
Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance


1
School of Geography and Development, University of Arizona, Tucson, AZ 85721, USA
2
School of Natural Resources and the Environment, University of Arizona, Tucson, AZ 85721, USA
*
Author to whom correspondence should be addressed.
Forests 2017, 8(7), 221; https://doi.org/10.3390/f8070221
Submission received: 19 April 2017
/
Revised: 13 June 2017
/
Accepted: 19 June 2017
/
Published: 22 June 2017
(This article belongs to the Special Issue Post-Disturbance Forest Management and Regeneration Dynamics)
Abstract
:Climate change is increasing the frequency and extent of high-severity disturbance, with potential to alter vegetation community composition and structure in environments sensitive to tipping points between alternative states. Shrub species display a range of characteristics that promote resistance and resilience to disturbance, and which yield differential post-disturbance outcomes. We investigated differences in shrub patch size and stem density in response to variations in fire severity, vegetation community, and post-disturbance reproductive strategies in Sky Island forested ecosystems in the southwestern United States. Patterns in shrub structure reflect the effects of fire severity as well as differences among species with alternate post-fire reproductive strategies. Increased fire severity correlates with larger patch sizes and greater stem densities; these patterns are observed across multiple fire events, indicating that disturbance legacies can persist for decades. High severity fire produces the largest shrub patches, and variance in shrub patch size increases with severity. High severity fire is likely to promote expansion of shrub species on the landscape, with implications for future community structure. Resprouting species have the greatest variability in patch structure, while seeding species show a strong response to disturbance: resprouting species dominate at low disturbance severities, and obligate seeders dominate high severity areas. Differential post-fire reproductive strategies are likely to generate distinct patterns of vegetation distribution following disturbance, with implications for community composition at various scales. Shrub species demonstrate flexible responses to wildfire disturbance severity that are reflected in shrub patch dynamics at small and intermediate scales.
1. Introduction
The resilience and resistance of vegetation to disturbance events is a primary influence on ecological succession. Post-disturbance vegetation response and recovery reflects uncertainties in community composition, structure, and recovery trajectories [1,2,3]. Wildland fire is an especially complicated disturbance agent because spatial and temporal variability in fire severity can compound one another to produce highly variable outcomes [3,4]. In addition, species-specific responses to disturbance influence the composition and structure of post-fire vegetation communities. In forests, shrubs are particularly vulnerable to pyrogenic effects, including direct combustion of canopy and supporting stems and convective heat damage to cambial tissue and roots, because of their low stature growth forms [2,5]. Perhaps because of their short stature and vulnerability to fire effects, shrub species have evolved a suite of life history strategies for recovery following disturbance, including germinating from seed, sprouting from surviving tissues, and combinations of seeding and sprouting [5,6,7,8]. These adaptations allow shrubs often to dominate early post-fire succession, sometimes persisting in situ for multiple decades after fire [9,10,11].
1.1. Ecological Resilience in Shrub Systems
Ecological resilience can be defined as the capacity of ecosystems to withstand disturbance without altering important structures and processes [12,13]. Shrub species have evolved a range of traits that promote resistance and resilience to disturbance, and which yield differential post-disturbance outcomes, including plasticity in growth form, post-disturbance regeneration strategies, and physiological characteristics that minimize damage from disturbance [1,6,7]. Shrub stands exemplify fire-resilient communities because of high fire tolerance and rapid post-disturbance recovery of many of the constituent species [10,14,15]. Shrub communities tend to be dynamically stable in the face of pyrogenic disturbance, until fire cessation or biological invasion alters community composition or structure. At this point, the reapplication of disturbance can cause a restructuring of species composition or growth forms that modifies future disturbance regimes.
The dynamics and stability of shrublands and forests are of particular interest to managers, ecologists, foresters and range specialists, and economic users of landscapes, especially in systems that are sensitive to disturbance-mediated tipping points between alternate structural types or stable states [13,16,17]. The composition, density, and structure of vegetation communities following disturbance can prove highly resilient in the face of later changes, and may resist reverting to pre-disturbance conditions because of altered ecosystem dynamics [18].
The capacity for fire resistance seen in many of the taller shrub species in southwestern North America (most markedly in Quercus) allows some shrubs to take single-stemmed, arboreal form, thereby lifting the canopy above the direct influence of flames and convective heat from low-intensity fire [2]. Because many species can take either tree or shrub form following injury by fire, the structure of shrub patches (defined here as the relationship between canopy area and stem density) has important implications for forest structure after wildfire disturbance. The plastic response of shrub species to disturbance has implications for understanding potential “tipping points” in the structural trajectory of an ecosystem after wildfire disturbance [17,19]. Ecological disturbance, especially when severity is high or disturbance is unusually frequent, can promote changes in species composition and vegetation structure [6,17]. Importantly, even when species composition is unaffected, disturbance can ‘tip’ ecological communities into alternate structural states, such as shrubby growth form versus arboreal form [9]. Given a sufficiently long fire-free period, single-stemmed shrub patches, most of which germinate from seed, are more likely to grow into monopodial trees, which can withstand repeated low- and moderate-intensity wildfire without converting to low-profile shrub form [20,21]. In contrast, multi-stemmed shrub patches tend to persist in the face of disturbance by resprouting as low-profile shrub patches [2,5]. Both of these outcomes in shrub species reflect varying degrees of resilience and resistance to disturbance. Consequently, shrub species are able to resist pyrogenic change by altering their structure to accommodate different fire regimes: low severity fire allows some shrub species to grow above the flame height and survive even frequent application of low-intensity fire, while some shrub species subjected to high-severity fire can either resprout or initiate from seeds following canopy mortality. Thus, the plasticity of growth form represents a form of resilience to disturbance, because some shrub species can recover from high-severity disturbance by changing structure into multiple-stemmed shrub patches.
1.2. Climate and Fire Severity in the North American Monsoon Region
In contrast to research on fire and climate effects of shrub species growing in Mediterranean ecosystems, the literature on shrub dynamics in forested ecosystems dominated by the North American Monsoon (NAM) is relatively sparse. Unlike Mediterranean ecosystems, which receive the bulk of precipitation in the winter, this NAM-dominated region experiences two wet seasons, with >50% of precipitation falling between July and September and most of the remaining precipitation falling between December and March [22]. The increase in humidity and precipitation during the summer influences fire extent and severity throughout the fire season (during the fore-monsoon drought versus during the monsoon), as well as post-fire recovery, regrowth, and germination. Whereas Mediterranean ecosystems tend to experience the greatest fire severities and burned-area extent in autumn, before winter rains commence [23,24], NAM-dominated ecosystems have the highest fire severities and greatest area burned in the fore-monsoon drought of May through early July, with many smaller fires ignited during the monsoon period [25,26].
Considerable literature on shrub dynamics in the NAM region has focused on shrub encroachment into grasslands (e.g., [27,28,29,30]), and on impacts of shrub evapotranspiration on desert hydrology (e.g., [31,32]). Less research has focused on the dynamics of shrub species in montane forest ecosystems, particularly as those shrub species are affected by wildfire disturbance. Generally, research into pyrogenic influences on shrub species in the NAM region evaluates the differential responses of conifers versus angiosperm species to fire severity and post-fire regeneration, with a focus on pre- and post-fire frequency and dominance, rather than investigating shrub-specific responses [9,11,33]. In the face of low-intensity fire, many shrub species exhibit resistance to fire injury [33,34], but at higher fire severities these species may persist following fire either by resprouting or by germinating from soil seed banks or from inseeding [35]. Shrub life history strategies reflect differential methods of gaining advantage after disturbance, including resprouting, germinating from seed, and combinations of the two strategies (facultative seeding; [1]).
The structure and composition of montane forests and woodlands has important implications for future fire severity, post-fire recovery trajectories, and associated resource management issues that include watershed function, habitat for species of special concern, and recreational amenities. Here, we describe the architectural responses of shrub species to fire injury across fire severities, with implications for potential tipping-point responses between forests and shrubfields in montane forest ecosystems. To that end, we investigate the dynamics of shrub structure (patch size and stem density) in response to variations in fire severity, legacies of prior disturbance, vegetation community, and post-disturbance reproductive strategies in Madrean Sky Island forested ecosystems, to address shrub species’ resistance and resilience to pyrogenic disturbance. Finally, we ask whether prior fire severity may influence shrub structure in the decades following disturbance.
2. Materials and Methods
2.1. Study Area
This study was conducted in the Chiricahua Mountains of southeastern Arizona, USA (Figure 1). The Chiricahuas are the largest and most biodiverse of Arizona’s montane Sky Islands, characterized by strong floristic affinity with Madrean archipelago forests that extend into the northern Mexican states of Sonora and Chihuahua, as well as across southern Arizona and the Mogollon Rim [36,37,38]. Climate in the Chiricahuas is fairly moderate for the latitude, with 14.1 °C mean annual temperature at 1660 m elevation, and mean winter temperature of 5.7 °C [39]. Mean annual precipitation at 1660 m is 442 mm, with the bulk of precipitation falling in the summer (56%) and winter (27%). Spring precipitation provides 8% of the annual total, and autumn contributes 9% [39]. The Chiricahuas span an elevational range of 1220–2975 m, and support a wide range of habitats along this gradient, with slope aspect providing a strong influence on moisture availability, and hence the vegetation type. Semi-desert grasslands and upland scrub ecosystems are found at the lowest elevations (1220–2000 m), and are replaced by pinyon-juniper-oak woodlands at elevations from 1450–2600 m. Lower pine-oak forest and woodland is common between 1700–2560 m, and upper conifer-oak forest and woodlands are dominant between 2070–2740 m, with aspect strongly influencing the elevations at which oaks or conifers are dominant. At the highest elevations, dry-mesic mixed conifer is the dominant vegetation type between 2280–2740 m, and the range is topped by aspen-mixed conifer forest between 2590–2795 m. South-facing slopes generally carry more xeric vegetation at any given elevation compared to shady, north-facing slopes [40,41]. Fires are common in forested vegetation types in the Chiricahuas, with pine-oak forests showing historical evidence of frequent fire (mean fire interval 4–6 years) [42]. High-elevation mixed conifer forests also experienced frequent fires (mean fire interval 10–16 years), with little evidence of stand-replacing fire [43]. The Chiricahuas have a long history of frequent, low-intensity fire, with tree-ring based fire histories from as early as the 1400s [44] demonstrating an historical dynamic interplay of climatic and anthropogenic influences on fire frequency [42,44,45]. Decades of fire suppression have produced structural changes to forests and fuels in the Chiricahuas that have led to a recent increase in fire severity and longer fire intervals [9,33,42,46].
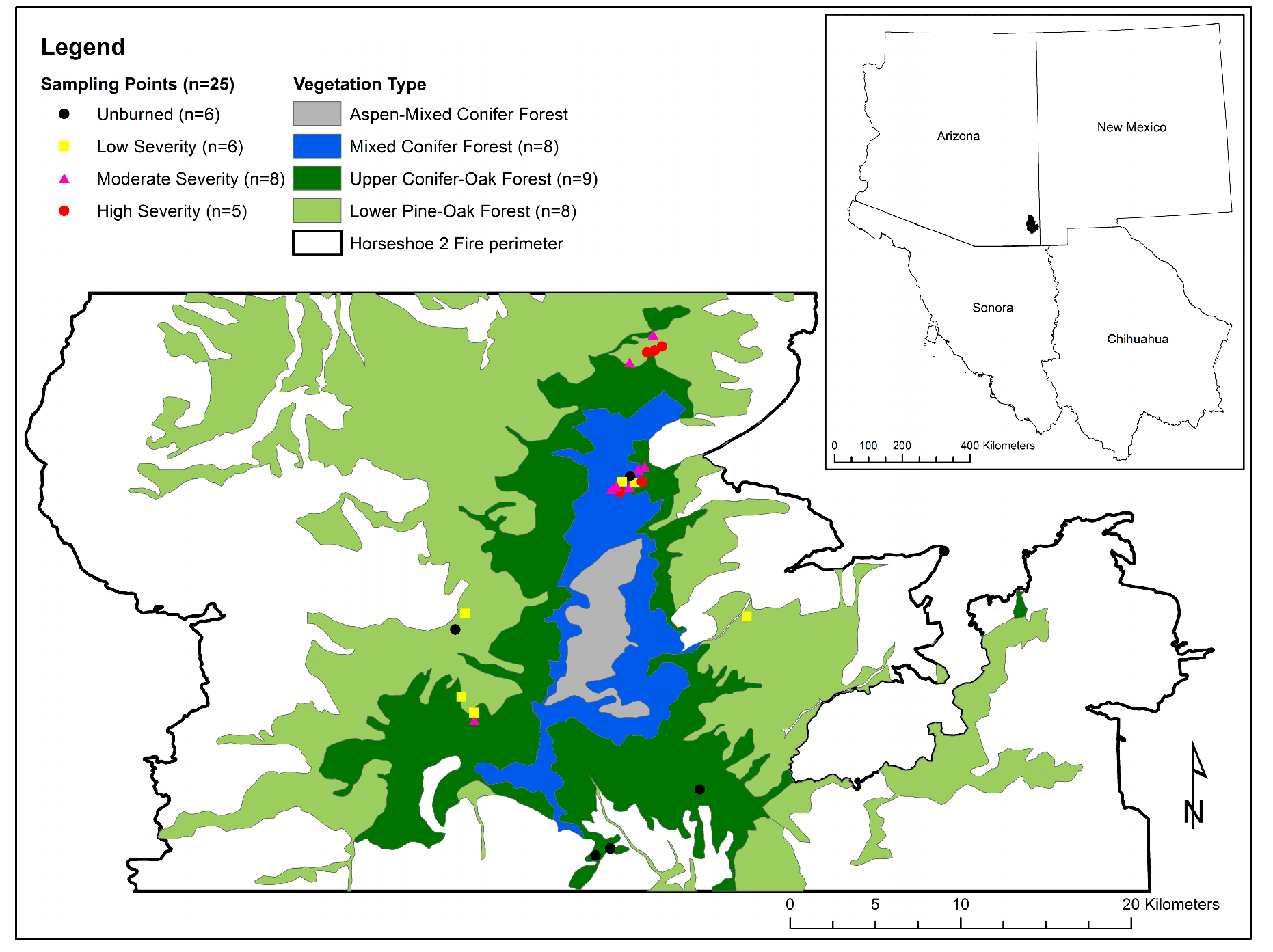
Twenty-five sampling plots were positioned in the Coronado National Forest (Figure 1) within the perimeter of the 2011 Horseshoe 2 Fire, which burned nearly 91,000 ha between 8 May and 25 June during the hot and dry fore-monsoon drought period. Almost 42% of the area affected by the Horseshoe 2 Fire burned at moderate and high severity [47]. These plots had been unburned since at least the 1994 Rattlesnake Fire, which burned 10,210 ha between 28 June–29 July. Unlike the Horseshoe 2 Fire, which consumed its fuels and was controlled before seasonal initiation of the NAM, the Rattlesnake Fire ignited during the fore-monsoon drought and burned into the more humid monsoon period, with 44% of the area burned at moderate and high severity [44]. Sixteen study plots were located within the perimeter of both fires, allowing for analysis of the effects of two mixed-severity fires on forest and shrub patch structure. Sampling locations were selected to represent a range of elevational settings, topographic variables, vegetation types, and Horseshoe 2 and Rattlesnake Fire soil burn severities. Soil burn severity was mapped using the Differenced Normalized Burn Ratio (dNBR), which is a refinement of Normalized Burn Ratio (NBR). NBR is an index of vegetation health that assesses healthy vegetation (high reflectance in the near-infrared (NIR) spectrum) in comparison to burned areas (high reflectance in the shortwave infrared (SWIR) spectrum): NBR = (NIR − SWIR)/(NIR + SWIR). To assess fire effects on vegetation, dNBR takes pre-fire and post-fire images and compares NBR values, and provides a good comparison of post-fire vegetation compared to pre-fire vegetation: dNBR = NBRpre-fire − NBRpost-fire [47].
We situated the 25 sampling locations in three ecological systems: Madrean Lower Pine-Oak Forest and Woodland; Madrean Upper Conifer-Oak Forest and Woodland; and Southern Rocky Mountain Dry-Mesic Montane Mixed Conifer Forest and Woodland [40,41]. These three ecological systems roughly correspond to the U.S. government’s LANDFIRE Existing Vegetation Type (EVT) classifications of Madrean Lower Pine-Oak Forest and Woodland and Southern Rocky Mountain Dry-Mesic Montane Mixed Conifer Forest and Woodland, respectively. The principle distinction between “ecological systems” and LANDFIRE EVTs is that Madrean Upper Conifer-Oak Forest and Woodland consists of >50% Douglas-Fir and large pines, but otherwise contains the same species assemblages as Madrean Lower Pine-Oak Forest and Woodland, albeit in different relative abundances [48]. For purposes of simplicity, the three vegetation types are referred to here as “Lower Pine-Oak”, “Upper Pine-Oak”, and “Mixed Conifer” (Figure 1).
2.2. Shrub Measurements
Shrub patch measurements were conducted in 25 1000 m2 (17.85 m radius) circular plots. Within each plot, the canopy area of every individual shrub and shrub patch (n = 867) was measured by taking the major and minor axis of each shrub patch to the nearest 0.1 m. Ellipses representing shrub patch area were calculated from these two axes. Within every shrub patch, the number of living stems per patch was counted. Using these two metrics of shrub structure (area and stem counts), we calculate stem density by dividing the number of stems in a patch by the canopy area. Shrub patches were measured to the edge of the plot in cases where the patch extended beyond plot boundaries, and shrubs were identified to species. In addition to species presented in this paper, measurements were taken of cactus, agave (Agave spp.), yucca (Yucca spp.), and beargrass (Nolina) species, as well as grass, fern, and Rubus species. Rubus species were not analyzed in this paper because their thicket-like growth form makes accurate stem counts difficult, and roseate shrubs such as agave, yucca, and beargrass are almost always single-stemmed. Data for shrub species not analyzed in this paper are presented elsewhere [46].
2.3. Shrub Classification
Shrub species have developed several distinct life history strategies for regenerating and propagating following disturbance by wildfire, including germinating from seed, sprouting from root collars or rhizomes, or combinations of these two strategies [7,8]. For purposes of analysis, we grouped the species identified in this study by post-fire regeneration strategy (Table A1 in Appendix A), obtained from the Fire Effects Information System [15]; species citations provided in Appendix B. Obligate seeders are species that must germinate from seeds after pyrogenic disturbance, whereas obligate resprouters are species that characteristically regenerate post-fire from existing stump or root collars or from root rhizomes, generally because their seeds are highly sensitive to damage from fire [49,50,51]. Facultative seeders can germinate from seed or resprout from preexisting stumps, roots, or rhizomes following disturbance by fire [1]. We excluded facultative seeding species from our analysis because an insufficient number were sampled for this study.
We additionally tested whether particular post-fire regeneration strategies are associated with predominance in patches of single stem individuals. We hypothesized that obligate seeding species would be observed more commonly as single-stemmed patches, because these shrubs require germination from seeds after fire disturbance. By contrast, obligate resprouters have the capability to send up multiple stems from root collars and rhizomes, which would produce shrub patches with continuous canopy supported by multiple stems.
2.4. Statistical Testing and Regression Analysis
The effects of outlier observations were evaluated using influence tests, and observations with a Cook’s D over 1.0 were discarded [52]. We used pairwise Student’s t-tests to evaluate significance (α = 0.05) of differences in shrub structure observed across variations in fire severity, vegetation type, and post-fire regeneration strategy. Because the raw shrub patch data were not normally distributed, data were log transformed to achieve normality and thus to conform to the assumptions of Student’s t-test. We used linear and nonlinear regression of shrub patch data to model the relationships between shrub patch area and the number of live stems supporting that canopy. Regressions were performed for individual species and combinations of species across independent variables including fire severity and vegetation type. Species combinations include the shrub species that co-occurred on plots, as well as species grouped by post-disturbance regeneration strategies. Regression residuals were checked for normalcy, and no trend was observed in the residuals. Z statistics were calculated to test significant relationships between proportional abundances, and coefficients of variation of shrub patch size and stem density were calculated to assess shrub variability in relation to the mean.
3. Results
3.1. Fire Severity Influences Shrub Patch Size and Stem Density
Antecedent fire severity plays a significant role in structuring shrub patch canopy area at moderate and high severities and in determining shrub patch stem density, especially in unburned, low-, and moderate-severity burn areas of the study sites (Table 1). While average shrub patch size remains roughly constant across fire severity classes, shrub canopies in high-severity burned areas are nearly twice as large on average (Table 1). Shrub species respond to increasing fire severity from unburned to moderate-severity fire with steady decreases in the mean and median patch sizes and with increases in the maximum patch size (Table 1).
Increasing fire severity also influences shrub stem density, steadily reducing the mean and maximum stem density per patch, with significant differences observed in all but low-severity burned areas (Table 1). We also find a reduction in the variation (C.V.) in stem density among plots with increasing fire intensity. Pairwise comparisons of fire severities (unburned vs. low, unburned vs. moderate, unburned vs. high, etc.) demonstrate that the canopy area of shrubs growing in unburned areas do not differ from shrubs in low-severity plots, but that canopy area differs across all the other comparisons (Table 2). Similarly, stem density does not differ between unburned and low-severity plots, or between unburned and high-severity plots, but all other relationships are significantly different (Table 2).
3.2. Shrub Variability across Vegetation Types
The distribution and dominance of shrub species varies by vegetation type, which transitions from Lower Pine-Oak to Upper Pine-Oak and Mixed conifer with increases in elevation. The three vegetation types contain roughly the same number of shrub species, and a number of species are present across the elevational gradient represented by these three vegetation types (Table 3). Several species tend to cluster in lower or higher elevational positions; the greatest number and overlap in species is found in the Upper Pine-Oak vegetation type, which occupies an intermediate elevational position between Lower Pine-Oak and Mixed Conifer forests. Pairwise testing of significance shows shrub patch structure differing significantly across vegetation types. Stem density per patch is significantly different across all vegetation types (p = 0.001 for Lower Pine-Oak vs. Upper Pine-Oak; p < 0.001 for Mixed Conifer vs. both Pine-Oak types), and patch area differs significantly between mixed conifer and the two pine-oak vegetation types (p < 0.001).
Lower Pine-Oak shares 80% of its shrub species with Upper Pine Oak, and the Mixed Conifer vegetation type shares 70% of its species with Upper Pine-Oak, while only 38% of the species are common to all three vegetation types. The overall dominance of shrub species declines with elevation (Table 3).
3.3. Post-Disturbance Regeneration Strategies Affect Shrub Patch Dynamics
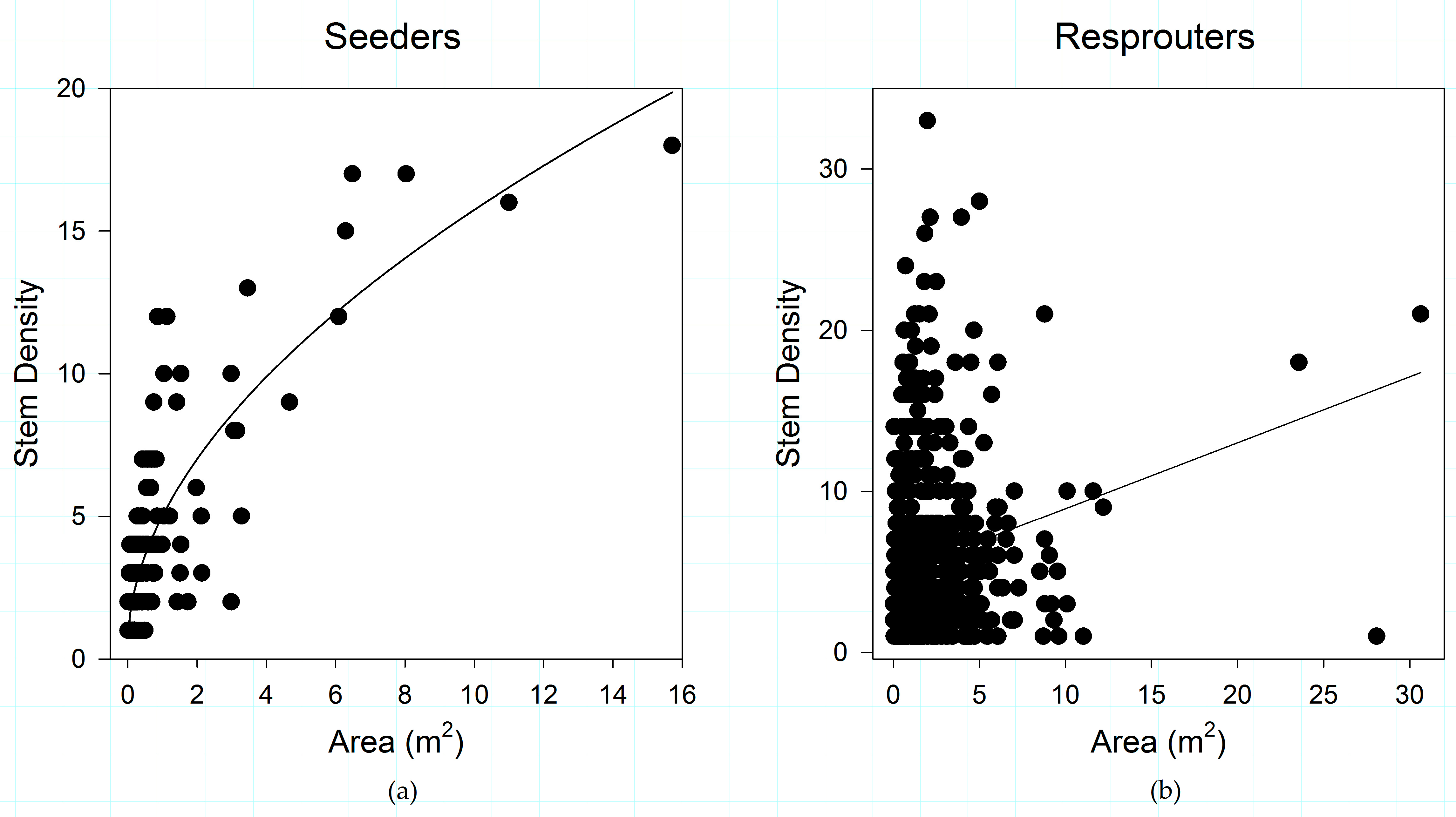
Post-fire regeneration strategy appears to play a role in structuring the relationships between shrub canopy and the number of stems supporting that canopy when all vegetation types and fire severities are combined (Figure 2). Obligate seeding species have a much stronger relationship (R2 = 0.704) between shrub patch area and number of stems than do obligate resprouters (R2 = 0.04). Differences observed in patch area and stem density between these post-fire regeneration strategies is significant (p < 0.001).
When shrubs are analyzed for dissimilarities in patch structure across variations in fire severity and regeneration strategy, a different pattern emerges. Obligate resprouters show only a weak relationship of shrub structure across variations in fire severity, with the strongest relationship emerging in oak species growing in unburned areas (R2 = 0.256; slope coefficient = 0.322; data not shown). When obligate sprouting species are analyzed across sprout source (root crown versus rhizome), no pattern in post-fire shrub patch structure emerges. This suggests that the sprout location (root versus rhizome) of post-fire shrub sprouts is independent of fire severity, and that fire effects do not promote one sprouting strategy after pyrogenic disturbance.
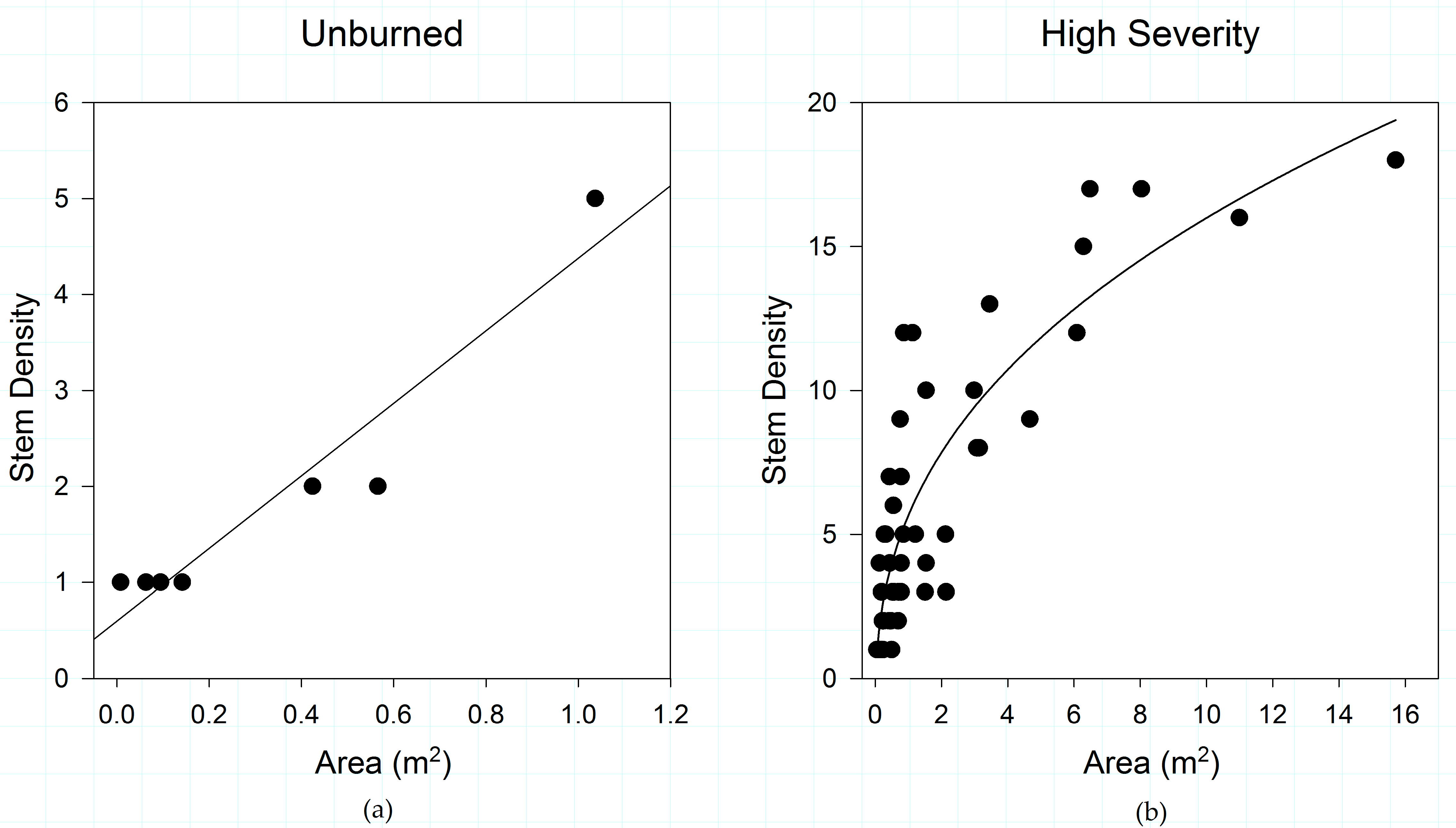
By contrast, obligate seeders show a variable response to shrub structure across fire severity classes. Among obligate seeders, the strongest relationships between shrub structure and fire severity are observed in unburned (R2 = 0.922; slope coefficient = 0.244) and high-severity areas (R2 = 0.754) (Figure 3), with weaker relationships in moderate (R2 = 0.398; slope coefficient = 0.135; data not shown) and low fire severity (R2 = 0.141; slope coefficient = 0.045; data not shown).
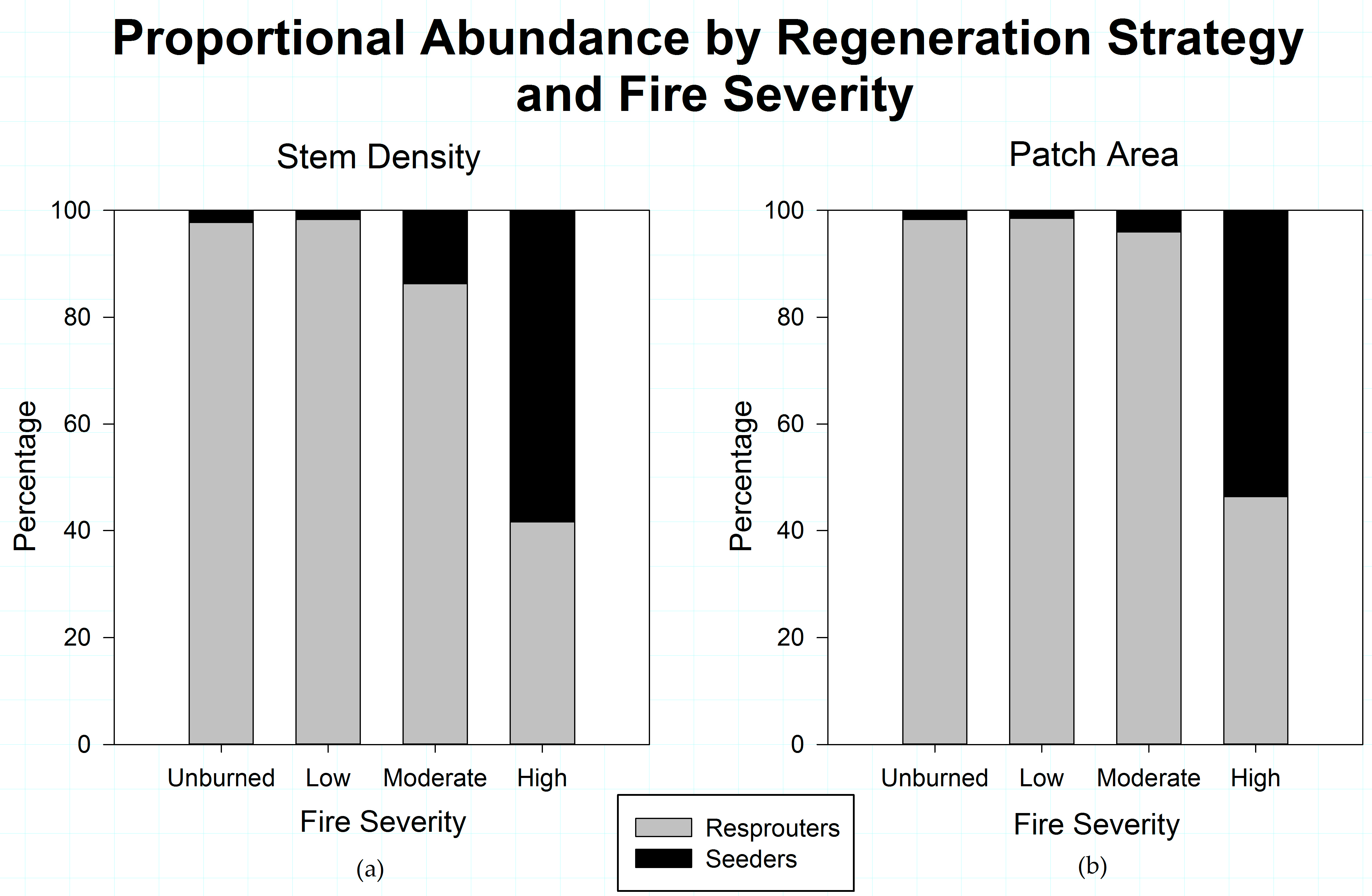
When regeneration strategy is evaluated across fire severities (Figure 4), obligate resprouters are proportionally much more common at lower fire severities. At high severities, obligate seeders are proportionally more abundant, whether measured by stem density or canopy area. The dominance of obligate post-fire seeders in high-severity burn areas is significant.
When regeneration strategy is evaluated across vegetation types, we see a strong tendency for obligate resprouters to dominate Pine-Oak ecosystems, and for obligate seeders to dominate in Mixed Conifer (Table 3 and Table 4). Lower and Upper Pine-Oak communities are significantly weighted towards obligate resprouters. In contrast, Mixed Conifer plots show significant dominance by obligate seeders (63.9%), with obligate resprouters representing 36.1%.
Of the shrub patches measured, 16% were composed of single-stemmed individuals. Of these, 61.0% of the single-stemmed shrub patches encountered were resprouters, and 38.3% were obligate seeders (data not shown). Only 15.2% of multi-stemmed shrub patches were comprised of obligate seeders, whereas 80.1% of multiple-stemmed shrub patches were composed of resprouters. Single-stemmed and multi-stemmed patches differ significantly (p < 0.001) from one another in patch size and stem density, and the variance in multi-stemmed patches (7.60) is greater than in single-stemmed patches (0.96).
3.4. Vegetation Type Does Not Predict Variability in Shrub Structure
Shrub species occur in differing abundances across vegetation types based on their post-fire regeneration strategy. Vegetation types, as a broad set of plant associations and a proxy for elevational position, do not display any characteristic patterns in terms of shrub structure, although significant differences are observed between the three vegetation types in stem density, and between mixed conifer and the pine-oak vegetation types for patch area. When all shrub species are combined within a vegetation type, relationships between shrub patch cover and stem density were weak in Lower Pine-Oak (R2 = 0.397, slope coefficient = 0.294, p < 0.001) and Upper Pine-Oak (R2 = 0.218, slope coefficient = 0.230, p < 0.001) plots, but stronger in Mixed Conifer plots (R2 = 0.576, slope coefficient = 0.948, p < 0.001) (data not shown).
3.5. Fire Severity Influences Shrub Structure
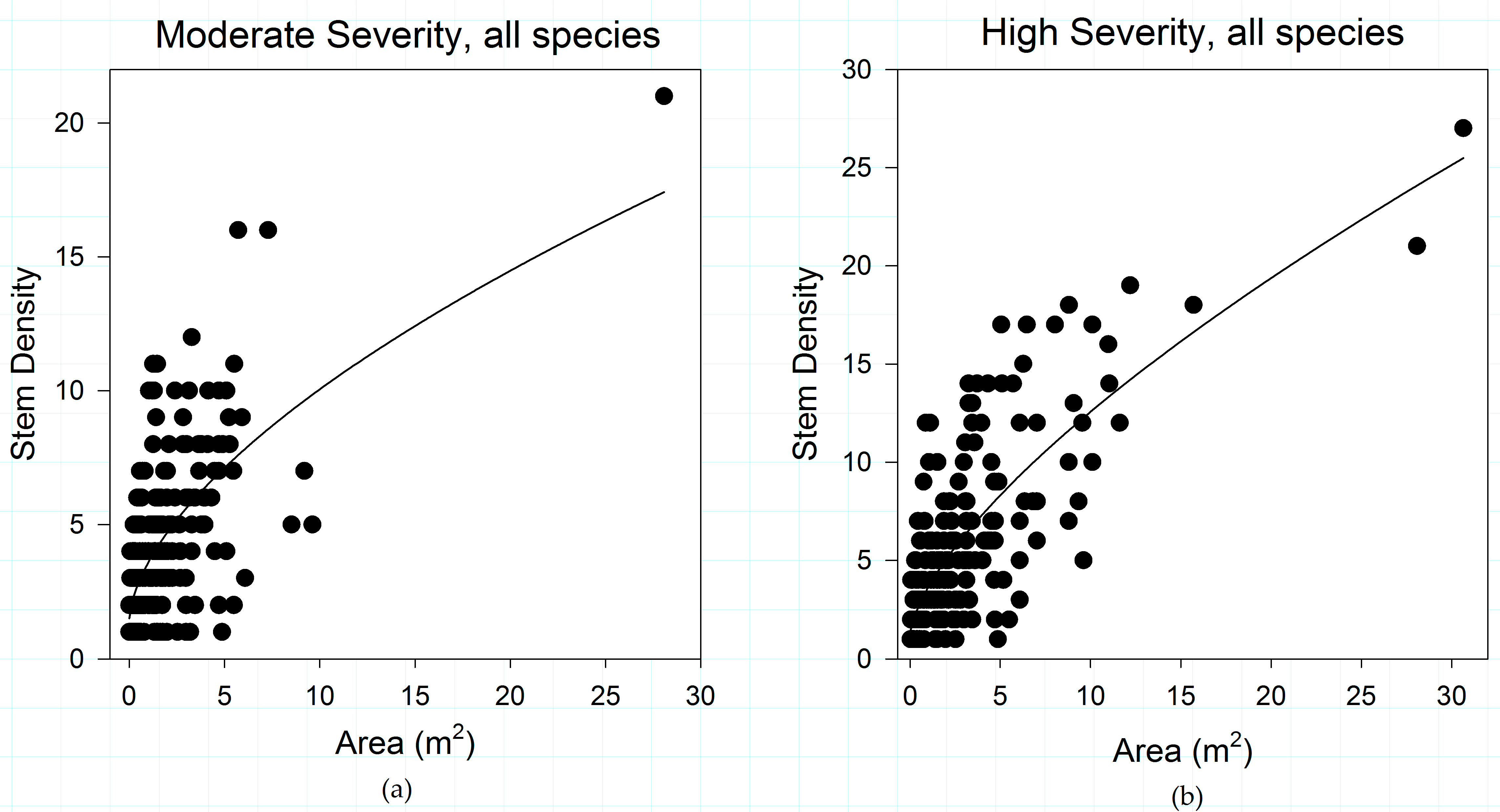
When the entire shrub community is considered, the relationship between stem density and patch area becomes stronger with increasing fire severity in successive fires (Figure 5). Unburned areas have the weakest relationship (R2 = 0.257, slope coefficient = 0.101) between stem density and area, with low severity areas (R2 = 0.334, slope coefficient = 0.229) reflecting weak relationships between patch area and stem density (data not shown). Higher fire severities strengthen the relationship between patch area and stem density, increasing in moderate-severity areas (R2 = 0.588), and strongest in high-severity areas (R2 = 0.662) (Figure 5).
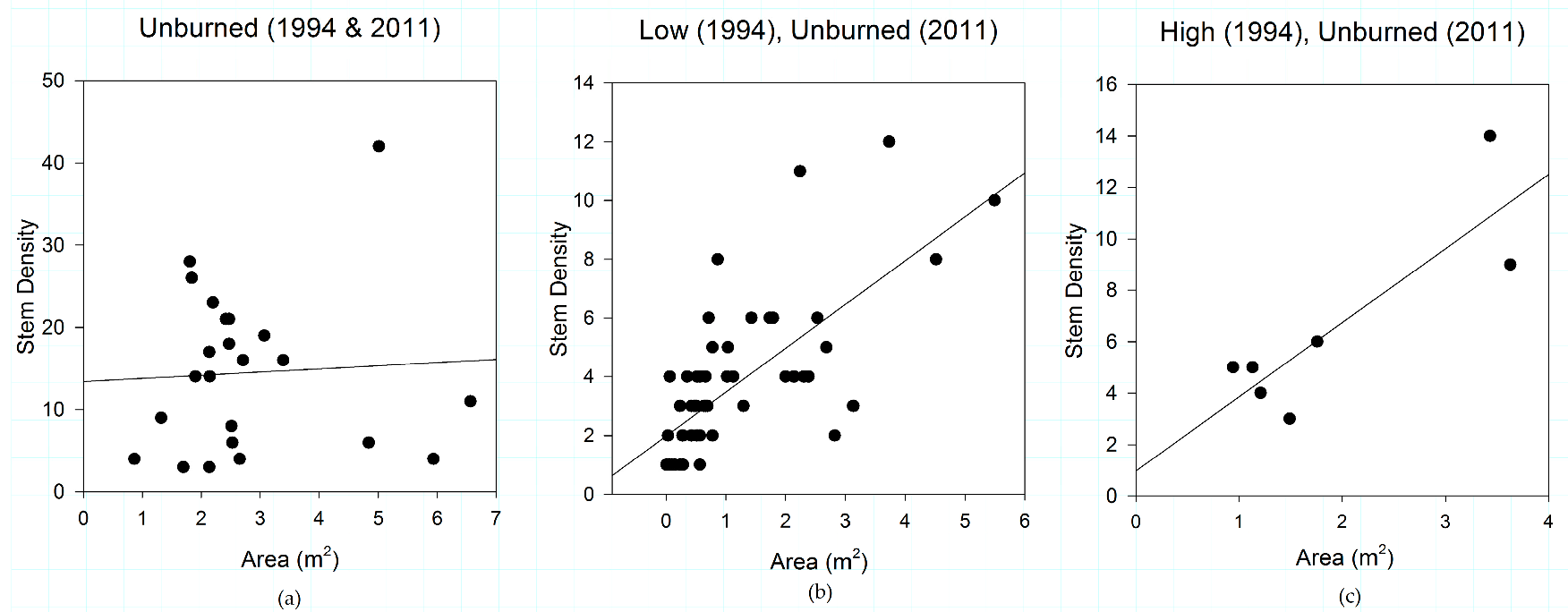
The organizing effects of antecedent fire severity on shrub patch dynamics are also stable through time. When plots that were unburned in 2011 are analyzed by prior (1994) fire severity, the same pattern illustrated in Figure 5 emerges. In plots unburned in 2011, 1994 fire severity plays a role in structuring shrub structure (Figure 6), with shrubs growing in areas that did not experience fire in 1994 and 2011 having a very weak pattern in terms of shrub structure (R2 = 0.003, slope coefficient = 0.008). The relationship between canopy area and stem density strengthens in plots that were burned at low severity (R2 = 0.503, slope coefficient = 0.337) and high severity (R2 = 0.727, slope coefficient = 0.252) in 1994 but went unburned in 2011 (Figure 6). An insufficient number of 1994 moderate-severity plots were sampled for this analysis.
4. Discussion
4.1. Fire Effects on Shrub Patch Structure
In Madrean Sky Island ecosystems, disturbance by wildfire produces predictable patterns in shrub stand structure in terms of canopy size and the relationship of canopy area to the stem density across fire severity classes. The organizing effect of fire severity can persist for decades, which suggests that the legacy effects of disturbance can endure long enough for subsequent disturbance to occur, thus providing a strong influence on post-disturbance recovery patterns. This is an important factor in post-disturbance recovery, because spatial distribution, species composition, and structural features of pre-disturbance vegetation influence post-disturbance recovery trajectories and influence community-level resilience and resilience to disturbance. Equally important, pre-disturbance vegetation patterns can have strong effects on subsequent disturbance severity. Recursive relationships between shrub responses to disturbance followed by vegetation influences on later disturbance severity provide important insights into important and management-relevant ecological dynamics in forested environments [5,9,53].
Increases in patch size variance and maximum patch size (see Table 1) with fire severity are likely attributable to the effect of increasing fire severity in removing and reducing overstory canopy, which opens space for enhanced germination and sprouting of shrub species, as well as for individual shrubs to merge and intersect into larger patches. This is probably the result of pyrogenic changes to overstory tree canopy, which is unchanged in unburned areas, removed entirely in high-severity areas, and affected moderately in low and moderate fire severity areas. Low-severity fire alters only understory vegetation components, with minimal overstory and canopy effects, while moderate-severity fire can remove overstory canopy and have large effects on understory composition. In low and some moderate fire severity areas, shrub regeneration may occur from both resprouting and seed germination, which might weaken the relationships observed in obligate reseeding species.
The variable outcomes of understory and overstory vegetation in low and moderate fire severity areas suggests the potential for divergence of fire effects on vegetation independent of soil burn severity, followed by post-fire response of the vegetation itself. The differences observed in shrub patch structure in low-severity burned areas compared to moderate- and high-severity plots may also reflect a distinction between patches composed of single individual shrubs versus consolidated patches. As disturbance effects from fire become more pronounced, the shrubs regenerating become more regular and predictable in their growth form. This trend is evident across a fairly large range in terms of both shrub canopy area and stem density. The regulation of shrub canopy by fire severity is likely due to the increasingly strong effects of moderate- and high-severity fire on overstory tree canopy. In the absence of competition from overstory canopy, shrub patches develop in a more regular fashion, with less variance in shrub patch structure.
Lower fire severities promote smaller patches of shrubs that are carried by proportionally greater stem densities. Increases in patch area are not due to increased stem density, and the overall correlation between patch size and the stem density is relatively high (Pearson’s r = 0.639). High-severity fire represents such a dramatic ecosystem reset that the shrubs that sprout or germinate can grow large canopies carried by a smaller number of stems. It is possible that high-severity burned areas do not share the trend in declining shrub stem density because the lower soil burn severity classes have upper bounds, but high severity does not; in effect, the range of fire effects can be greater in high-severity burned areas than in lower severity patches. In this sense, moderate-severity fire may not have a unique effect on shrub stem densities, but rather represent only the overall trend in fire effects on shrub patches along a severity gradient.
4.2. Post-Fire Regeneration Strategy Affects Post-Disturbance Shrub Patch Dynamics
Important variations in shrub patch structure occur because of post-fire regeneration strategies of shrub species. Shrub patch dynamics also vary across fire severity classes, demonstrating that increasing fire severity has an organizing effect on the relationship between shrub patch canopy cover and stem density. Moreover, the organizing effects of fire severity on shrub patch dynamics can persist for decades, even with the ongoing processes of succession, competition, and species-specific responses to age and time. In the absence of fire for long periods of time, shrubs appear to be structured differently under closed or nearly-closed overstory canopy compared to shrubs growing in more recently burned areas. This result suggests that the observed pattern of increasing fire severity strengthening the relationship between the components of shrub structure, and explaining progressively increasing amounts of variance in shrub canopy-to-stem ratios, may be stable over longer periods of time.
Fire severity, vegetation type, and the behavior of individual species are insufficient to explain variance in shrub structure among obligate resprouters. Sprouting appears to be an effective regeneration strategy that yields high variability in post-fire shrub structure. Obligate resprouting species can regenerate from rhizomes or from the root crown, and some species are capable of regenerating from both root crowns and rhizomes [7,8], and even of spreading asexually through adventitious rooting of aerial stems [54]. Adventitious rooting produces new sites at which post-fire sprouting could occur, and although this phenomenon was not directly observed in this study, it could have effects on shrub patch structure [54]. When regeneration strategy is evaluated across fire severities (Figure 4), obligate resprouters are proportionally much more common following lower fire severity, probably because unburned and low-severity burned areas are characterized by intact overstory canopy, and low-severity fire does not totally kill the root and stump rhizomes of sprouting species. In part, the stronger relationship observed between stems and shrub canopy area arises from the fact that obligate seeders germinate from seeds, whereas obligate resprouters could send up a number of shoots from a small area of a root or stump collar. However, in cases where tightly clumped seeds all germinate into a single patch, variable canopy areas could be sustained by differential numbers of stems. Despite that likelihood, the relationship and structure of shrub cover appears markedly different across post-fire regeneration strategy, indicating considerable plasticity in post-disturbance response.
Obligate seeding species are proportionally most abundant in the highest-elevation vegetation type and in high-severity burned areas. In part, this likely reflects large canopy openings as well as destruction of below-ground plant tissues from which resprouts could emerge, in part because resprouting vigor may be greatest following moderate-severity fire [55]. In high-severity patches, shrub regeneration frequently relies on seed rain from unburned or less-severely-burned areas because of soil seedbank mortality [35,56,57] and destruction of rhizome and root crown tissue [2,56]. Some post-fire seeding species may have a competitive advantage over resprouting species following intense wildfire because of their long-lived, heat-resistant seeds [58] and the tendency of seeds to be fire stimulated [59]. Germination from seed can be prompted by several fire cues, including charred wood [60], smoke [57], ash [57], and heat [56,57], all of which can confer an advantage on seeding species if severe fire effects have killed or decreased the resprouting vigor of competing plants [8,55]. The post-fire seeding species contained in this study rely on durable, heat-resistant seeds [58,61] and heat scarification to promote germination [58,62]. In low and moderate burned areas, post-fire resprouting appears to be a more effective strategy than seeding, based on proportional abundances of resprouters compared to seeders (Figure 4). This is likely a result of the competitive advantage obtained from undamaged pre-fire biomass [2,5,8] over germination.
Vegetation type does not exert an influence on patch structure, but the proportion of shrub patches differs across vegetation types. The decline in shrub cover across an elevational gradient of vegetation types signifies the considerable importance of shrub cover at lower-elevation sites, and the relatively lower importance of shrub cover as a component of higher-elevation mixed conifer ecosystems. Unburned and low-severity burned areas are colonized by very small patches of obligate seeding species. These small patches are generally emergent from single seeds that are spatially disaggregated (or else they would have been measured as multi-stemmed patches) as is evident in Figure 3. The more dramatic disturbance associated with high-severity fire creates much larger patches carried by numerous stems.
4.3. Climate Influences on Fire Effects on Shrub Species
Shrub species growing in NAM-dominated ecosystems may demonstrate different responses to disturbance by fire compared to Mediterranean ecosystem shrubs because of seasonal differences in soil moisture and humidity. The seasonality of burning can play an important role in shrub recovery following fire, although most understory species are not significantly affected by season of fire [63,64]. Because soil and fuel moistures influence fire severity, late dry-season fires in Mediterranean climates have a disproportionate effect on shrub cover and species richness, particularly at small scales [64]. In contrast, late-season fires in NAM-dominated forests coincide with greater humidity, higher soil and fuel moistures, and increased precipitation. Thus, late-season fires in NAM-dominated ecosystems may promote lower disturbance severity compared to fires in Mediterranean climates, with implications for post-disturbance recovery of shrub species. The seasonality of burning can affect resprouting species particularly strongly, with burning early in the growing season causing high mortality because non-structural carbohydrates have not yet been assimilated into plant tissues [65,66]. Thus, the phenology of certain shrub species may produce increased vulnerability to wildfire occurring at different times of year.
The bimodal distribution of seasonal precipitation in NAM-dominated climates influences disturbance timing and severity, with the spatially most extensive and ecologically most severe fires burning in May and June, and with fire size and intensity moderating with the onset of the monsoon season [67]. For this reason, the highest fire intensities observed over large spatial scales are likely to be experienced in the middle of the growing season, with growth-promoting rainfall falling before the growing season ceases. By contrast, Mediterranean-climate shrubs are reliant on soil moisture from cool-season precipitation, meaning that shrub recovery may be delayed until conditions are favorable for germination and resprouting [68]. Shrub species growing in Mediterranean climates tend to recover quickly from early season fires, which occur when soil and fuel moistures are highest [64].
4.4. Tipping Points between Forest and Shrubfields
Elevated disturbance severity in Madrean forested ecosystems is predicted to promote conversion of forest ecosystems to woodlands or shrubfields [9,18]. Many Madrean shrub and tree species display traits that make them variously more resistant to damage by wildfire, or resilient after disturbance [1]; Appendix B, which might confer community-scale resilience and resistance to disturbance, especially low and moderate severity wildfire.
Increased disturbance severity is likely to have differential effects on trees and shrubs, particularly since only one Madrean pine (Chihuahuan pine, P. leiophylla Scheide & Deppe) is resilient to crown fire through post-fire resprouting [69,70]. By contrast, all the Madrean oaks and many shrub species are capable of resprouting, either from rhizomes or root collars [71,72]; Appendix B. Oaks growing in regions characterized by mixed fire regimes invest in belowground carbohydrate stores, which aid in post-fire resprouting, and may delay allocations in bark thickness, which would confer additional resistance to fire [73]. For this reason, high-severity disturbance that kills monopodial trees can promote conversion of forest to shrubland, in the process producing stable vegetation types that can weather repeated moderate- and high-severity disturbance through resprouting. In this sense, type conversion of pine-oak forest into oak shrublands could represent a change to more resilient vegetation that is suited to this region’s likely future climate and disturbance regimes.
Type conversion of forest to shrubland can reflect changes in dominant plant functional types (e.g., loss of conifers) but does not necessarily herald changes to the relative abundance of species, as when arboreal-form oak trees are top-killed and resprout as low-profile shrubs. Shrubby species, with their lower canopies, are more susceptible to canopy injury from less-intense burning than tree-form species with lofted canopies [73]. Low- and moderate-severity fire has disproportionate effects on small-diameter stems, which promotes resprouting in species that are capable of it and simultaneously promotes persistence of large-diameter stems. Thus, subsequent disturbance of shrubfields and the vigorous post-fire recovery of shrub species is likely to produce shrub patches that are resistant to change in either species composition or structure and resilient in the face of repeated disturbance [9,33]. In this sense, shrub species are both resistant and resilient in the face of disturbance, which suggests that areas that are dominated by shrub species may remain so, especially under conditions of repeated moderate- and high-severity wildfire. Shrublands may persist until disturbance ceases for a sufficient time for later-seral stage species such as pines and other conifers to regenerate, or for management actions to alter community composition and structure to favor alternate forest states. Management actions may be necessary to influence the balance between conifers and oaks, because tree seedlings grow more slowly than post-disturbance sprouts [33] and can be rapidly outcompeted. Once established, many Madrean conifers are resistant to low and moderate fire severity, but are not high-severity disturbance, and tend not to endure except under specific disturbance regimes. Lengthy disturbance cessation favors certain pine seedlings, as their growth rate accelerates following more than a decade of establishment [74]. On the other hand, biomass accumulation from long fire-free periods, coupled with the growth of ladder fuels, increases fire intensity and severity and likely favors oak dominance.
In pine-oak woodlands, which may be sensitive to tipping between conifer dominance or oak dominance with altered disturbance regimes [75], high frequency of low- to moderate-intensity fire promotes abundant tree seedling regeneration, but paradoxically produces low survivorship of recently-established seedlings [9,76,77]. In the Madrean Sky Islands, high frequency, low-severity fire interacts with resilient pine and pine-oak forests, but these forests may not be resistant to elevated disturbance severity. Moderate-intensity disturbance will tend to favor fire-resistant species, at least initially, by top-killing plants with lower canopies. Species that can resprout rapidly or regenerate from seed following disturbance by fire can gain biomass relatively quickly and become co-dominant in mixed pine-oak systems. Stem elongation growth rates of resprouting oaks can exceed 50 cm per year [33], which could allow oak shrub patches to become the dominant canopy post-fire.
5. Conclusions
Disturbance by wildfire produces predictable patterns in shrub structure in terms of canopy size and the relationship of canopy area to stem density across fire severity classes. The organizing effect of fire severity on forest structure and composition can persist for decades, suggesting that the legacy effects of disturbance may endure long enough for subsequent disturbance to occur. This is an important factor in post-disturbance ecological recovery, because spatial distribution, species composition, and structural features of pre-disturbance vegetation all influence post-disturbance recovery trajectories. Equally important, these post-disturbance vegetation patterns can have strong effects on subsequent disturbance severity. Recursive relationships between shrub responses to disturbance followed by vegetation influences on later disturbance severity provide important insights into important and management-relevant ecological dynamics in forested environments.
In the Madrean Sky Islands, shrub species respond to fire disturbance through changes to patch size, total canopy area, and shrub patch structure, all of which vary along a gradient in fire severity. The post-fire regeneration strategy of shrub species influences post-fire recovery and relative dominance among shrub communities. The persistence of established vegetation means that recovery of shrub species to mixed-severity fire creates legacies that may persist for decades. This is especially crucial because pyrogenic changes to vegetation communities can alter the interactions of understory and overstory vegetation, with important management implications for forest and woodland ecosystems. The response of shrub species to fire severity in NAM-dominated ecosystems likely differs from species in Mediterranean ecosystems because of temporal divergences in rainfall and humidity. Differences in seasonal rainfall in NAM-dominated ecosystems can modulate the intensity of disturbance relative to the phenological responses of shrub species, with late spring or early summer fires causing overstory tree mortality and setting the stage for post-fire shrub recovery. NAM-dominated ecosystems generally experience the highest severity fires during the growing season, which allows seeding and resprouting species to recover following summertime rains. Additionally, fires occurring during the monsoon season generally burn at lower severities than in the fore-monsoon drought, and may not tend to alter the persistent structure of both tree and shrub species.
These findings provide new insights into the post-disturbance structure and dynamics of shrub species in NAM-dominated forested ecosystems, a climate region in which shrub species have received relatively little research attention. Ultimately, understanding the post-fire response of shrubs to variations in fire severity provides land and fire managers with valuable information not only for predicting immediate post-fire recovery and canopy structure of various shrub species, but also for forecasting potential vegetation structure for decades into the future. Widespread tree mortality and increasing fire size and severity combine to place additional importance on understanding the post-fire dynamics of shrub species in forested ecosystems.
Acknowledgments
Research funding was provided by the Coronado National Forest, through the Arizona FireScape program (www.azfirescape.org). Alessandro Marafino, Brett Meyers, and Joshua Sutter assisted with field measurements. Steve Yool provided helpful comments on the manuscript. We acknowledge Helen Poulos, Drew Barton, Craig Wilcox, and José Iniguez for their deep knowledge of Madrean Sky Island forests, and their helpful comments on natural history in the Chiricahuas. Publication costs were supported by the University of Arizona Libraries Open Access Publication Fund. We thank two anonymous reviewers for their helpful comments on this manuscript.
Author Contributions
Jesse Minor and Don A. Falk conceived and designed the experiments; Jesse Minor conducted the fieldwork and performed the analysis; Jesse Minor and Greg A. Barron-Gafford analyzed the data; Jesse Minor, Don A. Falk, and Greg A. Barron-Gafford wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Species identified in sampling plots, categorized by post-fire regeneration strategies based on the Fire Effects Information System (FEIS) [15].
Table A1.
Species identified in sampling plots, categorized by post-fire regeneration strategies based on the Fire Effects Information System (FEIS) [15].
| Study Species by Post-Fire Regeneration Strategy | Number of Patches (n = 867) |
|---|---|
| Obligate Seeder | 197 |
| Ceanothus fendleri (Rhamnaceae) | 143 |
| Ptelea trifoliata (Rutaceae) | 15 |
| Ribes pinetorum (Grossulariaceae) | 38 |
| Sambucus coerulea (Adoxaceae) | 1 |
| Obligate Resprouter | 670 |
| Arbutus arizonica (Ericaceae) | 1 |
| Cercocarpus betuloides (Rosaceae) | 2 |
| Mimosa biuncifera (Fabaceae) | 34 |
| Physocarpus monogynus (Rosaceae) | 2 |
| Quercus arizonica (Fagaceae) | 239 |
| Quercus emoryi (Fagaceae) | 40 |
| Quercus gambelii (Fagaceae) | 27 |
| Quercus hypoleucoides (Fagaceae) | 219 |
| Quercus rugosa (Fagaceae) | 11 |
| Frangula betulaefolia (Rhamnaceae) | 6 |
| Robinia neomexicana (Fabaceae) | 83 |
| Symphoricarpos rotundifolius (Caprifoliaceae) | 6 |
Appendix B
References used to categorize woody shrub species by post-fire regeneration strategy. Note: in some cases, classification into post-fire regeneration strategy was made on the basis of similar species within the same genus.
Arbutus arizonica (Gray) Sarg.
Pavek, D.S. Arbutus arizonica. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1993. Available online: https://www.fs.fed.us/database/feis/plants/tree/arbari/all.html (accessed on 20 January 2016).
Cercocarpus betuloides Nutt.
Gucker, C.L. Cercocarpus montanus. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 2006. Available: http://www.fs.fed.us/database/feis/plants/shrub/cermon/all.html (accessed on 20 January 2016).
Ceanothus fenderli
Zouhar, K.L. Ceanothus greggii. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 2000. Available online: https://www.fs.fed.us/database/feis/plants/shrub/ceaint/all.html (accessed on 20 January 2016).
Mimosa biuncifera
Uchytil, R.J. Mimosa aculeaticarpa var. biuncifera. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1990. Available online: https://www.fs.fed.us/database/feis/plants/shrub/mimacub/all.html (accessed on 20 January 2016).
Physocarpus monogynus
Habeck, R.J. Physocarpus malvaceus. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1992. Available online: https://www.fs.fed.us/database/feis/plants/shrub/phymal/all.html (accessed on 20 January 2016).
Ptelea trifoliata (L.) ssp. pallida (Greene)
USDA Natural Resources Conservation Service. Conservation Plant Characteristics. Ptelea trifoliata L., Common hoptree, PTTR. Available online: http://plants.usda.gov/java/charProfile?symbol=PTTR&format=print (accessed on 20 January 2016).
Quercus arizonica Sarg.
Pavek, D.S. Quercus arizonica. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1994. Available: https://www.fs.fed.us/database/feis/plants/tree/queari/all.html (accessed on 20 January 2016).
Quercus emoryi Torr.
Pavek, D.S. Quercus emoryi. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1994. Available online: https://www.fs.fed.us/database/feis/plants/tree/queemo/all.html (accessed on 20 January 2016).
Quercus gambelii Nutt.
Simonin, K.A. Quercus gambelii. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 2000. Available online: https://www.fs.fed.us/database/feis/plants/tree/quegam/all.html (accessed on 20 January 2016).
Rhamnus californica
McMurray, N.E. Frangula californica. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1990. Available online: http://www.fs.fed.us/database/feis/plants/shrub/fracal (accessed on 20 January 2016).
Ribes pinetorum
Marshall, K. A. Ribes montigenum. In: Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1995. Available online: https://www.fs.fed.us/database/feis/plants/shrub/ribmon/all.html (accessed on 20 January 2016).
Robinia neomexicana Gray.
Pavek, D.S. Robinia neomexicana. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1993. Available online: https://www.fs.fed.us/database/feis/plants/tree/robneo/all.html (accessed on 20 January 2016).
Sambucus coerulea
Crane, M.F. Sambucus nigra subsp. cerulea. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1989. Available online: https://www.fs.fed.us/database/feis/plants/shrub/samnigc/all.html (accessed on 20 January 2016).
Symphoricarpos rotundifolius
Scasta, J.D.; Engle, D.M.; Harr, R.N.; Debinski, D.M. Fire Induced Reproductive Mechanisms of a Symphoricarpos (Caprifoliaceae) Shrub after Dormant Season Burning. Bot. Stud. 2014, 55, 80.
Bates, J.D.; Sharp, R.N.; Davies, K.W. Sagebrush Steppe Recovery after Fire Varies by Development Phase of Juniperus occidentalis Woodland. Int. J. Wildland Fire 2014, 23, 117–130.
Aleksoff, K.C. Symphoricarpos oreophilus. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 1999. Available online: https://www.fs.fed.us/database/feis/plants/shrub/symore/all.html (accessed on 20 January 2016).
References
- Buhk, C.; Meyn, A.; Jentsch, A. The Challenge of Plant Regeneration after Fire in the Mediterranean Basin: Scientific Gaps in our Knowledge on Plant Strategies and Evolution of Traits. Plant Ecol. 2007, 192, 1–19. [Google Scholar] [CrossRef]
- Pratt, R.B.; Jacobsen, A.L.; Ramirez, A.R.; Helms, A.M.; Traugh, C.A.; Tobin, M.F.; Heffner, M.S.; Davis, S.D. Mortality of Resprouting Chaparral Shrubs after a Fire and during a Record Drought: Physiological Mechanisms and Demographic Consequences. Glob. Chang. Biol. 2014, 20, 893–907. [Google Scholar] [CrossRef] [PubMed]
- Stevens-Rumann, C.; Morgan, P. Repeated Wildfires Alter Forest Recovery of mixed-conifer Ecosystems. Ecol. Appl. 2016, 26, 1842–1853. [Google Scholar] [CrossRef] [PubMed]
- Shakesby, R.; Doerr, S. Wildfire as a Hydrological and Geomorphological Agent. Earth-Sci. Rev. 2006, 74, 269–307. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a Life History Strategy in Woody Plant Communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an Evolutionary Pressure Shaping Plant Traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Ackerly, D. Functional Strategies of Chaparral Shrubs in Relation to Seasonal Water Deficit and Disturbance. Ecol. Monogr. 2004, 74, 25–44. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.; Midgley, J.; Lamont, B.; Ojeda, F.; Burrows, G.; Enright, N.; Knox, K. Resprouting as a Key Functional Trait: How Buds, Protection and Resources Drive Persistence after Fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Barton, A.M. Intense Wildfire in Southeastern Arizona: Transformation of a Madrean oak-pine Forest to Oak Woodland. For. Ecol. Manag. 2002, 165, 205–212. [Google Scholar] [CrossRef]
- Barton, A. Fire Adaptations in Pines and Oaks: Tree Population Responses to Fire Suppression in Arizona's Madrean Forests. Gen. Tech. Rep. INT-GTR-320 (USA) 1995, 159–163. [Google Scholar]
- Guiterman, C.H.; Margolis, E.Q.; Swetnam, T.W. Dendroecological Methods for Reconstructing High-Severity Fire in Pine-Oak Forests. Tree-Ring Res. 2015, 71, 67–77. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and Stability of Ecological Systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Gunderson, L.H. Ecological resilience—In Theory and Application. Annu. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef]
- Brooks, M.L.; Chambers, J.C. Resistance to Invasion and Resilience to Fire in Desert Shrublands of North America. Rangel. Ecol. Manag. 2011, 64, 431–438. [Google Scholar] [CrossRef]
- Fire Effects Information System (FEIS). Available online: https://www.feis-crs.org/feis/ (accessed on 20 January 2016).
- Fukami, T.; Nakajima, M. Community Assembly: Alternative Stable States or Alternative Transient States? Ecol. Lett. 2011, 14, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.A. Mega-Fires, Tipping Points and Ecosystem Services: Managing Forests and Woodlands in an Uncertain Future. For. Ecol. Manag. 2013, 294, 250–261. [Google Scholar] [CrossRef]
- Falk, D.A. Are Madrean Ecosystems Approaching Tipping Points? Anticipating Interactions of Landscape Disturbance and Climate Change. In Proceedings of the Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III and 7th Conference on Research and Resource Management in the Southwestern Deserts, Tucson, AZ, USA, 1–5 May 2012; pp. 40–47. [Google Scholar]
- Law, R.; Morton, R.D. Alternative Permanent States of Ecological Communities. Ecology 1993, 74, 1347–1361. [Google Scholar] [CrossRef]
- VanderWeide, B.L.; Hartnett, D.C. Fire Resistance of Tree Species Explains Historical Gallery Forest Community Composition. For. Ecol. Manag. 2011, 261, 1530–1538. [Google Scholar] [CrossRef]
- Graves, S.J.; Rifai, S.W.; Putz, F.E. Outer Bark Thickness Decreases More with Height on Stems of Fire-Resistant than Fire-Sensitive Floridian Oaks (Quercus spp.; Fagaceae). Am. J. Bot. 2014, 101, 2183–2188. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.K.; Comrie, A.C. The North American Monsoon. Bull. Am. Meteorol. Soc. 1997, 78, 2197–2213. [Google Scholar] [CrossRef]
- Biswell, H.H. Prescribed Burning in California Wildlands Vegetation Management; University of California Press: Berkeley, CA, USA, 1989. [Google Scholar]
- Trouet, V.; Taylor, A.H.; Carleton, A.M.; Skinner, C.N. Fire-climate Interactions in Forests of the American Pacific Coast. Geophys. Res. Lett. 2006, 33, L18704. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Betancourt, J.L. Mesoscale Disturbance and Ecological Response to Decadal Climatic Variability in the American Southwest. J. Clim. 1998, 11, 3128–3147. [Google Scholar] [CrossRef]
- Westerling, A.L.; Gershunov, A.; Brown, T.J.; Cayan, D.R.; Dettinger, M.D. Climate and Wildfire in the Western United States. Bull. Am. Meteorol. Soc. 2003, 84, 595–604. [Google Scholar] [CrossRef]
- Barger, N.N.; Archer, S.R.; Campbell, J.L.; Huang, C.; Morton, J.A.; Knapp, A.K. Woody Plant Proliferation in North American Drylands: A Synthesis of Impacts on Ecosystem Carbon Balance. J. Geophys. Res. Biogeosci. 2011, 116, G00K07. [Google Scholar] [CrossRef]
- Knapp, A.K.; Briggs, J.M.; Collins, S.L.; Archer, S.R.; Bret-Harte, M.S.; Ewers, B.E.; Peters, D.P.; Young, D.R.; Shaver, G.R.; Pendall, E. Shrub Encroachment in North American Grasslands: Shifts in Growth Form Dominance Rapidly Alters Control of Ecosystem Carbon Inputs. Glob. Chang. Biol. 2008, 14, 615–623. [Google Scholar] [CrossRef]
- Notaro, M.; Mauss, A.; Williams, J.W. Projected Vegetation Changes for the American Southwest: Combined Dynamic Modeling and bioclimatic-envelope Approach. Ecol. Appl. 2012, 22, 1365–1388. [Google Scholar] [CrossRef] [PubMed]
- Petrie, M.; Collins, S.; Swann, A.; Ford, P.; Litvak, M. Grassland to Shrubland State Transitions Enhance Carbon Sequestration in the Northern Chihuahuan Desert. Glob. Chang. Biol. 2015, 21, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Huxman, T.E.; Wilcox, B.P.; Breshears, D.D.; Scott, R.L.; Snyder, K.A.; Small, E.E.; Hultine, K.; Pockman, W.T.; Jackson, R.B. Ecohydrological Implications of Woody Plant Encroachment. Ecology 2005, 86, 308–319. [Google Scholar] [CrossRef]
- Scott, R.L.; Huxman, T.E.; Williams, D.G.; Goodrich, D.C. Ecohydrological Impacts of woody-plant Encroachment: Seasonal Patterns of Water and Carbon Dioxide Exchange within a Semiarid Riparian Environment. Glob. Chang. Biol. 2006, 12, 311–324. [Google Scholar] [CrossRef]
- Barton, A.M. Pines Versus Oaks: Effects of Fire on the Composition of Madrean Forests in Arizona. For. Ecol. Manag. 1999, 120, 143–156. [Google Scholar] [CrossRef]
- Caprio, A.C.; Zwolinski, M.J. Fire and Vegetation in a Madrean Oak Woodland, Santa Catalina Mountains, Southeastern Arizona; USDA For. Serv. Gen. Tech. Rep. RM-218; USDA Forest Service: Fort Collins, CO, USA, 1994; pp. 389–398.
- Keeley, J.E.; Zedler, P.H. Reproduction of Chaparral Shrubs after Fire: A Comparison of Sprouting and Seeding Strategies. Am. Midl. Nat. 1978, 99, 142–161. [Google Scholar] [CrossRef]
- McLaughlin, S.P. An Overview of the Flora of the Sky Islands, Southeastern Arizona: Diversity, Affinities, and Insularity. In Biodiversity and Management of the Madrean Archipelago: The Sky Islands of Southwestern United States and Northwestern Mexico; USDA Forest Service General Technical Report RM-GTR 264; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 1994; pp. 60–70. [Google Scholar]
- Richardson, D.M. Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Van Devender, T.R.; Avila-Villegas, S.; Emerson, M.; Turner, D.; Flesch, A.D.; Deyo, N.S. Biodiversity in the Madrean Archipelago of Sonora, Mexico. In Proceedings of the Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III, Tucson, AZ, USA, 1–5 May 2012; pp. 10–16. [Google Scholar]
- Sawyer, D.A.; Kinraide, T.B. The Forest Vegetation at Higher Altitudes in the Chiricahua Mountains, Arizona. Am. Midl. Nat. 1980, 104, 224–241. [Google Scholar] [CrossRef]
- Malusa, J.; Falk, D.; Laing, L.; Gebow, B. Mapping Ecological Systems in Southeastern Arizona. In Proceedings of the Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III and 7th Conference on Research and Resource Management in the Southwestern Deserts, Tucson, AZ, USA, 1–5 May 2012; pp. A1–A7. [Google Scholar]
- Arizona FireScape. Chiricahua Ecological Systems Index. Available online: https://azfirescape.org/chiricahua/ecosystems_descriptions (accessed on 10 May 2015).
- Kaib, M.; Baisan, C.H.; Grissino-Mayer, H.D.; Swetnam, T.W. Fire History of the Gallery Pine-Oak Forests and Adjacent Grasslands of the Chiricahua Mountains of Arizona; United States Department of Agriculture Forest Service General Technical Report RM; United States Department of Agriculture Forest Service: Fort Collins, CO, USA; 1996; pp. 253–264.
- Morino, K.A.; Baisan, C.H.; Swetnam, T.W. Historical Fire Regimes in the Chiricahua Mountains, Arizona: An Examination of Fire along an Elevation Gradient and in Mixed-Confier Forest: Final Report; Laboratory of Tree-Ring Research, The University of Arizona: Tucson, AZ, USA, 2000. [Google Scholar]
- Swetnam, T.W.; Baisan, C.H.; Caprio, A.C.; McCord, A.; Brown, P.M. Fire and Flood in a Canyon Woodland: The Effects of Floods and Debris Flows on the Past Fire Regime of Rhyolite Canyon, Chiricahua National Monument; Report to the National Park Service; The University of Arizona: Tucson, AZ, USA, 1990; pp. 1–51. [Google Scholar]
- Seklecki, M.T.; Grissino-Mayer, H.D.; Swetnam, T.W. Fire History and the Possible Role of Apache-Set Fires in the Chiricahua Mountains of Southeastern Arizona; United States Department of Agriculture Forest Service General Technical Report RM; United States Department of Agriculture Forest Service: Fort Collins, CO, USA, 1996; pp. 238–246.
- Minor, J.J.; (University of Arizona, Tucson, AZ, USA). Personal communication, 2017.
- Monitoring Trends in Burn Severity: MTBS Project. MTBS Data Access: Fire Level Geospatial Data. Available online: http://mtbs.gov/dataquery/individualfiredata.html (accessed on 20 May 2015).
- LANDFIRE. LANDFIRE: Existing Vegetation Type. Available online: http://landfire.cr.usgs.gov/viewer/ (accessed on 12 May 2015).
- Pavek, D.S. Quercus arizonica. 1994. Available online: https://www.fs.fed.us/database/feis/plants/tree/queari/all.html (accessed on 20 January 2016).
- Aleksoff, K.C. Symphoricarpos oreophilus. 1999. Available online: https://www.fs.fed.us/database/feis/plants/shrub/symore/all.html (accessed on 20 January 2016).
- Simonin, K.A. Quercus gambelii. 2000. Available online: https://www.fs.fed.us/database/feis/plants/tree/quegam/all.html (accessed on 20 January 2016).
- Cook, R.D.; Weisberg, S. Residuals and Influence in Regression; Chapman and Hall: New York, NY, USA, 1982. [Google Scholar]
- Lavorel, S. Ecological Diversity and Resilience of Mediterranean Vegetation to Disturbance. Divers. Distrib. 1999, 5, 3–13. [Google Scholar] [CrossRef]
- Scasta, J.D.; Engle, D.M.; Harr, R.N.; Debinski, D.M. Fire Induced Reproductive Mechanisms of a Symphoricarpos (Caprifoliaceae) Shrub after Dormant Season Burning. Bot. Stud. 2014, 55, 80. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, Á.; Prior, L.D.; Perry, G.L.; Bowman, D.M. High Post-Fire Mortality of Resprouting Woody Plants in Tasmanian Mediterranean-Type Vegetation. Int. J. Wildland Fire 2017, 26, 532–537. [Google Scholar] [CrossRef]
- Keeley, J.E. Role of Fire in Seed Germination of Woody Taxa in California Chaparral. Ecology 1987, 68, 434–443. [Google Scholar] [CrossRef]
- Keeley, J.E.; Fotheringham, C. Role of Fire in Regeneration from Seed. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI: Wallingford, UK, 2000; pp. 311–330. [Google Scholar]
- Zouhar, K.L. Ceanothus greggii. 2000. Available online: https://www.fs.fed.us/database/feis/plants/shrub/ceagre/all.html (accessed on 20 January 2016).
- Quick, C.R. Notes on the Germination of Ceanothus Seeds. Madroño 1935, 3, 135–140. [Google Scholar]
- Sweeney, J.R. Responses of Vegetation to Fire: A Study of the Herbaceous Vegetation Following Chaparral Fires; University of California Publications in Botany; University of California Press: Berkeley, CA, USA, 1956; Volume 28, pp. 143–216. [Google Scholar]
- Crane, M.F. Sambucus nigra subsp. cerulea. 1989. Available online: https://www.fs.fed.us/database/feis/plants/shrub/samnigc/all.html (accessed on 20 January 2016).
- Quick, C.R. Resurgence of a Gooseberry Population after Fire in Mature Timber. J. For. 1962, 60, 100–103. [Google Scholar]
- Sparks, J.C.; Masters, R.E.; Engle, D.M.; Palmer, M.W.; Bukenhofer, G.A. Effects of Late growing-season and Late dormant-season Prescribed Fire on Herbaceous Vegetation in Restored pine-grassland Communities. J. Veg. Sci. 1998, 9, 133–142. [Google Scholar] [CrossRef]
- Knapp, E.E.; Schwilk, D.W.; Kane, J.M.; Keeley, J.E. Role of Burning Season on Initial Understory Vegetation Response to Prescribed Fire in a Mixed Conifer Forest. Can. J. For. Res. 2006, 37, 11–22. [Google Scholar] [CrossRef]
- Garrison, G.A. Carbohydrate Reserves and Response to Use; USDA For. Serv. Gen. Tech. Rep. GTR-INT-1; United States Department of Agriculture Forest Service: Fort Collins, CO, USA, 1972.
- Harrington, M.G. Notes: The Effects of Spring, Summer, and Fall Burning on Gambel Oak in a Southwestern Ponderosa Pine Stand. For. Sci. 1985, 31, 156–163. [Google Scholar]
- Swetnam, T.W.; Betancourt, J.L. Fire-Southern Oscillation Relations in the Southwestern United States. Science 1990, 249, 1017. [Google Scholar] [CrossRef] [PubMed]
- Enright, N.; Fontaine, J.; Westcott, V.; Lade, J.; Miller, B. Fire Interval Effects on Persistence of Resprouter Species in Mediterranean-Type Shrublands. Plant Ecol. 2011, 212, 2071–2083. [Google Scholar] [CrossRef]
- Stone, E.; Stone, M. Root Collar Sprouts in Pine. J. For. 1954, 52, 487–491. [Google Scholar]
- McCune, B. Ecological Diversity in North American Pines. Am. J. Bot. 1988, 353–368. [Google Scholar] [CrossRef]
- Rowe, J.S. Concepts of Fire Effects on Plant Individuals and Species. In The Role of Fire in Northern Circumpolar Ecosystems; Rowe, R.W., MacLean, D.A., Eds.; John Wiley & Sons: Chichester, UK, 1983; pp. 135–154. [Google Scholar]
- Babb, G.D. Sprouting Response of Quercus arizonica and Quercus emoryi Following Fire. Master’s Thesis, The University of Arizona, Tucson, AZ, USA, 1992. [Google Scholar]
- Schwilk, D.W.; Gaetani, M.S.; Poulos, H.M. Oak Bark Allometry and Fire Survival Strategies in the Chihuahuan Desert Sky Islands, Texas, USA. PLoS ONE 2013, 8, e79285. [Google Scholar] [CrossRef] [PubMed]
- Barton, A.M. Factors Controlling Plant Distributions: Drought, Competition, and Fire in Montane Pines in Arizona. Ecol. Monogr. 1993, 63, 367–397. [Google Scholar] [CrossRef]
- Poulos, H.M. A Review of the Evidence for Pine-Oak Niche Differentiation in the American Southwest. J. Sustain. For. 2009, 28, 92–107. [Google Scholar] [CrossRef]
- White, A.S. Presettlement Regeneration Patterns in a Southwestern Ponderosa Pine Stand. Ecology 1985, 66, 589–594. [Google Scholar] [CrossRef]
- Brown, D.J.; Mali, I.; Forstner, M.R. Wildfire and Postfire Restoration Action Effects on Microclimate and Seedling Pine Tree Survivorship. J. Fish Wildl. Manag. 2014, 5, 174–182. [Google Scholar] [CrossRef]
Figure 1.
Plot locations within three Ecological Systems in the Chiricahua Mountains, Arizona, USA. White areas within the Horseshoe 2 Fire perimeter correspond to xeric grassland, savanna, and woodland vegetation types that were not included in this analysis. Plot frequencies are indicated by both fire severity and vegetation type. No plots were sampled within the high-elevation Aspen-Mixed Conifer vegetation type.
Figure 1.
Plot locations within three Ecological Systems in the Chiricahua Mountains, Arizona, USA. White areas within the Horseshoe 2 Fire perimeter correspond to xeric grassland, savanna, and woodland vegetation types that were not included in this analysis. Plot frequencies are indicated by both fire severity and vegetation type. No plots were sampled within the high-elevation Aspen-Mixed Conifer vegetation type.
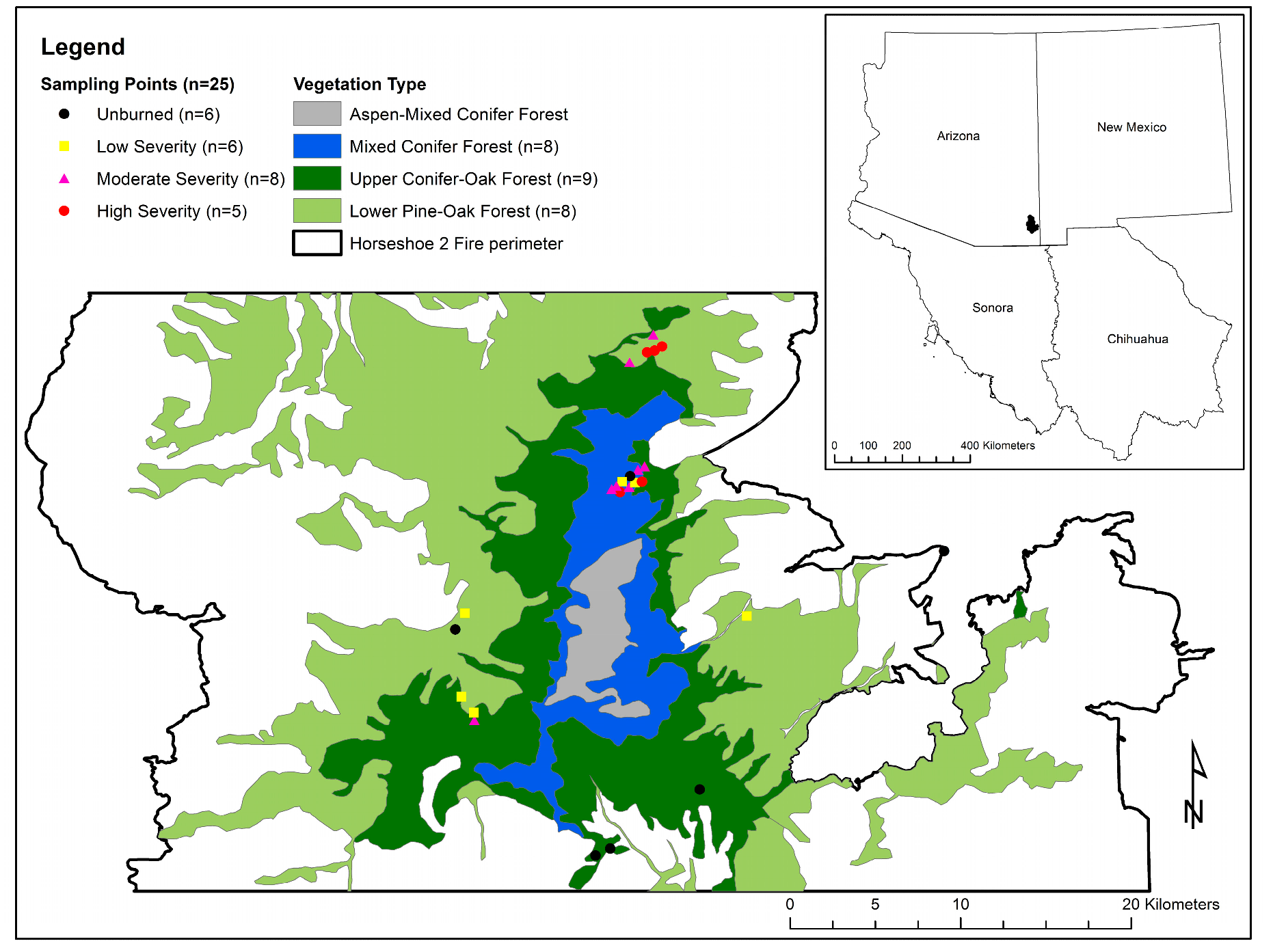
Figure 2.
(a) Shrub patch structure of obligate post-fire reseeding species; and (b) shrub patch structure of obligate post-fire resprouting species, in the study area. Shrub species show a differential response in the relationships between shrub patch area and stem density across regeneration strategies.
Figure 2.
(a) Shrub patch structure of obligate post-fire reseeding species; and (b) shrub patch structure of obligate post-fire resprouting species, in the study area. Shrub species show a differential response in the relationships between shrub patch area and stem density across regeneration strategies.
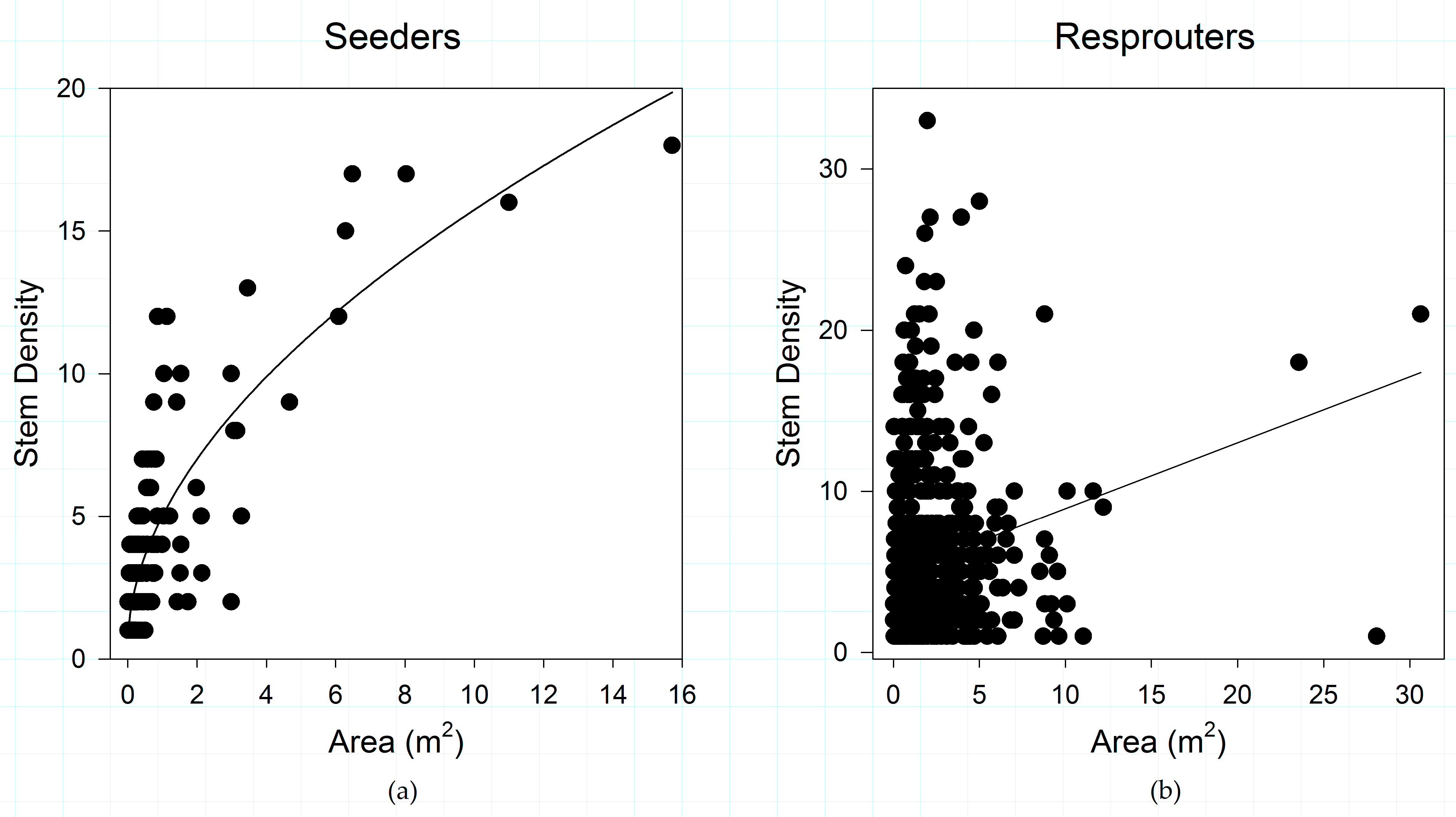
Figure 3.
Fire severity effects on obligate seeding species across fire severities in: (a) unburned plots; and (b) high severity plots, in the study area. Obligate sprouting species show no pattern in shrub structure in any fire severity class, but obligate seeders growing in unburned, moderate-severity, and high-severity burned areas demonstrate characteristic patterns in shrub patch structure.
Figure 3.
Fire severity effects on obligate seeding species across fire severities in: (a) unburned plots; and (b) high severity plots, in the study area. Obligate sprouting species show no pattern in shrub structure in any fire severity class, but obligate seeders growing in unburned, moderate-severity, and high-severity burned areas demonstrate characteristic patterns in shrub patch structure.
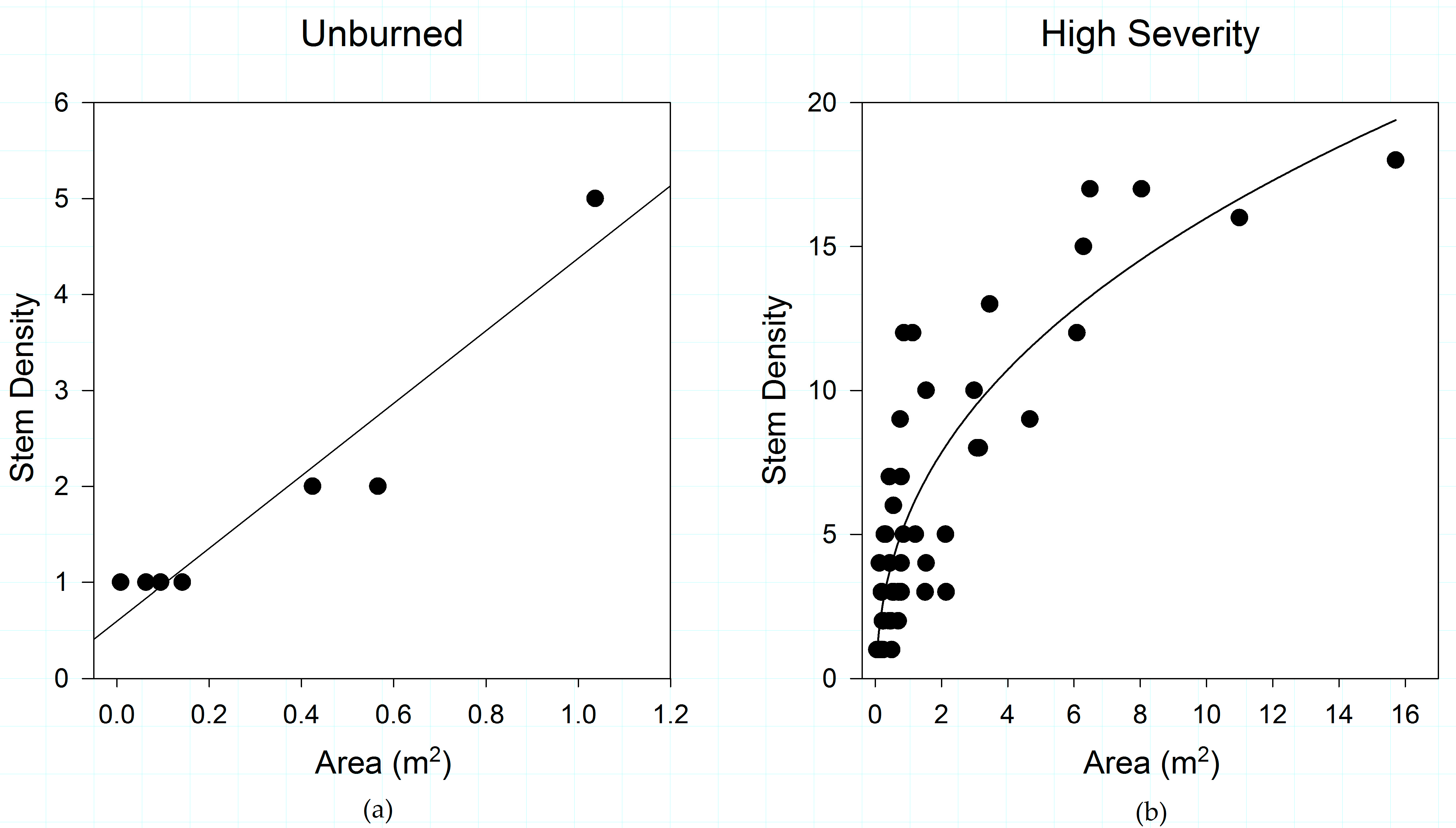
Figure 4.
(a) Proportional abundance of stem density among shrub species by post-fire regeneration strategy across fire severity classes; and (b) Proportion of patch area among shrub species by post-fire regeneration strategy across fire severity classes, in the study area. The pattern of proportional dominance is similar regardless of which metric of shrub patch structure is used. Obligate seeding and obligate resprouting species are significantly different at every fire severity in terms of both stem density (Unburned p = 0.001; Low p < 0.001, Moderate p < 0.001; High p = 0.049) and patch area (p < 0.001 for all severities).
Figure 4.
(a) Proportional abundance of stem density among shrub species by post-fire regeneration strategy across fire severity classes; and (b) Proportion of patch area among shrub species by post-fire regeneration strategy across fire severity classes, in the study area. The pattern of proportional dominance is similar regardless of which metric of shrub patch structure is used. Obligate seeding and obligate resprouting species are significantly different at every fire severity in terms of both stem density (Unburned p = 0.001; Low p < 0.001, Moderate p < 0.001; High p = 0.049) and patch area (p < 0.001 for all severities).
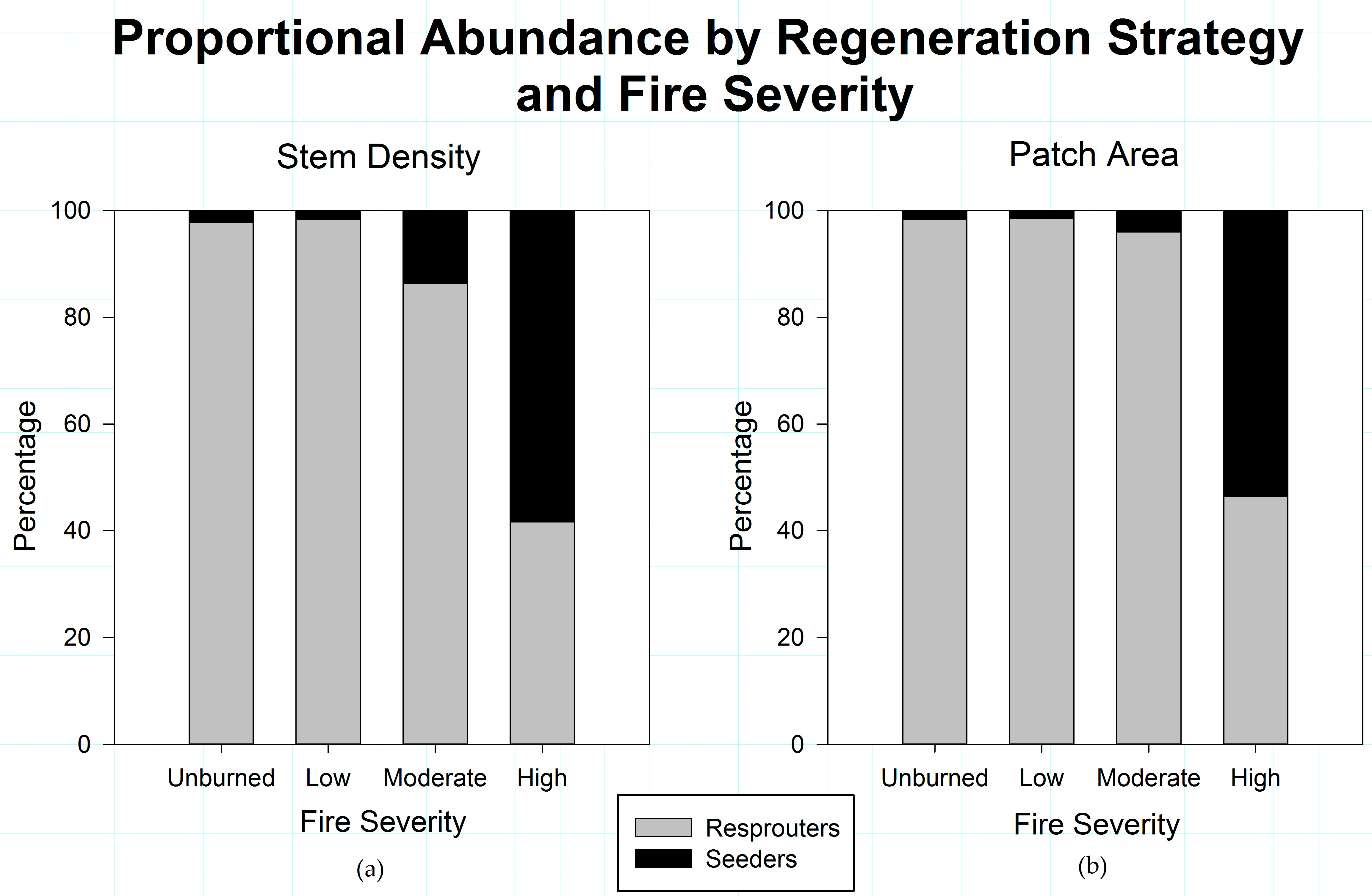
Figure 5.
(a) Shrub patch structural relationships for moderate-severity fire; and (b) Shrub patch structural relationships for high-severity fire, in the study area. Fire severity has an organizing effect on shrub patch structure. Increases in fire severity show a strengthening in the relationship between shrub canopy area and stem density.
Figure 5.
(a) Shrub patch structural relationships for moderate-severity fire; and (b) Shrub patch structural relationships for high-severity fire, in the study area. Fire severity has an organizing effect on shrub patch structure. Increases in fire severity show a strengthening in the relationship between shrub canopy area and stem density.
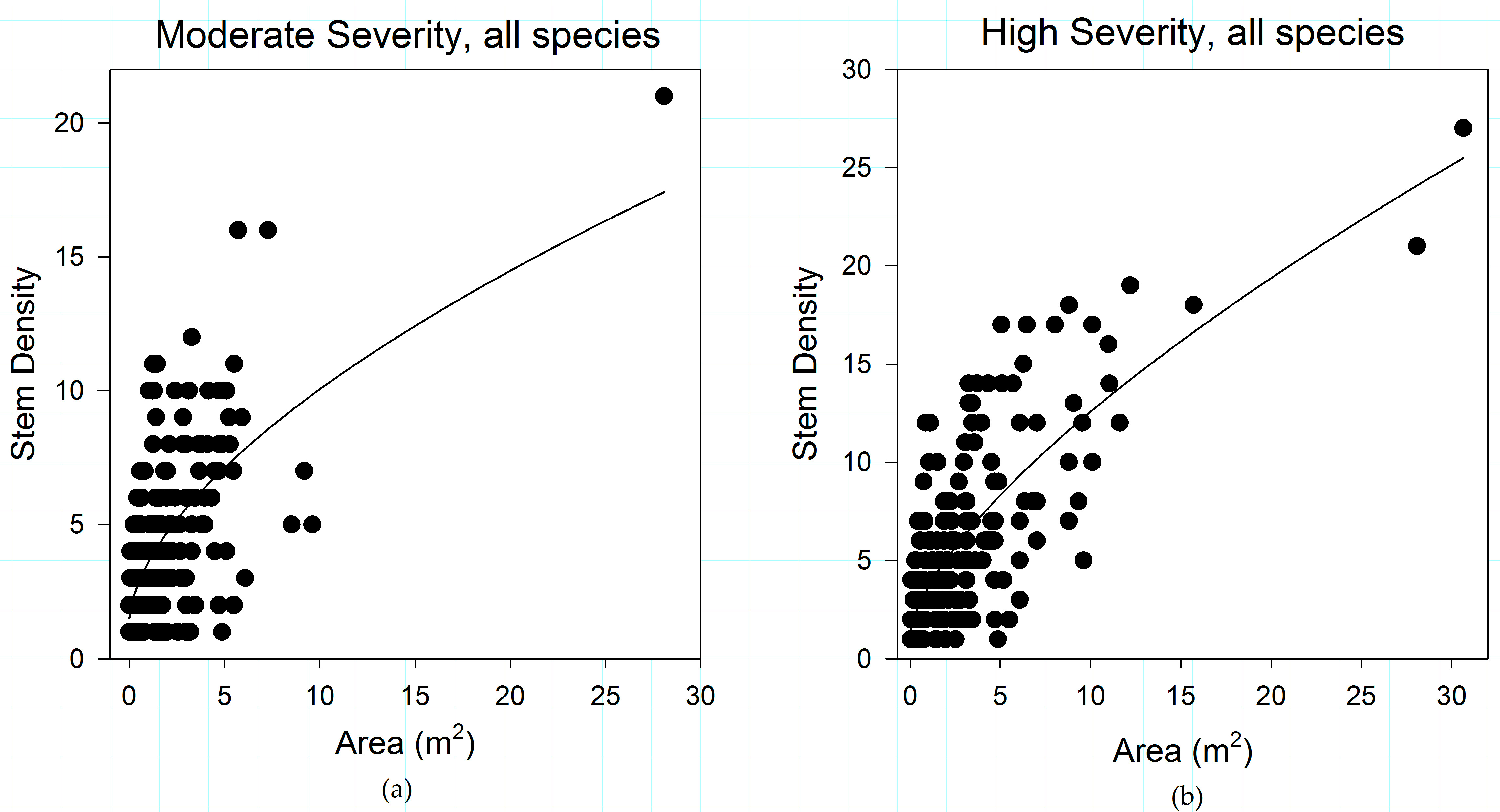
Figure 6.
Shrub patch structural relationships of (a) plots that did not experience fire in 1994 or 2011; (b) plots that experienced low-severity fire in 1994 and went unburned in 2011; and (c) plots that experienced high-severity fire in 1994 and went unburned in 2011, in the study area. The organizing effect of increasing fire severity observed in plots burned in 2011 (Figure 3 and Figure 5) can be observed in plots burned in 1994 and unburned in 2011. The organizing effect of fire severity on shrub patch structure may persist for decades.
Figure 6.
Shrub patch structural relationships of (a) plots that did not experience fire in 1994 or 2011; (b) plots that experienced low-severity fire in 1994 and went unburned in 2011; and (c) plots that experienced high-severity fire in 1994 and went unburned in 2011, in the study area. The organizing effect of increasing fire severity observed in plots burned in 2011 (Figure 3 and Figure 5) can be observed in plots burned in 1994 and unburned in 2011. The organizing effect of fire severity on shrub patch structure may persist for decades.
Table 1.
Descriptive statistical characteristics of shrub patch structure (patch area and stem density) across fire severity categories.
Table 1.
Descriptive statistical characteristics of shrub patch structure (patch area and stem density) across fire severity categories.
| Fire Severity | Patch Area (m2) | Stem Density (Stems/m2) | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean | Median | Maximum | C.V. | Mean | Median | Maximum | C.V. | |
| Unburned (n = 84) | 1.7 | 1.3 | 6.5 | 0.9 | 4.1 * | 3.1 | 6.4 | 1.2 |
| Low (n = 286) | 1.5 | 0.9 | 26.4 | 1.6 | 3.9 | 4.6 | 2.3 | 0.6 |
| Moderate (n = 312) | 1.4 * | 0.6 | 28.1 | 1.6 | 2.6 * | 4.8 | 0.7 | 0.5 |
| High (n = 184) | 2.7 * | 1.6 | 30.6 | 1.4 | 2.0 * | 2.6 | 0.9 | 0.6 |
n = number of patches sampled, in the study area. Coefficients of variation (C.V.) is the ratio of standard deviation to the mean. Asterisk (*) indicates significance of the mean at α = 0.05.
Table 2.
Mean shrub patch size and mean stem density across pairwise fire severity comparisons, in the study area.
Table 2.
Mean shrub patch size and mean stem density across pairwise fire severity comparisons, in the study area.
| Fire Severity | Stem Density (Stems/m2) | ||||
|---|---|---|---|---|---|
| Unburned | Low | Moderate | High | ||
| Canopy Area (m2) | Unburned | 8.7 | 10.9 ** | 5.6 | |
| Low | 1.5 | 10.0 ** | 7.0 * | ||
| Moderate | 1.4 * | 1.3 * | 8.7 ** | ||
| High | 2.4 * | 2.0 ** | 2.0 ** | ||
Values above the blank diagonal represent shrub stem density, and values below the blank diagonal represent shrub canopy area. Relevant comparisons are made in the columns. For example, in column 1, Unburned-Low compares unburned and moderate-severity fire in terms of canopy area. Significance was tested using pairwise Student’s t-Tests; a single asterisk (*) indicates significance at α = 0.05 and double asterisk (**) indicates significance at α = 0.001.
Table 3.
Relative dominance of shrub species by vegetation type within the study area located in the Chiricahua Mountains, Arizona, USA.
Table 3.
Relative dominance of shrub species by vegetation type within the study area located in the Chiricahua Mountains, Arizona, USA.
| Vegetation Type | Species | Total Shrub Patch Area (m2) | Measured Shrub Cover (%) | Measured Patches (n) |
|---|---|---|---|---|
| Lower Pine-Oak | Quercus arizonica | 435.6 | 47.8% | 199 |
| Quercus hypoleucoides | 325.3 | 35.7% | 139 | |
| Ceanothus fenderli † | 50.7 | 5.6% | 116 | |
| Quercus emoryi | 36.7 | 4.0% | 40 | |
| Quercus rugosa | 23.1 | 2.5% | 6 | |
| Mimosa biuncifera | 21.5 | 2.4% | 34 | |
| Quercus gambelii | 8.0 | 0.9% | 5 | |
| Symphoricarpos rotundifolius | 6.3 | 0.7% | 2 | |
| Robinia neomexicana | 3.7 | 0.4% | 4 | |
| Physocarpus monogynus | 0.2 | 0.1% | 1 | |
| Total | 911.1 | - | 546 | |
| Upper Pine-Oak | Quercus hypoleucoides | 142.0 | 27.0% | 76 |
| Robinia neomexicana | 134.5 | 25.6% | 56 | |
| Quercus arizonica | 93.8 | 17.9% | 39 | |
| Ceanothus fenderli † | 63.3 | 12.1% | 25 | |
| Quercus gambelii | 49.7 | 9.5% | 22 | |
| Quercus rugosa | 21.9 | 4.2% | 5 | |
| Cercocarpus betuloides | 8.9 | 1.7% | 2 | |
| Frangula betulaefolia | 3.8 | 0.7% | 5 | |
| Physocarpus monogynus | 1.7 | 0.3% | 1 | |
| Arbutus arizonica | 1.5 | 0.3% | 1 | |
| Symphoricarpos rotundifolius | 0.7 | 0.1% | 3 | |
| Ribes pinetorum † | 0.2 | 0.1% | 2 | |
| Total | 522 | - | 237 | |
| Mixed Conifer | Robinia neomexicana | 73.7 | 65.0% | 23 |
| Ribes pinetorum † | 17.6 | 16.2% | 35 | |
| Ptelea trifoliata † | 5.6 | 5.7% | 15 | |
| Quercus hypoleucoides | 3.6 | 3.9% | 4 | |
| Sambucus coerulea † | 3.0 | 3.4% | 1 | |
| Frangula betulaefolia | 2.2 | 2.8% | 1 | |
| Ceanothus fenderli † | 0.2 | 1.1% | 1 | |
| Symphoricarpos rotundifolius | 0.05 | 1.0% | 1 | |
| Quercus arizonica | 0.01 | 0.9% | 1 | |
| Total | 106.0 | - | 84 |
Shrub dominance declines across the vegetation types used in this study in terms of both total shrub patch area and the number of patches measured. Shrub cover often represents a small fraction of the total area of measured plots, but certain species tend to dominate the distribution of shrub patch area. Within each vegetation type, species are sorted by total measured patch area and proportional dominance. Post-fire seeding species are indicated with †.
Table 4.
Proportional abundance of shrub species by post-fire reproductive strategy, in the study area.
Table 4.
Proportional abundance of shrub species by post-fire reproductive strategy, in the study area.
| Vegetation Type | Obligate Seeders | Obligate Resprouters | Total n |
|---|---|---|---|
| Lower Pine-Oak | 21.2 | 78.8 * | 546 |
| Upper Pine-Oak | 11.4 | 88.6 * | 237 |
| Mixed Conifer | 63.9 * | 36.1 | 84 |
Significant differences (indicated by asterisk (*)) are observed between dominance of obligate seeders in Mixed Conifer and obligate resprouters in Pine-Oak vegetation types (p < 0.001; Z = 9.35).
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Minor, J.; Falk, D.A.; Barron-Gafford, G.A. Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance. Forests 2017, 8, 221. https://doi.org/10.3390/f8070221
AMA Style
Minor J, Falk DA, Barron-Gafford GA. Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance. Forests. 2017; 8(7):221. https://doi.org/10.3390/f8070221
Chicago/Turabian StyleMinor, Jesse, Donald A. Falk, and Greg A. Barron-Gafford. 2017. "Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance" Forests 8, no. 7: 221. https://doi.org/10.3390/f8070221
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.