
A Tree Species Effect on Soil That Is Consistent Across the Species’ Range: The Case of Aspen and Soil Carbon in North America


1
Natural Resources Canada, Canadian Forest Service, Laurentian Forestry Centre, 1055 du P.E.P.S., P.O. Box 10380, Stn. Sainte-Foy, Québec QC G1V 4C7, Canada
2
Department of Wildland Resources and Ecology Center, Utah State University, 5230 Old Main Hill, Logan, UT 84322-5230, USA
*
Author to whom correspondence should be addressed.
Forests 2017, 8(4), 113; https://doi.org/10.3390/f8040113
Submission received: 15 February 2017
/
Revised: 31 March 2017
/
Accepted: 5 April 2017
/
Published: 8 April 2017
(This article belongs to the Special Issue Tree Species, as Major Drivers of Forest Ecosystems Functioning)
Abstract
:Trembling aspen covers a large geographic range in North America, and previous studies reported that a better understanding of its singular influence on soil properties and processes is of high relevance for global change questions. Here we investigate the potential impact of a shift in aspen abundance on soil carbon sequestration and soil carbon stability at the continental scale by conducting a systematic literature review using 23 published studies. Our review shows that aspen’s effect on soil carbon is relatively consistent throughout the species range. Aspen stores less C in the forest floor but similar amounts in the mineral soil relative to conifers. However, a robust set of indicators of soil C stability, for example, degree of organo-mineral associations, proportion of readily-available or labile C estimated during long-term soil incubations or using hot-water extraction, pattern of soil C distribution, and temperature sensitivity of soil heterotrophic respiration, reveals that the soil organic carbon (SOC) stock under aspen is more stable, rendering it more protected against environmental changes and soil disturbances. Therefore, our continental-scale analysis highlights that an increase in the abundance of trembling aspen in North American forests may increase the resistance and resilience of soil C stocks against global changes.
1. Introduction
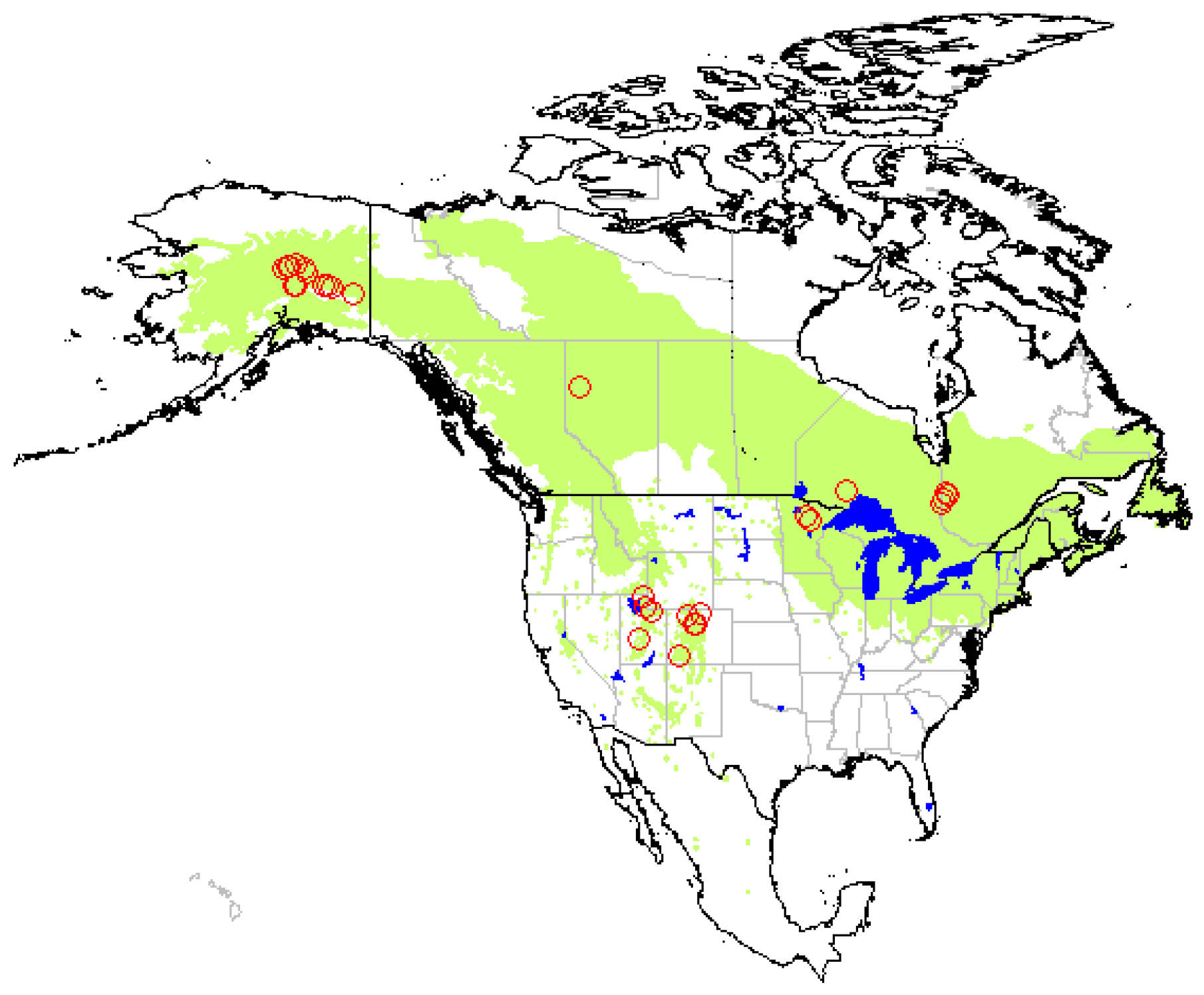
Trembling aspen (Populus tremuloides Michx.) covers a large geographic range in North America (Figure 1), and is found on a great variety of soils, mainly Alfisols, Spodosols, Inceptisols [1], and Mollisols [2,3]. Its abundance within a forest stand has been found to positively affect stand productivity, total aboveground biomass, soil respiration, and nutrient cycling in boreal mixedwood forests of various composition [4,5,6]. In western and midwestern USA, aspen has also been associated with increased levels of understory plant species diversity [5,7]. Enhanced levels of avifauna, mammal, and macroinvertebrate biodiversity have also been reported in aspen stands compared to adjacent conifer stands [8,9]. Furthermore, in the semi-arid western USA, aspen has a positive effect on snow accumulation and water retention [10], an issue of major importance in regions that depend on snow as a water source.
As a pioneer species, aspen has developed mechanisms for quickly occupying suitable areas through fast seed dispersal and vegetative suckering, which are often enhanced by disturbances like fire [11]. Because of its widespread distribution and its positive effects on many ecosystem services, aspen has received considerable attention in the scientific literature, even as early as 1925 [12]. At the southern end of its distribution range, reported cases of aspen decline became a research focus as early as 1942 [13], and this has continued throughout the last century [14,15,16,17]. Overall, herbivory by defoliators, like the forest tent caterpillar and ungulate browsing, changes in disturbance regimes (such as fire exclusion), and a drier climate can affect aspen distribution and abundance. A study by Worrall et al. [18] recently concluded that aspen distribution might change in the future due to aspen disappearing from areas currently on the edge of its climatic niche where water availability will no longer be sufficient to ensure its survival. This is mostly predicted to happen in its southern distribution range, but the northern areas are also expected to see climate-induced changes. While a drier climate will limit the survival of aspen in many areas (especially in the southwest), in areas with more suitable edaphic conditions (mostly in the north), a drier climate will most likely promote more fires [19], thereby, in fact, creating more favourable conditions for aspen stand establishment [20,21,22,23]. Furthermore, increased rates of insect disturbance or forest harvesting can also potentially benefit aspen abundance in such areas [24,25].
Apart from aboveground forest characteristics, the effect of aspen on soil characteristics is less well understood. In view of potential climate and management induced changes in forest composition, the effect of tree species soil organic carbon (SOC) storage has received increased attention in recent years [26,27]. Considering the large amount of carbon stored in soil, understanding species induced SOC patterns can give us crucial insights into the potential impact of a shift in aspen abundance on SOC sequestration and the continental carbon balance.
The objective of this paper is to assess whether trembling aspen has an effect on the amount and nature of soil carbon relative to conifers, and whether this effect is consistent across the species’ continental range, such that it can be extrapolated across North America. We also aim at identifying factors potentially responsible for inconsistencies among studies regarding the magnitude or the direction of the observed aspen effects (none, positive, negative). In our analysis, soil properties and processes other than carbon (e.g., nutrient availability, pH) were also considered when possible, but the main focus was to assess the effect of aspen on soil carbon storage and stability against C losses (i.e., the relative persistence of that C in the soil). To achieve this, we conducted a systematic literature review, defined as a transparent and impartial summary of existing research, using 23 published studies conducted in North America, and summarized the outcomes.
2. Material and Methods
2.1. Study Area
Trembling aspen is the most widely distributed tree in North America, growing from Alaska to central Mexico [1]. The species is widespread across all provinces of Canada, in Alaska, in north-eastern United States as well as in western states, where it grows mainly at high altitudes (Figure 1). The trembling aspen distribution range on the North American continent was taken as the study limits for our literature review.
2.2. Data Collection
The available literature on the influence of aspen on soil carbon was compiled using Scopus, Web of Science, ResearchGate, as well as Google Scholar. Keywords used during the bibliographic search were: aspen, soil carbon, soil organic matter. In order to be included in our analysis, studies had to meet three criteria: (1) report soil organic carbon (or organic matter) concentration, content (or stock), and/or a stability indicator; (2) compare aspen with one or more coniferous species established on similar site conditions (paired-sites preferably); (3) perform statistical analysis and report results. Care was taken to avoid duplicating results from similar datasets reported in different publications. Due to the low number of studies reporting the influence of aspen on comparable SOC variables (concentration vs. content, forest floor vs. mineral horizons, different soil sampling depths, etc.), we were unable to apply statistical methods to our dataset (meta-analysis). Indeed, only six studies reported forest floor C concentration and twelve reported C content. In addition, the different types of soil carbon stability indicators used among studies precluded direct comparison of response ratios. For example, it is impossible to compare Q10 values (proxy for temperature sensitivity of soil respiration and microbial decomposition) with proportion of mineral-associated carbon or with SOC that is more easily decomposable (incubation) or soluble (hot-water extraction). The systematic literature review yielded 23 references, totalling 93 cases (Table A1). The widespread geographic distribution of studies across North America was considered a good coverage of the trembling aspen range (Figure 1).
3. Results
Soil C concentration remains largely unaffected by the presence of aspen relative to conifers. All cases reporting C concentrations in forest floor and mineral soil show no singular effect of aspen, except in one study where forest floor C concentration is lower under aspen than under conifer (Table 1). Forest floor C content is lower under aspen than under conifer in two thirds of the studies (14 out of 21 cases), while in most cases no vegetation effects are detected in the SOC content of the mineral soil (18 out of 25). The stability of SOC, as expressed by the variety of C stability indicators used in the different studies, is for the majority of the cases higher under aspen than under conifer, especially in the mineral soil (13 out of 19). Total SOC storage (FF+MIN) is generally not different among vegetation types. These indicators are degree of organo-mineral associations, proportion of readily-available or labile C estimated during long-term soil incubations or using hot-water extraction, pattern of soil C distribution, and temperature sensitivity of soil heterotrophic respiration. There are also cases (two out of five) pointing toward a higher stability in the forest floor, albeit less pronounced than in the mineral soil. None of the studies show a decline in SOC stability for aspen in either soil layer.
The potential factors that could modulate aspen effect were not fully independent over the full geographic extent of the study. For example conifer species distribution co-varied strongly with climate (Table A1). Nevertheless, the most robust trends that were observed, specifically the absence of a species effect on C concentrations and on mineral soil C content; the negative effect of aspen on forest floor C stocks as well its positive effect on mineral soil C stability, were found over the full geographical range covered by the dataset (Table A1). Moreover, studies with opposite results regarding aspen effects were frequently found within a same geographic area (Table A1). These results suggest that the direction and importance of the aspen effects are dependent on local conditions or specificity of the experimental designs such as site history, local drainage conditions or time since stand establishment. Unfortunately the lack of large scale geographic patterns in aspen effects does not allow to identify factors responsible for the importance or direction of these effects.
4. Discussion
4.1. Aspen Does Not Increase Soil C But Promotes Its Stability
The size of the SOC stock is generally not significantly different between aspen and conifer stands. While aspen tends to store less C in the forest floor relative to conifers, mineral soil C storage is generally similar, making total SOC storage not significantly different among forest types, given the large variability of soil carbon contents in the mineral soil and the relatively low contribution of forest floor to total soil carbon storage. A notable exception to this pattern is found in the Intermountain West, where SOC stocks are higher under aspen in sites in Northern and Southern Utah [3,28]. However, there is strong evidence that aspen promotes SOC stability in the entire soil profile, but especially in the mineral soil. Indeed, C stability in the mineral soil is clearly higher in aspen stands relative to conifer stands in 70% of the studies, with no studies showing a decline in SOC stability associated with the presence of aspen. A multitude of indicators serve as proxies for determining this higher level of C protection in aspen soils that point at the mechanisms of stabilization. The CO2 released during long-term soil incubations at high temperature, and expressed as a fraction of total SOC, serve as the most direct indicator of SOC readily available to microbes (i.e., labile SOC). Numerous studies show that SOC under aspen is indeed less decomposable relative to conifers [3,29,30], although two studies did not detect any differences among stand types [31,32]. A recent multisite study in Utah [28] further supports lower SOC lability under aspen, as indicated by hot water extractions. The temperature sensitivity of soil heterotrophic respiration, relevant for predicting soil C losses in a warmer world, is also lower in aspen than in spruce stands, indicating that enhanced soil C losses in response to climate warming may be less pronounced in aspen stands than in coniferous ones [33].
Across the various SOC fractionation techniques, SOC associated with the clay and silt particles is considered most stable, as the organo-mineral associations confer one of the strongest chemical protection mechanisms against C losses via microbial decomposition, leaching, and erosion [34,35,36,37]. The light fraction or uncomplexed organic matter fraction, on the other hand, has no chemical bond between C and mineral surfaces and is considered to be more responsive to environmental changes and soil disturbances [34,36]. When comparing SOC fractions between aspen and conifer soils, considerably more mineral-associated SOC (up to one-third) is found in the top soil under aspen [28,38] in seasonally dry montane sites of western USA, while conifers store a slightly higher proportion of SOC in the light fraction [28]. Similarly, in the more mesic climate of eastern Canada, aspen store proportionally less SOC in the uncomplexed organic matter fraction [39].
Moreover, the location of the C stored within the soil profile also has implications for long-term stability of SOC. Deep soil layers are generally colder, less aerated, sometimes waterlogged, and generally more nutrient-limited relative to surface layers, thereby limiting microbial biomass, microbial activity, and associated C losses [37]. The SOC located in the forest floor, in contrast, is more vulnerable to environmental perturbations, fire, and management activities [40,41,42]. Thus, soil depth becomes critical in assessing SOC storage potential [43].
While there are cases pointing toward a higher C stability of aspen in the forest floor, albeit less pronounced than in the mineral soil (two cases out of five), it should be noted that these data points are derived from the same study conducted in different regions of Canada with different coniferous species [30].
Collectively, the available data show that aspen does not consistently alter the size of the total SOC stock. Presence of aspen is often associated with a decrease in forest floor C content, and a shift of SOC to lower depths in the soil profile compared to nearby conifers. This is consistent with published synthesis reports indicating that broadleaves store less C in the forest floor layer relative to conifers, while effects in the mineral soil are inconclusive [26,27,44]. Aspen presence, however, facilitates SOC stability, which may increase the resistance and resilience of soil C stocks to global changes.
4.2. Drivers of Aspen Effects on Soil Carbon Stability
4.2.1. Litter Quantity
The productivity of the forest ecosystem and the related fluxes of detritus from various sources into the soil may influence SOC accumulation and stabilization. Aspen is often characterized by higher productivity than other species growing in the same environment [6,45], resulting in a greater influx of aboveground and belowground detritus into the soil. A recent comparative study in Utah indicates annual overstory foliage C inputs of 724 ± 175 kg C·ha−1 for aspen vs. 475 ± 82 kg C·ha−1 for conifers [28]. This is similar to observations in central-eastern Canada, comparing total aboveground inputs (foliar + woody litter): in the Ontario sites, average annual inputs are 1325 ± 36 kg C·ha−1 for aspen, 1136 ± 67 kg C·ha−1 for jack pine, and 1119 ± 51 kg C·ha−1 for mixed coniferous stands, whereas in the Quebec sites, annual inputs are 1233 ± 57 kg C·ha−1 for aspen and 926 ± 129 kg C·ha−1 for black spruce stands [30]. While litterfall under aspen is substantially higher than under conifers, the link between litterfall quantity and SOC stability in the mineral soil is not straightforward. In fact, considering the growing evidence of higher substrate quality positively affecting C stabilization in mineral soil (e.g., [46,47]), it is difficult to differentiate between the effects of quantity and quality when comparing aspen and conifer. In addition, doubling litter inputs without changing litter quality in a 20-year controlled experiment had no effect on SOC stability [48] in a Northern hardwood forest, suggesting that litter quality is a stronger driver than litter amounts.
Much less is known about root detritus inputs. Fine root production constitutes an important portion (one-third to three-quarters) of annual net primary productivity (), and due to their short lifespan, roots continuously contribute to the soil organic matter () pool [49]. Aspen exhibits large intraspecific variation in root morphological traits due to genetic, biotic, and abiotic factors [50,51,52]. When comparing aspen fine root biomass to that of conifers, the findings are not consistent. For example, Bauhus and Messier [53] and Laganière et al. [6] show no differences between aspen and conifer stands in the Canadian boreal forest, as does Ayres et al. [54] for the top 10 cm of soil at a site in Colorado, while Boča and Van Miegroet [28] report much higher fine root biomass under conifers in Utah.
In the study by Steele et al. [51], dead roots comprise from 24% to 47% of total fine root biomass, and fine root turnover is not different between aspen, jack pine, and black spruce. Since soil conditions were not similar among stands, these results must be evaluated with caution. Boča and Van Miegroet [28] report that dead root biomass comprises 32% of aspen and 36% of conifer total fine root biomass, while Laganière et al. [6] indicate a higher proportion of dead vs. total fine root biomass under aspen stands (29%) relative to jack pine (18%) or mixed conifer stands (17%). Finér et al. [55] report that aspen fine roots decompose significantly faster than those of adjacent conifers. Block et al. [56] also reported that aspen fine roots are relatively short-lived, with lifespans ranging from 30 to 300 days. Considering similar or lower total root biomass (although with variable proportion of dead roots) and the similar turnover rates (root production/total root biomass), root detritus input per se is unable to explain the more stable SOC in aspen soils.
4.2.2. Litter Quality
The chemistry of aboveground and belowground detrital inputs varies among tree species and associated understory, and can affect SOC stability via many mechanisms, including biochemical recalcitrance or chemical sorption. When aspen foliage or root litter is compared to that of conifers, a generally consistent pattern emerges at the continental scale, pointing toward higher quality of C inputs under aspen. Indeed, the C/N ratio, the acid-insoluble residue or Klason lignin, as well as the lignin monomer V phenol are significantly lower in aspen foliage litter than in conifer needles [28,54,57,58,59], although litter chemistry may vary depending on biotic and abiotic conditions [60,61]. Lower C/N ratio of aspen root litter also indicates higher detritus quality and faster degradation potential [28,29]. Multiple experiments have shown that litter from aspen decomposes at a higher rate than that from conifers, at least in early stages of decomposition [59,62]. Also, the forest floor material under aspen has been calculated to have a lower mean residence time than litter under conifers [28,63]. It has been hypothesized that the faster a litter is decomposed the more recalcitrant and complex the remaining organic matter will be [64,65]. Indeed, Giardina et al. [29] reported that high quality aspen litter can eventually result in low quality SOC relative to lodgepole pine, as indicated by cumulative carbon emissions in incubated soils from northern Colorado. Long-term aspen-conifer foliar litter decomposition experiments also tend to support this hypothesis. Aspen foliage initially decomposes at a faster rate than conifer litter, but decomposition rates converge after a few years, so that mass remaining of aspen is comparable to conifer species or higher in some cases [57,62]. While the quality of litter inputs affects SOC stability in the forest floor via recalcitrance, other mechanisms may operate more indirectly, conferring greater stability to SOC in the mineral soil under aspen.
A striking difference between aspen and conifer forests observed in the northern regions is the higher pH and higher Ca content of aspen litter, generating higher soil pH and exchangeable base cation conditions in the vicinity of aspen trees [4,66,67]. Redistribution of soil nutrients from relatively deep mineral soil horizons to the upper forest floor layer via an abundant litterfall has often been documented, and aspen has been identified as a Ca pump [68,69]. Calcium can play a role in aggregate stability [36,70]. In addition, higher Ca levels have been found to reduce the output of dissolved organic carbon (DOC) and mineralizable SOC [71,72].
Moreover, the general higher palatability, pH, and nutrient contents of broadleaf litter relative to conifer needles are favourable to macrofauna [73,74]. For example, a microcosm experiment demonstrated that aspen litter, as well as humus derived from aspen trees, was more favourable to earthworms [59]. Earthworms participate actively in the formation and stabilization of soil aggregates [75,76,77]. Therefore, the more favourable soil conditions for macrofaunal organisms under aspen may be conducive to greater SOC stabilisation. It is noteworthy that these species-induced soil chemistry effects are not observed in the southern part of the aspen range, in seasonally dry montane sites. The lack of meaningful differences in pH of aspen and conifer soils in the Intermountain West can potentially be explained by the lack of excessive leaching and base stripping in these seasonally dry soils, or the input of dust from the surrounding desert areas [78]. Also, as noted by the lack of any sightings of earthworms in this area (e.g., [54]), earthworm activity does not seem to play a role in SOC redistribution and stabilization at these locations, probably due to the dry soil conditions.
Finally, higher quality substrates, while leading to faster forest floor decomposition rates, also could result in higher microbial utilization, and could, therefore, contribute to greater SOC stabilisation within the stable mineral-associated fraction due to higher microbial biomass, and more microbial by-products [79]. Indeed, spectral analysis of Utah soils seems to indicate that the C compounds of mineral-associated SOC are rather simple in structure (e.g., polysaccharides rather than aromatics) and likely of microbial origin (Román Dobarco, Unpublished data; see S1 for Materials and Methods).
Thus, in the long-term, SOC derived from high-quality litter can result in greater SOC storage and stabilization. The documented faster decomposition of aspen foliar and root litter, potentially due to a lower C/N ratio and lower acid-insoluble residues, coupled with the findings that aspen soils lose less SOC through heterotrophic degradation during incubations [3,29,30] support this hypothesis for aspen-conifer forests.
4.2.3. The Importance of Understory
Another important aspect of tree species influence on SOC stability is mediated through the associated understory cover. Understory plants growing in aspen forests can constitute an important above- and belowground C flux to the soil system [28]. Aspen stands have been shown to support higher understory plant species biomass, diversity, and productivity compared with adjacent conifers [6,7,80]. Understory composition can have a strong impact on soil characteristics that may be even greater than that imparted by overstory composition. For example, the bryophyte layer usually established under conifers is a major constituent of soil organic matter in boreal forests [81,82,83]. Bryophytes alter microclimate conditions by increasing soil moisture content, and reducing soil temperature as well as litter decomposition rate [84,85]. An increasing abundance of aspen in conifer-dominated forests has been associated with a decrease in bryophyte abundance, reduction of the forest floor thickness and soil moisture, and an increase in soil pH [73,86]. The development of a bryophyte derived organic layer under boreal conifer stands creates a SOC reservoir of low stability. Indeed, in spite of remarkably low decomposition rates [87,88,89], bryophyte derived organic layer has little contact with mineral particles because, unlike vascular plants, it does not grow roots into the mineral soil and hence there is little possibility for organo-mineral interactions [39]. In addition, the accumulation of this layer can generate cool and wet soil conditions that slow down organic matter decomposition [30]. However, such conditions that maintain SOC may not be resilient to a disturbance or a change in species composition that would prevent bryophyte growth.
Interestingly, the above-mentioned effects mediated by the associated understory vegetation could not easily have been observed in common garden studies or in intensive plantations, where the understory plant composition is usually strongly altered. This further underscores the high value and the complementary role of using natural forests or extensive plantations when studying the effect of tree species on soil properties and processes.
4.2.4. Rooting Pattern and Rhizosphere Processes
The vertical distribution of root biomass is closely linked to the distribution of SOC with depth [90]. Therefore, differences in rooting depth patterns among tree species may affect the vertical placement of SOC within the soil profile, with implications for SOC stability. While not much data are available on rooting patterns, Strong and La Roi [91] documented the root-system morphology of some boreal species, showing that aspen roots are well distributed throughout the soil profile and can reach 1 m in depth. By comparison, the roots of black spruce rarely go beyond a depth of 30 cm, and most of them are located at the organic-mineral interface. This may explain, in part, why the presence of aspen is associated with a decrease in forest floor C content and proportionally higher amounts of SOC in deeper, more protected soil horizons. However, in Utah, Boča and Van Miegroet [28] found root biomass C to be positively correlated with the light fraction (i.e., the less stable fraction of SOC).
We are not aware of a study that has explored the extent to which aspen and conifer trees directly affect soil C turnover and microbial biomass through their rhizodeposits. Higher specific root length (SRL) of aspen compared to conifers [50,51,53,92,93] suggests a higher volume of soil exploited and affected by rhizosphere processes like root exudation and detritus input [94]. This could suggest more dynamic belowground processes under aspen.
Root dynamics are affected by the type of associated mycorrhiza; for example, the decomposition rate of ectomycorrhizal (ECM) roots has been reported to be slower than that of non-ECM roots [95]. Most conifers have ECM associations, while aspen have ECM and arbuscular mycorrhizal (AM) associations, with ECM dominating in the topsoil closest to the litter layer, and AM more dominant in the deeper soil [96]. Brzostek et al. [97] found differences in root exudation and form of N produced when comparing two ECM and two AM tree species. The quantification and characterization of root exudates, and the microbial activity around them, as well as detritus turnover, and the interactions between rhizosphere and detritusphere, present directions for future research aimed at better understanding the mechanisms of how aspen belowground processes affect the formation and preservation of more stable SOC than conifers.
4.3. Factors Responsible for Inconsistencies
From the available data, we are able to detect a clear and consistent pattern of enhanced soil C stability in aspen soils throughout North America, and several potential drivers of SOC stability can be identified. At the same time, it appears that patterns of SOC stabilization can vary geographically, suggesting that ecosystem drivers of SOC storage and stabilization are modulated by several abiotic factors, in particular climate and intrinsic soil properties (geology, texture, soil depth) in line with the concept of interacting soil forming factors [98].
The role of climate can play out in many ways. In the western US water is the main limiting factor of aspen productivity, while it is temperature at its northern range [99]. This has several implications. For example, the role of aspen as a calcium pump and the higher concentration of this element in fresh litter relative to that of conifer litter, which can enhance soil pH, in turn affecting the sorption and desorption of organic compounds [100], is not observed in the dry portion of its range. Because of the limited vertical water movement in this seasonally dry area, soils generally have abundant Ca reserves and a high pH, and very few differences emerge between aspen and conifer soil pH and base saturation [3,32]. On the other hand, indicators of litter quality, such as higher nutrient to C ratios and lower lignin concentrations for aspen, are clearer in the western US [28,54]. Woldeselassie [101] further observed that even within the same watershed in northern Utah, SOC stocks in the mineral soil under aspen can vary considerably with aspect, presumably due to microclimate differences (temperature, moisture availability), and their impact on C inputs and losses through decomposition. Specifically, SOC to a depth of 40 cm was significantly different (p < 0.05) among transects with west and north facing aspects, which were wetter and had lower soil C stocks; while the drier south and east facing sites had the highest SOC. This suggests that drier conditions in this region in effect can favour greater C storage. By contrast, in the northern regions, the development of a moss layer is common under conifers and has a temperature isolating effect on soil, which delays the degradation of organic matter, slowing down its microbial transformation towards more stable forms—a mechanism not possible in dry regions lacking a bryophyte cover.
Geographic differences in forest productivity rates are an obvious explanatory factor for the variability in overall SOC storage among the studies used in this analysis. Differences in forest structure within the same site also seem to influence SOC storage, whether directly through differences in productivity or indirectly through its influence on understory abundance or composition. This is illustrated in the studies from Northern Utah and Southern Utah, which showed that aspen tree density or tree size (an index for canopy openness) significantly impact SOC storage under pure aspen [101], while mineral-associated (stable) SOC concentration is positively correlated with aspen abundance in mixed aspen, as expressed by live basal area of aspen [38].
Mineral soil characteristics could also influence the tree species effect. Román Dobarco and Van Miegroet [38] showed that soil mineral characteristics can override or amplify vegetation effects, as they observed that differences in C stocks between aspen and conifers were most pronounced at high silt and clay contents (40–70% clay + silt), while differences remained less pronounced in coarser soil textures (<40% clay + silt). Boča and Van Miegroet [28] showed a positive correlation between C concentration in the soils and extractable Fe and Al that could explain differences in SOC pools between a northern and southern Utah site. Overall, these findings underscore the interplay between abiotic (edaphic) and biological factors in creating stable SOC pools.
5. Conclusions
Our systematic literature review shows that aspen effect on soil carbon is relatively consistent throughout the species range. It shows that aspen stores less C in the forest floor but similar amounts in the mineral soil relative to conifers. However, the expression of a vegetation effect on mineral SOC storage may be constrained geographically by the importance of other biotic and abiotic factors. Nevertheless, a robust set of indicators of soil C stability, such as degree of organo-mineral associations, proportion of readily-available or labile C estimated during long-term soil incubations or using hot-water extraction, pattern of soil C distribution, and temperature sensitivity of soil heterotrophic respiration, reveals that SOC under aspen is more stable and would be thus more protected against environmental changes and soil disturbances. Therefore, our continental-scale analysis highlights that an increase in the abundance of trembling aspen in North American forests could be beneficial to the resistance and resilience of belowground soil C stocks against global changes. Disentangling the relative role of vegetation vs. abiotic effects on SOC stabilization, or identifying and quantifying the relative influence of individual drivers, remains a challenge that should be taken up by scientists.
Supplementary Files
Supplementary File 1Acknowledgments
We thank Julien Beguin for assistance with the generation of Figure 1 and Heather Alexander for providing coordinates for the Alaska sites. We thank Laurent Augusto for the opportunity to submit an invited paper for this special issue of Forests. We are grateful to three reviewers whose constructive comments significantly improved this manuscript.
Author Contributions
J.L., D.P., and H.V.M. designed the study. J.L. and A.B. conducted the literature review and collected the data. All authors wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
Table A1.
Description of the studies included this systematic literature review and summary of results. Aspen causes either an increase (+), a decrease (−), or no change (0) in forest floor (FF) and mineral (MIN) soil C concentration, content, and stability relative to conifers. When a similar effect (+, −, or 0) is observed more than once per reference, the number of cases showing this effect is indicated in brackets next to the effect sign.
Table A1.
Description of the studies included this systematic literature review and summary of results. Aspen causes either an increase (+), a decrease (−), or no change (0) in forest floor (FF) and mineral (MIN) soil C concentration, content, and stability relative to conifers. When a similar effect (+, −, or 0) is observed more than once per reference, the number of cases showing this effect is indicated in brackets next to the effect sign.
| Reference | Location | Conifer Species 1 | Stand Age 2 (Year) | Sampling Depth (cm) | Soil Type | MAT 3 (°C) | MAP 4 (mm) | N | C Concentration | C Content | C Stability | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FF | MIN | FF | MIN | FF+MIN | FF | MIN | Indicators 5 | |||||||||
| Alban 1982 [68] | MN | Pg, Pr, Pb | 39–40, 41–49 | FF, 0–61, 0–80 | Alfisols | 4 | 610 | 1 | 0, -(5) | 0(2), -(4) | ||||||
| Alexander and Mack 2016 [102] | AK | Pm | 20–100 | FF, 0–10 | Gelisols to Inceptisols | n.a. | 286 | 2–14 | 0(3) | 0, -(2) | 0(3) | |||||
| Ayres et al. 2009 [54] | CO | Pc, Pe | n.a. | 0–10 | n.a. | n.a. | n.a. | 4 | 0(2) | 0(2) | ||||||
| Bauhus et al. 1998 [103] | QC | Ab+Pg | 50, 124 | FF, 0–10 | Spodosols, Alfisols | 0.6 | 823 | 1 | 0(4) | 0(4) | ||||||
| Boča and Van Miegroet 2017 [28] | UT | Ptm+Al+Ac+Pf | mature | FF, 0–50 | Alfisols, Mollisols | 3.2, 4.5 | 1031, 823 | 4 | - | + | 0 | +(3) | A, B, C | |||
| Buck and St. Clair 2012 [104] | UT | Al+Pe+Ptm | n.a. | FF(OA), 0–10 | n.a. | 10.1 | 201 | 10 | 0 | 0 | ||||||
| Côté et al. 2000 [31] | QC | Ab+Pg | 50, 124 | FF, 0–10 | Spodosols, Alfisols | 0.6 | 823 | 4 | 0 | 0 | 0 | 0 | 0 | D | ||
| Dobarco and Van Miegroet 2014 [38] | UT | Ptm+Al+Ac+Pf | mature | 0–15 | Alfisols, Mollisols | 3.2, 4.5 | 1197, 812 | 6, 5 | 0(2) | 0(2) | 0(4), +(2) | A, B, D | ||||
| Giardina et al. 2001 [29] | CO | Pc | 40–250 | 0–15 | n.a. | 0.5 | 700–850 | 6 | + | D | ||||||
| Hannam et al. 2004 [105] | AB | Pg | 80–140 | FF | Alfisols | −0.6 | 433 | 3 | - | 0 | ||||||
| Hannam et al. 2005 [106] | AB | Pg | 80–140 | FF | Alfisols | −0.6 | 433 | 3 | - | 0 | ||||||
| Jerabkova et al. 2006 [107] | AB | Pg mainly | 70–125 | FF, 0–7 | Alfisols | −0.6 | 433 | 3 | 0 | 0 | 0 | 0 | ||||
| Kishchuk et al. 2014 [108] | AB | Pg mainly | 62–124 | 10–17 | Alfisols | −0.6 | 431 | 3 | 0 | |||||||
| Laganière et al. 2009 [73] | QC | Pm | 79–89 | FF | Alfisols | 0.8 | 890 | 3 | - | |||||||
| Laganière et al. 2011 [39] | QC | Pm | 90 | 0–15 | Alfisols | 0.7 | 890 | 8 | + | A | ||||||
| Laganière et al. 2012 [33] | QC | Pm | 90 | n/a | Alfisols | 0.7 | 890 | 8 | + | E | ||||||
| Laganière et al. 2013 [30] | QC, ON | Pm, Pb | 90, 83 | FF, 0–55 | Alfisols, Inseptisols | 0.7, 2.5 | 890, 712 | 8, 4 | 0, - | 0(2), - | 0(2) | +(2) | +(2) | C, D | ||
| Lamarche et al. 2004 [109] | QC | Pg+Pm+Ab | 57–131 | FF | Alfisols, Spodosols | 0.8 | 857 | 18 | 0 | |||||||
| Olsen and Van Miegroet 2010 [32] | UT | Al+Pe | mature | 0–30 | Alfisols | 7 | 950 | 3 | 0 | 0 | D | |||||
| Paré and Bergeron 1996 [4] | QC | Pg | 49–123 | FF, 0–10 | Alfisols | 0.6 | 823 | 8 | - | 0 | ||||||
| Ste-Marie et al. 2007 [69] | QC | Pb | 59–89 | FF, 0–20 | Alfisols, Spodosols | 0.7 | 890 | 3 | 0, + | 0(2) | ||||||
| Weishampel et al. 2009 [110] | MN | Pb+Pr+Ab | 20–58 | FF, 0–40 | Alfisols | 3 | 785 | 3–15 | - | 0 | 0 | |||||
| Woldeselassie et al. 2012 [3] | UT | Ptm+Pe+Al+Pc | mature | FF, 0–60 | variable | 4.5 | 890–950 | 6 | - | 0, + | + | +(3) | A, C, D | |||
1 Ab, Abies balsamea; Ac, Abies concolor; Al, Abies lasiocarpa; Pb, Pinus banksiana; Pc, Pinus contorta; Pe, Picea engelmannii; Pf, Pinus flexilis; Pg, Picea glauca; Pm, Picea mariana, Pr, Pinus resinosa; Ptm, Pseudotsuga menziesii; 2 n.a., information not available; 3 MAT, mean annual temperature; 4 MAP, mean annual precipitation; 5 Stability indicators: A, degree of organo-mineral associations; B, water-extractable C; C, pattern of soil C distribution; D, proportion of readily-available or labile C estimated during long-term soil incubations; E, temperature sensitivity of soil heterotrophic respiration.
References
- Burns, R.; Honkala, B. Silvics of North America: 2. Hardwoods. Agriculture Handbook 654.; Department of Agriculture, Forest Service: Washington, DC, USA, 1990.
- Bartos, D.L.; Amacher, M.C. Soil Properties Associated with Aspen to Conifer Succession. Rangelands 1998, 20, 25–28. [Google Scholar]
- Woldeselassie, M.; Van Miegroet, H.; Gruselle, M.-C.; Hambly, N. Storage and Stability of Soil Organic Carbon in Aspen and Conifer Forest Soils of Northern Utah. Soil Sci. Soc. Am. J. 2012, 76, 2230. [Google Scholar] [CrossRef]
- Paré, D.; Bergeron, Y. Effect of colonizing tree species on soil nutrient availability in clay soil of the boreal mixedwood. Can. J. For. Res. 1996, 26, 1022–1031. [Google Scholar] [CrossRef]
- Reich, P.B.; Bakken, P.; Carlson, D.; Frelich, L.E.; Friedman, S.K.; Grigal, D.F. Influence of Logging, Fire, and Forest Type on Biodiversity and Productivity in Southern Boreal Forests. Ecology 2001, 82, 2731–2748. [Google Scholar] [CrossRef]
- Laganière, J.; Cavard, X.; Brassard, B.W.; Paré, D.; Bergeron, Y.; Chen, H.Y.H. The influence of boreal tree species mixtures on ecosystem carbon storage and fluxes. For. Ecol. Manag. 2015, 354, 119–129. [Google Scholar] [CrossRef]
- Kuhn, T.J.; Safford, H.D.; Jones, B.E.; Tate, K.W. Aspen (Populus tremuloides) stands and their contribution to plant diversity in a semiarid coniferous landscape. Plant Ecol. 2011, 212, 1451–1463. [Google Scholar] [CrossRef]
- DeByle, N.V.; Winokur, R.P. Aspen: Ecology and Management in the Western United States. General Technical Report RM-119. USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1985; Volume 119. [Google Scholar]
- Griffis-Kyle, K.L.; Beier, P. Small isolated aspen stands enrich bird communities in southwestern ponderosa pine forests. Biol. Conserv. 2003, 110, 375–385. [Google Scholar] [CrossRef]
- LaMalfa, E.M.; Ryle, R. Differential Snowpack Accumulation and Water Dynamics in Aspen and Conifer Communities: Implications for Water Yield and Ecosystem Function. Ecosystems 2008, 11, 569–581. [Google Scholar] [CrossRef]
- Long, J.N.; Mock, K. Changing perspectives on regeneration ecology and genetic diversity in western quaking aspen: Implications for silviculture. Can. J. For. Res. 2012, 42, 2011–2021. [Google Scholar] [CrossRef]
- Baker, F.S. Aspen in the Central Rocky Mountain Region. U. S. Dep. Agric. Bull. 1925, 1291. [Google Scholar] [CrossRef]
- Packard, F.M. Wildlife and Aspen in Rocky Mountain National Park, Colorado. Ecology 1942, 23, 478–482. [Google Scholar] [CrossRef]
- Bartos, D.L. Landscape dynamics of aspen and conifer forests. In Sustaining Aspen in Western Landscapes: Symposium Proceedings; Shepperd, W.D., Binkley, D., Bartos, D.L., Stohlgren, T.J., Eskew, L.G., Eds.; USDA Forest Service., Rocky Mountain Research Station: Fort Collins, CO, USA, 2001; pp. 5–14. [Google Scholar]
- Rogers, P. Using Forest Health Monitoring to assess aspen forest cover change in the southern Rockies ecoregion. For. Ecol. Manag. 2002, 155, 223–236. [Google Scholar] [CrossRef]
- Di Orio, A.P.; Callas, R.; Schaefer, R.J. Forty-eight year decline and fragmentation of aspen (Populus tremuloides) in the South Warner Mountains of California. For. Ecol. Manag. 2005, 206, 307–313. [Google Scholar] [CrossRef]
- Kulakowski, D.; Veblen, T.T.; Drinkwater, S. The Persistence of Quaking Aspen (Populus tremuloides) in the Grand Mesa Area, Colorado. Ecol. Appl. 2004, 14, 1603–1614. [Google Scholar] [CrossRef]
- Worrall, J.J.; Rehfeldt, G.E.; Hamann, A.; Hogg, E.H.; Marchetti, S.B.; Michaelian, M.; Gray, L.K. Recent declines of Populus tremuloides in North America linked to climate. For. Ecol. Manag. 2013, 299, 35–51. [Google Scholar] [CrossRef]
- Boulanger, Y.; Gauthier, S.; Gray, D.R.; Le Goff, H.; Lefort, P.; Morissette, J. Fire regime zonation under current and future climate over eastern Canada. Ecol. Appl. 2013, 23, 904–923. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.H.; Vasiliauskas, S.; Kayahara, G.J.; Ilisson, T. Wildfire promotes broadleaves and species mixture in boreal forest. For. Ecol. Manag. 2009, 257, 343–350. [Google Scholar] [CrossRef]
- Lafleur, B.; Cazal, A.; Leduc, A.; Bergeron, Y. Soil organic layer thickness influences the establishment and growth of trembling aspen (Populus tremuloides) in boreal forests. For. Ecol. Manag. 2015, 347, 209–216. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Goetz, S.J.; Mack, M.C.; Alexander, H.D.; Jin, Y.; Randerson, J.T.; Loranty, M.M. The impacts and implications of an intensifying fire regime on Alaskan boreal forest composition and albedo. Glob. Chang. Biol. 2011, 17, 2853–2866. [Google Scholar] [CrossRef]
- Kulakowski, D.; Matthews, C.; Jarvis, D.; Veblen, T.T. Compounded disturbances in sub-alpine forests in western Colorado favour future dominance by quaking aspen (Populus tremuloides). J. Veg. Sci. 2013, 24, 168–176. [Google Scholar] [CrossRef]
- Ilisson, T.; Chen, H.Y.H. Response of six boreal tree species to stand replacing fire and clearcutting. Ecosystems 2009, 12, 820–829. [Google Scholar] [CrossRef]
- Boucher, Y.; Auger, I.; Noël, J.; Grondin, P.; Arseneault, D. Fire is a stronger driver of forest composition than logging in the boreal forest of eastern Canada. J. Veg. Sci. 2017, 28, 57–68. [Google Scholar] [CrossRef]
- Augusto, L.; De Schrijver, A.; Vesterdal, L.; Smolander, A.; Prescott, C.; Ranger, J. Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests. Biol. Rev. Camb. Philos. Soc. 2015, 9, 444–466. [Google Scholar] [CrossRef] [PubMed]
- Vesterdal, L.; Clarke, N.; Sigurdsson, B.D.; Gundersen, P. Do tree species influence soil carbon stocks in temperate and boreal forests? For. Ecol. Manag. 2013, 309, 1–15. [Google Scholar] [CrossRef]
- Boča, A.; Van Miegroet, H. Can carbon fluxes explain differences in soil organic carbon storage under aspen and conifer forest overstories? Forests 2017. accepted. [Google Scholar]
- Giardina, C.P.; Ryan, M.G.; Hubbard, R.M.; Binkley, D. Tree Species and Soil Textural Controls on Carbon and Nitrogen Mineralization Rates. Soil Sci. Soc. Am. J. 2001, 65, 1272–1279. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bergeron, Y.; Chen, H.Y.H.; Brassard, B.W.; Cavard, X. Stability of Soil Carbon Stocks Varies with Forest Composition in the Canadian Boreal Biome. Ecosystems 2013, 16, 852–865. [Google Scholar] [CrossRef]
- Côté, L.; Brown, S.; Paré, D.; Fyles, J.; Bauhus, J. Dynamics of carbon and nitrogen mineralization in relation to stand type, stand age and soil texture in the boreal mixedwood. Soil Biol. Biochem. 2000, 32, 1079–1090. [Google Scholar] [CrossRef]
- Olsen, H.R.; Van Miegroet, H. Factors Affecting Carbon Dioxide Release from Forest and Rangeland Soils in Northern Utah. Soil Sci. Soc. Am. J. 2010, 74, 282. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bergeron, Y.; Chen, H.Y.H. The effect of boreal forest composition on soil respiration is mediated through variations in soil temperature and C quality. Soil Biol. Biochem. 2012, 53, 18–27. [Google Scholar] [CrossRef]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Mikutta, R.; Kleber, M.; Torn, M.S.; Jahn, R. Stabilization of Soil Organic Matter: Association with Minerals or Chemical Recalcitrance? Biogeochemistry 2006, 77, 25–56. [Google Scholar] [CrossRef]
- Von Lützow, M.V.; Kogel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Dungait, J.A.J.; Hopkins, D.W.; Gregory, A.S.; Whitmore, A.P. Soil organic matter turnover is governed by accessibility not recalcitrance. Glob. Chang. Biol. 2012, 18, 1781–1796. [Google Scholar] [CrossRef]
- Román Dobarco, M.; Van Miegroet, H. Soil Organic Carbon Storage and Stability in the Aspen-Conifer Ecotone in Montane Forests in Utah State, USA. Forests 2014, 5, 666–688. [Google Scholar] [CrossRef]
- Laganière, J.; Angers, D.A.; Paré, D.; Bergeron, Y.; Chen, H.Y.H. Black Spruce Soils Accumulate More Uncomplexed Organic Matter than Aspen Soils. Soil Sci. Soc. Am. J. 2011, 75, 1125. [Google Scholar] [CrossRef]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Fire effects on temperate forest soil C and N storage. Ecol. Appl. 2011, 21, 1189–1201. [Google Scholar] [CrossRef] [PubMed]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Kishchuk, B.E.; Morris, D.M.; Lorente, M.; Keddy, T.; Sidders, D.; Quideau, S.; Thiffault, E.; Kwiaton, M.; Maynard, D. Disturbance intensity and dominant cover type influence rate of boreal soil carbon change: A Canadian multi-regional analysis. For. Ecol. Manag. 2016, 381, 48–62. [Google Scholar] [CrossRef]
- Harrison, R.B.; Footen, P.W.; Strahm, B.D. Deep Soil Horizons: Contribution and Importance to Soil Carbon Pools and in Assessing Whole-Ecosystem Response to Management and Global Change. For. Sci. 2011, 57, 67–76. [Google Scholar]
- Boča, A.; Van Miegroet, H.; Gruselle, M.-C. Forest Overstory Effect on Soil Organic Carbon Storage: A Meta-analysis. Soil Sci. Soc. Am. J. 2014, 78, S35–S47. [Google Scholar] [CrossRef]
- Gower, S.T.; Hunter, A.; Campbell, J.; Vogel, J.; Veldhuis, H.; Harden, J.; Trumbore, S.; Norman, J.M.; Kucharik, C.J. Nutrient dynamics of the southern and northern BOREAS boreal forests. Ecoscience 2000, 7, 481–490. [Google Scholar] [CrossRef]
- Castellano, M.J.; Mueller, K.E.; Olk, D.C.; Sawyer, J.E.; Six, J. Integrating plant litter quality, soil organic matter stabilization, and the carbon saturation concept. Glob. Chang. Biol. 2015, 21, 3200–3209. [Google Scholar] [CrossRef] [PubMed]
- Cyle, K.T.; Hill, N.; Young, K.; Jenkins, T.; Hancock, D.; Schroeder, P.A.; Thompson, A. Substrate quality influences organic matter accumulation in the soil silt and clay fraction. Soil Biol. Biochem. 2016, 103, 138–148. [Google Scholar] [CrossRef]
- Lajtha, K.; Bowden, R.D.; Nadelhoffer, K. Litter and Root Manipulations Provide Insights into Soil Organic Matter Dynamics and Stability. Soil Sci. Soc. Am. J. 2014, 78, S261. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Bergeron, Y. Influence of Environmental Variability on Root Dynamics in Northern Forests. CRC Crit. Rev. Plant Sci. 2009, 28, 179–197. [Google Scholar] [CrossRef]
- Hajek, P.; Hertel, D.; Leuschner, C. Intraspecific variation in root and leaf traits and leaf-root trait linkages in eight aspen demes (Populus tremula and P. tremuloides). Front. Plant Sci. 2013, 4, 415. [Google Scholar] [CrossRef] [PubMed]
- Steele, S.J.; Gower, S.T.; Vogel, J.G.; Norman, J.M. Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskatchewan and Manitoba, Canada. Tree Physiol. 1997, 17, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Brassard, B.W.; Chen, H.Y.H.; Bergeron, Y.; Paré, D. Differences in fine root productivity between mixed- and single-species stands. Funct. Ecol. 2011, 25, 238–246. [Google Scholar] [CrossRef]
- Bauhus, J.; Messier, C. Soil exploitation strategies of fine roots in different tree species of the southern boreal forest of eastern Canada. Can. J. For. Res. 1999, 29, 260–273. [Google Scholar] [CrossRef]
- Ayres, E.; Steltzer, H.; Berg, S.; Wallenstein, M.D.; Simmons, B.L.; Wall, D.H. Tree species traits influence soil physical, chemical, and biological properties in high elevation forests. PLoS ONE 2009, 4, e5964. [Google Scholar] [CrossRef] [PubMed]
- Finér, L.; Messier, C.; De Grandpré, L. Fine-root dynamics in mixed boreal conifer - broad-leafed forest stands at different successional stages after fire. Can. J. For. Res. 1997, 27, 304–314. [Google Scholar] [CrossRef]
- Block, R.M.A.; Van Rees, K.C.J.; Knight, J.D. A Review of Fine Root Dynamics in Populus Plantations. Agrofor. Syst. 2006, 67, 73–84. [Google Scholar] [CrossRef]
- Moore, T.R.; Trofymow, J.A.; Prescott, C.E.; Fyles, J.; Titus, B.D. Patterns of carbon, nitrogen and phosphorus dynamics in decomposing foliar litter in Canadian forests. Ecosystems 2006, 9, 46–62. [Google Scholar] [CrossRef]
- Strukelj, M.; Brais, S.; Quideau, S.A.; Oh, S.-W. Chemical transformations of deadwood and foliar litter of mixed boreal species during decomposition. Can. J. For. Res. 2012, 42, 772–788. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bradley, R.L. How does a tree species influence litter decomposition? Separating the relative contribution of litter quality, litter mixing, and forest floor conditions. Can. J. For. Res. 2010, 40, 465–475. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, Y. Carbon:nitrogen stoichiometry in forest ecosystems during stand development. Glob. Ecol. Biogeogr. 2011, 20, 354–361. [Google Scholar] [CrossRef]
- Preston, C.M.; Bhatti, J.S.; Flanagan, L.B.; Norris, C. Stocks, chemistry, and sensitivity to climate change of dead organic matter along the Canadian boreal forest transect case study. Clim. Chang. 2006, 74, 233–251. [Google Scholar] [CrossRef]
- Prescott, C.E.; Zabek, L.M.; Staley, C.L.; Kabzems, R. Decomposition of broadleaf and needle litter in forests of British Columbia: Influences of litter type, forest type, and litter mixtures. Can. J. For. Res. 2000, 30, 1742–1750. [Google Scholar] [CrossRef]
- Gower, S.T.; Vogel, J.G.; Norman, M.; Kucharik, C.J.; Steele, S.J. Carbon distribution and aboveground net primary production in aspen, jack pine, and black spruce stands in Saskatchewan and Manitoba, Canada. J. Geophys. Res. Biogeosci. 1997, 102, 29029–29041. [Google Scholar] [CrossRef]
- Smith, J.L.; Bell, J.M.; Bolton, H.; Bailey, V.L. The initial rate of C substrate utilization and longer-term soil C storage. Biol. Fertil. Soils 2007, 44, 315–320. [Google Scholar] [CrossRef]
- Yang, H.S.; Janssen, B.H. Relationship between substrate initial reactivity and residues ageing speed in carbon mineralization. Plant Soil 2002, 239, 215–224. [Google Scholar] [CrossRef]
- Légaré, S.; Paré, D.; Bergeron, Y. Influence of aspen on forest floor properties in black spruce-dominated stands. Plant Soil 2005, 275, 207–220. [Google Scholar] [CrossRef]
- Ste-Marie, C.; Houle, D. Forest floor gross and net nitrogen mineralization in three forest types in Quebec, Canada. Soil Biol. Biochem. 2006, 38, 2135–2143. [Google Scholar] [CrossRef]
- Alban, D.H. Effects of Nutrient Accumulation by Aspen, Spruce, and Pine on Soil Properties. Soil Sci. Soc. Am. J. 1982, 46, 853–861. [Google Scholar] [CrossRef]
- Ste-Marie, C.; Paré, D.; Gagnon, D. The contrasting effects of aspen and jack pine on soil nutritional properties depend on parent material. Ecosystems 2007, 10, 1299–1310. [Google Scholar] [CrossRef]
- Wuddivira, M.N.; Camps-Roach, G. Effects of organic matter and calcium on soil structural stability. Eur. J. Soil Sci. 2007, 58, 722–727. [Google Scholar] [CrossRef]
- Baldock, J.; Skjemstad, J. Role of the soil matrix and minerals in protecting natural organic materials against biological attack. Org. Geochem. 2000, 31, 697–710. [Google Scholar] [CrossRef]
- Whittinghill, K.A.; Hobbie, S.E. Effects of pH and calcium on soil organic matter dynamics in Alaskan tundra. Biogeochemistry 2012, 111, 569–581. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bradley, R.L. Linking the abundance of aspen with soil faunal communities and rates of belowground processes within single stands of mixed aspen–black spruce. Appl. Soil Ecol. 2009, 41, 19–28. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Lal, R. Mechanisms of Carbon Sequestration in Soil Aggregates. CRC Crit. Rev. Plant Sci. 2004, 23, 481–504. [Google Scholar] [CrossRef]
- Bossuyt, H.; Six, J.; Hendrix, P.F. Protection of soil carbon by microaggregates within earthworm casts. Soil Biol. Biochem. 2005, 37, 251–258. [Google Scholar] [CrossRef]
- Lubbers, I.M.; van Groenigen, K.J.; Fonte, S.J.; Six, J.; Brussaard, L.; van Groenigen, J.W. Greenhouse-gas emissions from soils increased by earthworms. Nat. Clim. Chang. 2013, 3, 187–194. [Google Scholar] [CrossRef]
- Munroe, J.S.; Attwood, E.C.; O’Keefe, S.S.; Quackenbush, P.J.M. Eolian deposition in the alpine zone of the Uinta Mountains, Utah, USA. Catena 2015, 124, 119–129. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Stam, B.R.; Malechek, J.C.; Bartos, D.L.; Bowns, J.E.; Godfrey, E.B. Effect of Conifer Encroachment Into Aspen Stands on Understory Biomass. Rangel. Ecol. Manag. 2008, 61, 93–97. [Google Scholar] [CrossRef]
- Bisbee, K.E.; Gower, S.T.; Norman, J.M.; Nordheim, E.V. Environmental controls on ground cover species composition and productivity in a boreal black spruce forest. Oecologia 2001, 129, 261–270. [Google Scholar] [CrossRef]
- O’Connell, K.E.B.; Gower, S.T.; Norman, J.M. Net ecosystem production of two contrasting boreal black spruce forest communities. Ecosystems 2003, 6, 248–260. [Google Scholar] [CrossRef]
- Bona, K.A.; Shaw, C.H.; Fyles, J.W.; Kurz, W.A. Modelling moss-derived carbon in upland black spruce forests. Can. J. For. Res. 2016, 534, 520–534. [Google Scholar] [CrossRef]
- Gornall, J.L.; Jónsdóttir, I.S.; Woodin, S.J.; Van Der Wal, R. Arctic mosses govern below-ground environment and ecosystem processes. Oecologia 2007, 153, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.I.; Cornelissen, J.H.C.; Klahn, T.; Van Logtestijn, R.S.P.; Broekman, R.; Schweikert, W.; Aerts, R. An experimental comparison of chemical traits and litter decomposition rates in a diverse range of subarctic bryophyte, lichen and vascular plant species. J. Ecol. 2009, 97, 886–900. [Google Scholar] [CrossRef]
- Royer-Tardif, S.; Bradley, R.L. Forest floor properties across sharp compositional boundaries separating trembling aspen and jack pine stands in the southern boreal forest. Plant Soil 2011, 345, 353–364. [Google Scholar] [CrossRef]
- Harden, J.W.; O’Neill, K.P.; Trumbore, S.E.; Veldhuis, H.; Stocks, B.J. Moss and soil contributions to the annual net carbon flux of a maturing boreal forest. J. Geophys. Res. 1997, 102, 28805–28816. [Google Scholar] [CrossRef]
- Wickland, K.P.; Neff, J.C. Decomposition of soil organic matter from boreal black spruce forest: Environmental and chemical controls. Biogeochemistry 2008, 87, 29–47. [Google Scholar] [CrossRef]
- Fenton, N.J.; Bergeron, Y.; Paré, D. Decomposition rates of bryophytes in managed boreal forests: Influence of bryophyte species and forest harvesting. Plant Soil 2010, 336, 499–508. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Strong, W.L.; La Roi, G.H. Root-system morphology of common boreal forest trees in Alberta, Canada. Can. J. For. Res. 1983, 13, 1164–1173. [Google Scholar] [CrossRef]
- Pinno, B.D.; Wilson, S.D.; Steinaker, D.F.; Rees, K.C.J.; McDonald, S.A. Fine root dynamics of trembling aspen in boreal forest and aspen parkland in central Canada. Ann. For. Sci. 2010, 67, 710. [Google Scholar] [CrossRef]
- Yanai, R.D.; McFarlane, K.J.; Lucash, M.S.; Kulpa, S.E.; Wood, D.M. Similarity of nutrient uptake and root dimensions of Engelmann spruce and subalpine fir at two contrasting sites in Colorado. For. Ecol. Manag. 2009, 258, 2233–2241. [Google Scholar] [CrossRef]
- Eissenstat, D.M. Costs and benefits of constructing roots of small diameter. J. Plant Nutr. 1992, 15, 763–782. [Google Scholar] [CrossRef]
- Langley, A.J.; Chapman, S.K.; Hungate, B.A. Ectomycorrhizal colonization slows root decomposition: The post-mortem fungal legacy. Ecol. Lett. 2006, 9, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Neville, J.; Tessier, J.; Morrison, I.; Scarratt, J.; Canning, B.; Klironomos, J. Soil depth distribution of ecto- and arbuscular mycorrhizal fungi associated with Populus tremuloides within a 3-year-old boreal forest clear-cut. Appl. Soil Ecol. 2002, 19, 209–216. [Google Scholar] [CrossRef]
- Brzostek, E.R.; Greco, A.; Drake, J.E.; Finzi, A.C. Root carbon inputs to the rhizosphere stimulate extracellular enzyme activity and increase nitrogen availability in temperate forest soils. Biogeochemistry 2013, 115, 65–76. [Google Scholar] [CrossRef]
- Jenny, H. Factors of Soil Formation. A System of Quantitative Pedology; Dover Publications: New York, NY, USA, 1994. [Google Scholar]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Strahm, B.D.; Harrison, R.B. Controls on the Sorption, Desorption and Mineralization of Low-Molecular-Weight Organic Acids in Variable-Charge Soils. Soil Sci. Soc. Am. J. 2008, 72, 1653. [Google Scholar] [CrossRef]
- Woldeselassie, M.K. Soil Organic Carbon and Site Characteristics in Aspen and Evaluation of the Potential Effects of Conifer Encroachment on Soil Properties in Northern Utah, Master’s Thesis, Utah State University, Logan, UT, USA, 2009. [Google Scholar]
- Alexander, H.D.; Mack, M.C. A Canopy Shift in Interior Alaskan Boreal Forests: Consequences for Above- and Belowground Carbon and Nitrogen Pools during Post-fire Succession. Ecosystems 2016, 19, 98–114. [Google Scholar] [CrossRef]
- Bauhus, J.; Paré, D.; Côté, L. Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Buck, J.R.; St. Clair, S.B. Aspen Increase Soil Moisture, Nutrients, Organic Matter and Respiration in Rocky Mountain Forest Communities. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Hannam, K.D.; Quideau, S.A.; Oh, S.-W.; Kishchuk, B.E.; Wasylishen, R.E. Forest Floor Composition in Aspen- and Spruce-Dominated Stands of the Boreal Mixedwood Forest. Soil Sci. Soc. Am. J. 2004, 68, 1735. [Google Scholar] [CrossRef]
- Hannam, K.D.; Quideau, S.A; Kishchuk, B.E.; Oh, S.-W.; Wasylishen, R.E. Forest-floor chemical properties are altered by clear-cutting in boreal mixedwood forest stands dominated by trembling aspen and white spruce. Can. J. For. Res. 2005, 35, 2457–2468. [Google Scholar] [CrossRef]
- Jerabkova, L.; Prescott, C.E.; Kishchuk, B.E. Effect of variable-retention harvesting on soil nitrogen availability in boreal mixedwood forests. Can. J. For. Res. 2006, 36, 3029–3038. [Google Scholar] [CrossRef]
- Kishchuk, B.E.; Quideau, S.; Wang, Y.; Prescott, C. Long-term soil response to variable-retention harvesting in the EMEND (Ecosystem Management Emulating Natural Disturbance) experiment, northwestern Alberta. Can. J. Soil Sci. 2014, 94, 263–279. [Google Scholar] [CrossRef]
- Lamarche, J.; Bradley, R.L.; Pare, D.; Legare, S.; Bergeron, Y. Soil parent material may control forest floor properties more than stand type or stand age in mixedwood boreal forests. Ecoscience 2004, 11, 228–237. [Google Scholar] [CrossRef]
- Weishampel, P.; Kolka, R.; King, J.Y. Carbon pools and productivity in a 1-km2 heterogeneous forest and peatland mosaic in Minnesota, USA. For. Ecol. Manag. 2009, 257, 747–754. [Google Scholar] [CrossRef]
Figure 1.
Geographic distribution of trembling aspen (Populus tremuloides Michx.) and location of the study sites covered by this literature review.
Figure 1.
Geographic distribution of trembling aspen (Populus tremuloides Michx.) and location of the study sites covered by this literature review.
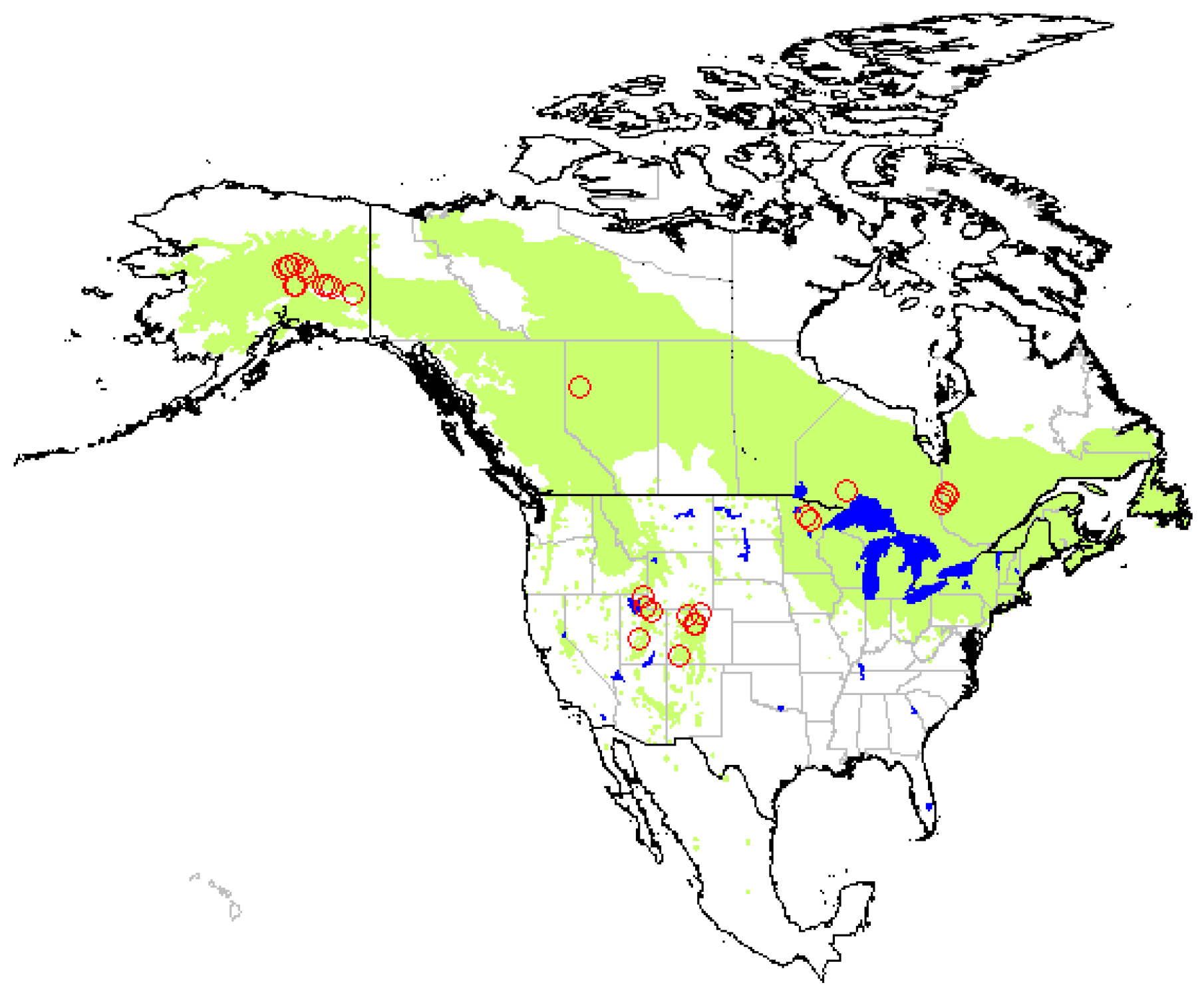
Table 1.
Number of cases where the presence of aspen caused an increase, a decrease, or no change in forest floor (FF) and mineral (MIN) soil C concentration, content, and stability relative to conifers.
Table 1.
Number of cases where the presence of aspen caused an increase, a decrease, or no change in forest floor (FF) and mineral (MIN) soil C concentration, content, and stability relative to conifers.
| Effect | C Concentration | C Content | C Stability | ||||
|---|---|---|---|---|---|---|---|
| FF | MIN | FF | MIN | FF+MIN | FF | MIN | |
| Increase | 0 | 0 | 1 | 2 | 1 | 2 | 13 |
| Decrease | 1 | 0 | 14 | 5 | 0 | 0 | 0 |
| No effect | 10 | 12 | 6 | 18 | 5 | 3 | 6 |
| Total number of cases | 11 | 12 | 21 | 25 | 6 | 5 | 19 |
Bold highlights the main effect in each column. Data extracted from 23 papers.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Laganière, J.; Boča, A.; Van Miegroet, H.; Paré, D. A Tree Species Effect on Soil That Is Consistent Across the Species’ Range: The Case of Aspen and Soil Carbon in North America. Forests 2017, 8, 113. https://doi.org/10.3390/f8040113
AMA Style
Laganière J, Boča A, Van Miegroet H, Paré D. A Tree Species Effect on Soil That Is Consistent Across the Species’ Range: The Case of Aspen and Soil Carbon in North America. Forests. 2017; 8(4):113. https://doi.org/10.3390/f8040113
Chicago/Turabian StyleLaganière, Jérôme, Antra Boča, Helga Van Miegroet, and David Paré. 2017. "A Tree Species Effect on Soil That Is Consistent Across the Species’ Range: The Case of Aspen and Soil Carbon in North America" Forests 8, no. 4: 113. https://doi.org/10.3390/f8040113
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.