
Novelty and Its Ecological Implications to Dry Forest Functioning and Conservation


1
International Institute of Tropical Forestry, USDA Forest Service, 1201 Ceiba, Jardín Botánico sur, Río Piedras 00926-1115, Puerto Rico
2
Consulting Research Ecology, 2947 NE 31st Ave., Portland, OR 97212, USA
*
Author to whom correspondence should be addressed.
Forests 2017, 8(5), 161; https://doi.org/10.3390/f8050161
Submission received: 24 February 2017
/
Revised: 12 April 2017
/
Accepted: 6 May 2017
/
Published: 10 May 2017
(This article belongs to the Special Issue Tropical Forest Ecology and Management for the Anthropocene)
Abstract
:Tropical and subtropical dry forest life zones support forests with lower stature and species richness than do tropical and subtropical life zones with greater water availability. The number of naturalized species that can thrive and mix with native species to form novel forests in dry forest conditions in Puerto Rico and the US Virgin Islands is lower than in other insular life zones. These novel dry forests are young (<60 years) with low structural development, high species dominance, and variable species density. Species density is low during initial establishment and increases with age. At the 1-ha scale, novel forests can have greater species density than mature native forests. Species groups, such as nitrogen-fixing species, and other naturalized species that dominate novel dry forests, have a disproportional influence on forest element stoichiometry. Novel dry forests, compared to the mean of all forest species assemblages island-wide, tend to have fallen leaf litter with lower than average manganese and sodium concentrations and lower than average C/N and C/P ratios. After accounting for significant differences in stand age, geology, and or precipitation, novel dry forests compared to native dry forests have higher C anomalies, lower Ca and Na anomalies, and lower C/N ratio anomalies. Taken together, these characteristics may influence litter decomposition rates and the species composition, diversity, and food web dynamics in litter and soil. Novel dry forests also contribute to the conservation of native plant species on highly degraded lands.
1. Introduction
Humans are attracted to tropical and subtropical dry forest life zones (sensu lato [1]; dry life zone(s) from now on) because the climate is favorable to their health, agricultural activity, and fuelwood production [2,3]. The consequences of human activity to dry forests are well documented, as these forests are converted to non-forest land covers such as pastures or agriculture, or their aboveground stem wood biomass is unsustainably removed to satisfy fuelwood demand [2,4]. Soil degradation in the form of compacted, nutrient-depleted, or eroded soils, is a common outcome of intensive human activity in dry life zones [4]. This activity tends to fragment the landscapes of dry life zones [5].
When land use pressure is reduced on deforested dry forest landscapes, successional processes allow for the re-establishment of dry forests, as has been observed in Central America, the Caribbean, and other tropical countries [6,7]. In Puerto Rico, recurring island-wide forest inventories uncovered the phenomena that forest succession after abandonment of agricultural use resulted in forest stands dominated by introduced species [8]. Hobbs et al. [9] identified these forests as novel forests, because they are a consequence of human activity, result in new species combinations, and are expanding in land cover throughout the world [10]. Novel forests are “the new wild” of the Anthropocene Epoch [11], and represent a “new world order” [10]; in Puerto Rico, 75 percent of the forest cover is now novel [12].
One of the conservation challenges of the Anthropocene is to characterize novel forests and identify their structure and functioning. Our objective is to assess the ecological implications of novelty in Puerto Rico’s dry forests through a synthesis of published information coupled with a new analysis of island-wide fallen leaf chemistry, first reported in Erickson et al. [13]. To accomplish this goal, we need to place dry forests in environmental and historical contexts so that the adaptive role of novelty can be revealed. Therefore, we first consider the effects of climate and land use history on Puerto Rican dry forests and then summarize novel dry forest structure and leaf litter chemistry in relation to the species composition of stands. We end with a discussion on the implications of novelty to dry forest functioning and conservation.
2. Methods
We review the dry forest literature for Puerto Rico and the Caribbean with particular attention to forest structure and functioning. To display the climatic conditions of dry forest assemblages, we used a moisture availability index applied to forests throughout Puerto Rico and the US Virgin Islands by Brandeis et al. in their study of forest species assemblages in those islands [14]. The index (C/mm) is the quotient of air temperature in degrees centigrade (C) and rainfall in millimeters (mm). It was based on 30-year average annual rainfall and air temperature for each of the 22 species assemblages (used here interchangeably with “forest”) in their analysis. Brandeis et al. [14] analyzed the forest communities of Puerto Rico and the US Virgin Islands using the results of island-wide forest inventories. From their Tables 8 and 9 [14], we selected nine novel and eight native dry forest species assemblages for comparisons of forest structure. These assemblages were all successional and of similar age (<60 years), the main difference being their species composition.
Species Importance Value (IV) curves (sensu Whittaker [15,16]) are used to establish species dominance in forests and infer levels of stress. Ranking species according to their IV, which is an index that includes the species basal area and stem density relative to those of the stands where they occur, assesses species dominance. Whittaker [15,16] showed that the steep IV curves approach geometric series, while the flatter IV curves approach lognormal distributions and suggested that steeper curves reflected communities under stress. Another structural index that we used to assess dry forest stature was the Holdridge Complexity Index. Holdridge [1] used this Index (based on forest structural measures and number of species) to show that forest complexity increased with moisture availability.
For the analysis of element chemistry and stoichiometry of novel and native dry forests, we build on the study of Erickson et al. [13] who analyzed fallen leaf litter mass and chemistry (11 elements) in 140 plots located across Puerto Rico within 14 of the species assemblages described by Brandeis et al. [14]. Using the same data set as in Erickson et al. [13], we selected the five driest forest communities for a total of 41 plots. Although these communities are commonly found in the Dry Forest Life Zone (sensu Holdridge), each community contains plots located in wetter life zones. We note that there are novel forest communities with greater mean annual precipitation than these five that are considered in the Erickson et al. [13] paper but not here, where the emphasis is on drier novel and native communities. Modeled mean annual precipitation (cf. [13]) for individual plots in this study ranges from 787 mm to 2322 mm. Although the species assemblages used in this analysis are identified by the dominant tree species, the leaf litter samples that were chemically analyzed represent the litter of all the species in the stand, not necessarily a monospecific leaf litter.
To calculate concentration anomalies for the five forest assemblages (three novel and two native), for each element and ratio we subtracted a mean value based on all plots within the 14 island-wide assemblages (roughly 139, depending on element) from the mean of each assemblage. Thus, the concentration anomalies of fallen leaves establish the stoichiometry of dry forests (novel and native) in relation to the corresponding mean for all plots in all species assemblages island-wide. These concentration anomaly comparisons are conservative given that the island-wide averages include the dry forest means. Concentration anomalies for all novel plots together and all native plots together were compared to island-wide means (zero on the anomaly graphs) for each element using T-Tests (SAS version 9.4, SAS Institute, Cary, NC, USA).
We tested for individual differences in concentration anomalies among the five forest assemblages, which would suggest idiosyncratic community-scale responses, using ANOVA followed by a post-hoc Tukey–Kramer analysis (SAS Institute, Cary, NC, USA). We also tested whether anomalies differed between novel and native dry forest assemblages using contrast statements. Variables, except for carbon, nitrogen, sulfur, calcium, magnesium and the N/P ratio, which were normally distributed, were log-transformed to meet assumptions of tests. Occasional extreme outliers were removed to further improve normality. The presence of karst has been shown to influence fallen leaf C, Ca, Mn, Al, and Fe chemistry [13], and only about a third of novel plots were located on karst compared to about 75 percent of the native plots. Similarly, forest assemblages differed in mean annual precipitation and mean midpoint age (cf. [13]), also shown to influence fallen leaf chemistry for some elements [13]. Accordingly, we retained mean annual precipitation and stand age as co-variates and accounted for presence/absence of karst in models if significant at the 0.05 level.
3. Results
3.1. Dry Forests in General
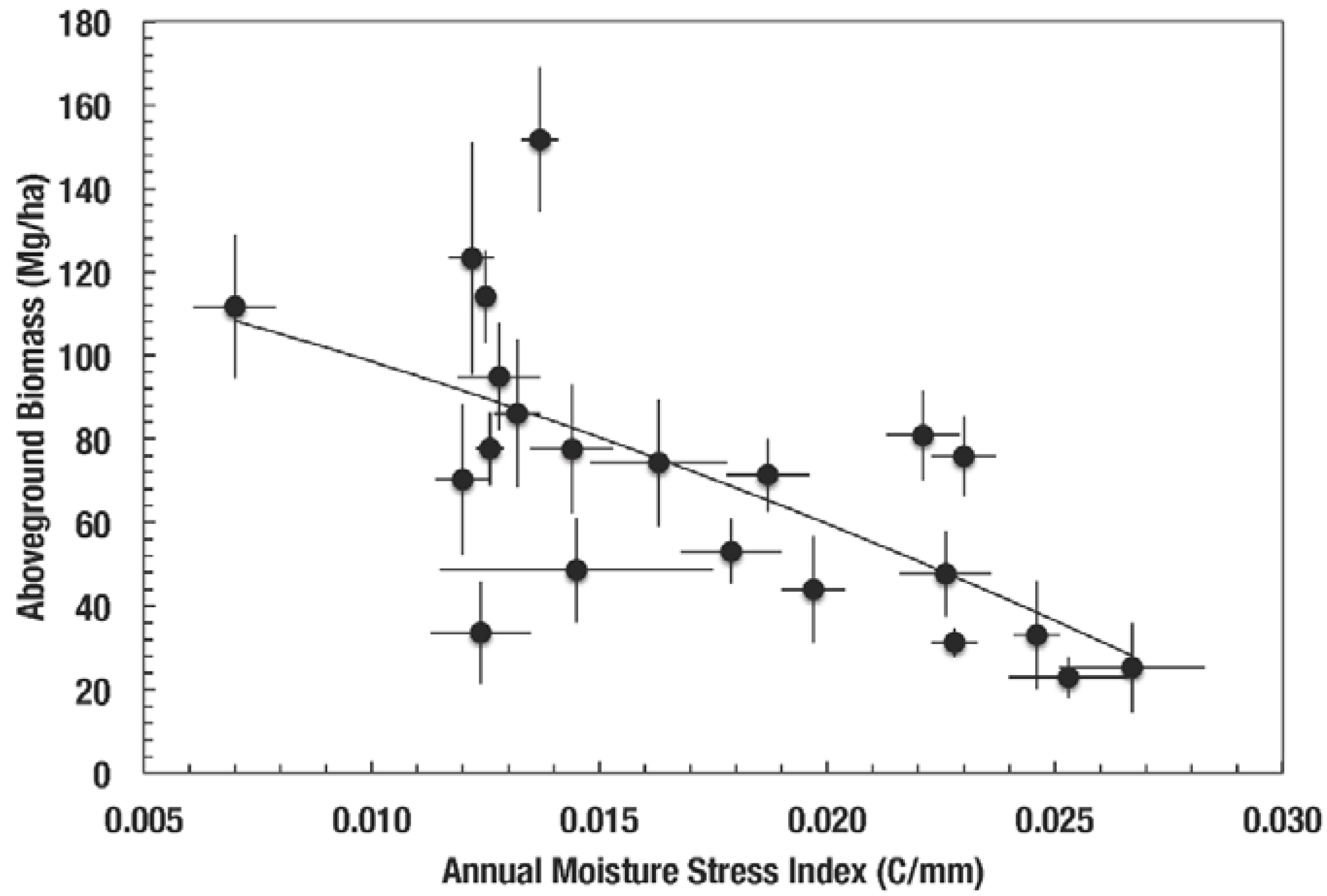
Dry forest environmental conditions affect forest structural development and species composition. Dry forests have the lowest complexity, species richness, and stature among tropical forests. Brown and Lugo [17] showed that carbon accumulation in vegetation and soil and litterfall were lower in tropical and subtropical dry forests compared to tropical and subtropical moist, wet, and rain forests. Martínez Yrizar [18] found a positive relationship between rainfall and aboveground biomass and litterfall for dry forests from different tropical locations. For Puerto Rico, biomass was low in forests with the lower moisture availability conditions (Figure 1). Moreover, Gentry [19] found that the dry forest flora of the Neotropics was less diverse than that of moist and wet forests, and that Caribbean dry forests were at the lower end of the species richness gradient of dry forests. He thought that the climate of dry forests coupled to limestone substrates in the Caribbean limited the diversification of its dry forest flora.
3.2. Naturalized Species and the Structure of Novel Dry Forests
Fifteen percent of the plant species of the Caribbean are introduced species [20], most of which naturalize, i.e., establish self-sustaining wild populations. Introduced species can represent between 43 to 110 percent of the native insular flora of individual oceanic islands in the Pacific [21]. In Puerto Rico, the number of naturalized tree species varies with life zone, peaking in the moist and wet forests with lower numbers in rain and dry forest climates. Francis and Liogier [22] listed 118 tree species as naturalized to Puerto Rico (about 18 percent of the tree flora). Of these, approximately 29 species grow in the dry life zone (annual rainfall below 1000 mm), compared to about 43 in the moist life zone (between 1000 and 2000 mm annual rainfall). Thirty-one other tree species grow in an annual rainfall range between 1500 and 3800 mm. This means that the number of tree species available to colonize degraded sites and remix with native species to form novel forests is reduced in the dry life zone compared to moist or wet life zones.
Novel dry forests in Puerto Rico and the US Virgin Islands are characterized by lower tree density, basal area, aboveground biomass, and tree height than native dry forests of similar age (<60 years) (Table 1). The age of dry forest native stands tends to be on the higher end of the age range but they are also secondary forests recovering from agricultural disturbance. The age category for most novel forest stands in Brandeis et al. [14] was 23 to 49 years, which helps explain the low level of structural development. The level of dominance of the top ranked species was similar in both native and novel dry forests, but native stands tended to have more species than novel ones [14,23,24].
Early succession novel dry forests (<60 years), such as those studied by Molina Colón et al. [23], support few species with very high dominance (up to 90 percent), reflecting the initial colonization of deforested sites by a few species. Successional dry novel forests exhibit higher dominance than nearby mature historic dry forests (>80 years), and their species density is lower [24,25]. Older novel dry forests support more species than mature native dry forests at the 1-ha scale, but have less species density than mature historic forests at smaller sampling scales [26]. Native species that are unable to colonize degraded sites are able to grow in sites colonized by non-native tree species, thus increasing the diversity of novel forests [24,25,26].
3.3. Stoichiometry of Leaf Litter
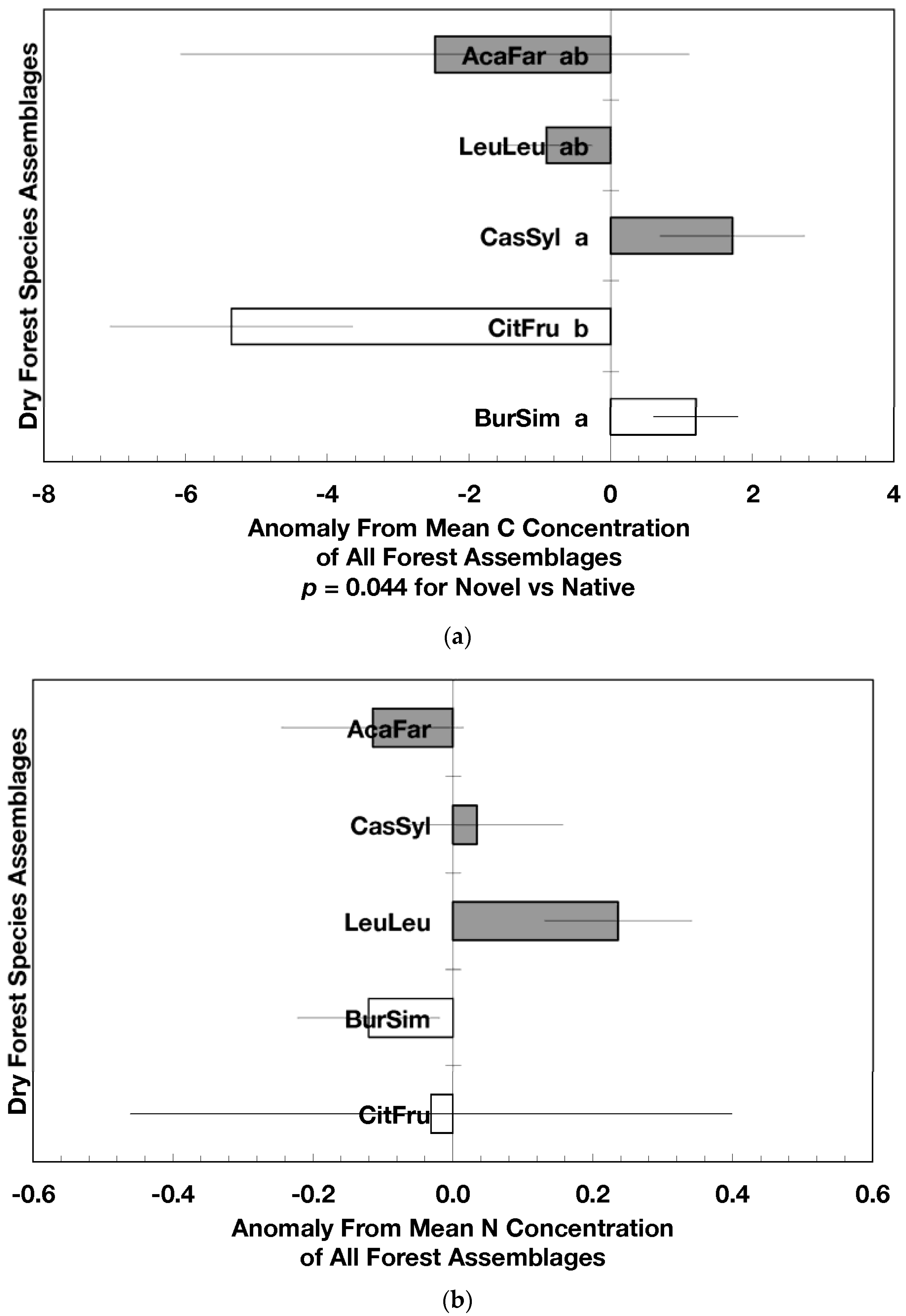
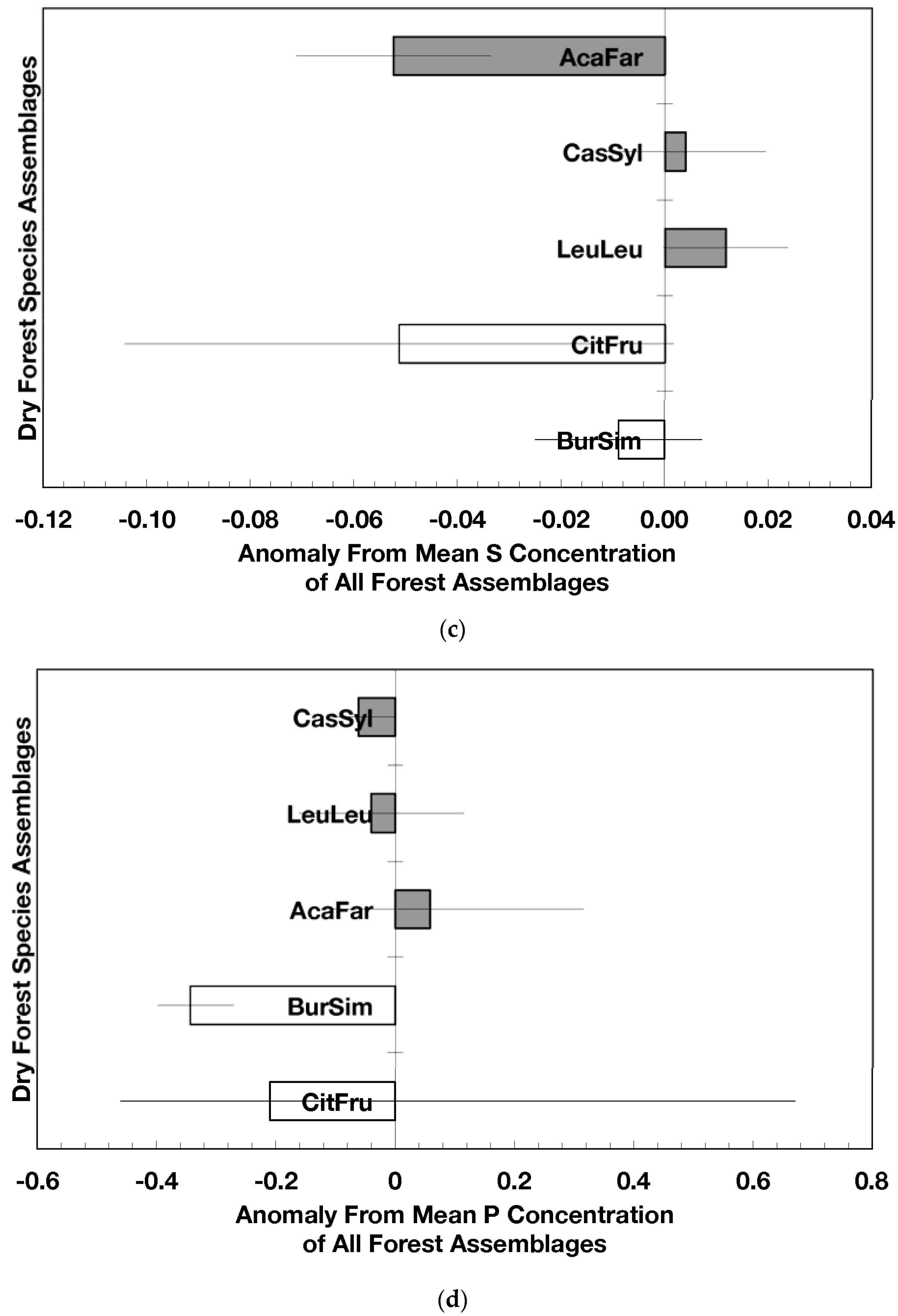
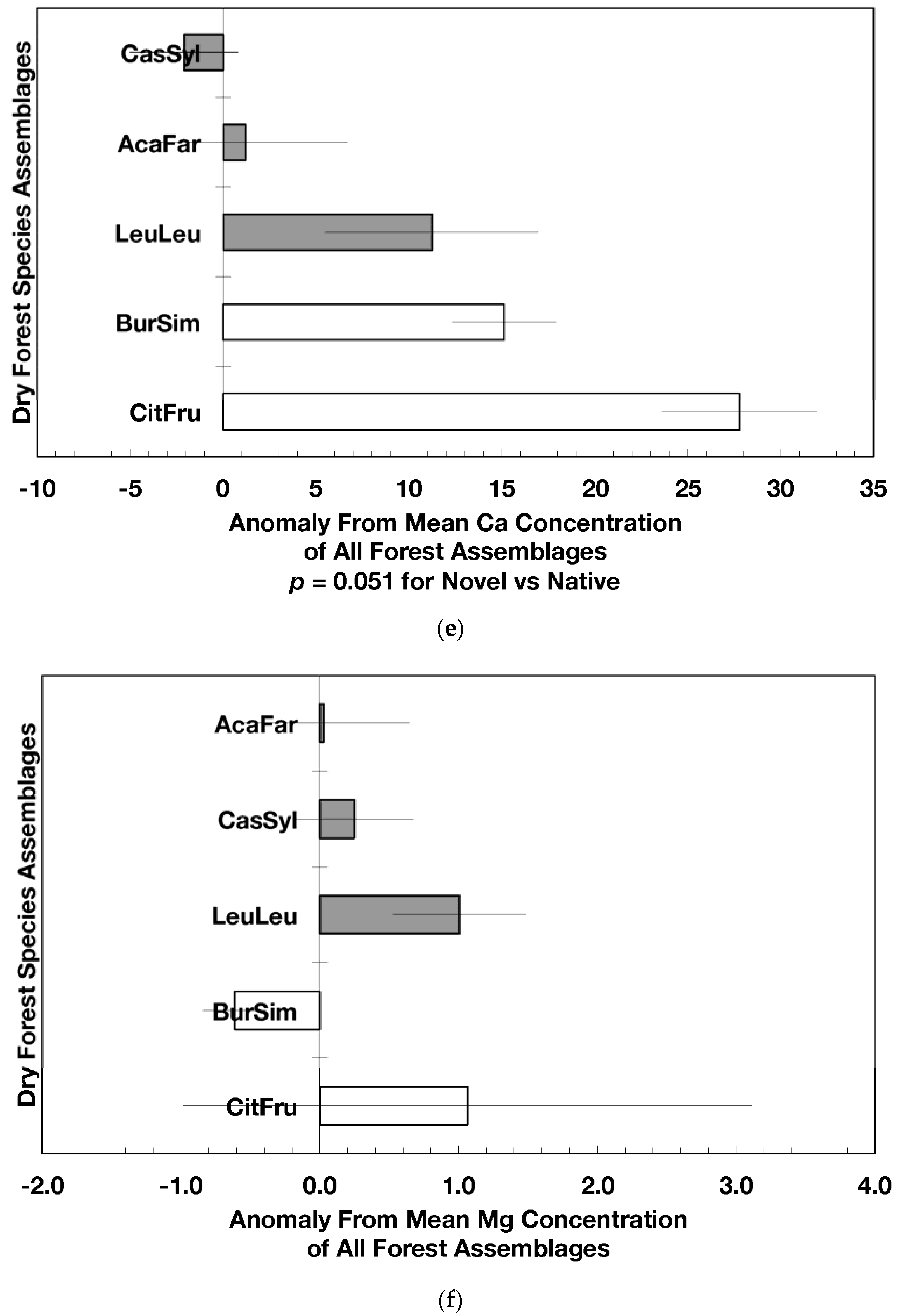
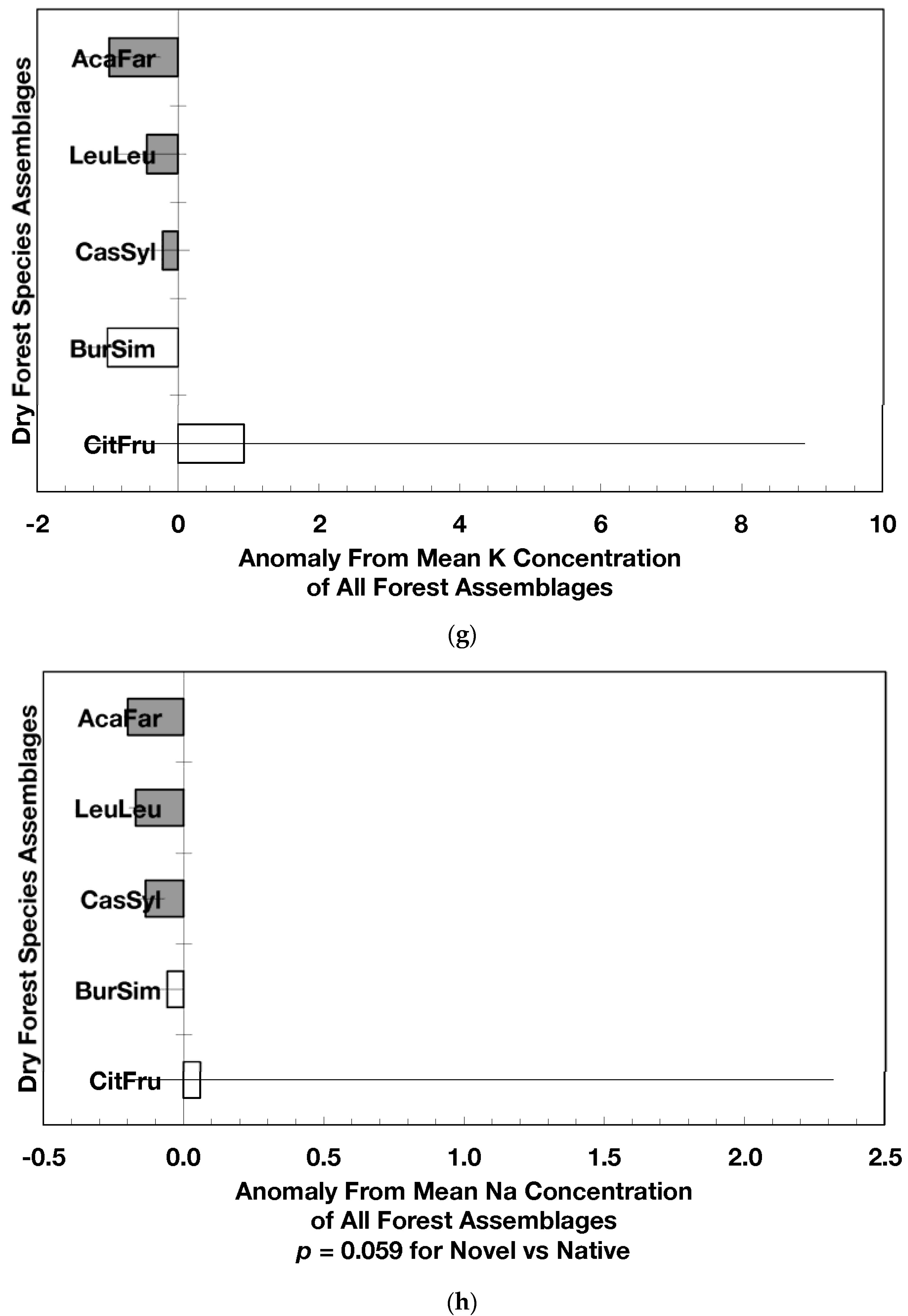
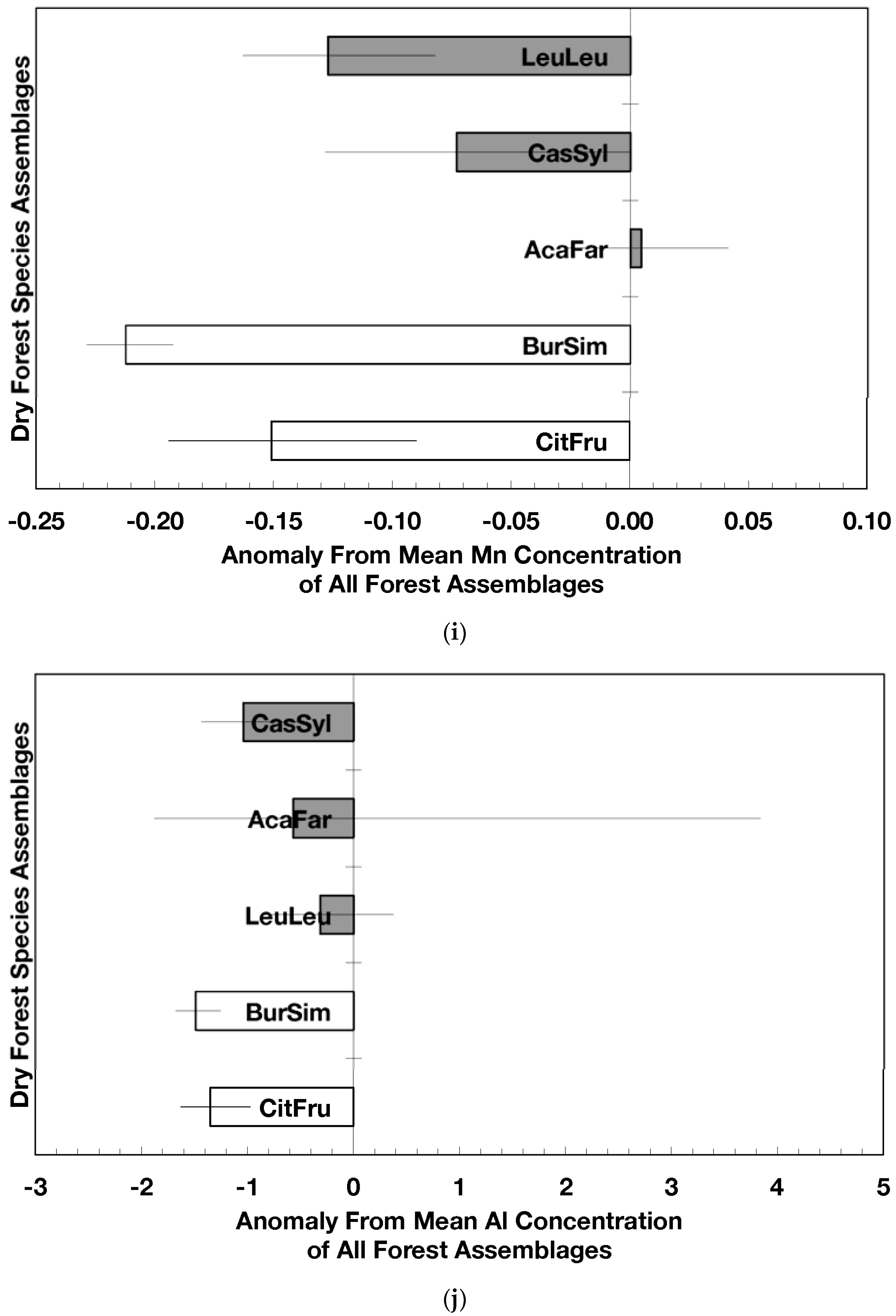
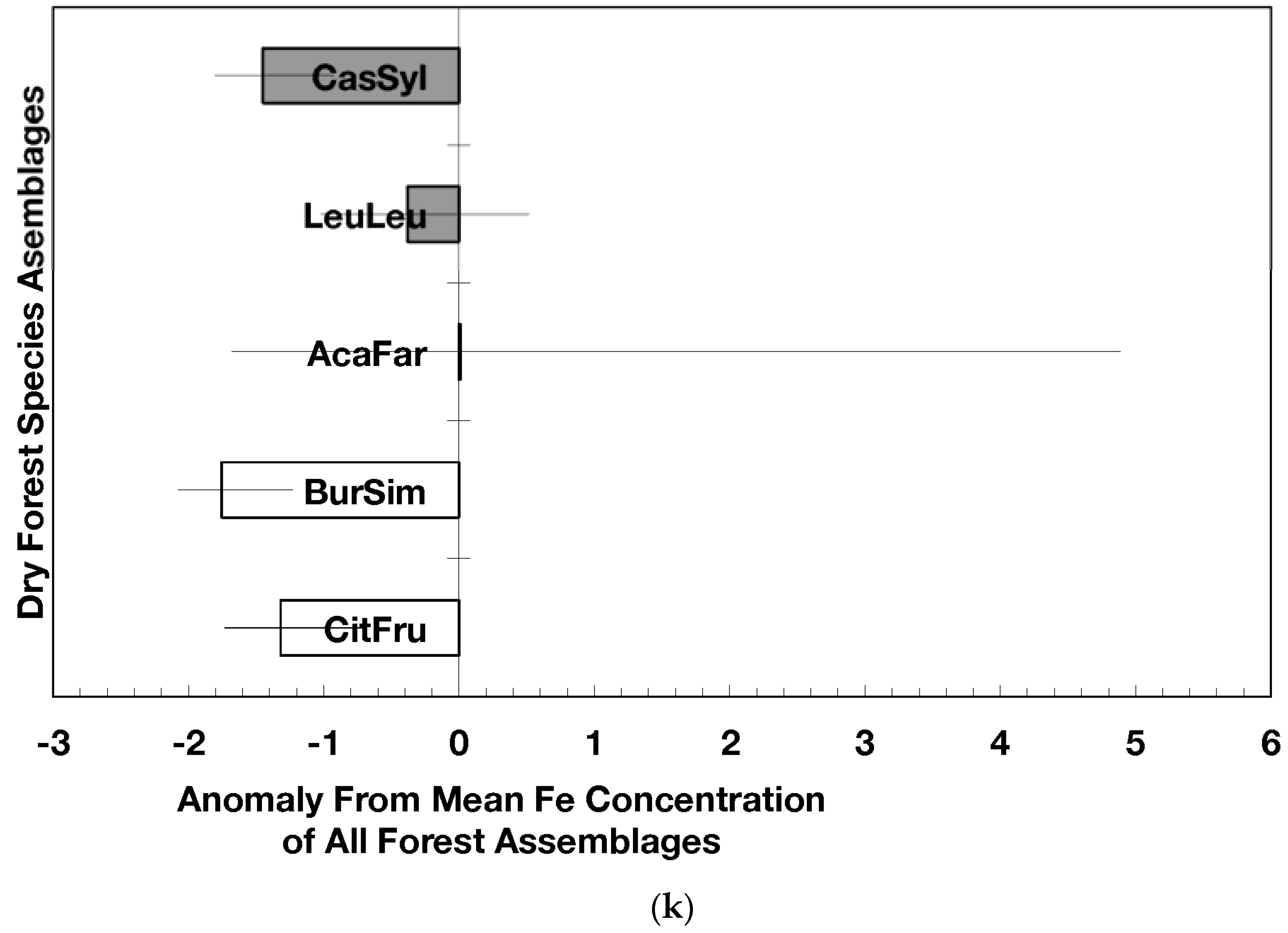
The overarching pattern in the comparisons of element concentration anomalies between novel and native dry forests and with island-wide forests is the absence of a consistent pattern among elements. After accounting for differences in stand age, C concentration anomalies (p = 0.044) were greater in novel dry forests than in native dry forests (Figure 2a) although differences among individual assemblages also existed. For example, native stands dominated by Citharexylum had significantly lower carbon concentration anomalies (p = 0.004, Figure 2a) than other assemblages. Novel dry forests tended to show lower Ca (p = 0.051) and Na (p = 0.059) anomalies than native dry forests Figure 2e,h). Nitrogen, sulfur, phosphorus, magnesium, potassium, manganese, aluminum, and iron concentration anomalies did not differ among the forest assemblages or between novel and native dry forests (Figure 2b–d,f,g,i–k).
Native dry forest concentration anomalies tended to differ from island-wide averages more than novel dry forest concentration anomalies. Native forests had significantly lower P, Al, and Fe concentration anomalies than island-wide averages (p = 0.015, 0.0007, and 0.026, respectively), while novel forest anomalies for these elements did not differ from island-wide averages (Figure 2d,j,k). Native dry forests had higher Ca concentration anomalies than the island-wide average (p < 0.0001, Figure 2e). Both native and novel dry forests had lower Mn anomalies (p = 0.0001 and 0.053, respectively) than the island-wide average Figure 2i). Novel dry forests had significantly lower Na anomalies compared to island-wide averages (p = 0.0006, Figure 2h). These comparisons to island-wide averages do not take into account differences in geology, precipitation, or mean stand age among the assemblages. For example, around 75 percent of the native plots compared to a third of the novel plots were located on karst substrates, which potentially explains their positive Ca and negative P, Al, and Fe anomalies (cf. [13]). As well, native plots tended (p = 0.073) to be older than novel plots (28 versus 15 years). While older plots island-wide have been shown to have lower Al, Fe, and P concentrations in fallen leaves than younger plots [13]; these older plots are greater than 50 years, and only four of the native and three of the novel plots were in this age class. Nonetheless, these tests show how plot location influences, to some degree, the differences that may exist between novel and native forest stands within this dataset; the ANOVA results on the other hand account for effects of the co-variables.
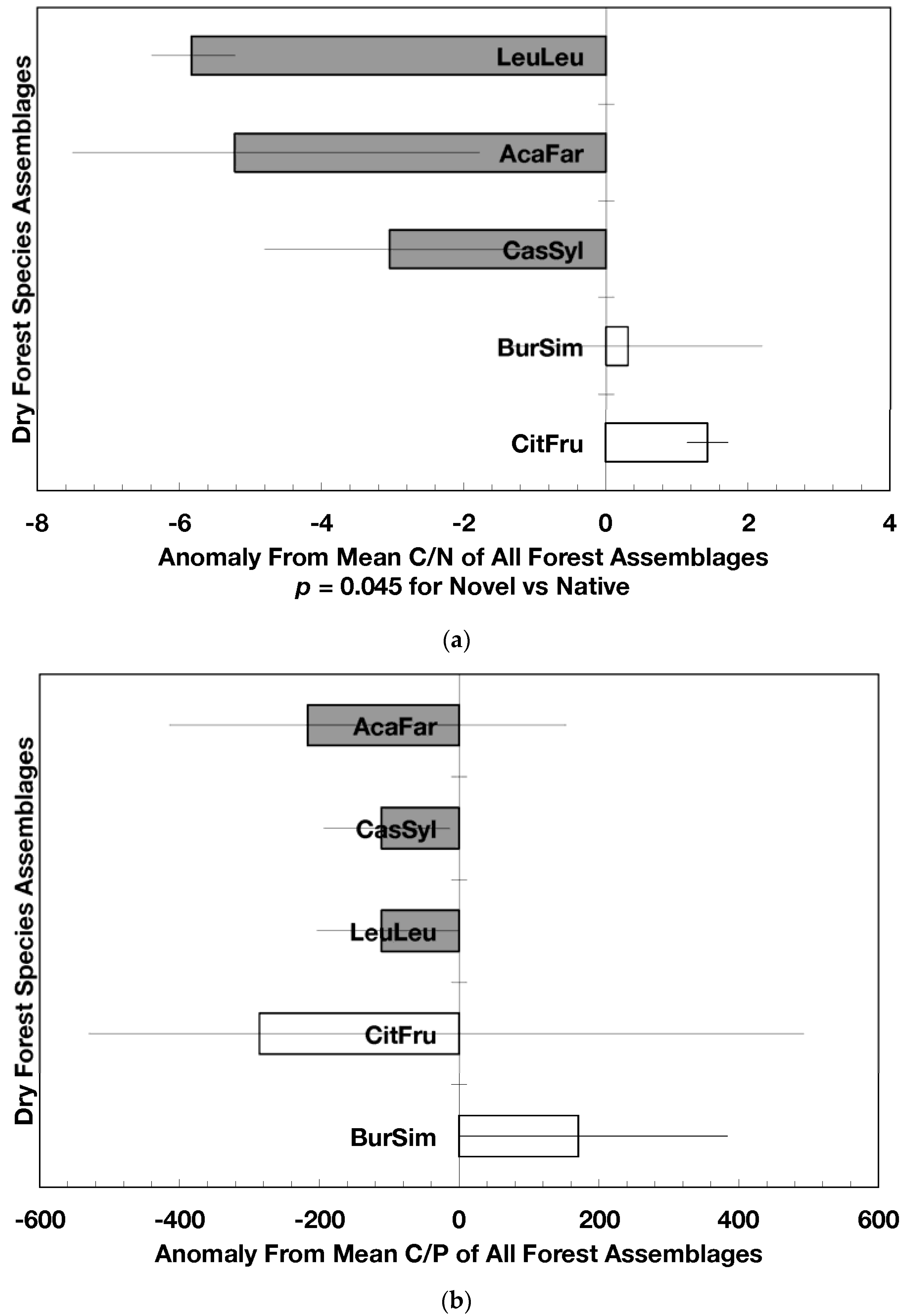
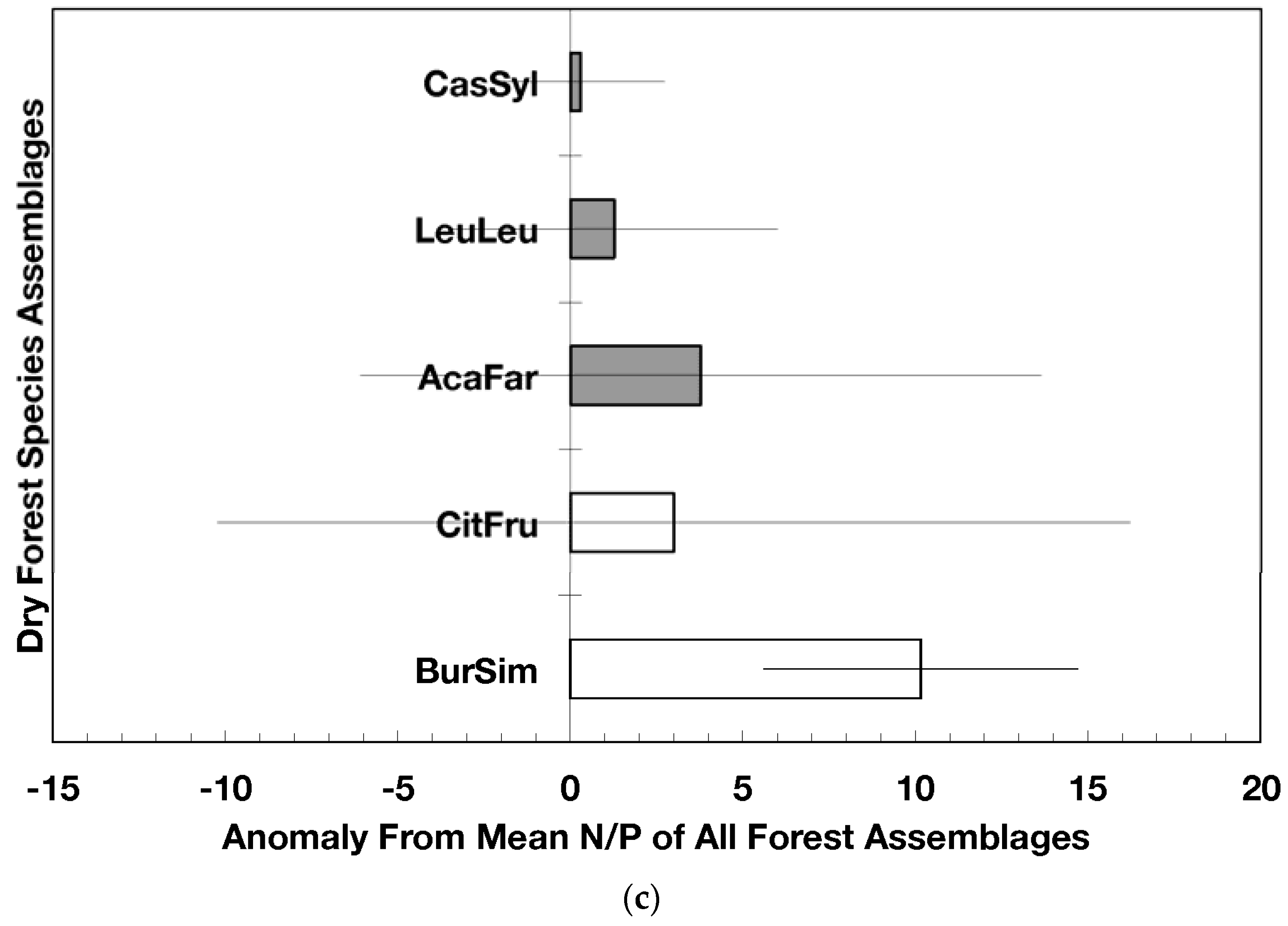
Novel dry forest assemblages had lower C/N ratio anomalies than native assemblages (p = 0.045, Figure 3a); these anomalies were also significantly lower than the island-wide average (p = 0.0017) while those from native dry forests did not differ from the island-wide average. The C/P ratio anomalies did not differ between novel and native dry forest assemblages but novel dry forest assemblages as a group did show significantly lower anomalies (p = 0.0013) than the island-wide average (Figure 3b). The N/P ratio anomalies did not differ between novel and native dry forests (nor between the forest assemblages) though there was a slight indication that native forest N/P anomalies were higher than the island-wide average (p = 0.061; Figure 3c).
Table 2 shows the variation of element and mass accumulation by dry forest species assemblage reported by Erickson et al. [13]. They found no differences in forest floor or fallen leaf mass among species assemblages, but differences in concentration among assemblages (their Table 2) made a difference in the absolute accumulation of phosphorus, calcium, manganese, and aluminum.
4. Discussion
4.1. Species Dominance, Density, and Novelty
Broadly speaking, the forests of Puerto Rico and the Caribbean are characterized by high species dominance and low species density [27]. Typically, the IV curves for Puerto Rican forests are steep with a short tail (few species) in contrast to those of continental tropical forests, which are less steep and have longer tails. Research in Puerto Rico shows that tree species dominance increases with increasing environmental stress such as low or excessive moisture availability or nutrient limitations [27]. As stands mature, tree species dominance decreases through succession. However, even in undisturbed and mature native forests, tree species dominance is higher in Puerto Rico than measured in Amazonian forests [28] but similar to other Caribbean islands [29].
While biogeographical isolation plays a role in the ability of species to disperse from continental regions to islands [30], also at play in the sharp differences in community structure and species density between the Caribbean and Amazonia is the recurrent hurricane disturbance regime of the Caribbean [31]. Lugo [31] argued that hurricane disturbances are partially responsible for the low and similar level of tree species density across all forest types in Puerto Rico (about 45 to 55 tree species per ha), and for the high dominance of species in insular forests [27,32]. However, irrespective of explanation, the empiric reality is that the IV curves for historic Puerto Rican forests are steep with short tails, reflecting high species dominance by a few species, and low species density. How do these characteristics change with novelty?
Novel forests contain novel mixtures of native and non-native tree species, while the naturalized species component is missing in the native stands. The addition of naturalized species increases the species density in novel systems at the 1-ha scale. The age of forest stands also affects species dominance and density. Tree species dominance decreases and species density increases in novel and native dry forests as they age, reflecting the accumulation of mostly native species through succession [23,24,25].
Puerto Rican novel forests in dry and wetter life zones are younger (<60 years) than island-wide historic native forests (>80 years) [33] because their establishment follows land abandonment, which island-wide started in the 1960s, accelerated over the succeeding decades, and finally slowed down at the onset of the 21st century [34]. Geographic scale is important because species area curves of forests in Puerto Rico are steep [26,27], which means that species density changes rapidly with area sampled. However, the species area curve for novel forests is initially less steep than that of historic forests, but reaches a higher plateau than historic forests at the 1-ha scale [26]. We believe that more research is needed in the species/area relationships during the colonizing phases of novel forests, as well as the landscape aspects of native species persistence in novel landscapes.
4.2. The Importance of Species
As moisture conditions change from wet to moist to dry, fewer species are available to colonize degraded sites and those that colonize exert more dominance over their communities. Dominant species occupy more space and process and accumulate more resources than non-dominant species. Thus, the characteristics of novel dry forests will be more dependent on the characteristics of naturalized species colonizing abandoned, degraded sites, because a small group of those species will become very abundant and dominant in the emerging forests.
The ways in which individual species can influence the structure and functioning of forests is through their natural history traits, including their physiognomy, growth characteristics, phenological rhythms, and stoichiometry. Hulshof et al. [35] discuss how species functional traits vary and influence dry forest structure and functioning, as did Lopezaraiza-Mikel et al. [36] for the phenology of dry forest tree species. We illustrate the role of species in forest ecosystem functioning with the stoichiometry of fallen leaf litter.
4.3. Stoichiometry of Novel and Native Dry Forests
The results on the influence of species assemblages on forest-level characteristics (fallen leaf chemistry) underscore the point made earlier that the species composition and age of forests can have relevance to mass and nutrient fluxes of forests. The high chemical quality of fallen leaf litter in novel dry forests (low C/N and C/P) compares favorably with the chemical quality of species assemblages known for their high primary productivity and rapid nutrient cycling [37]. Thus, the fallen leaf chemistry of novel dry forests reflects substrates that are favorable for rapid decomposition and recycling (Table 2).
Erickson et al. [13] found that all dry forests (novel or native) in the dry life zone had significant differences in fallen leaf chemistry when compared to forests in other life zones of Puerto Rico. The dry forests fallen leaves had the lowest concentration of carbon on the island (42 percent), the highest concentrations of calcium (29 mg/g) and magnesium (3.6 mg/g), and the lowest concentrations of sodium (0.11 mg/g). When Puerto Rico’s dry forest results are compared to ten leaf litter chemistry values reported by Jaramillo and Sanford [38] for dry forests in Mexico, India, Belize, Australia, and Puerto Rico (site not included in Erickson et al. [13]), the following patterns emerge: the potassium and magnesium concentrations in Puerto Rico’s dry life zone are lower than those in all but two locations, while the calcium, nitrogen, and phosphorus concentrations are higher than those of all but one location. This comparison suggests more labile leaf litter in Puerto Rican dry forests given the higher nitrogen and phosphorus concentrations. Jaramillo and Sanford [38] also report total mass and nutrient accumulation in litter for the same locations discussed above. Comparisons show lower mass accumulation and similar nutrient content in the leaf litter of Puerto Rican dry forests. A lower leaf litter mass is consistent with more labile leaf litter.
Erickson et al. [13] also found that the youthful age of forests (<60 years) resulted in higher phosphorus, iron, and aluminum concentrations and lower carbon concentrations and C/P and N/P in fallen leaf litter compared to older stands island-wide. They concluded that the availability of nitrogen (discussed below) and phosphorus might be greater in today’s island’s forests than before deforestation. Do novel dry forests exhibit the same element concentrations as native dry forests? The answer to this question is not categorical because the pattern of element concentration for fallen leaf litter at times varied with forest assemblage, mean stand age, geological substrate, or precipitation. Nevertheless, we did find trends and significant differences in element concentrations and ratios between novel and native dry forests in Puerto Rico (Figure 2 and Figure 3). These differences suggest potential acceleration of nutrient and mass fluxes due to the faster decomposition of leaf litter with lower C/N and Ca, i.e., more labile leaf litter. Similarly, the concentration and ratio anomalies for the dry forest communities compared to the grand mean of all plots across the island also suggest potential acceleration of nutrient and mass fluxes as a result of differences between species assemblages. For example, the lower than average C/N and C/P in novel dry forests (Figure 3a,b) support the notion of a faster leaf decomposition rate, which is reflected in a lower accumulation of leaf litter in these forests (Table 2). As we discuss below, species composition and dominance in relation to age and geographic scale play a role not only in the stoichiometry of forest stands but also in their influence in the restoration of degraded sites and continuing process of community assembly.
4.4. Implications of Novelty to Functioning and Conservation of Dry Forests
The landscape of Puerto Rico, like Anthropocene landscapes everywhere, is dynamic and in constant change as a result of human activity [39]. Nevertheless, long-term processes such as the forest transition from a deforested to a forested landscape exert their influence on the age (young) of forests and the particular successional stage in which they find themselves. Molina Colón et al. [23] found that the early stages of dry forest establishment result in patches of dissimilar novel species assemblages (similarity index of 26 percent), whose species composition is dictated by seed sources and their dispersal vectors. The spatial diversity of forest patches may share common species, such as Leucaena, a nitrogen fixing tree, that grows throughout the dry life zone independently of the type of past land use [40]. However, the combination of all patches support more species than found in individual patches, even when they are in close proximity to each other.
Ramjohn et al. [5] found that up to 86 percent of the tree species in a protected area of the dry life zone, were found in novel forest fragments dispersed through the landscape. Fragments of 0.04 ha contained about 45 percent of the protected area tree species while fragments of 33 ha contained 75 percent of the reference species. Thus, the dissimilarity of species combinations among patches counteracts the high dominance and low species density of individual forest patches so that the landscape as a whole conserves the species richness of the dry forest formation. Therefore, novelty of species composition contributes to the colonization of degraded sites and the conservation of native species that otherwise could not colonize degraded sites abandoned after agricultural activities.
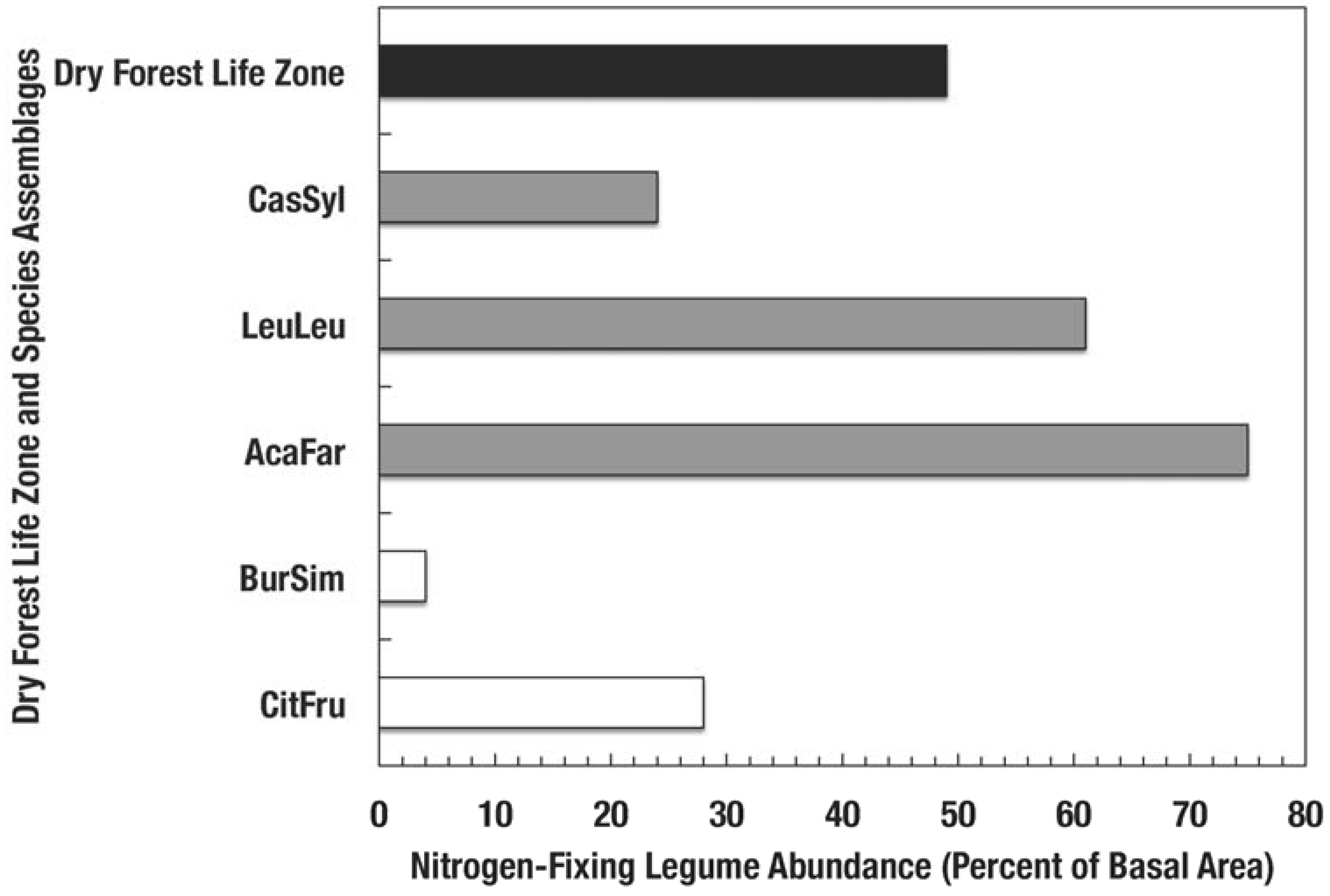
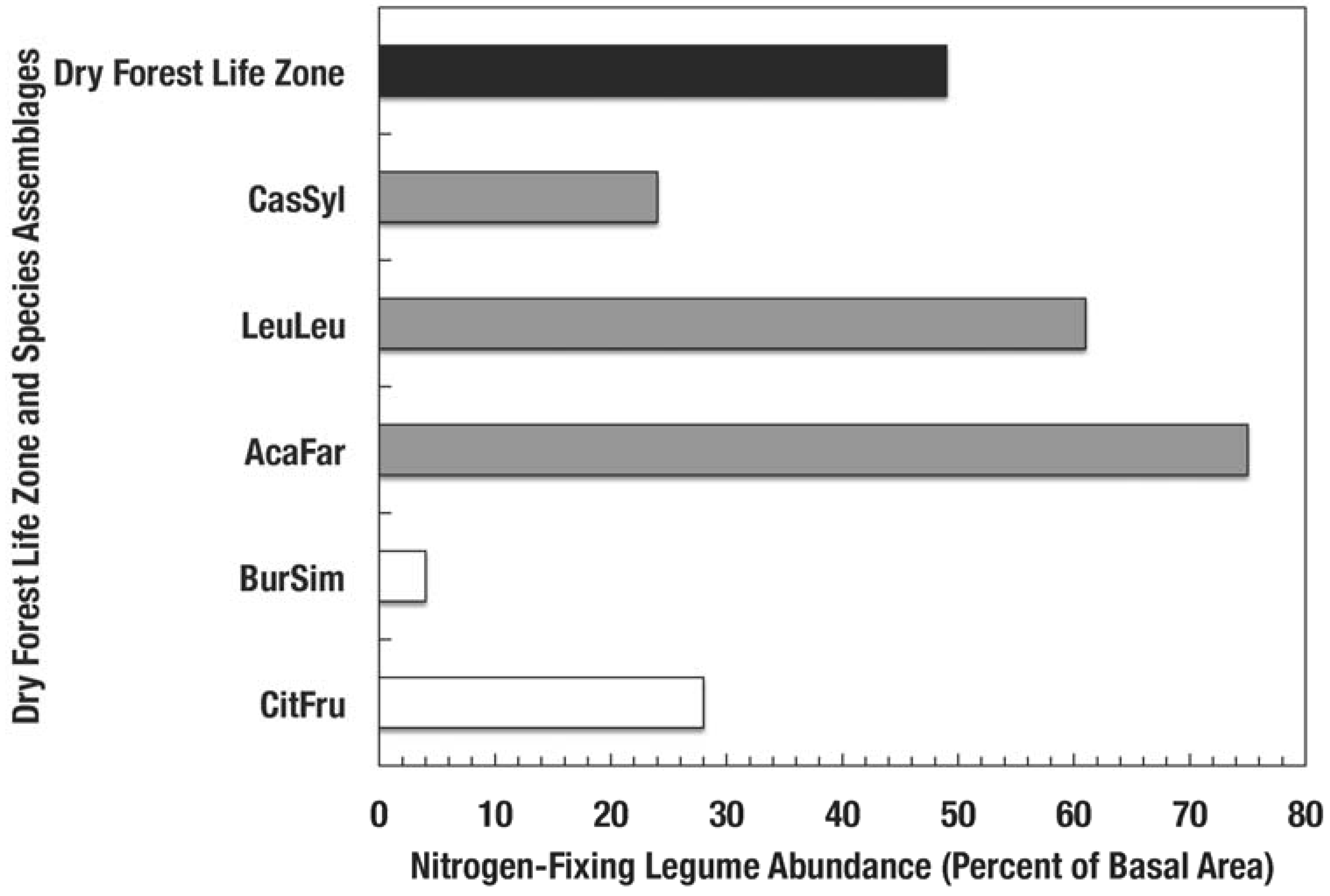
Nitrogen-fixing species such as Leucaena have the capacity to colonize highly degraded soils and re-start arrested succession [13,40,41]. Erickson et al. [13] found that the basal area of nitrogen fixing species in 14 species assemblages in Puerto Rico was positively related to leaf litter phosphorus concentration and negatively correlated with leaf litter C/N ratio. Compared to the native dry forests, the lower C/N ratios (Figure 3a) found in fallen leaf litter from novel forests suggest faster litter decomposition rates [42], which illustrate how species stoichiometry influences ecosystem processes. Moreover, novel dry forests contain more nitrogen fixing species than native dry forests (Figure 4). Because of the abundance of nitrogen fixers and high dominance of naturalized species in novel forests, an individual or group of species with particular concentrations of nutrients can determine the nutrient and carbon fluxes and stoichiometric ratios in forests (Figure 3).
There is no a priori reason to expect that species from different geographical origins would have particular nutrient concentrations or even unique life history traits. However, naturalized species that dominate novel forests do so under strong environmental selective pressure. In dry forests, these include the low moisture conditions (which limit the number of species that overcome this limitation), a deforested landscape (which selects for pioneer species), and degraded soil conditions, which select for particular nutrient use efficiencies and life history strategies. Thus, the stoichiometry of a species can set in motion a cascade effect through nutrient pathways that can influence rate of carbon fluxes (e.g., decomposition or primary productivity) and associated faunal and microbial organisms. This has already been shown in monospecific dry forest stands under extreme drought conditions by Barberena Arias [43].
Barberena Arias [43] found that the air temperature and humidity condition under the canopy of individual dry forest species was particular to the species. The tree species also regulated the amount and concentration of elements accumulating on the forest floor. These two conditions, in turn, had an effect on the richness, species composition, and trophic condition of soil arthropods decomposing litter. The N/P ratio was inversely related to arthropod density. On a landscape with a patchwork of dissimilar novel and native forest communities, the “priming” chemical effect of the dominant species might create “islands” with diverse nutrient levels and proportions that, in turn, affect soil organisms and ecosystem processes. This is a hypothesis to be tested with future research.
In summary, the implications of novelty to the ecology and conservation of Puerto Rico’s dry forests include (1) increases in species dominance and density at the 1-ha scale; (2) restoring forest conditions on degraded sites; (3) facilitating the regeneration and growth of native species on degraded sites; (4) establishing litter patches with diverse concentrations of chemical elements; (5) increasing the presence of nitrogen-fixing species and the availability of nitrogen in forest stands; (6) increasing the concentration of some elements in leaf litter above those observed in native forest stands; (7) decreasing the C/N ratio below those observed in native dry forest stands; and (8) potentially accelerating the flux of mass and nutrients by accelerating litter decomposition or primary productivity rates.
Acknowledgments
This study was conducted in cooperation with the University of Puerto Rico. We benefited from the collaborative research and discussions with numerous students and colleagues. Gisel Reyes collaborated with the literature search. We thank Ernesto Medina for substantial comments on the manuscript.
Author Contributions
Both authors contributed equally to the text and interpretation of results with Lugo focusing on the literature review and Erickson on the statistical analyses.
Conflicts of Interest
Authors declare no conflict of interests.
References
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San José, Costa Rica, 1967. [Google Scholar]
- Murphy, P.G.; Lugo, A.E. Ecology of tropical dry forest. Annu. Rev. Ecol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Murphy, P.G.; Lugo, A.E. Dry forests of Central America and the Caribbean. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 9–34. [Google Scholar]
- Maass, J.M. Conversion of tropical dry forest to pasture and agriculture. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 399–422. [Google Scholar]
- Ramjohn, I.A.; Murphy, P.G.; Burton, T.M.; Lugo, A.E. Survival and rebound of Antillean dry forests: Role of forest fragments. Forest Ecol. Manag. 2012, 284, 124–132. [Google Scholar] [CrossRef]
- Rudel, T.K.; Lugo, M.P.; Zichal, H. When fields revert to forest: Development and spontaneous reforestation in post-war Puerto Rico. Prof. Geogr. 2000, 52, 386–397. [Google Scholar] [CrossRef]
- Rudel, T.K. Tropical Forests: Regional Paths of Destruction and Regeneration in the Late Twentieth Century; Columbia University Press: New York, NY, USA, 2005. [Google Scholar]
- Lugo, A.E.; Helmer, E. Emerging forests on abandoned land: Puerto Rico’s new forests. For. Ecol. Manag. 2004, 190, 145–161. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Arico, S.; Aronson, J.; Baron, J.S.; Bridgewater, P.; Cramer, V.A.; Epstein, P.R.; Ewel, J.J.; Klink, C.A.; Lugo, A.E.; et al. Novel ecosystems: Theoretical and management aspects of the new ecological world order. Glob. Ecol. Biogeogr. 2006, 15, 1–7. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.S.; Hall, C.M. (Eds.) Novel Ecosystems: Intervening in the New Ecological World Order; Willey-Blackwell: West Sussex, UK, 2013. [Google Scholar]
- Pearce, F. The New Wild: Why Invasive Species Will Be Nature's Salvation; Beacon Press: Boston, MA, USA, 2015. [Google Scholar]
- Martinuzzi, S.; Lugo, A.E.; Brandeis, T.J.; Helmer, E.H. Geographic distribution and level of novelty of Puerto Rican forests. In Novel Ecosystems: Intervening in the New Ecological World Order; Hobbs, R.J., Higgs, E.S., Hall, C., Eds.; Wiley: Oxford, UK, 2013; pp. 81–87. [Google Scholar]
- Erickson, H.E.; Helmer, E.H.; Brandeis, T.J.; Lugo, A.E. Controls of fallen leaf chemistry and forest floor element masses in native and novel forests across a tropical island. Ecosphere 2014, 5, 48. [Google Scholar] [CrossRef]
- Brandeis, T.J.; Helmer, E.; Vega, H.M.; Lugo, A.E. Climate shapes the novel plant communities that form after deforestation in Puerto Rico and the U.S. Virgin Islands. For. Ecol. Manag. 2009, 258, 1704–1718. [Google Scholar] [CrossRef]
- Whittaker, R.H. Dominance and diversity in land plant communities: Numerical relations of species express the importance of competition in community function and evolution. Science 1965, 147, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.H. Communities and Ecosystems; The Macmillan Company: Toronto, ON, Canada, 1970. [Google Scholar]
- Brown, S.; Lugo, A.E. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 1982, 14, 161–187. [Google Scholar] [CrossRef]
- Martínez, Y.A. Biomass distribution and primary productivity of tropical dry forests. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 326–345. [Google Scholar]
- Gentry, A.H. Diversity and floristic composition of neotropical dry forests. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 146–194. [Google Scholar]
- Acevedo-Rodríguez, P.; Strong, M.T. Floristic richness and affinities in the West Indies. Bot. Rev. 2008, 74, 5–36. [Google Scholar] [CrossRef]
- Kiehn, M. Invasive alien species and islands. In The Biology of Island Floras; Bramwell, D., Caujapé-Castells, J., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 365–384. [Google Scholar]
- Francis, J.K.; Liogier, H.A. Naturalized Exotic Tree Species in Puerto Rico; General Technical Report SO-82; USDA, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1991. [Google Scholar]
- Molina Colón, S.; Lugo, A.E.; Ramos, O. Novel dry forests in southwestern Puerto Rico. For. Ecol. Manag. 2011, 262, 170–177. [Google Scholar] [CrossRef]
- Molina-Colón, S.; Lugo, A.E. Recovery of a subtropical dry forest after abandonment of different land uses. Biotropica 2006, 38, 354–364. [Google Scholar] [CrossRef]
- Molina Colón, S. Long-Term Recovery of a Caribbean Dry Forest after Abandonment of Different Land Uses in Guánica, Puerto Rico. Dissertation, University of Puerto Rico, Río Piedras, Puerto Rico, 1998. [Google Scholar]
- Lugo, A.E.; Brandeis, T.A. New mix of alien and native species coexist in Puerto Rico’s landscapes. In Biotic Interactions in the Tropics. Their Role in the Maintenance of Species Diversity; Burslem, D., Pinard, M., Hartley, S., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 484–509. [Google Scholar]
- Lugo, A.E. Los bosques. In Biodiversidad de Puerto Rico. Vertebrados Terrestres y Ecosistemas; Joglar, R.L., Ed.; Editorial del Instituto de Cultura Puertorriqueña: San Juan, PR, USA, 2005; pp. 395–548. [Google Scholar]
- Jardim, F.C.S.; Hosokawa, R.T. Estrutura da floresta equa- torial úmida da Estação Experimental de Silvicultura Tropical do INPA. Acta Amazon. 1986, 16–17, 411–508. [Google Scholar]
- Beard, J.S. The Natural Vegetation of the Windward and Leeward Islands. Oxford Forestry Memoirs 21; The Clarendon Press: Oxford, UK, 1949. [Google Scholar]
- Whittaker, R.J. Island Biogeography: Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Lugo, A.E. Visible and invisible effects of hurricanes on forest ecosystems: An international review. Austral Ecol. 2008, 33, 368–398. [Google Scholar] [CrossRef]
- Lugo, A.E. Dominancia y diversidad de plantas en Isla de Mona. Acta Cient. 1991, 5, 65–71. [Google Scholar]
- Kennaway, T.; Helmer, E.H. The forest types and ages cleared for land development in Puerto Rico. GISci. Remote Sens. 2007, 44, 356–382. [Google Scholar] [CrossRef]
- Brandeis, T.J.; Turner, J.A. Puerto Rico’s Forests, 2009; USDA Forest Service Resource Bulletin SRS-191; Southern Research Station: Ashville, NC, USA, 2013. [Google Scholar]
- Hulshof, C.M.; Yrízar, A.M.; Burques, A.; Boyle, B.; Enquist, B.J. Plant functional trait variation in tropical dry forests: A review and synthesis. In Tropical Dry Forests in the Americas: Ecology, Conservation, and Management; Sánchez-Azofeifa, A., Powers, J.S., Fernandes, G.W., Quesada, M., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2014; pp. 129–140. [Google Scholar]
- Lopezaraiza-Mikel, M.; Quesada, M.; Álvarez-Añorve, M.; Ávila-Cabadilla, L.; Martén-Rodriguez, S.; Calvo-Alvarado, J.; do-Santo, M.M.; Fernandes, G.W.; Sánchez-Azofeifa, A.; de J. Aguilar-Aguilar, M.; et al. Phenological patterns of tropical dry forests along latitudinal and successional gradients in the Neotropics. In Tropical Dry Forests in the Americas; Sánchez-Azofeifa, A., Powers, J.S., Fernandes, G.W., Quesada, M., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2014; pp. 101–128. [Google Scholar]
- Abelleira Martínez, O.; Lugo, A.E. Post sugarcane succession in moist alluvial sites in Puerto Rico. In Post-Agricultural Succession in the Neotropics; Myster, R.W., Ed.; Springer: New York, NY, USA, 2008; pp. 73–92. [Google Scholar]
- Jaramillo, V.J.; Sanford, R.L. Nutrient cycling in tropical deciduous forests. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 346–361. [Google Scholar]
- Lugo, A.E. Can we manage tropical landscapes? An answer from the Caribbean perspective. Landsc. Ecol. 2002, 17, 601–615. [Google Scholar] [CrossRef]
- Parrotta, J.A. Leucaena leucocephala (Lam.) de Wit. Leucaena; SO-ITF-SM-52; International Institute of Tropical Forestry, USDA Forest Service: Río Piedras, Puerto Rico, 1992. [Google Scholar]
- Santiago García, R.J.; Colón, S.M.; Sollins, P.; van Bloem, S.J. The role of nurse trees in mitigating fire effects on tropical dry forest restoration: A case study. Ambio 2008, 37, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Berg, B.; McClaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin, Germany, 2003. [Google Scholar]
- Barberena-Arias, M.F. Single Tree Species Effects on Temperature, Nutrients and Arthropod Diversity in Litter and Humus in the Guánica Dry Forest. Ph.D. Thesis, University of Puerto Rico, Río Piedras, Puerto Rico, 2008. [Google Scholar]
Figure 1.
Moisture Availability Index and aboveground biomass of Puerto Rican forests [14]. Vertical and horizontal bars are standard error of the mean reported by Brandeis et al. [14].

Figure 2.
Mean (±standard error) element concentration anomalies in fallen leaves of dry forest assemblages of Puerto Rico. Units are in percent for carbon, nitrogen, and sulfur and mg/g for all others. Elements include carbon (a), nitrogen (b), sulfur (c), phosphorus (d), calcium (e), magnesium (f), potassium (g), sodium (h), manganese (i), aluminum (j) and iron (k). Codes for species assemblages and the number of replicates in parenthesis are (novel assemblages in bold and shaded bars): CitFru (3)—Citharexylum fruticosum, BurSim (9)—Bursera simaruba, CasSyl (14)—Casearia sylvestris, LeuLeu (10)—Leucaena leucocephala, and AcaFar (5)—Acacia farnesiana. Anomalies are based on a mean of all plots within 14 island-wide species assemblages. Anomalies with the same letter in (a) indicate forest assemblages that are not significantly different from each other. Means and standard errors in (d), (g), (h), (i), and (k) were back-transformed from ln-transformed data.
Figure 2.
Mean (±standard error) element concentration anomalies in fallen leaves of dry forest assemblages of Puerto Rico. Units are in percent for carbon, nitrogen, and sulfur and mg/g for all others. Elements include carbon (a), nitrogen (b), sulfur (c), phosphorus (d), calcium (e), magnesium (f), potassium (g), sodium (h), manganese (i), aluminum (j) and iron (k). Codes for species assemblages and the number of replicates in parenthesis are (novel assemblages in bold and shaded bars): CitFru (3)—Citharexylum fruticosum, BurSim (9)—Bursera simaruba, CasSyl (14)—Casearia sylvestris, LeuLeu (10)—Leucaena leucocephala, and AcaFar (5)—Acacia farnesiana. Anomalies are based on a mean of all plots within 14 island-wide species assemblages. Anomalies with the same letter in (a) indicate forest assemblages that are not significantly different from each other. Means and standard errors in (d), (g), (h), (i), and (k) were back-transformed from ln-transformed data.
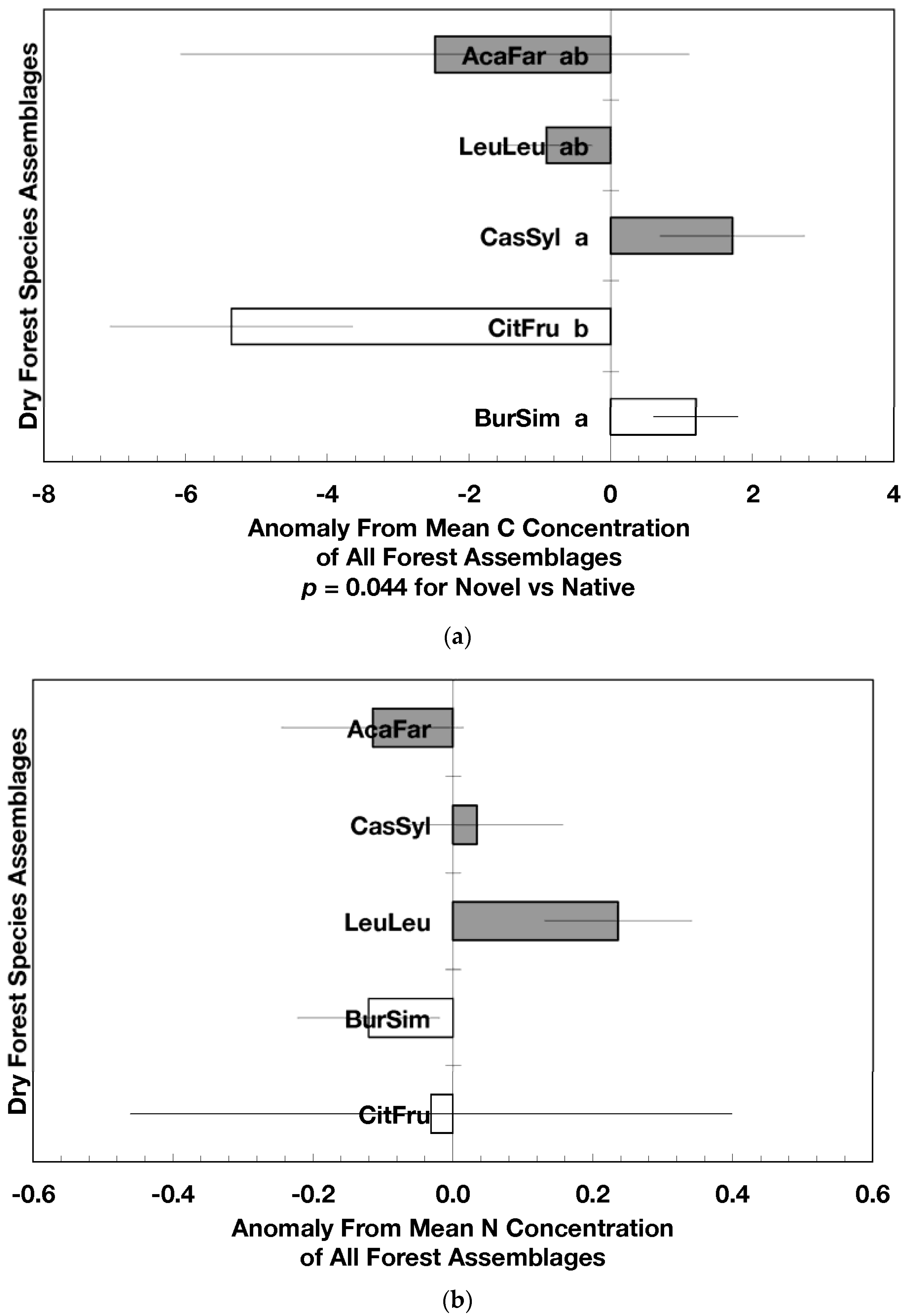
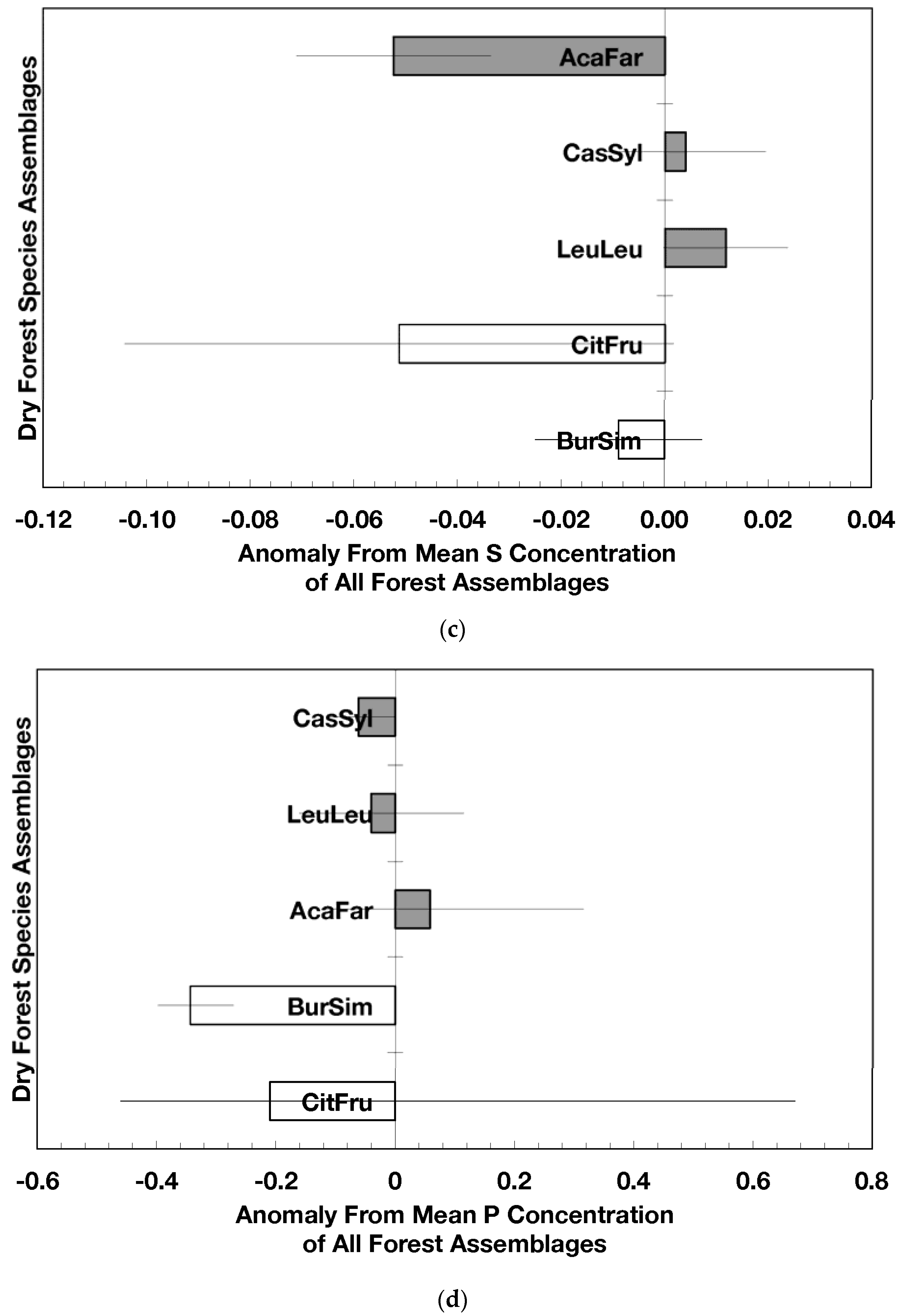

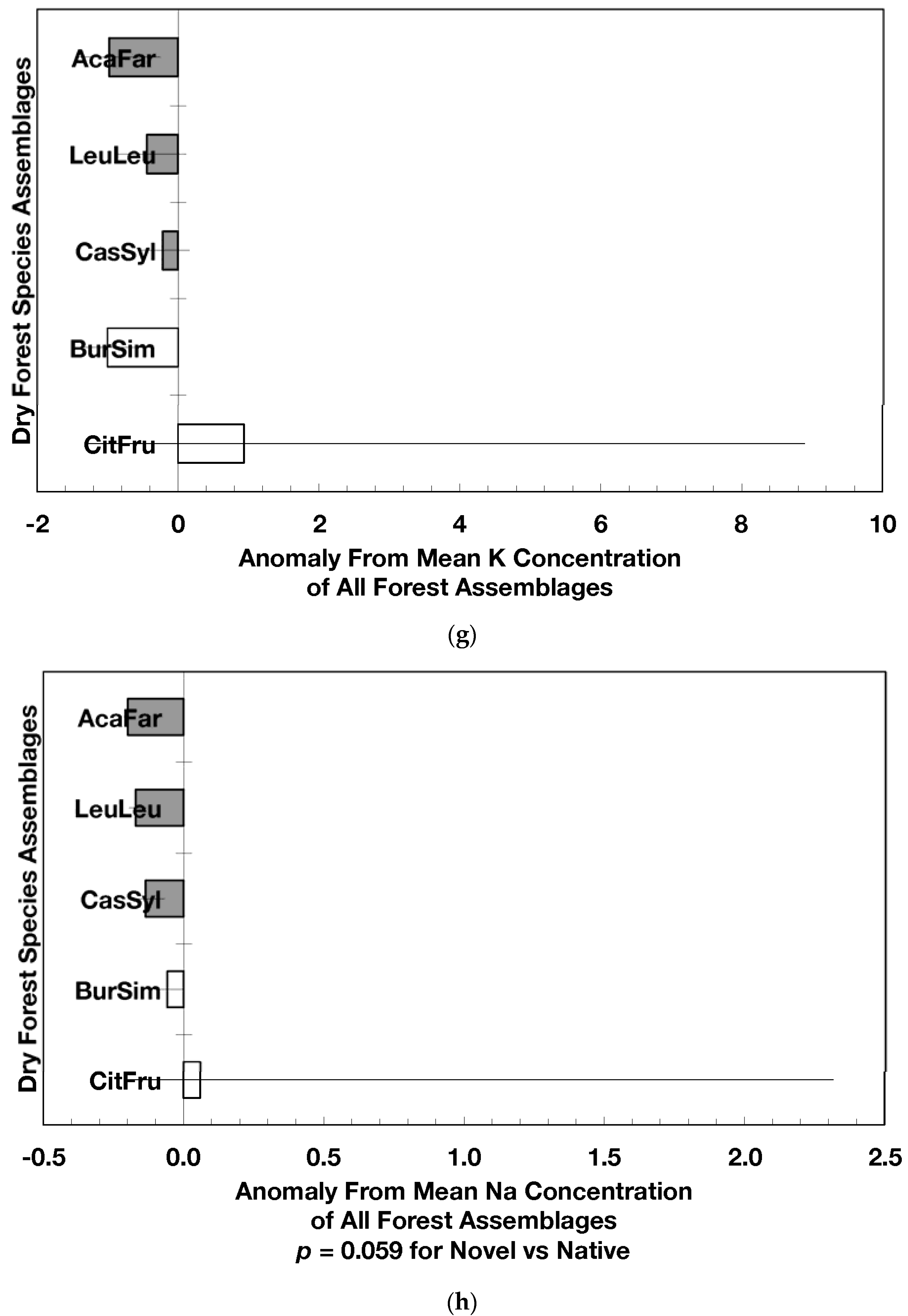
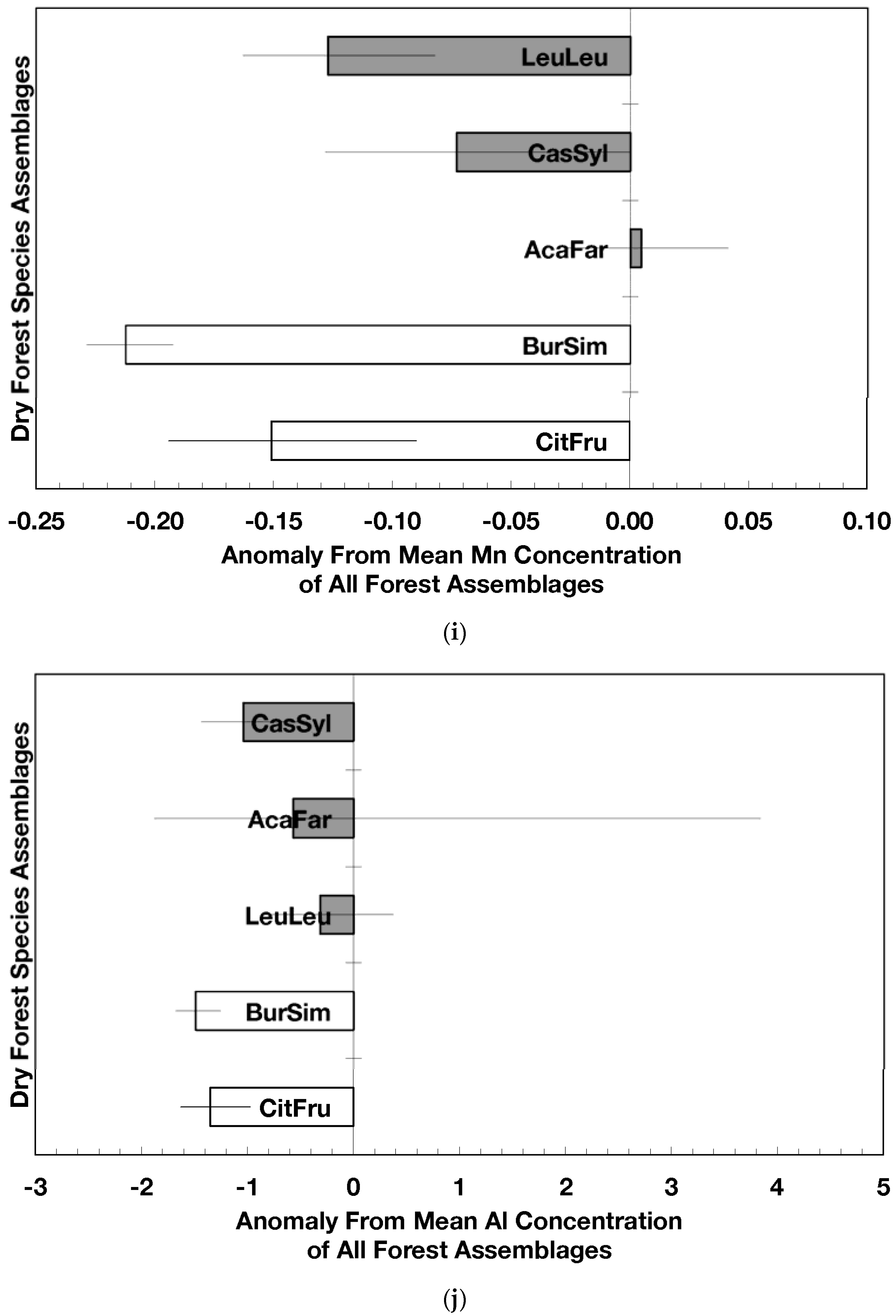
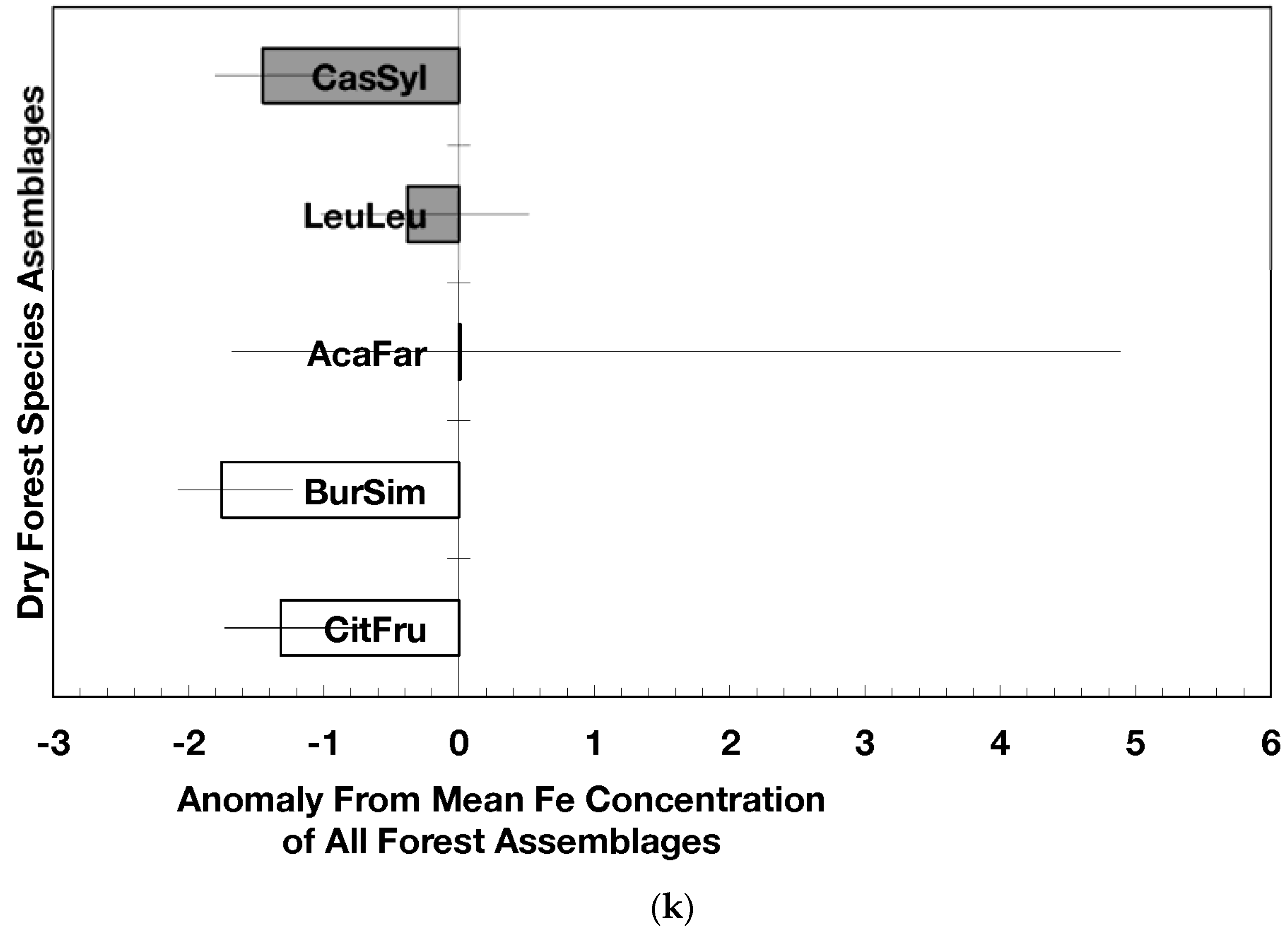
Figure 3.
Anomalies for element ratios in dry forests assemblages of Puerto Rico: carbon to nitrogen (a), carbon to phosphorus (b), and nitrogen to phosphorus (c). Values are the mean (±standard error). Codes for species assemblages and the number of replicates in parenthesis are (novel assemblages in bold and shaded bars): CitFru (3)—Citharexylum fruticosum, BurSim (9)—Bursera simaruba, CasSyl (15)—Casearia sylvestris, LeuLeu (10)—Leucaena leucocephala, and AcaFar (5)—Acacia farnesiana. Anomalies are based on a mean of all plots within 14 island-wide species assemblages. Means and standard errors in (a) and (b) were back-transformed from ln-transformed data.
Figure 3.
Anomalies for element ratios in dry forests assemblages of Puerto Rico: carbon to nitrogen (a), carbon to phosphorus (b), and nitrogen to phosphorus (c). Values are the mean (±standard error). Codes for species assemblages and the number of replicates in parenthesis are (novel assemblages in bold and shaded bars): CitFru (3)—Citharexylum fruticosum, BurSim (9)—Bursera simaruba, CasSyl (15)—Casearia sylvestris, LeuLeu (10)—Leucaena leucocephala, and AcaFar (5)—Acacia farnesiana. Anomalies are based on a mean of all plots within 14 island-wide species assemblages. Means and standard errors in (a) and (b) were back-transformed from ln-transformed data.
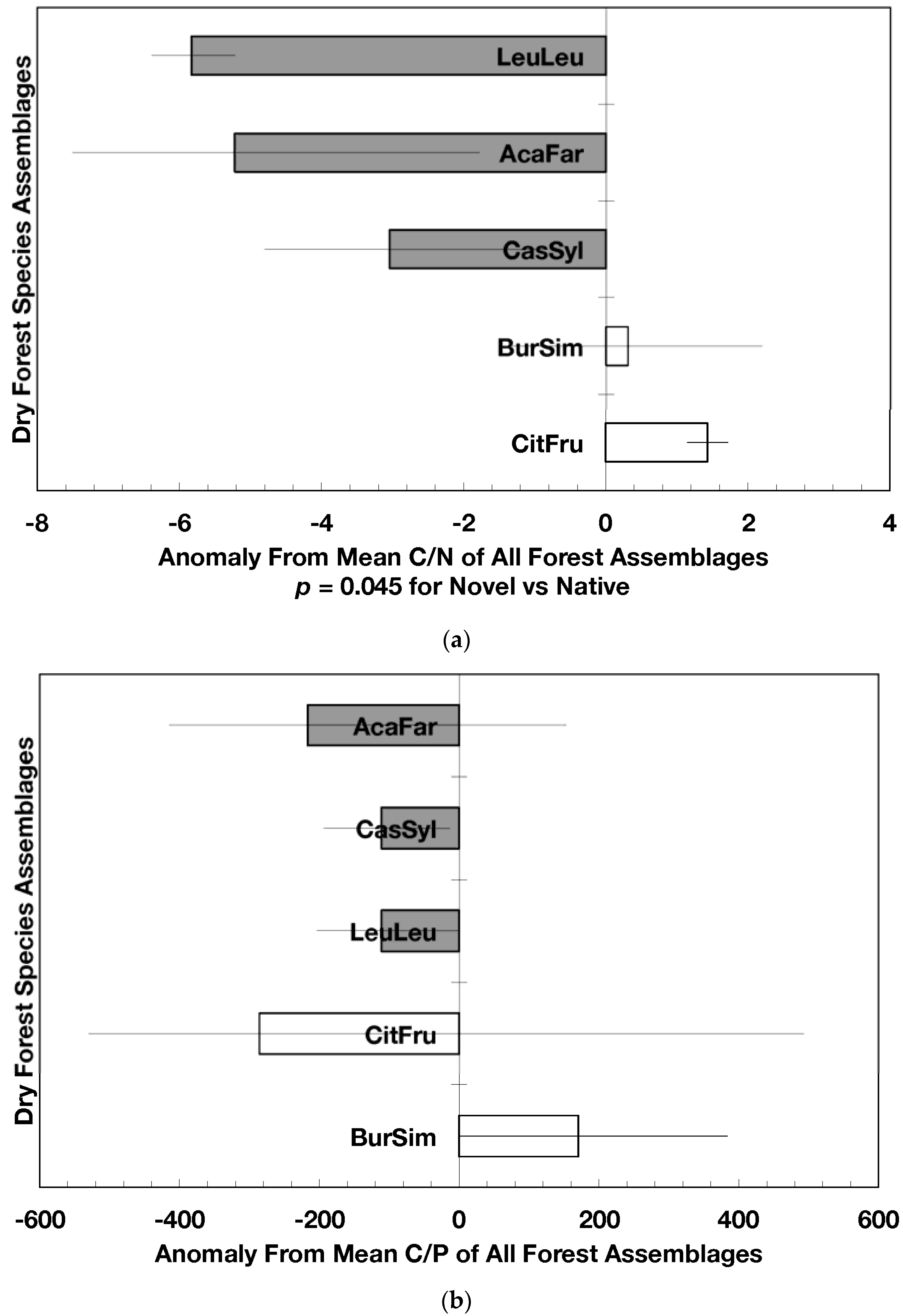
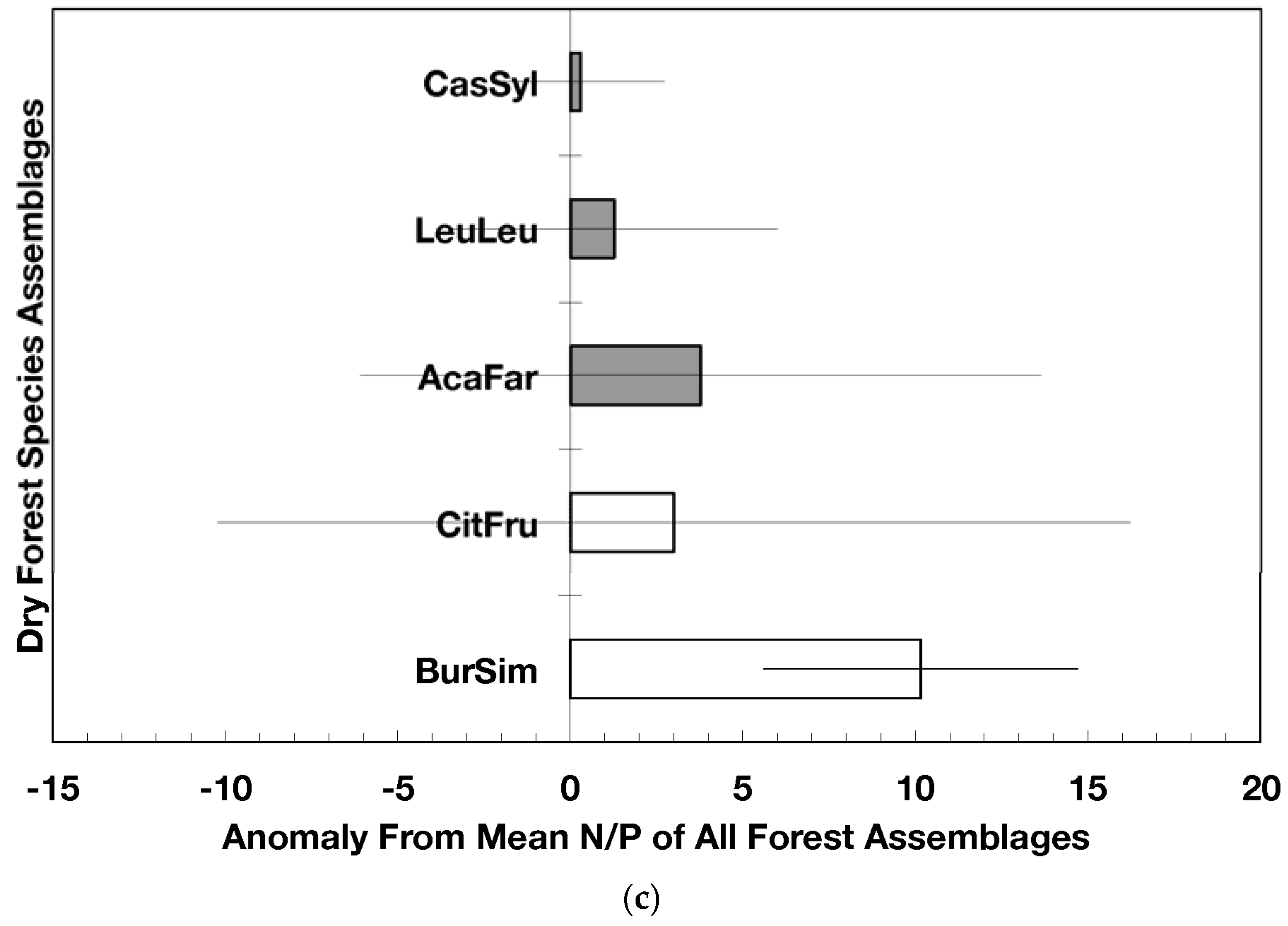
Figure 4.
Percent of stand basal area of nitrogen-fixing species in dry forest assemblages of Puerto Rico [13]. Codes for species assemblages are (shaded bars correspond to novel assemblages identified in bold below): CitFru—Citharexylum fruticosum, BurSim—Bursera simaruba, CasSyl—Casearia sylvestris, LeuLeu—Leucaena leucocephala, and AcaFar—Acacia farnesiana. Clear bars are native dry forests and the solid bar represents forests of the subtropical dry forest life zone.
Figure 4.
Percent of stand basal area of nitrogen-fixing species in dry forest assemblages of Puerto Rico [13]. Codes for species assemblages are (shaded bars correspond to novel assemblages identified in bold below): CitFru—Citharexylum fruticosum, BurSim—Bursera simaruba, CasSyl—Casearia sylvestris, LeuLeu—Leucaena leucocephala, and AcaFar—Acacia farnesiana. Clear bars are native dry forests and the solid bar represents forests of the subtropical dry forest life zone.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Average structural parameters of dry forests in Puerto Rico and the US Virgin Islands [14]. Averages are based on nine novel and eight native species assemblages of similar age. The p value denotes the level of significance in a t-test comparison of averages.
Table 1.
Average structural parameters of dry forests in Puerto Rico and the US Virgin Islands [14]. Averages are based on nine novel and eight native species assemblages of similar age. The p value denotes the level of significance in a t-test comparison of averages.
| Forest Status | Stem Density (stems/ha) | Basal Area (m2/ha) | Aboveground Biomass (Mg/ha) | Tree Height (m) |
|---|---|---|---|---|
| Novel | 3103 | 9.71 | 36.43 | 5.83 |
| Native | 4348 | 16.4 | 70.33 | 7.3 |
| p value | 0.012 | 0.011 | 0.0002 | 0.002 |
Table 2.
Mass (g/m2) of forest floor litter (Total), fallen leaf mass, and chemical element mass (g/m2) in total litter for dry forest species assemblages (those in bold are novel assemblages), and subtropical dry forest life zone forests. Other codes for species assemblages are: CitFru—Citharexylum fruticosum, BurSim—Bursera simaruba, CasSyl—Casearia sylvestris, LeuLeu—Leucaena leucocephala, AcaFar—Acacia farnesiana. Data are from Erickson et al. [13].
Table 2.
Mass (g/m2) of forest floor litter (Total), fallen leaf mass, and chemical element mass (g/m2) in total litter for dry forest species assemblages (those in bold are novel assemblages), and subtropical dry forest life zone forests. Other codes for species assemblages are: CitFru—Citharexylum fruticosum, BurSim—Bursera simaruba, CasSyl—Casearia sylvestris, LeuLeu—Leucaena leucocephala, AcaFar—Acacia farnesiana. Data are from Erickson et al. [13].
| Species Assemblage and Replicates | Total Mass | Leaf Mass | C | N | S | P | Ca | Mg | K | Na | Mn | Al | Fe |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CitFru (3) | 285 | 94 | 131 | 4.2 | 1.2 | 0.18 | 7.9 | 0.79 | 0.55 | 0.07 | 0.04 | 0.49 | 0.65 |
| BurSim (9) | 666 | 434 | 333 | 10.1 | 1.9 | 0.26 | 19.2 | 1.39 | 0.9 | 0.23 | 0.09 | 0.87 | 0.63 |
| CasSyl (14) | 465 | 234 | 224 | 7.5 | 1.4 | 0.29 | 8.1 | 1.29 | 1.06 | 0.09 | 0.10 | 0.98 | 0.83 |
| LeuLeu (10) | 691 | 225 | 325 | 11.7 | 1.9 | 0.44 | 16.7 | 2.05 | 1.33 | 0.08 | 0.11 | 0.69 | 0.46 |
| AcaFar (5) | 325 | 89 | 154 | 4.8 | 1.2 | 0.15 | 7.6 | 0.66 | 0.44 | 0.03 | 0.03 | 0.68 | 0.70 |
| Life Zone (12) | 543 | 145 | 253 | 6.7 | 1.4 | 0.21 | 14.1 | 1.22 | 0.69 | 0.04 | 0.05 | 0.76 | 0.82 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lugo, A.E.; Erickson, H.E. Novelty and Its Ecological Implications to Dry Forest Functioning and Conservation. Forests 2017, 8, 161. https://doi.org/10.3390/f8050161
AMA Style
Lugo AE, Erickson HE. Novelty and Its Ecological Implications to Dry Forest Functioning and Conservation. Forests. 2017; 8(5):161. https://doi.org/10.3390/f8050161
Chicago/Turabian StyleLugo, Ariel E., and Heather E. Erickson. 2017. "Novelty and Its Ecological Implications to Dry Forest Functioning and Conservation" Forests 8, no. 5: 161. https://doi.org/10.3390/f8050161
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.