
Tree Climbing Techniques and Volume Equations for Eschweilera (Matá-Matá), a Hyperdominant Genus in the Amazon Forest

Abstract
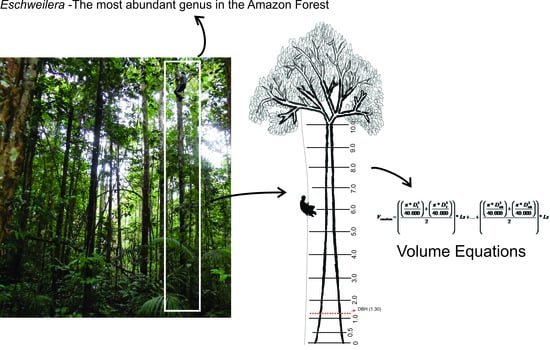
:
1. Introduction
2. Materials and Methods
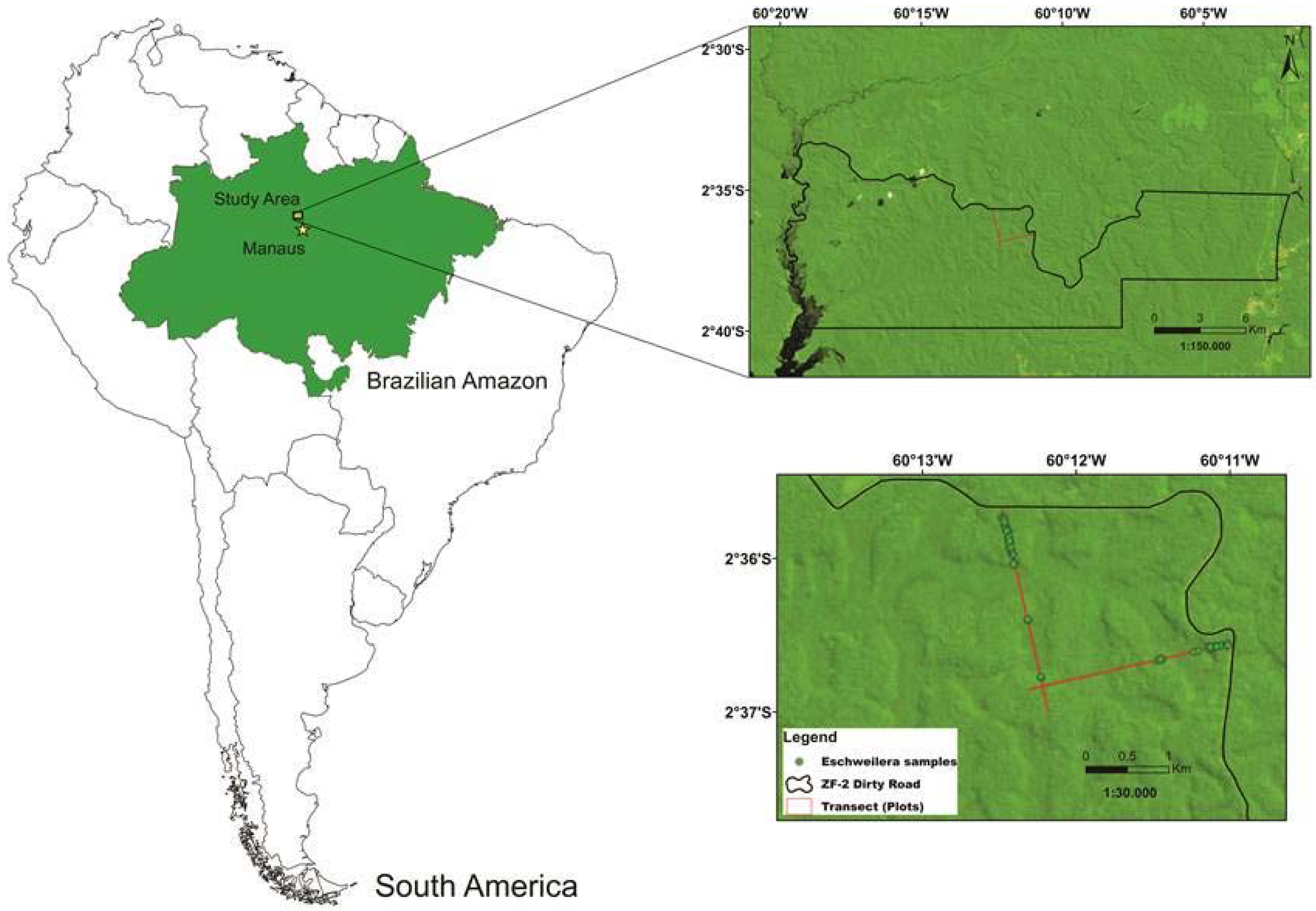
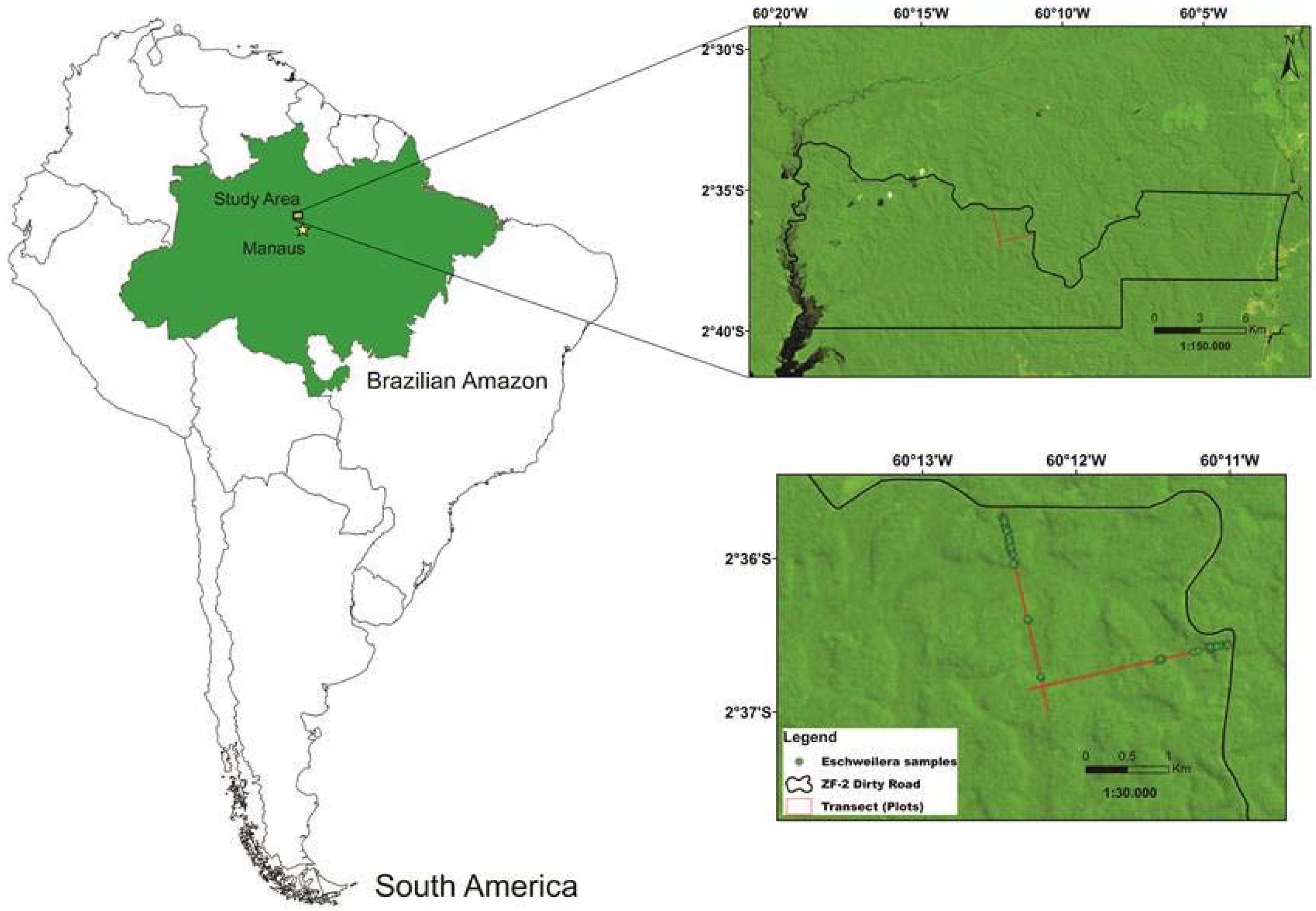
2.1. Study Area
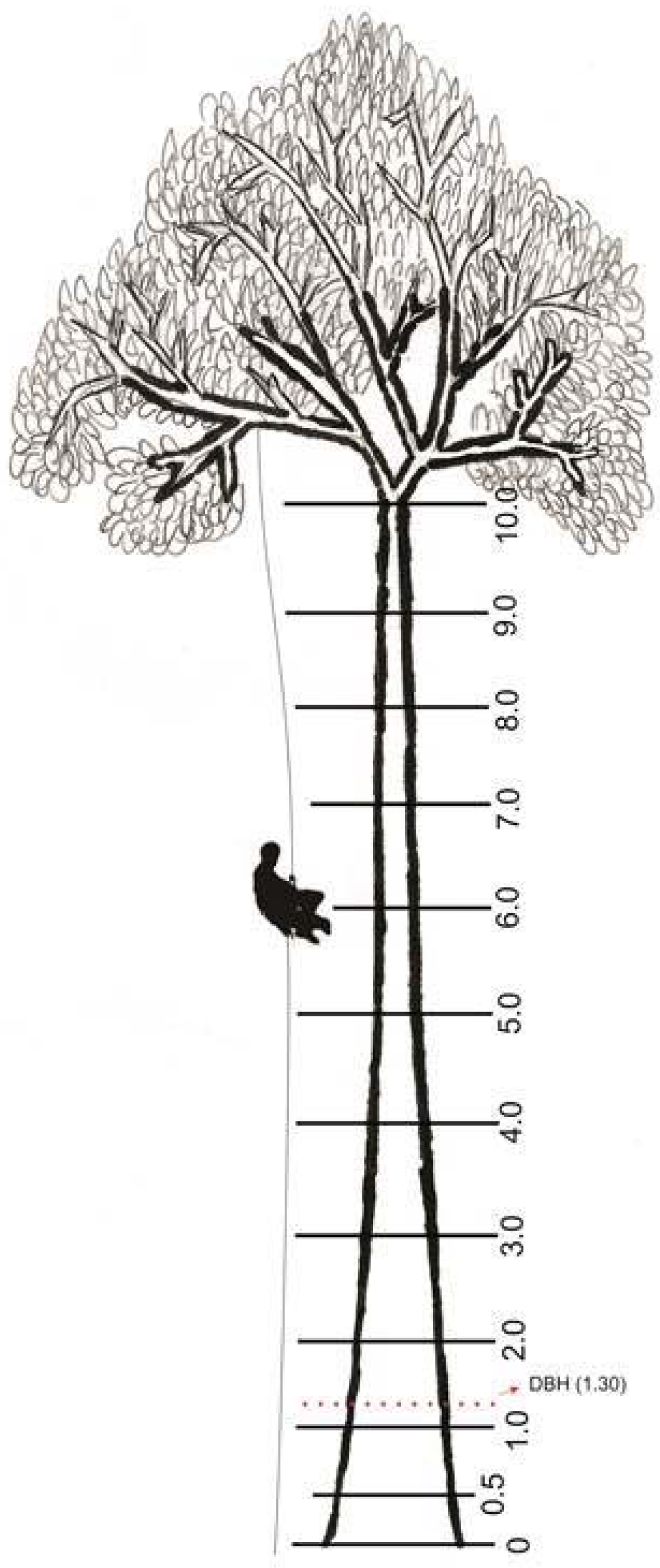
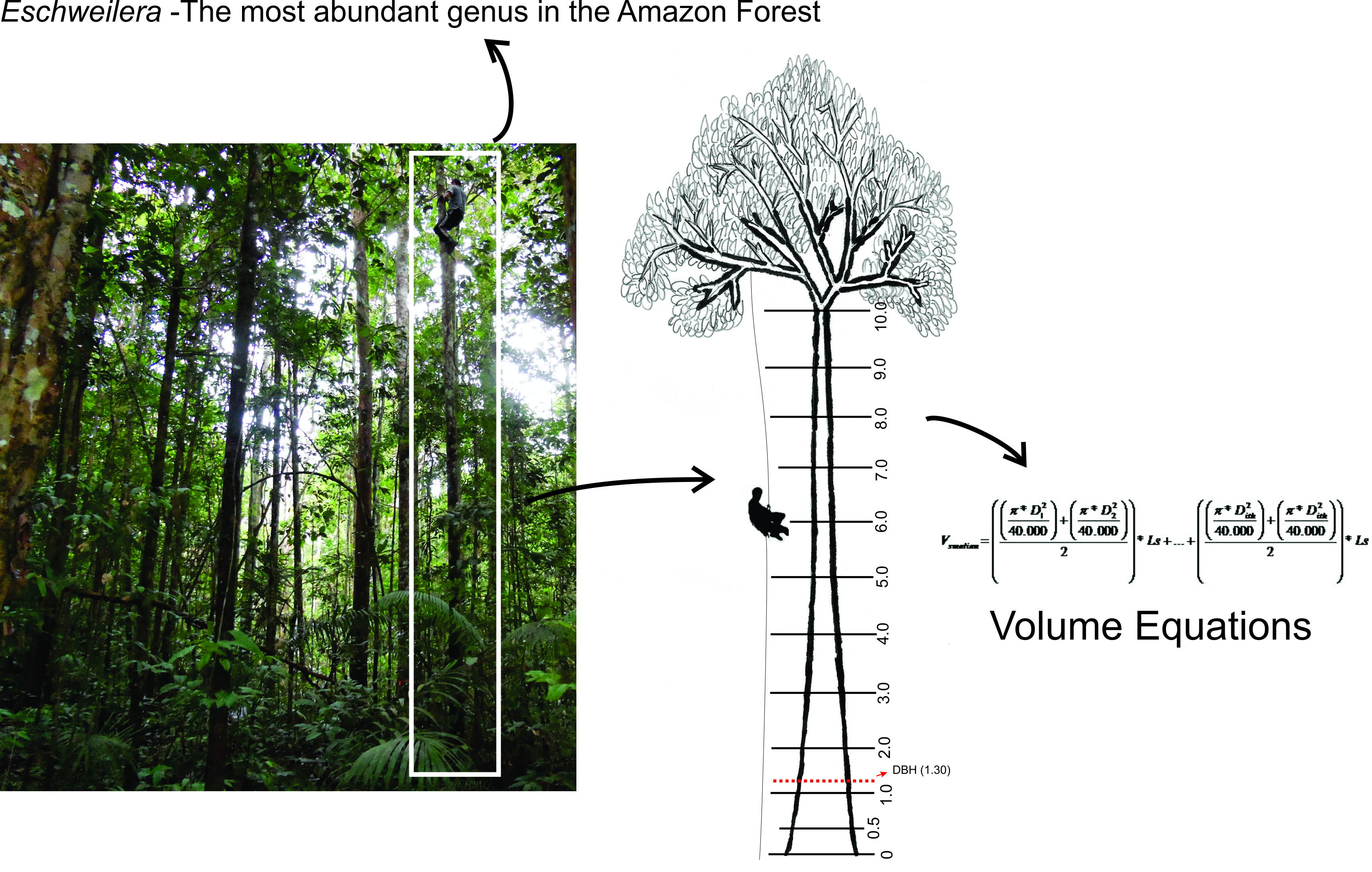
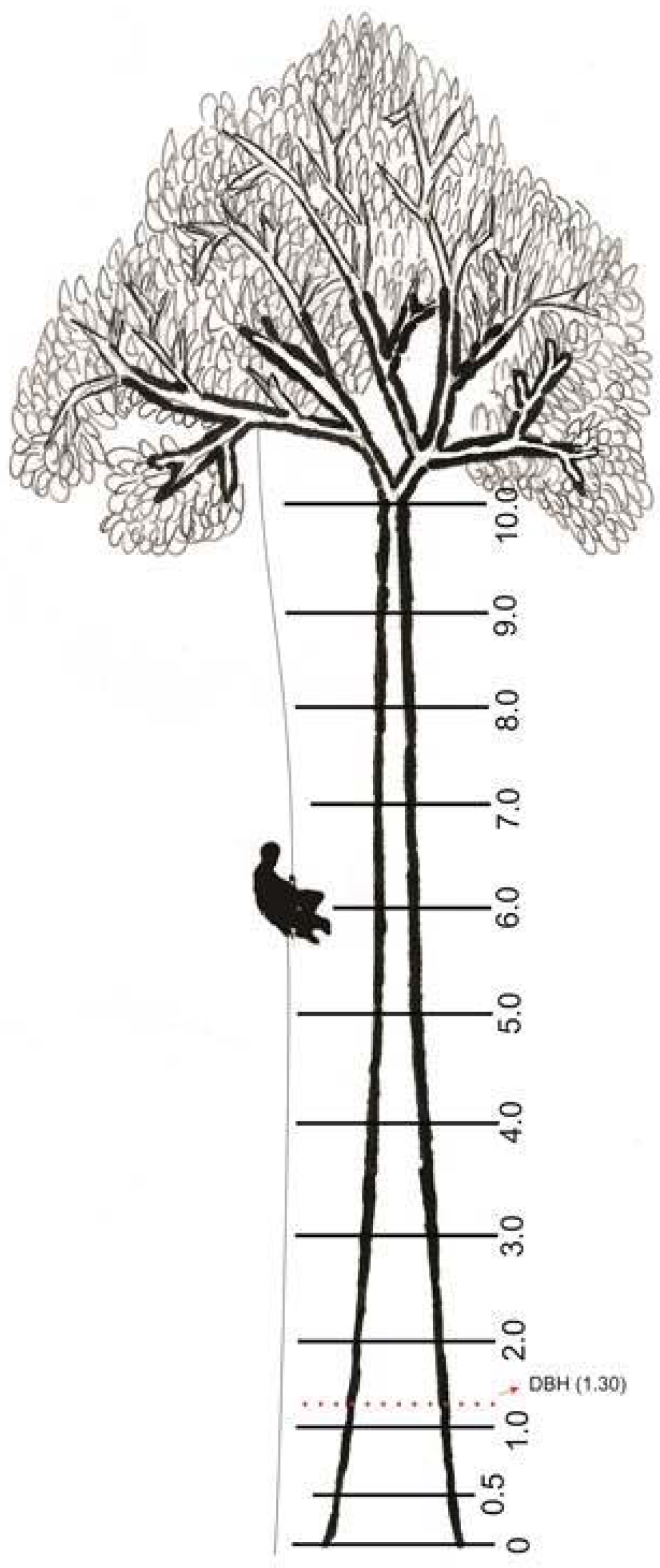
2.2. Climbing Technique
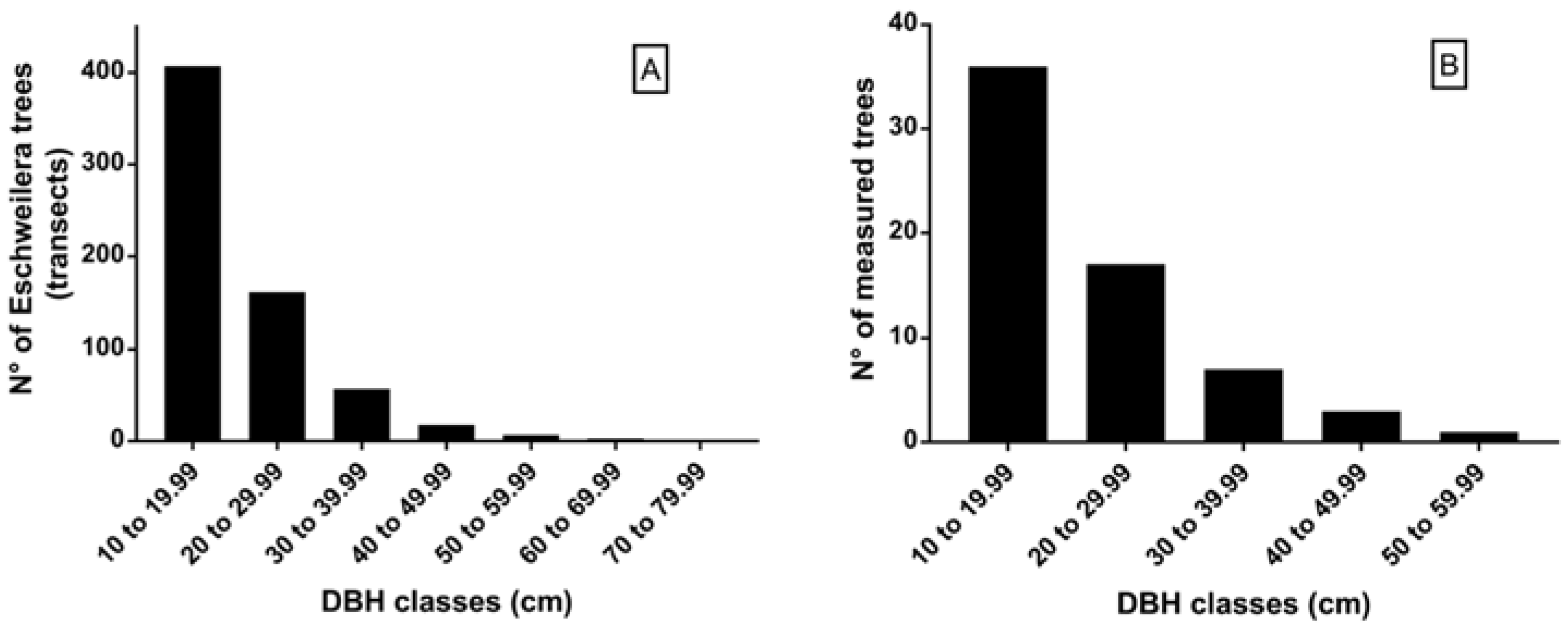
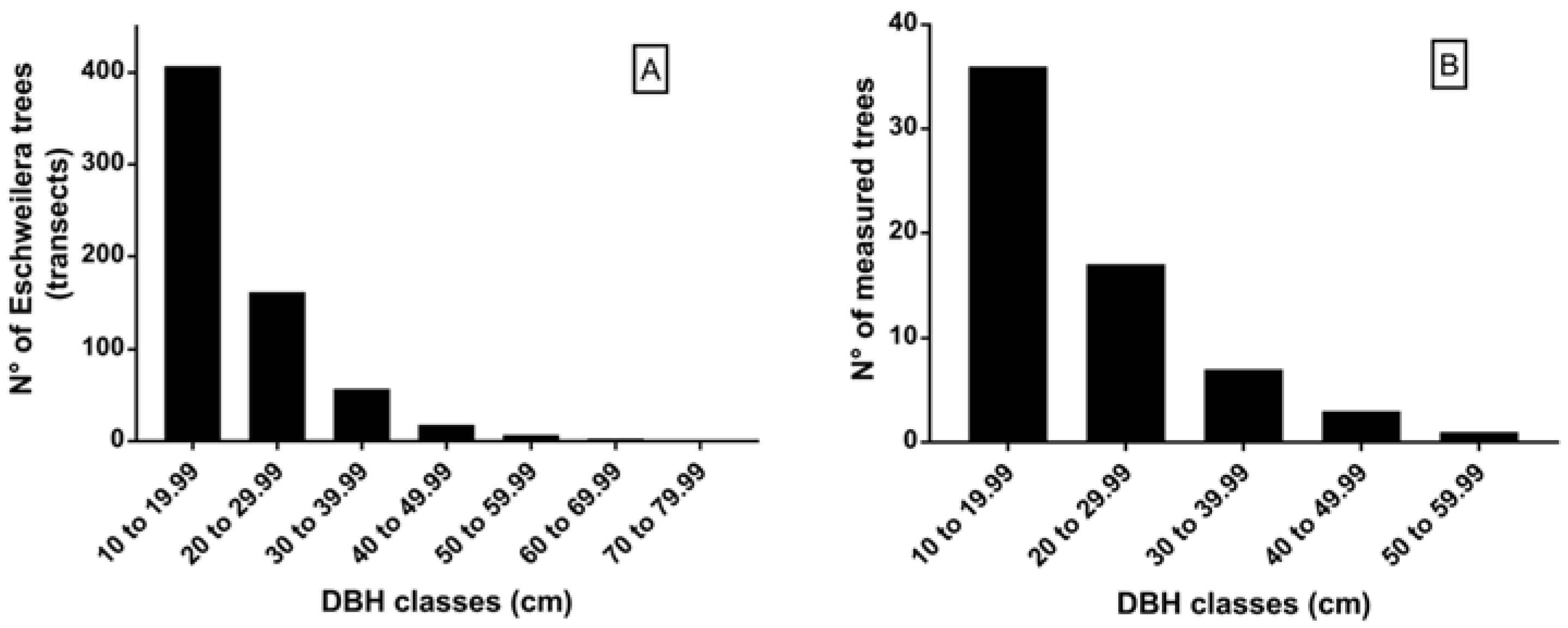
2.3. Selection Method of Eschweilera Trees
2.4. Tested Models
2.5. Form Factor
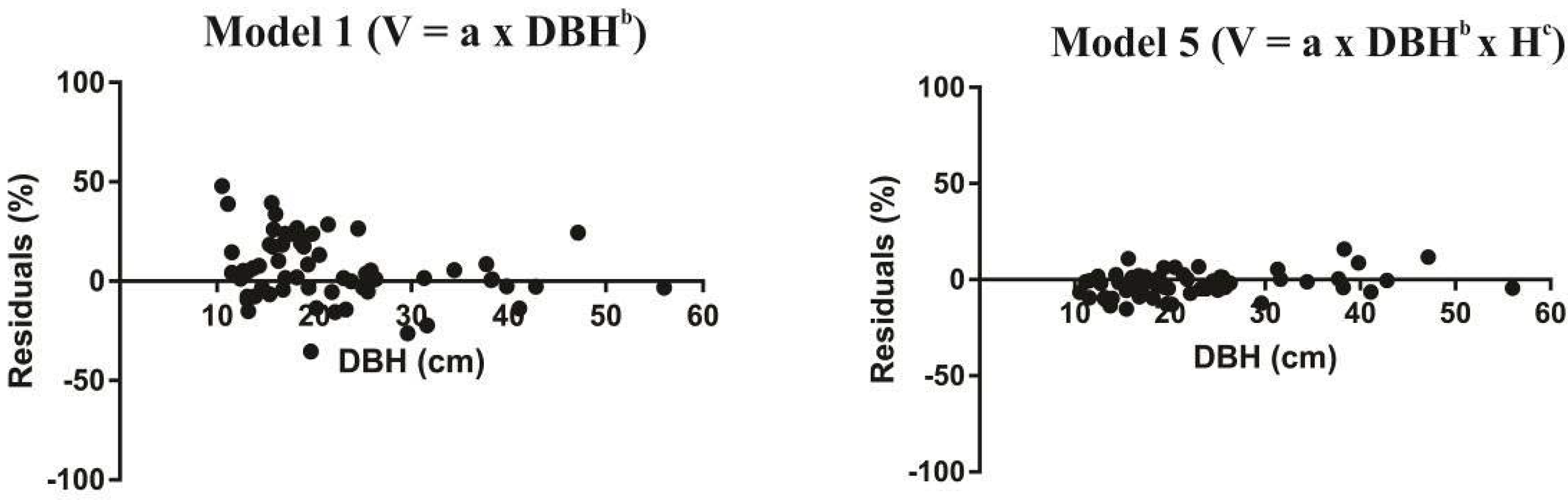
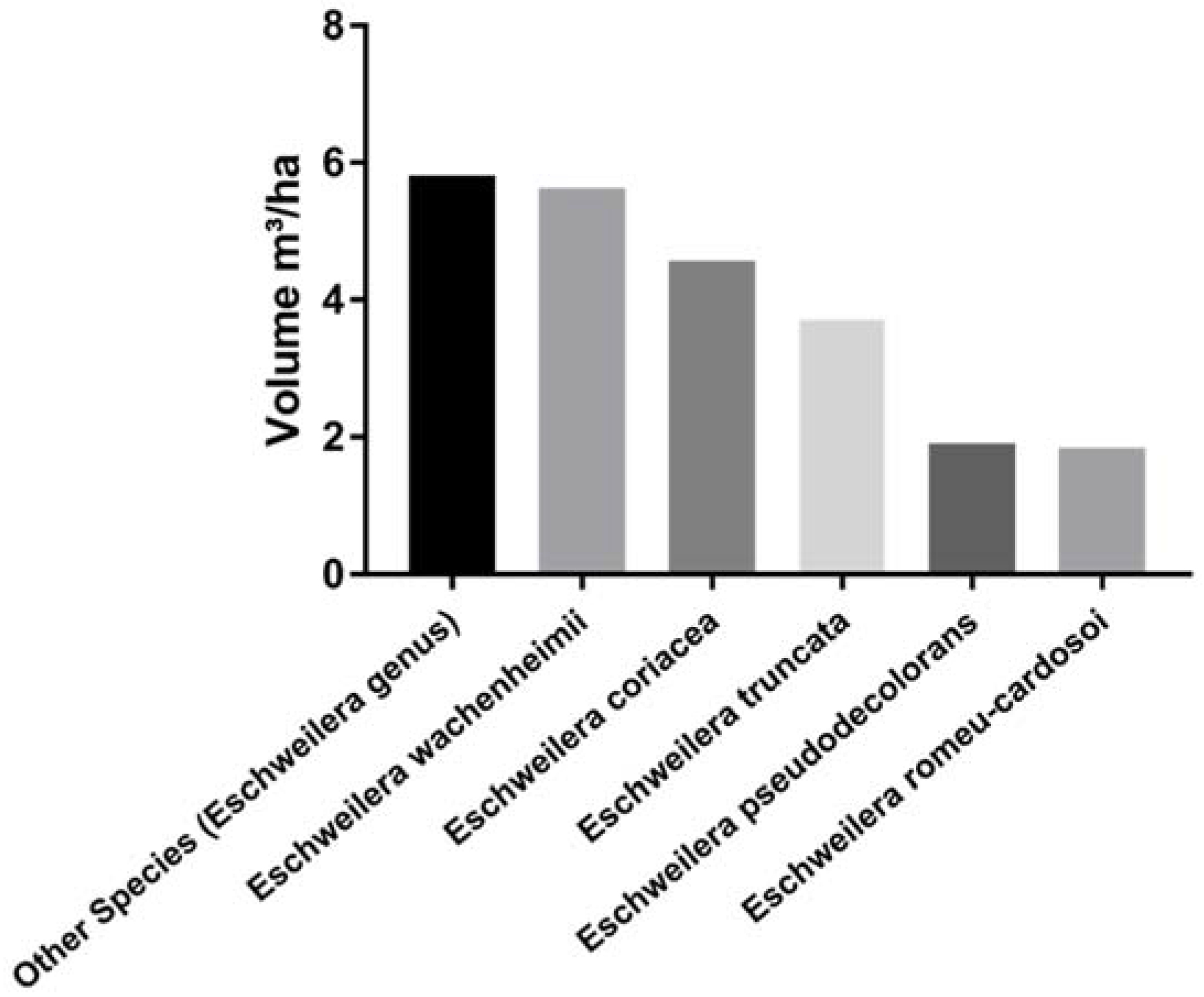
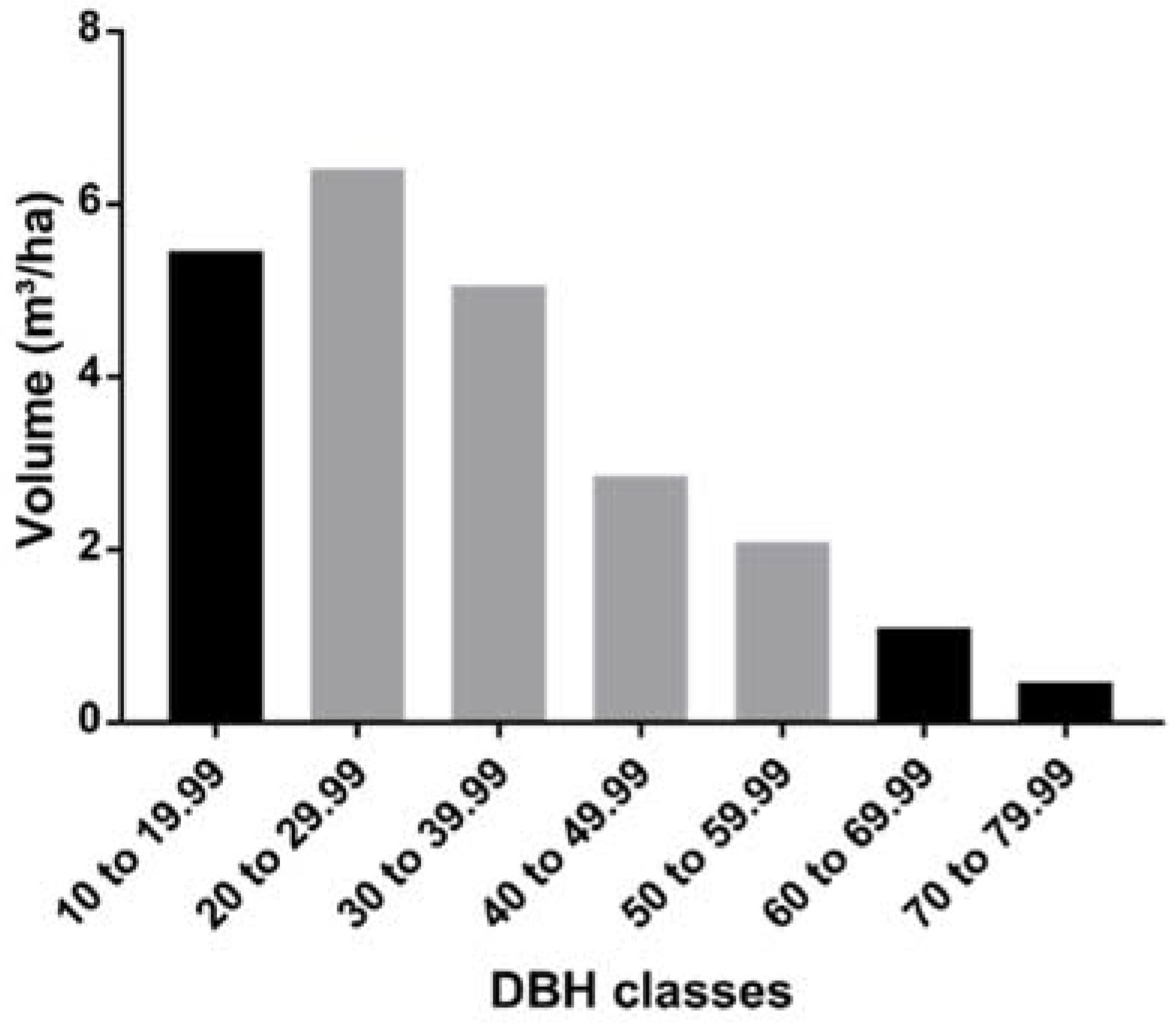
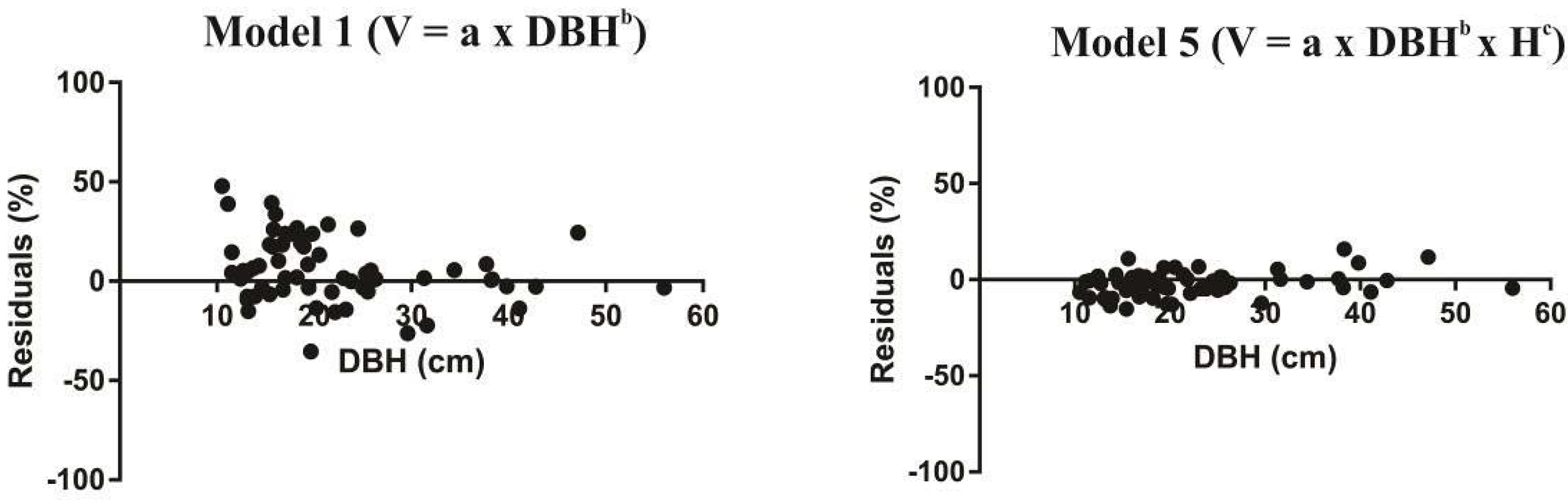
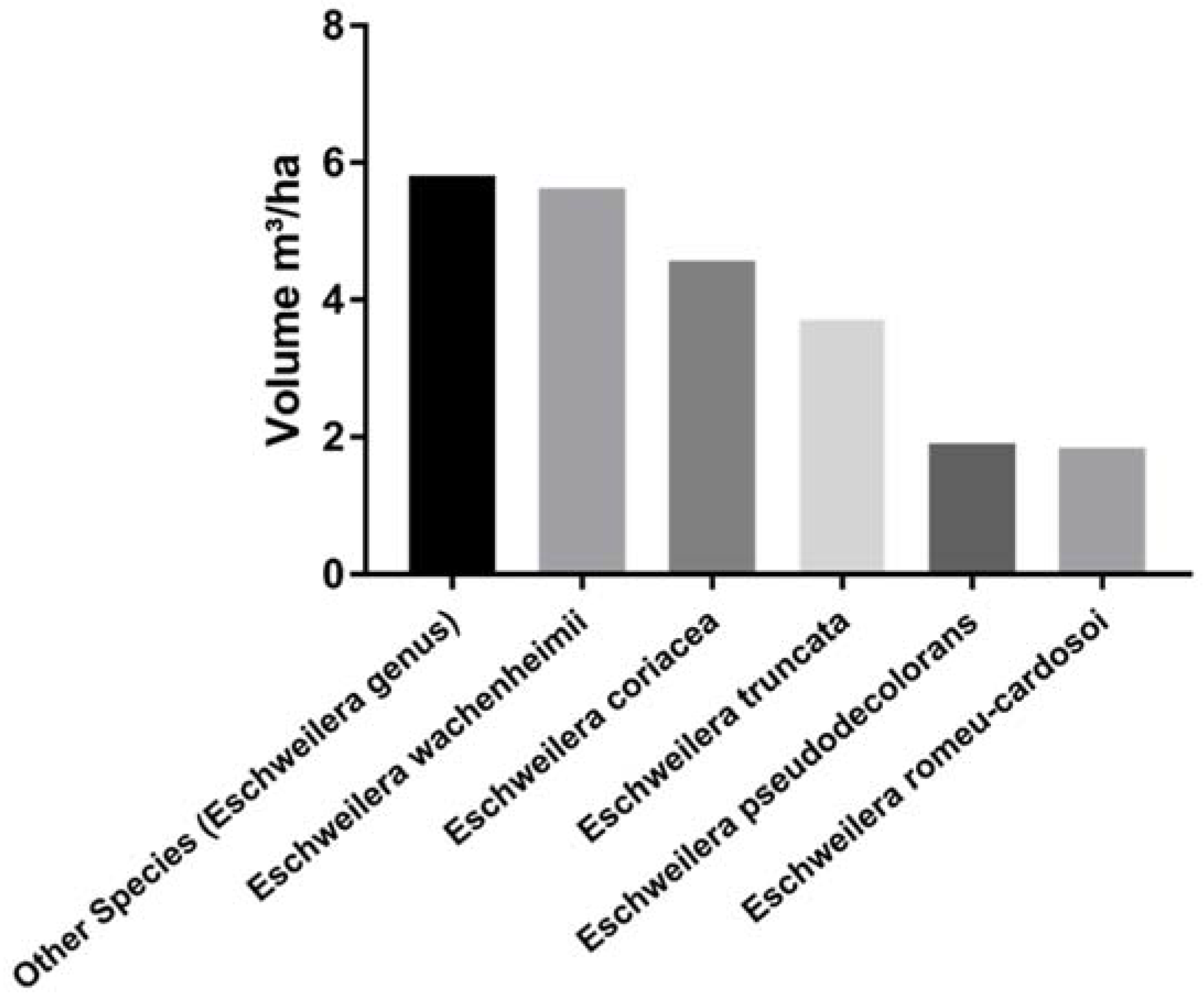
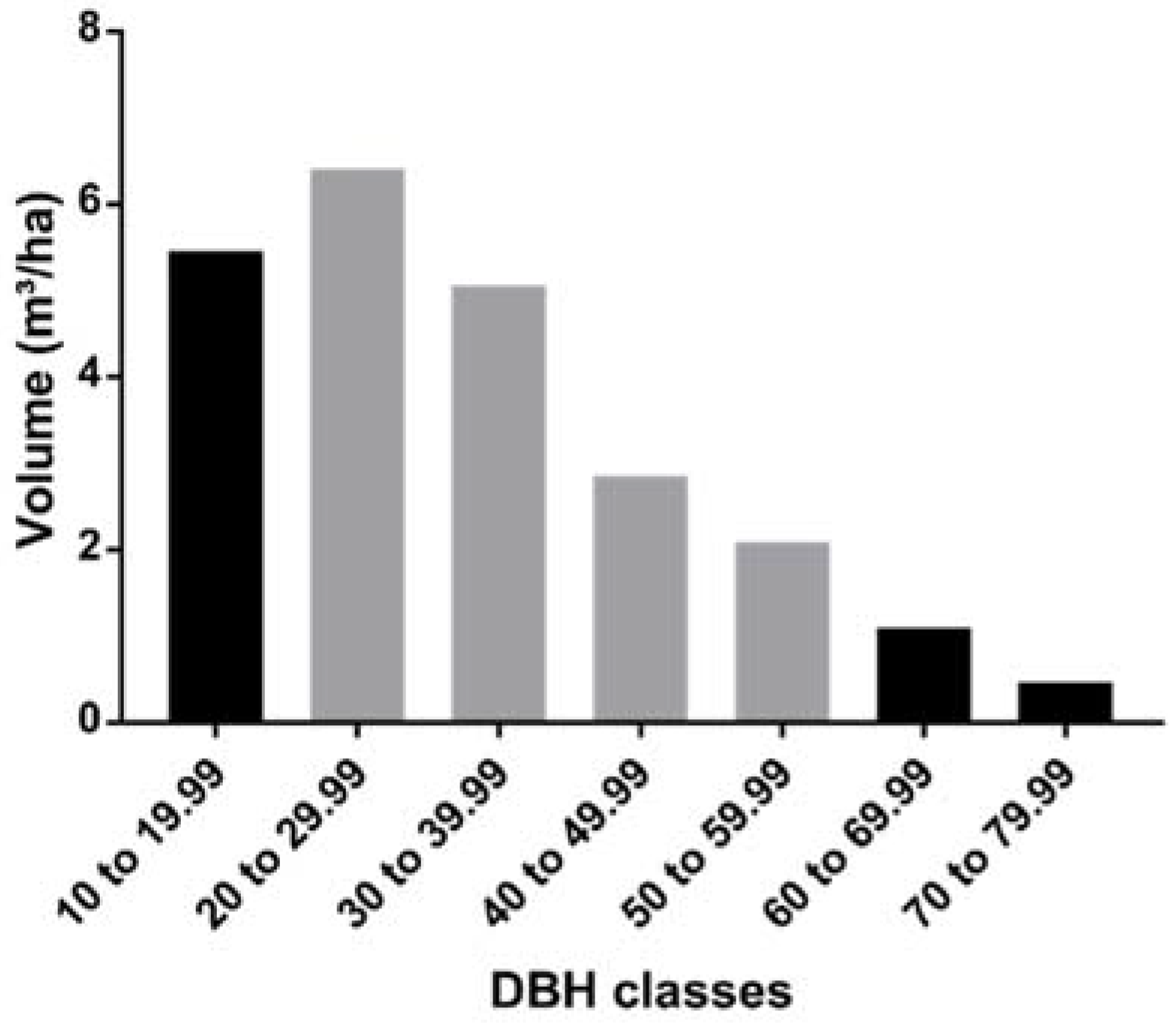
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Higuchi, N.; dos Santos, J.; Ribeiro, R.J.; Minette, L.; Biot, Y. Biomassa da parte aérea davegetação da floresta tropical úmida de terra-firme da amazonia. Acta Amazon. 1998, 28, 153–166. [Google Scholar] [CrossRef]
- Magnabosco Marra, D.; Higuchi, N.; Trumbore, S.E.; Ribeiro, G.H.P.M.; dos Santos, J.; Carneiro, V.M.C.; Lima, A.J.N.; Chambers, J.Q.; Negrón-Juárez, R.I.; Holzwarth, F.; et al. Predicting biomass of hyperdiverse and structurally complex central Amazonian forests—A virtual approach using extensive field data. Biogeosciences 2016, 13, 1553–1570. [Google Scholar] [CrossRef]
- Ngomanda, A.; Engone Obiang, N.L.; Lebamba, J.; Moundounga Mavouroulou, Q.; Gomat, H.; Mankou, G.S.; Loumeto, J.; Midoko Iponga, D.; Kossi Ditsouga, F.; Zinga Koumba, R.; et al. Site-specific versus pantropical allometric equations: Which option to estimate the biomass of a moist central African forest? For. Ecol. Manag. 2014, 312, 1–9. [Google Scholar] [CrossRef]
- Nelson, B.W.; Mesquita, R.; Pereira, J.L.G.; Garcia Aquino de Souza, S.; Teixeira Batista, G.; Bovino Couto, L. Allometric regressions for improved estimate of secondary forest biomass in the central Amazon. For. Ecol. Manag. 1999, 117, 149–167. [Google Scholar] [CrossRef]
- Alder, D.; Synnott, T.J. Permanent Sample Plot Techniques for Mixed Tropical Forest; Oxford Forestry Institute: Oxford, UK, 1992; p. 124. [Google Scholar]
- Felfili, J.M. Dynamics of the natural regeneration in the Gama gallery forest in central Brazil. For. Ecol. Manag. 1997, 91, 235–245. [Google Scholar] [CrossRef]
- Laurance, W.F.; Ferreira, L.V.; Rankin-de Merona, J.M.; Laurance, S.G.; Hutchings, R.W.; Lovejoy, T.E. Effects of forest fragmentation on recruitment patterns in Amazonian tree communities. Conserv. Biol. 1998, 12, 460–464. [Google Scholar] [CrossRef]
- Laman, T.G. Safety recommendations for climbing rain forest trees with “single rope technique”. Biotropica 1995, 27, 406–409. [Google Scholar] [CrossRef]
- Perry, D.R. A method of access into the crows of emergent and canopy trees. Biotropica 1978, 10, 155–157. [Google Scholar] [CrossRef]
- Ter Steege, H.; Cornelissen, J.H.C. Collecting and studying bryophytes in the canopy of standing rain forest trees. J. Hattori Bot. Lab. 1988, 285, 285–290. [Google Scholar]
- Whitacre, D.F. Additional techniques and safety hints for climbing tall trees, and some equipment and information sources. Biotropica 1981, 13, 286–291. [Google Scholar] [CrossRef]
- de Castilho, C.V.; Magnusson, W.E.; Oliveira de Araujo, R.N.; da Costa Pereira, E.; de Souza, S.S. The use of french spikes to collect botanical vouchers in permanent plots: Evaluation of potential impacts. Biotropica 2006, 38, 555–557. [Google Scholar] [CrossRef]
- James, S.A.; Meinzer, F.C.; Goldstein, G.; Woodruff, D.; Jones, T.; Restom, T.; Mejia, M.; Clearwater, M.; Campanello, P. Axial and radial water transport and internal water storage in tropical forest canopy trees. Oecologia 2003, 134, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.G.; Hubbard, R.M.; Clark, D.A.; Sanford, R.L. Woody-tissue respiration for Simarouba amara and Minquartia guianensis, two tropical wet forest trees with different growth habits. Oecologia 1994, 100, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.A.; Lepsch-Cunha, N. Lecythidaceae of a Central Amazonian moist forest. Mem. N. Y. Bot. Gard. 1995, 75, 1–55. [Google Scholar]
- Ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomão, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.-F.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef] [PubMed]
- Durgante, F.M.; Higuchi, N.; Almeida, A.; Vicentini, A. Species Spectral Signature: Discriminating closely related plant species in the Amazon with Near-Infrared Leaf-Spectroscopy. For. Ecol. Manag. 2013, 291, 240–248. [Google Scholar] [CrossRef]
- Moutinho, V.H.P.; Couto, A.M.; Lima, J.T.; De Aguiar, O.J.R.; De Oliveira Gonçalves Nogueira, M. Energetic characterization of Matá-matá wood from the Brazilian rainforest (Eschweilera Mart Ex Dc). Sci. For. Sci. 2011, 39, 457–461. [Google Scholar]
- Bernardo, A.L.; Reis, M.G.F.; Reis, G.G.; Harrison, R.B.; Firme, D.J. Effect of spacing on growth and biomass distribution in Eucalyptus camaldulensis, E. pellita and E. urophylla plantations in southeastern Brazil. For. Ecol. Manag. 1998, 104, 1–13. [Google Scholar] [CrossRef]
- Calegario, N.; Daniels, R.F.; Maestri, R.; Neiva, R. Modeling dominant height growth based on nonlinear mixed-effects model: A clonal Eucalyptus plantation case study. For. Ecol. Manag. 2005, 204, 11–20. [Google Scholar] [CrossRef]
- Gama, F.F.; dos Santos, J.R.; Mura, J.C. Eucalyptus biomass and volume estimation using interferometric and polarimetric SAR Data. Remote Sens. 2010, 2, 939–956. [Google Scholar] [CrossRef]
- Higuchi, N.; Ramm, W. Developing bole wood volume equations for a group of tree species of central Amazon (Brazil). Commonw. For. Rev. 1985, 64, 33–41. [Google Scholar]
- Da Silva, J.A.A.; Borders, B.E.; Brister, G.H. Estimating tree volume using a new form factor. Commonw. For. Rev. 1994, 73, 14–17. [Google Scholar]
- Larson, P.R. Stem form development of forest trees. For. Sci. 1963, 9, a0001. [Google Scholar]
- Da Silva, R.P.; Dos Santos, J.; Tribuzy, E.S.; Chambers, J.Q.; Nakamura, S.; Higuchi, N. Diameter increment and growth patterns for individual tree growing in Central Amazon, Brazil. For. Ecol. Manag. 2002, 166, 295–301. [Google Scholar] [CrossRef]
- Carneiro, V.M.C. Composição Florística e Análise Estrutural na Floresta Primária de Terra Firme da Bacia do rio Cuieiras, Manaus-AM; Instituto Nacional de Pesquisas da Amazônia (INPA): Manaus, Brazil, 2004; p. 67. [Google Scholar]
- De Assis, R.L.; Wittmann, F. Forest structure and tree species composition of the understory of two central Amazonian várzea forests of contrasting flood heights. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 251–260. [Google Scholar] [CrossRef]
- Lopes, M.A. Population structure of Eschweilera coriacea (DC.) S.A. Mori in forest fragments in eastern Brazilian Amazonia. Rev. Bras. Bot. 2007, 30, 509–519. [Google Scholar] [CrossRef]
- Zent, E.L.; Zent, S. Floristic composition, structure, and diversity of four forest plots in the Sierra Maigualida, Venezuelan Guayana. Biodivers. Conserv. 2004, 13, 2453–2484. [Google Scholar] [CrossRef]
- Schöngart, J. Growth-Oriented Logging (GOL): A new concept towards sustainable forest management in Central Amazonian várzea floodplains. For. Ecol. Manag. 2008, 256, 46–58. [Google Scholar] [CrossRef]
- Husch, B.; Beers, T.W.; Kershaw, J.A., Jr. Forest Mensuration; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Spurr, S.H. Forest Inventory; Ronald Press Co.: New York, NY, USA, 1952; p. 476. [Google Scholar]
- Hohenadl, W. Neue Grundlagen der holzmessung. Forstwiss. Cent. 1922, 44, 184–194. [Google Scholar] [CrossRef]
- Krenn, K.; Prodan, M. Die Bestimmung der Echten Schaftholzformzahl und Ausbauchungsreihe aus Dem Echten Formquotienten; Frankfurt am Main: Frankfurt, Germany, 1944. [Google Scholar]
- Schumacher, F.X.; Dos Santos Hall, F. Logarithmic Expression of Timber-Tree Volume; US Government Printing Office: Washington, WA, USA, 1934.
- Gray, H.R. The Form and Taper of Forest-Tree Stems; Imperial Forestry Institute: Oxford, UK, 1956.
- Danielli, F.E.; Gimenez, B.O.; de Oliveira, C.K.A.; dos Santos, J.; Higuchi, N. Yield modeling in sawing logs of Manilkara spp. (Sapotaceae) in sawmill in the state of Roraima, Brazil. Sci. For. 2016, 44, 641–651. [Google Scholar]
- Brazil CONAMA. Resolução No 406 de 02 de Fevereiro de 2009. Available online: http://www.mma.gov.br/port/conama/legiabre.cfm?codlegi=597 (accessed on 1 May 2017).
- Fauset, S.; Johnson, M.O.; Gloor, M.; Baker, T.R.; Monteagudo, M.A.; Brienen, R.J.W.; Feldpausch, T.R.; Lopez-Gonzalez, G.; Malhi, Y.; ter Steege, H.; et al. Hyperdominance in Amazonian forest carbon cycling. Nat. Commun. 2015, 6, 6857. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.N.; do Amaral, I.L.; Ramos, M.B.P.; Nobre, A.D.; Couto, L.B.; Sahdo, R.M. Composição e diversidade florístico-estrutural de um hectare de floresta densa de terra firme na Amazônia Central, Amazonas, Brasil. Acta Amazon. 2008, 38, 627–641. [Google Scholar] [CrossRef]
- Guariguata, M.R.; Pinard, M.A. Ecological knowledge of regeneration from seed in neotropical forest trees: Implications for natural forest management. For. Ecol. Manag. 1998, 112, 87–99. [Google Scholar] [CrossRef]
- Da Silva, F.; Suwa, R.; Kajimoto, T.; Ishizuka, M.; Higuchi, N.; Kunert, N. Allometric equations for estimating biomass of euterpe precatoria, the most abundant palm species in the Amazon. Forests 2015, 6, 450–463. [Google Scholar] [CrossRef]
- Nogueira, E.M.; Fearnside, P.M.; Nelson, B.W.; Barbosa, R.I.; Keizer, E.W.H. Estimates of forest biomass in the Brazilian Amazon: New allometric equations and adjustments to biomass from wood-volume inventories. For. Ecol. Manag. 2008, 256, 1853–1867. [Google Scholar] [CrossRef]
- Higuchi, N. O desmatamento insustentável na Amazônia. Ciência Hoje 2006, 39, 67–71. [Google Scholar]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Kanninen, M. Allometric models for tree volume and total aboveground biomass in a tropical humid forest in Costa Rica1. Biotropica 2005, 37, 2–8. [Google Scholar] [CrossRef]
- Kitahara, F.; Mizoue, N.; Yoshida, S. Effects of training for inexperienced surveyors on data quality of tree diameter and height measurements. Silva Fenn. 2015, 44, 657–667. [Google Scholar] [CrossRef]
- De Almeida, D.R.A.; Nelson, B.W.; Schietti, J.; Gorgens, E.B.; Resende, A.F.; Stark, S.C.; Valbuena, R. Contrasting fire damage and fire susceptibility between seasonally flooded forest and upland forest in the Central Amazon using portable profiling LiDAR. Remote Sens. Environ. 2016, 184, 153–160. [Google Scholar] [CrossRef]
- Schietti, J.; Martins, D.; Emilio, T.; Souza, P.F.; Levis, C.; Baccaro, F.B.; da Pinto, J.L.P.V.; Moulatlet, G.M.; Stark, S.C.; Sarmento, K.; et al. Forest structure along a 600 km transect of natural disturbances and seasonality gradients in central-southern Amazonia. J. Ecol. 2016, 104, 1335–1346. [Google Scholar] [CrossRef]
- Gimenez, B.O.; Danielli, F.E.; De Oliveira, C.K.A.; Dos Santos, J.; Higuchi, N. Volume equations for merchantable timber species of Southern Roraima state|Equações volumétricas para espécies comerciais madeireiras do sul do estado de Roraima. Sci. For. Sci. 2015, 43, 291–301. [Google Scholar]
- Hunter, M.O.; Keller, M.; Victoria, D.; Morton, D.C. Tree height and tropical forest biomass estimation. Biogeosciences 2013, 10, 8385–8399. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No. of Trees |
|---|---|
| Eschweilera atropetiolata S.A.Mori | 1 |
| Eschweilera bracteosa (Poepp. ex O.Berg) Miers | 1 |
| Eschweilera coriacea (DC.) S.A.Mori | 6 |
| Eschweilera grandiflora (Aubl.) Sandwith | 1 |
| Eschweilera micrantha (O.Berg) Miers | 1 |
| Eschweilera pedicellata (Rich.) S.A.Mori | 4 |
| Eschweilera pseudodecolorans S.A.Mori | 5 |
| Eschweilera sp. | 7 |
| Eschweilera truncata A.C.Sm | 8 |
| Eschweilera wachenheimii (Benoist) Sandwith | 30 |
| Total | 64 |
| No. | Model | Author |
|---|---|---|
| 1 | Vi = a × DBHib + ε | Husch [31] |
| 2 | Ln (Vi) = a + b × ln DBHi + ε | Husch [31] |
| 3 | Vi = a + b × DBHi2 + ε | Spurr [32] |
| 4 | Vi = a + b × DBHi + c × DBHi2 + ε | Hohenadl/Kreen and Prodan [33,34] |
| 5 | Vi = a × DBHib × Hic + ε | Schumacher and Hall [35] |
| 6 | Ln (Vi) = a + b × ln DBHi + c × ln Hi + ε | Schumacher and Hall [35] |
| 7 | Vi = a + b × (DBHi2 × Hi) + ε | Spurr [32] |
| 8 | Vi = a × (DBHi2 × Hi)b + ε | Spurr [32] |
| ID | Model | Coefficients | p | R2 | S.E.E (m3) | |
|---|---|---|---|---|---|---|
| 1 | V = a × DBHb | a | 0.00032 | 0 | 0.97 | 0.07 |
| b | 2.242681 | 0 | ||||
| c | - | - | ||||
| 2 | Ln (V) = a + b × ln DBH | a | −8.413954 | 0 | 0.97 | 0.08 |
| b | 2.348828 | 0 | ||||
| c | - | - | ||||
| 3 | V = a + b × DBH2 | a | −0.068854 | 0 | 0.97 | 0.07 |
| b | 0.000841 | 0 | ||||
| c | - | - | ||||
| 4 | V = a + b × DBH + c × DBH2 | a | 0.019768 | 0.743 | 0.97 | 0.07 |
| b | −0.00712 | 0.135 | ||||
| c | 0.00096 | 0 | ||||
| 5 | V = a × DBHb × Hc | a | 0.000089 | 0 | 0.99 | 0.04 |
| b | 2.107657 | 0 | ||||
| c | 0.70496 | 0 | ||||
| 6 | Ln (V) = a + b × ln DBH + c × ln H | a | −9.18928 | 0 | 0.99 | 0.05 |
| b | 2.023415 | 0 | ||||
| c | 0.769948 | 0 | ||||
| 7 | V = a + b × (DBH2 × H) | a | 0.003958 | 0.657 | 0.98 | 0.05 |
| b | 0.000062 | 0 | ||||
| c | - | - | ||||
| 8 | V = a × (DBH2 × H)b | a | 0.000052 | 0 | 0.98 | 0.05 |
| b | 1.018465 | 0 | ||||
| c | - | - | ||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gimenez, B.O.; Dos Santos, L.T.; Gebara, J.; Celes, C.H.S.; Durgante, F.M.; Lima, A.J.N.; Dos Santos, J.; Higuchi, N. Tree Climbing Techniques and Volume Equations for Eschweilera (Matá-Matá), a Hyperdominant Genus in the Amazon Forest. Forests 2017, 8, 154. https://doi.org/10.3390/f8050154
Gimenez BO, Dos Santos LT, Gebara J, Celes CHS, Durgante FM, Lima AJN, Dos Santos J, Higuchi N. Tree Climbing Techniques and Volume Equations for Eschweilera (Matá-Matá), a Hyperdominant Genus in the Amazon Forest. Forests. 2017; 8(5):154. https://doi.org/10.3390/f8050154
Chicago/Turabian StyleGimenez, Bruno O., Leandro T. Dos Santos, Jonas Gebara, Carlos H. S. Celes, Flávia M. Durgante, Adriano J. N. Lima, Joaquim Dos Santos, and Niro Higuchi. 2017. "Tree Climbing Techniques and Volume Equations for Eschweilera (Matá-Matá), a Hyperdominant Genus in the Amazon Forest" Forests 8, no. 5: 154. https://doi.org/10.3390/f8050154