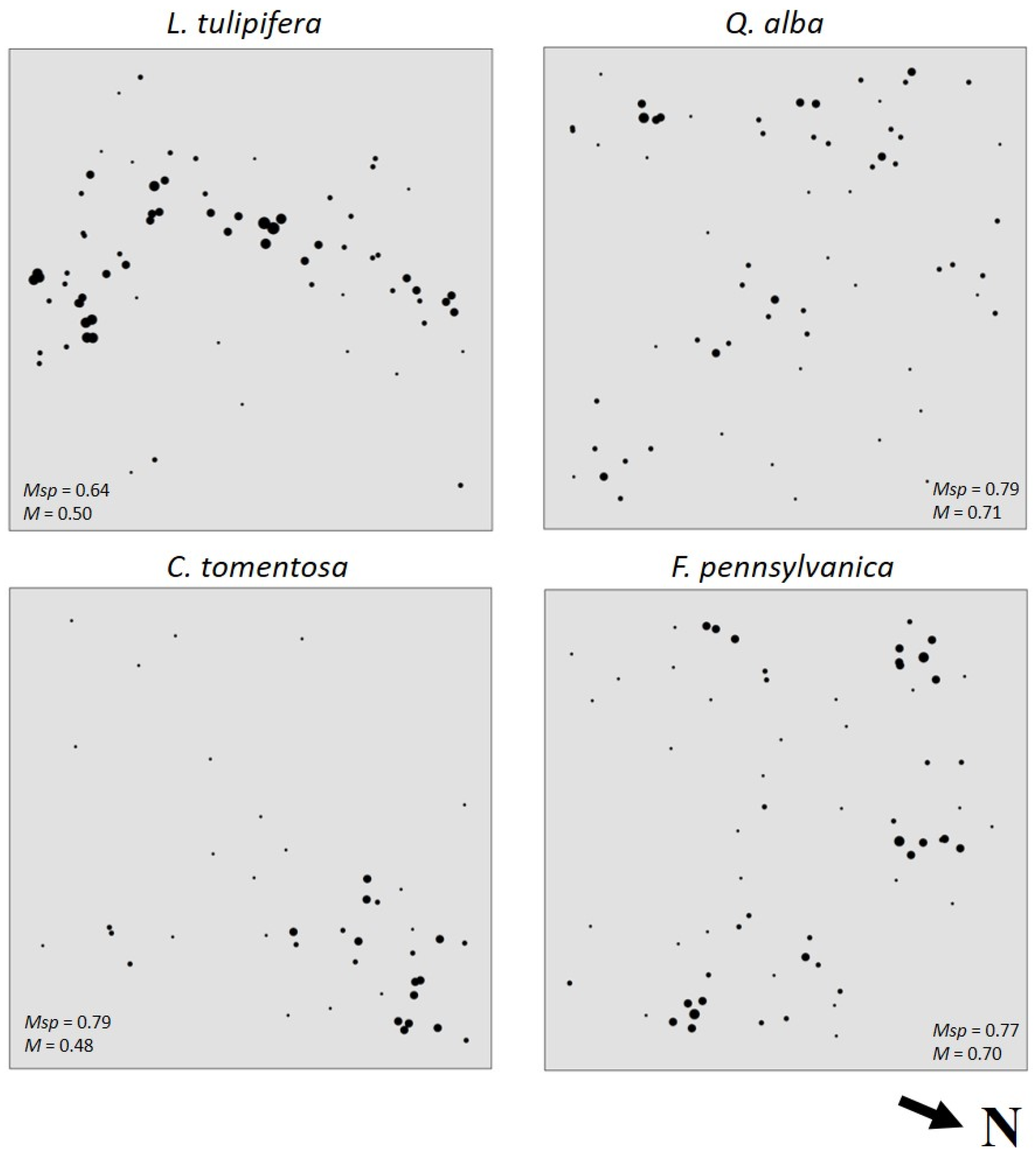
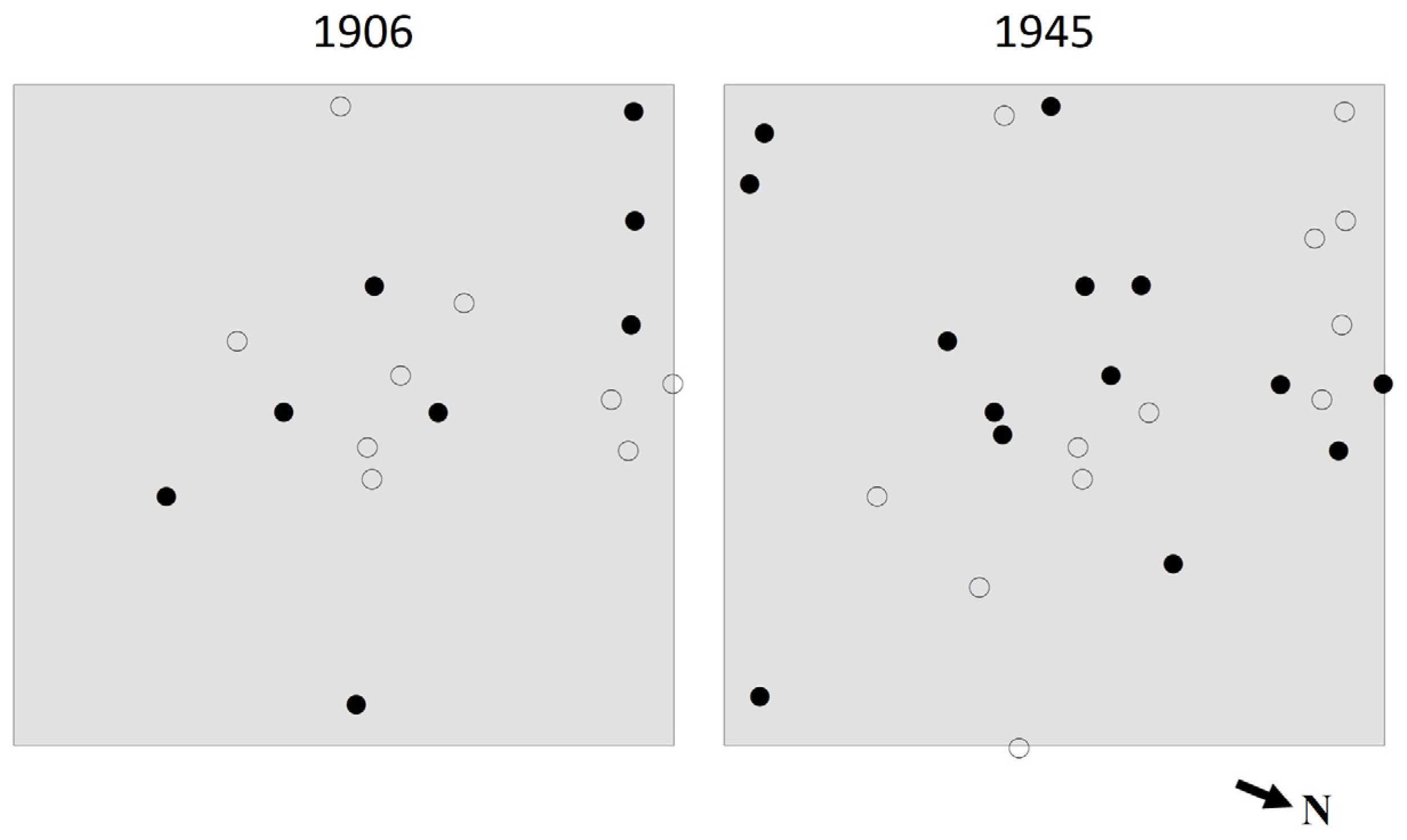
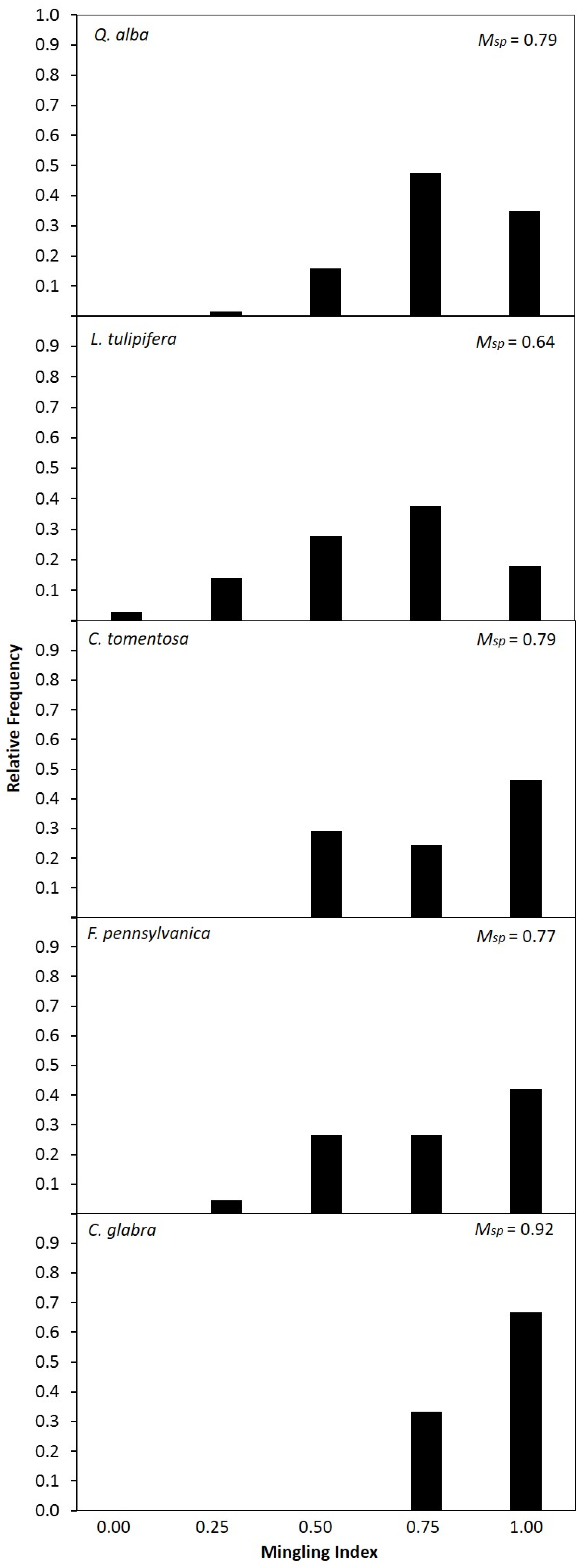
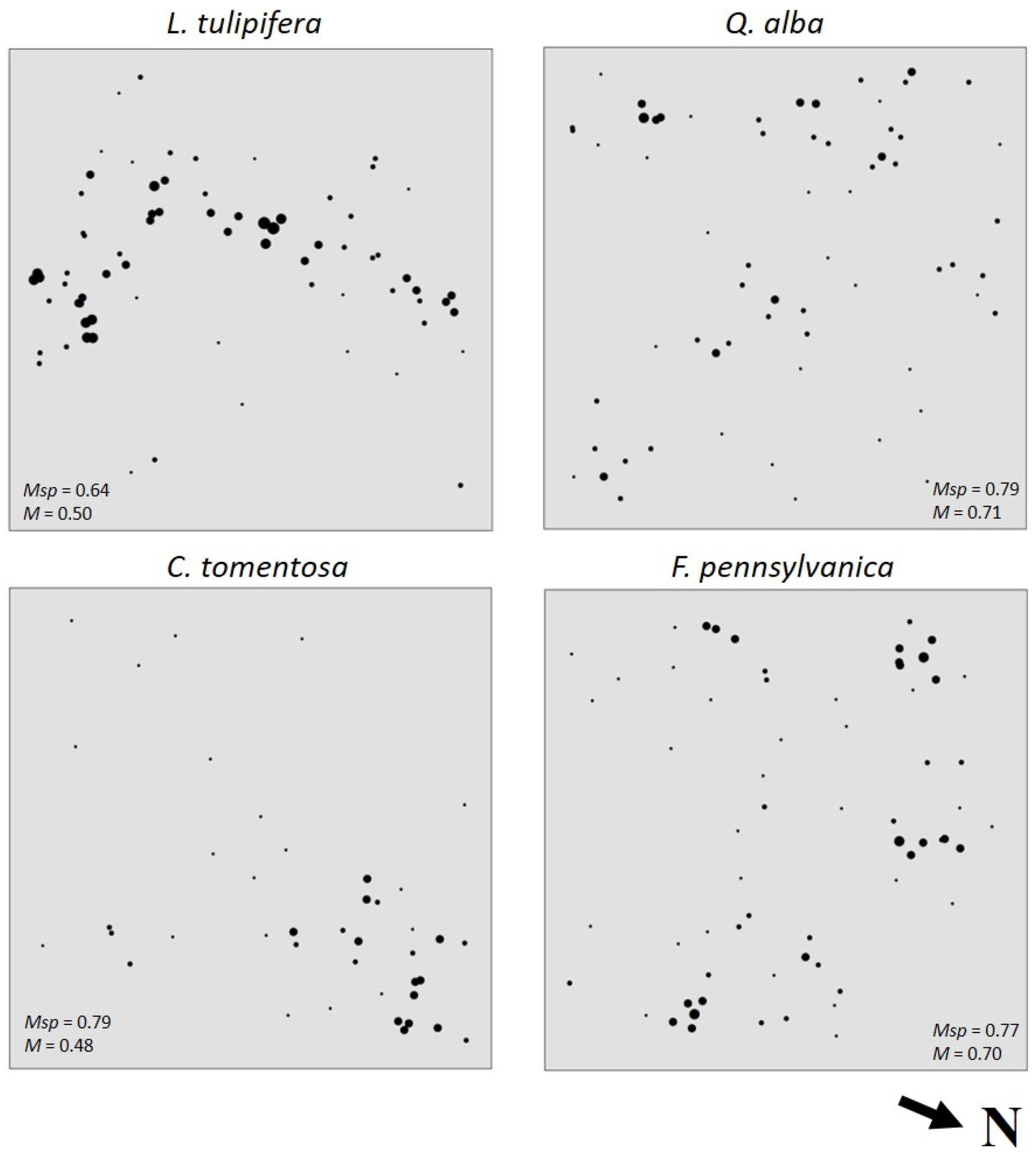
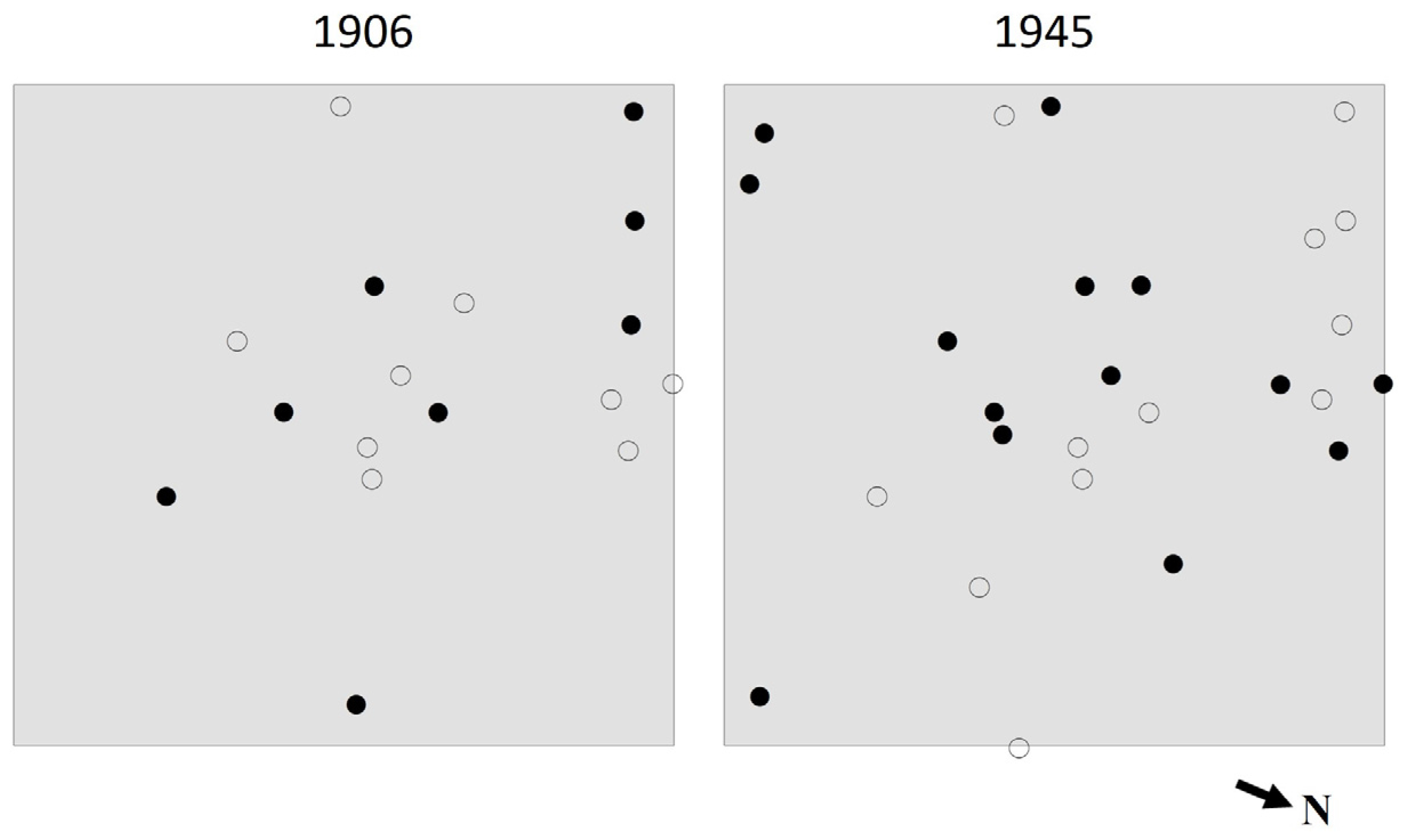
3.3. Species Intermingling and Spatial Patterns
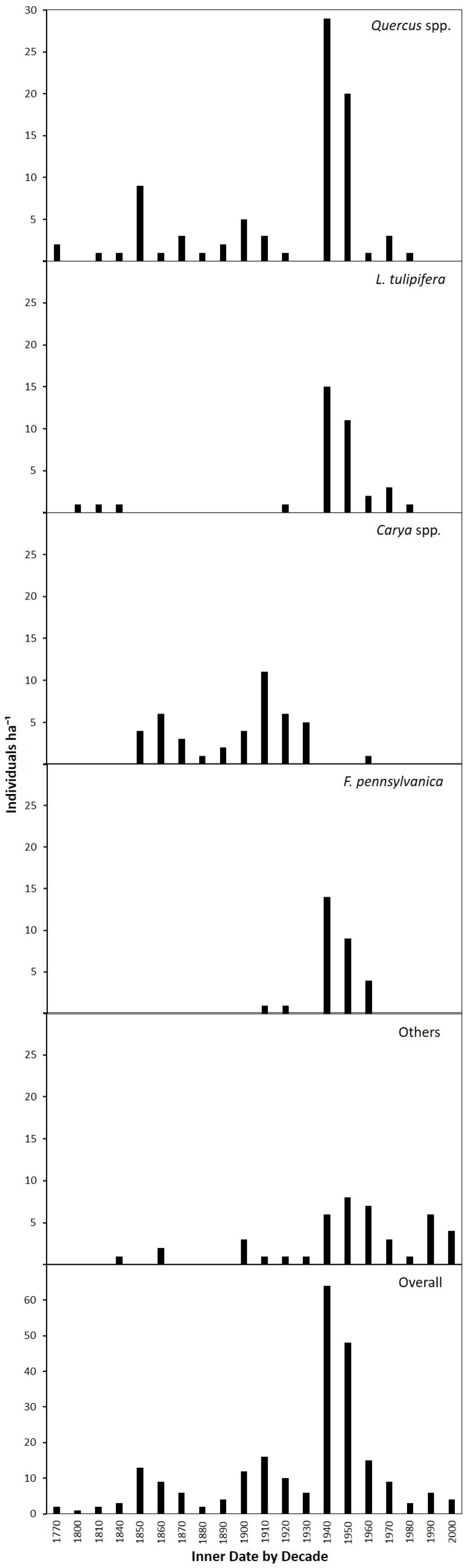
3.4. Age Structure and Canopy Disturbance History
4. Discussion
4.1. Forest Composition
4.2. Forest Structure
4.3. Species Intermingling and Spatial Patterns
4.4. Canopy Disturbance History and Stand Development
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hansen, A.J.; Spies, T.A.; Swanson, F.J.; Ohmann, J.L. Conserving biodiversity in managed forests. BioScience 1991, 41, 382–392. [Google Scholar] [CrossRef]
- Aplet, G.H. Beyond even-vs. uneven-aged management: Toward a cohort-based silviculture. J. Sustain. For. 1994, 2, 423–433. [Google Scholar] [CrossRef]
- Hanson, J.J.; Lorimer, C.G. Forest structure and light regimes following moderate wind storms: Implications for multi-cohort management. Ecol. Appl. 2007, 17, 1325–1340. [Google Scholar] [CrossRef] [PubMed]
- Puettmann, K.J.; Coates, K.D.; Messier, C. A Critique of Silviculture: Managing for Complexity; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef]
- O'Hara, K.L.; Ramage, B.S. Silviculture in an uncertain world: Utilizing multi-aged management systems to integrate disturbance. Forestry 2013, 86, 401–410. [Google Scholar] [CrossRef]
- Halpin, C.R.; Lorimer, C.G. Trajectories and resilience of stand structure in response to variable disturbance severities in northern hardwoods. For. Ecol. Manag. 2016, 365, 69–82. [Google Scholar] [CrossRef]
- Shifley, S.R.; Moser, W.K.; Nowak, D.J.; Miles, P.D.; Butler, B.J.; Aguilar, F.X.; DeSantis, R.D.; Greenfield, E.J. Five anthropogenic factors that will radically alter forest conditions and management needs in the northern United States. For. Sci. 2014, 60, 914–925. [Google Scholar] [CrossRef]
- Long, J.N. Emulating natural disturbance regimes as a basis for forest management: A North American view. For. Ecol. Manag. 2009, 257, 1868–1873. [Google Scholar] [CrossRef]
- Keeton, W.S. Managing for late-successional/old-growth characteristics in northern hardwood-conifer forests. For. Ecol. Manag. 2006, 235, 129–142. [Google Scholar] [CrossRef]
- O’Hara, K. Multiaged Silviculture: Managing for Complex Forest stand Structures; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Churchill, D.J.; Larson, A.J.; Dahlgreen, M.C.; Franklin, J.F.; Hessburg, P.F.; Lutz, J.A. Restoring forest resilience: From reference spatial patterns to silvicultural prescriptions and monitoring. For. Ecol. Manag. 2013, 291, 442–457. [Google Scholar] [CrossRef]
- Dickinson, Y.; Pelz, K.; Giles, E.; Howie, J. Have we been successful? Monitoring horizontal forest complexity for forest restoration projects. Restor. Ecol. 2016, 24, 8–17. [Google Scholar] [CrossRef]
- Oliver, C.D. Forest development in North America following major disturbances. For. Ecol. Manag. 1980, 3, 153–168. [Google Scholar] [CrossRef]
- Martin, W.H. Characteristics of old-growth mixed mesophytic forests. Nat. Areas J. 1992, 12, 127–135. [Google Scholar]
- Johnson, P.S.; Shifley, S.R.; Rogers, R. The Ecology and Silviculture of Oaks, 2nd ed.; CABI: Oxfordshire, UK, 2009. [Google Scholar]
- Seymour, R.S.; White, A.S.; de Maynadier, P.G. Natural disturbance regimes in northeastern North America: Evaluating silvicultural systems using natural scales and frequencies. For. Ecol. Manag. 2002, 155, 357–367. [Google Scholar] [CrossRef]
- Henry, J.D.; Swan, J.M.A. Reconstructing Forest History from Live and Dead Plant Material—An Approach to the Study of Forest Succession in Southwest New Hampshire. Ecology 1974, 55, 772–783. [Google Scholar] [CrossRef]
- Oliver, C.D.; Stephens, E.P. Reconstruction of a Mixed-Species Forest in Central New England. Ecology 1977, 58, 562–572. [Google Scholar] [CrossRef]
- Lorimer, C.G. Methodological considerations in the analysis of forest disturbance history. Can. J. For. Res. 1985, 15, 200–213. [Google Scholar] [CrossRef]
- Lorimer, C.G.; Frelich, L.E. A methodology for estimating canopy disturbance frequency and intensity in dense temperate forests. Can. J. For. Res. 1989, 19, 651–663. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; Wiley: New York, NY, USA, 1996. [Google Scholar]
- Whitney, G.G. From Coastal Wilderness to Fruited Plain: A History of Environmental Change in Temperate North America from 1500 to the Present; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Lorimer, C.G. Historical and ecological roles of disturbance in eastern North American forests: 9000 years of change. Wildl. Soc. Bull. 2001, 29, 425–439. [Google Scholar]
- Burrascano, S.; Keeton, W.S.; Sabatini, F.M.; Blasi, C. Commonality and variability in the structural attributes of moist temperate old-growth forests: A global review. For. Ecol. Manag. 2013, 291, 458–479. [Google Scholar] [CrossRef]
- Fenneman, N.M. Physiography of Eastern United States; McGraw-Hill: New York, NY, USA, 1938; p. 714. [Google Scholar]
- Griffith, G.E.; Omernik, J.M.; Comstock, J.A.; Lawrence, S.; Martin, G.; Goddard, A.; Hulcher, V.J.; Foster, T. Ecoregions of Alabama and Georgia (Color Poster with Map, Descriptive Text, Summary Tables, and Photographs; Map Scale 1:17,000,000); US Geological Survey: Reston, VA, USA, 2001.
- Shankman, D.; Hart, J.L. The fall line: A physiographic-forest vegetation boundary. Geogr. Rev. 2007, 97, 502–519. [Google Scholar] [CrossRef]
- Cox, L.E.; Hart, J.L. Two Centuries of Forest Compositional and Structural Changes in the Alabama Fall Line Hills. Am. Midl. Nat. 2015, 174, 218–237. [Google Scholar] [CrossRef]
- Thornthwaite, C.W. An approach toward rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- PRISM Climate Group. Northwest Alliance for Computational Science and Engineering. Available online: http://www.prism.oregonstate.edu/ (accessed on 13 December 2016).
- USDA NRCS (United States Department of Agriculture, Natural Resources Conservation Service). Web Soil Survey. Available online: http://websoilsurvey.nrcs.usda.gov/ (accessed on 6 December 2016).
- FIA. Forest Inventory and Analysis National Program. 3.0 Phase 3 Field Guide–Down Woody Materials. Available online: https://www.fia.fs.fed.us/library/field-guides-methods-proc/ (accessed on 25 January 2017).
- Buchanan, M.L.; Hart, J.L. A methodological analysis of canopy disturbance reconstructions using Quercus alba. Can. J. For. Res. 2011, 41, 1359–1367. [Google Scholar] [CrossRef]
- Stokes, M.A.; Smiley, T.C. An introduction to Tree-Ring Dating; University of Arizona Press: Tuscon, AZ, USA, 1996. [Google Scholar]
- Orvis, K.H.; Grissino-Mayer, H.D. Standardizing the reporting of abrasive papers used to surface tree-ring samples. Tree Ring Res. 2002, 58, 47–50. [Google Scholar]
- Nowacki, G.J.; Abrams, M.D. Radial-growth averaging criteria for reconstructing disturbance histories from presettlement-origin oaks. Ecol. Monogr. 1997, 67, 225–249. [Google Scholar] [CrossRef]
- Rubino, D.L.; McCarthy, B.C. Comparative analysis of dendroecological methods used to assess disturbance events. Dendrochronologia 2004, 21, 97–115. [Google Scholar] [CrossRef]
- Hart, J.L.; Grissino-Mayer, H.D. Vegetation patterns and dendroecology of a mixed hardwood forest on the Cumberland Plateau: Implications for stand development. For. Ecol. Manag. 2008, 255, 1960–1975. [Google Scholar] [CrossRef]
- Hart, J.L.; Cowden, M.M.; Torreano, S.J.; Vestal, J.P.R. Disturbance, succession, and structural development of an upland hardwood forest on the Interior Low Plateau, Tennessee. Nat. Areas J. 2015, 35, 557–573. [Google Scholar] [CrossRef]
- Lorimer, C.G. Age structure and disturbance history of a southern Appalachian virgin forest. Ecology 1980, 61, 1169–1184. [Google Scholar] [CrossRef]
- Fraver, S.; White, A.S. Identifying growth releases in dendrochronological studies of forest disturbance. Can. J. For. Res. 2005, 35, 1648–1656. [Google Scholar] [CrossRef]
- Hart, J.L.; van de Gevel, S.L.; Grissino-Mayer, H.D. Forest dynamics in a natural area of the southern Ridge and Valley, Tennessee. Nat. Areas J. 2008, 28, 275–289. [Google Scholar] [CrossRef]
- Pommerening, A. Approaches to quantifying forest structures. Forestry 2002, 75, 305–324. [Google Scholar] [CrossRef]
- Kint, V.; van Meirvenne, M.; Nachtergale, L.; Geudens, G.; Lust, N. Spatial methods for quantifying forest stand structure development: A comparison between nearest-neighbor indices and variogram analysis. For. Sci. 2003, 49, 36–49. [Google Scholar]
- Saunders, M.R.; Wagner, R.G. Long-term spatial and structural dynamics in Acadian mixedwood stands managed under various silvicultural systems. Can. J. For. Res. 2008, 38, 498–517. [Google Scholar] [CrossRef]
- Pastorella, F.; Paletto, A. Stand structure indices as tools to support forest management: An application in Trentino forests (Italy). J. For. Sci. 2013, 59, 159–168. [Google Scholar]
- Pommerening, A.; Stoyan, D. Edge-correction needs in estimating indices of spatial forest structure. Can. J. For. Res. 2006, 36, 1723–1739. [Google Scholar] [CrossRef]
- Graz, F.P. The behaviour of the species mingling index Msp in relation to species dominance and dispersion. Eur. J. For. Res. 2004, 123, 87–92. [Google Scholar] [CrossRef]
- Graz, F.P. Spatial diversity of dry savanna woodlands. In Forest Diversity and Management; Hawksworth, D.L., Bull, A.T., Eds.; Springer Science and Business Media: Dordrecht, The Netherlands, 2006; pp. 83–97. [Google Scholar]
- Robison, S.A.; McCarthy, B.C. Potential factors affecting the estimation of light availability using hemispherical photography in oak forest understories. J. Torrey Bot. Soc. 1999, 126, 344–349. [Google Scholar] [CrossRef]
- Busing, R.T. Disturbance and the population dynamics of Liriodendron tulipifera: Simulations with a spatial model of forest succession. J. Ecol. 1995, 83, 45–53. [Google Scholar] [CrossRef]
- Lorimer, C.G. Development of the red maple understory in northeastern oak forests. For. Sci. 1984, 30, 3–22. [Google Scholar]
- Abrams, M.D. The red maple paradox. BioScience 1998, 48, 355–364. [Google Scholar] [CrossRef]
- McEwan, R.W.; Dyer, J.M.; Pederson, N. Multiple interacting ecosystem drivers: Toward an encompassing hypothesis of oak forest dynamics across Eastern North America. Ecography 2011, 34, 244–256. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. Community, edaphic, and historical analysis of mixed oak forests of the Ridge and Valley Province in central Pennsylvania. Can. J. For. Res. 1992, 22, 790–800. [Google Scholar] [CrossRef]
- Richards, J.D.; Hart, J.L. Canopy gap dynamics and development patterns in secondary Quercus stands on the Cumberland Plateau, Alabama, USA. For. Ecol. Manag. 2011, 262, 2229–2239. [Google Scholar]
- White, S.D.; Hart, J.L.; Schweitzer, C.J.; Dey, D.C. Altered structural development and accelerated succession from intermediate-severity wind disturbance in Quercus stands on the Cumberland Plateau, USA. For. Ecol. Manag. 2015, 336, 52–64. [Google Scholar] [CrossRef]
- USDA APHIS (United States Department of Agriculture, Animal and Plant Health Inspection Service). Cooperative Emerald Ash Borer Project: EAB County Detections. Available online: Https://www.aphis.usda.gov/plant_health/plant_pest_info/emerald_ash_b (accessed on 7 December 2016).
- Sobek-Swant, S.; Kluza, D.A.; Cuddington, K.; Lyons, D.B. Potential distribution of emerald ash borer: What can we learn from ecological niche models using Maxent and GARP? For. Ecol. Manag. 2012, 281, 23–31. [Google Scholar]
- Parker, G.R. Old-growth forests of the central hardwood region. Nat. Areas J. 1989, 9, 5–11. [Google Scholar]
- Hart, J.L.; Clark, S.L.; Torreano, S.J.; Buchanan, M.L. Composition, structure, and dendroecology of an old-growth Quercus forest on the tablelands of the Cumberland Plateau, USA. For. Ecol. Manag. 2012, 266, 11–24. [Google Scholar] [CrossRef]
- Kupfer, J.A.; Kirsch, S.W. Heterogeneity of forest characteristics in primary and secondary forest stands on the third Chickasaw loess bluff, Tennessee. Phys. Geogr. 1998, 19, 35–54. [Google Scholar]
- Runkle, J.R. Gap regeneration in some old-growth forests of the eastern United States. Ecology 1981, 62, 1041–1051. [Google Scholar]
- Runkle, J.R. Disturbance regimes in temperate forests. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: New York, NY, USA, 1985; pp. 17–33. [Google Scholar]
- Runkle, J.R. Canopy tree turnover in old-growth mesic forests of eastern North America. Ecology 2000, 81, 554–567. [Google Scholar] [CrossRef]
- Cox, L.E.; Hart, J.L.; Dey, D.C.; Schweitzer, C.J. Composition, structure, and intra-stand spatial patterns along a disturbance severity gradient in a Quercus stand. For. Ecol. Manag. 2016, 381, 305–317. [Google Scholar] [CrossRef]
- Ruffner, C.M.; Abrams, M.D. Relating land-use history and climate to the dendroecology of a 326-year-old Quercus prinus talus slope forest. Can. J. For. Res. 1998, 28, 347–358. [Google Scholar] [CrossRef]
- Abrams, M.D.; Nowacki, G.J. Historical variation in fire, oak recruitment, and post-logging accelerated succession in central Pennsylvania. Bull. Torrey Bot. Club 1992, 119, 19–28. [Google Scholar]
- Holzmueller, E.J.; Gibson, D.J.; Suchecki, P.F. Accelerated succession following an intense wind storm in an oak-dominated forest. For. Ecol. Manag. 2012, 279, 141–146. [Google Scholar] [CrossRef]
- Lafon, C.W.; Naito, A.T.; Grissino-Mayer, H.D.; Horn, S.P.; Waldrop, T.A. Fire History of the Appalachian Region: A Review and Synthesis; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2017; p. 97.
- Hart, J.L.; Buchanan, M.L. History of fire in eastern oak forests and implications for restoration. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, USDA Forest Service General Technical Report GTR-NRS, Springfield, MO, USA, 17–19 May 2011; p. 102.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Density (stems·ha−1) | Relative Density (%) | Dominance (m2·ha−1) | Relative Dominance (%) | Importance | Mingling (Msp) |
|---|---|---|---|---|---|---|
| Quercus alba L. | 79 | 15.7 | 7.1 | 26.8 | 21.3 | 0.79 |
| Liriodendron tulipifera L. | 83 | 16.5 | 6.7 | 25.3 | 20.9 | 0.64 |
| Carya tomentosa (Lam.) Nutt. | 50 | 9.9 | 2.7 | 10.2 | 10.0 | 0.79 |
| Fraxinus pennsylvanica Marshall | 78 | 15.5 | 1.0 | 3.7 | 9.6 | 0.77 |
| Carya glabra (Mill.) Sweet | 25 | 5.0 | 2.5 | 9.4 | 7.2 | 0.92 |
| Oxydendrum arboreum (L.) DC. | 31 | 6.2 | 0.7 | 2.6 | 4.4 | 0.83 |
| Quercus rubra L. | 15 | 3.0 | 1.3 | 4.7 | 3.8 | 0.94 |
| Nyssa sylvatica Marshall | 17 | 3.4 | 1.0 | 3.7 | 3.6 | 1.00 |
| Pinus echinata Mill. | 8 | 1.6 | 1.4 | 5.4 | 3.5 | 1.00 |
| Frangula caroliniana (Walter) A. Gray | 27 | 5.4 | 0.1 | 0.3 | 2.8 | 0.81 |
| Quercus stellata Wangenh. | 7 | 1.4 | 0.8 | 2.9 | 2.2 | 0.75 |
| Cornus florida L. | 17 | 3.4 | 0.1 | 0.4 | 1.9 | 1.00 |
| Fagus grandifolia Ehrh. | 13 | 2.6 | 0.2 | 0.6 | 1.6 | 0.56 |
| Liquidambar styraciflua L. | 13 | 2.6 | 0.1 | 0.5 | 1.5 | 1.00 |
| Quercus velutina Lam. | 7 | 1.4 | 0.4 | 1.4 | 1.4 | 1.00 |
| Sassafras albidum (Nutt.) Nees | 7 | 1.4 | 0.1 | 0.3 | 0.8 | 0.79 |
| Tilia americana L. | 5 | 1.0 | 0.0 | 0.1 | 0.6 | 1.00 |
| Halesia carolina L. | 5 | 1.0 | 0.0 | 0.1 | 0.5 | 0.90 |
| Quercus coccinea Münchh. | 2 | 0.4 | 0.2 | 0.6 | 0.5 | 1.00 |
| Prunus alabamensis C. Mohr | 4 | 0.8 | 0.0 | 0.1 | 0.5 | 1.00 |
| Crataegus sp. | 4 | 0.8 | 0.0 | 0.0 | 0.4 | 1.00 |
| Diospyros virginiana L. | 1 | 0.2 | 0.1 | 0.5 | 0.4 | - |
| Cercis canadensis L. | 3 | 0.6 | 0.0 | 0.1 | 0.3 | 0.75 |
| Ostrya virginiana (Mill.) K. Koch | 1 | 0.2 | 0.0 | 0.0 | 0.1 | 1.00 |
| Acer rubrum L. | 1 | 0.2 | 0.0 | 0.0 | 0.1 | 1.00 |
| Vaccinium arboreum Marshall | 1 | 0.2 | 0.0 | 0.0 | 0.1 | 1.00 |
| Total | 504 | 100.0 | 26.6 | 100.0 | 100.0 |
| Seedlings | Saplings | |||
|---|---|---|---|---|
| Species | Density·ha−1 | Relative Density | Density·ha−1 | Relative Density |
| Acer rubrum | 20 | 0.1 | 0 | 0.0 |
| Aesculus pavia L. | 80 | 0.3 | 10 | 0.6 |
| Asimina triloba (L.) Dunal | 830 | 3.4 | 0 | 0.0 |
| Callicarpa americana L. | 290 | 1.2 | 180 | 10.2 |
| Calycanthus floridus L. | 290 | 1.2 | 0 | 0.0 |
| Carya glabra | 2000 | 8.1 | 0 | 0.0 |
| Carya tomentosa | 1070 | 4.3 | 30 | 1.7 |
| Cercis canadensis | 470 | 1.9 | 40 | 2.3 |
| Celtis occidentalis L. | 1460 | 5.9 | 120 | 6.8 |
| Chionanthus virginicus L. | 0 | 0.0 | 10 | 0.6 |
| Cornus florida | 50 | 0.2 | 0 | 0.0 |
| Crataegus sp. | 60 | 0.2 | 70 | 4.0 |
| Diospyros virginiana | 650 | 2.6 | 50 | 2.8 |
| Frangula caroliniana | 1110 | 4.5 | 150 | 8.5 |
| Fraxinus pennsylvanica | 100 | 0.4 | 20 | 1.1 |
| Halesia carolina | 1280 | 5.2 | 800 | 45.5 |
| Hamamelis virginiana L. | 110 | 0.4 | 10 | 0.6 |
| Hydrangea arborescens L. | 360 | 1.5 | 0 | 0.0 |
| Liriodendron tulipifera | 30 | 0.1 | 10 | 0.6 |
| Nyssa sylvatica | 530 | 2.1 | 60 | 3.4 |
| Oxydendrum arboreum | 0 | 0.0 | 30 | 1.7 |
| Pinus L. | 20 | 0.1 | 0 | 0.0 |
| Prunus alabamensis | 10 | 0.0 | 10 | 0.6 |
| Prunus umbellata Elliott | 10 | 0.0 | 0 | 0.0 |
| Quercus alba | 11,300 | 45.7 | 80 | 4.5 |
| Quercus coccinea | 10 | 0.0 | 0 | 0.0 |
| Quercus falcata Michx. | 110 | 0.4 | 0 | 0.0 |
| Quercus nigra L. | 20 | 0.1 | 0 | 0.0 |
| Quercus rubra | 430 | 1.7 | 0 | 0.0 |
| Quercus stellata | 90 | 0.4 | 0 | 0.0 |
| Quercus velutina | 1040 | 4.2 | 30 | 1.7 |
| Sassafras albidum | 230 | 0.9 | 0 | 0.0 |
| Tilia americana | 360 | 1.5 | 20 | 1.1 |
| Vaccinium arboreum | 20 | 0.1 | 30 | 1.7 |
| Viburnum acerifolium L. | 270 | 1.1 | 0 | 0.0 |
| Total | 24,710 | 100.0 | 1760 | 100.0 |
| Species | Psp | Msp | 1 − Msp | M (Psp/1 − Msp) |
|---|---|---|---|---|
| Liriodendron tulipifera | 0.18 | 0.64 | 0.36 | 0.50 |
| Fraxinus pennsylvanica | 0.16 | 0.77 | 0.23 | 0.70 |
| Quercus alba | 0.15 | 0.79 | 0.21 | 0.71 |
| Carya tomentosa | 0.10 | 0.79 | 0.21 | 0.48 |
| Carya glabra | 0.05 | 0.92 | 0.08 | 0.63 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ford, S.; Kleinman, J.; Hart, J. Spatial Patterns of Canopy Disturbance, Structure, and Species Composition in a Multi-Cohort Hardwood Stand. Forests 2017, 8, 93. https://doi.org/10.3390/f8030093
Ford S, Kleinman J, Hart J. Spatial Patterns of Canopy Disturbance, Structure, and Species Composition in a Multi-Cohort Hardwood Stand. Forests. 2017; 8(3):93. https://doi.org/10.3390/f8030093
Chicago/Turabian StyleFord, Scott, Jonathan Kleinman, and Justin Hart. 2017. "Spatial Patterns of Canopy Disturbance, Structure, and Species Composition in a Multi-Cohort Hardwood Stand" Forests 8, no. 3: 93. https://doi.org/10.3390/f8030093