
Earth System Model Needs for Including the Interactive Representation of Nitrogen Deposition and Drought Effects on Forested Ecosystems

1
Environmental Science Division, Argonne National Laboratory, Argonne, IL 60493, USA
2
Department of Biological Sciences, University of Illinois, Chicago, IL 60607, USA
*
Author to whom correspondence should be addressed.
Forests 2017, 8(8), 267; https://doi.org/10.3390/f8080267
Submission received: 9 May 2017
/
Revised: 15 July 2017
/
Accepted: 24 July 2017
/
Published: 27 July 2017
(This article belongs to the Special Issue Climate Change Impacts on the Dynamics of Forest Ecosystems)
Abstract
:One of the biggest uncertainties of climate change is determining the response of vegetation to many co-occurring stressors. In particular, many forests are experiencing increased nitrogen deposition and are expected to suffer in the future from increased drought frequency and intensity. Interactions between drought and nitrogen deposition are antagonistic and non-additive, which makes predictions of vegetation response dependent on multiple factors. The tools we use (Earth system models) to evaluate the impact of climate change on the carbon cycle are ill equipped to capture the physiological feedbacks and dynamic responses of ecosystems to these types of stressors. In this manuscript, we review the observed effects of nitrogen deposition and drought on vegetation as they relate to productivity, particularly focusing on carbon uptake and partitioning. We conclude there are several areas of model development that can improve the predicted carbon uptake under increasing nitrogen deposition and drought. This includes a more flexible framework for carbon and nitrogen partitioning, dynamic carbon allocation, better representation of root form and function, age and succession dynamics, competition, and plant modeling using trait-based approaches. These areas of model development have the potential to improve the forecasting ability and reduce the uncertainty of climate models.
1. Introduction
Earth system models (ESMs) have been used to predict the climate’s response to increased CO2 emissions (or concentrations), but uncertainty in land carbon (C) feedbacks results in a wide spread of uncertainty in model results [1]. Part of this uncertainty lies in a general lack of knowledge of the physical processes responsible for the land feedbacks on the C cycle, which makes estimating the land C sink difficult. Adding to this uncertainty is the response of ecosystems in the face of multiple stressors, the impacts of which may be non-additive and will certainly be highly variable across ecosystems. One prime example is the current increase in nitrogen (N) deposition and the predicted increase in drought [2]. Most field studies isolate one environmental component (e.g., drought, elevated CO2, N deposition) to study plant behavior. This results in an incomplete knowledge of an ecosystem’s reaction to multiple stressors, which limits our forecasting capability. Vegetation responses have a strong influence on C storage. As plants adjust their partitioning strategy for C and other nutrients to optimize uptake while limiting costs to ensure survival, the quantity and quality of C stored and the way nutrients are recycled will change.
Anthropogenic production of reactive N from fossil fuel combustion and fertilizer synthesis has led to an increased availability of reactive N in ecosystems, and this has become a global problem [3]. The current N deposition rate in some systems is over 10 kg ha−1 year−1 [4], which is well over the deposition rate from natural sources at 0.5 kg ha−1 year−1 [5]. Nitrogen deposits from industrial and agriculture activities have led to significant N loading in soils, particularly in regions of Europe and the eastern United States [6,7]. These N additions are within the critical load of N for sensitive ecosystems [8,9]. This N loading may be further exacerbated by increases in N availability from climate influences (i.e., warming and elevated CO2) on internal N cycling, which can increase decomposition, mineralization, and biological nitrogen fixation (BNF) [10,11,12]. The consequences of N loading are well known and include changes in biodiversity [7,13,14], composition [15,16], productivity [17], leaching [18,19], and possibly nitrification [17]. Since N is strongly tied to C, understanding the impact of increased N availability in ecosystems in the context of climate change is crucial if we are to understand whether increased N availability will strengthen or weaken the land’s C sink capacity.
Understanding the vegetative response to N deposition is increasingly difficult due a wide variability of responses between and within species and across climate and soil regimes, as well as contrasting behavior when faced with other elements of climate change (e.g., elevated CO2 or drought). For example, increased N availability generally results in an increase in aboveground woody biomass and a decrease in root biomass, but experiments with drought indicate an increase in root biomass at the expense of aboveground tissue [20,21,22,23]. This has led to the conclusion that N deposition may increase ecosystem vulnerability to drought [24], but some studies find that N deposition may enhance the recovery of some species post drought [25]. Therefore, a large uncertainty exists for vegetation response to increased N deposition under drought, especially because many of these antagonistic responses exhibit nonlinear behavior.
Predictions of climate change indicate an increase in the frequency and severity of drought [2] in many regions that are already water stressed. For example, in the Amazon, the dry season is expected to lengthen and intensify, and the area affected by seasonal drought is expected to expand by up to 0.75 million km2 by the end of the century [26]. The southwest and central United States are forecast to experience increases in drought severity that exceed the severest mega drought events at millennial scales [27]. Areas that do not experience a change in total precipitation may still experience changes in the timing and intensity of rainfall events. Plants under drought stress can experience hydraulic failure, C starvation, and increased vulnerability to disease, pests, and fire. Drought during peak growth periods will likely be more detrimental; juvenile and shallow-rooted plants are most susceptible to drought, whereas plants that are able to modify biogeochemical feedbacks will increase their chance of survival [28]. In general, drought results in decreased stomatal conductance and decreased net primary productivity [27]. In the past, drought events over the Amazon have reduced C storage through mortality and slowed growth [29]; increased events in the future could result in a weaker C sink of boreal, temperate, and tropical biomes across the globe [30]. This would contrast with the increased productivity that can result from high N deposition. Most ecosystems have experienced an extreme drought in the past, but extreme drought in combination with other climate impacts such as N deposition can put unprecedented amounts of stress on ecosystems. Given that the co-occurrence of drought and increased N availability are highly probable, these climate effects must be considered together to predict ecosystem behavior.
To answer questions about the future of the terrestrial C sink, we need to understand the behavior of ecosystems under long-term chronic N deposition and drought, as well as the interactions between these phenomena. ESMs are important tools for exploring the relationships between climate and vegetation responses, but current models might not be able to capture these processes.
The ESM community has made great strides in improving the C cycle in land surface models (LSMs) due to improvements to biogeochemical and hydrological cycles. Some of these advances constrain land C sequestration by imposing N limitations on C fertilization [31], water table, and inundation dynamics [32] and even hydraulic redistribution [33]. However, these models still cannot capture mortality in trees from drought, changes in biomass partitioning due to nutrient limitations, or even acclimation [34]. For example, Ukkola et al. [35] found that 14 LSMs overestimate seasonal drought, due partly to how models handle soil hydrology and plant moisture stress. De Kauwe et al. [36] also found that LSMs exaggerate drought effects because most models assume the same drought sensitivity for all vegetation types. Adding the N cycle to LSMs results in productivity decreases that range from 7% to 64% [37], but the feedbacks between C and N are weak once a steady state is reached [38]. This suggests that ESMs still lack key processes. In order to simulate N deposition-drought interactions, we hypothesize that additional model developments are necessary to mimic ecosystem stress responses, particularly if ESMs are to represent the C cycle accurately.
This review seeks to determine whether the current ESM framework of the C cycle is sufficient to capture the vegetation response to the combined effects of drought and growing N loading in ecosystems. To that end, this paper will review the observed impacts on C uptake and allocation due to increased N availability, drought, and their combined effects. Next will be a review of how current ESMs represent the coupled carbon-nitrogen model for C uptake and partitioning. We will present recent developments in several state-of-the-art LSMs, including the limitations of the representations for capturing interactions from N deposition and drought stress. Finally, we will discuss which additional processes should be the focus of future model developments to reduce uncertainty and improve predictive power.
2. Observations of N Impacts
Nitrogen is a critical component in ecosystems; it drives productivity through photosynthesis processes [39]. Increased N deposition results in increased production because N is generally the most limiting nutrient for growth [37,40]. Bala et al. [41] estimate that an additional 175 PgC has been stored since the pre-industrial period due to increased N deposition. This increase results mostly from changes in photosynthesis (i.e., increase in foliar N) or shifts in C allocation to increase light availability (Figure 1 and Table S1). Several mechanisms will allow plants to respond to changes in N such as the ability to increase photosynthetic capacity, plasticity to alter C partitioning of resources, and the presence or absence of other limiting factors in the system (e.g., water, P, mycorrhizal associations, etc.). An overview of some of the consequences of N deposition in ecosystems is included in Figure 1.
Nitrogen deposition studies frequently find an increase in foliar N that results in decreased C:N ratios of leaves [42,43,44]. This relationship between N deposition and foliar N content has even been used to estimate critical loads of N in Europe [45]. Pregitzer et al. [42] suggested that the increase in foliar N is responsible for the increase in aboveground net primary productivity (ANPP) that is found under elevated N availability, considering the relationship N plays in photosynthetic capacity and C assimilation [39,46]. However, reductions in N use efficiency and C allocation shifts away from mycorrhizae may also result in increases in ANPP [47]. Increases in productivity can also lead to increases in litter production that can ultimately lead to increased N in soils.
Gains in productivity can be the result of changes in patterns of C partitioning within the plant, and they often seem to be correlated with increases in above ground biomass [42,48,49]. Most increases in aboveground biomass from increased N deposition are allocated to stems [42,50,51], particularly for small-diameter trees [52]. This response results in faster biomass accumulation, which produces taller, skinnier trees [52]. Although this is more likely to affect young trees or seedlings exposed to elevated N, Du and Fang [53] also found weak growth in a mature forest. This may increase the mortality of young trees as a result of light limitation or the respiratory costs of early rapid growth [52]. However, the increase in growth is not consistent or linear for all species. For example, de Vries et al. [50] found that tropical forests had the least response to N deposition compared to temperate and boreal forests, and in some cases a negative growth relationship exists at high N inputs [49]. Herbaceous plants also experience biomass increases from N deposition [48,54], and under low levels of N deposition they can increase aboveground biomass more than trees [51].
The increase in aboveground biomass is expected to come at the expense of belowground inputs [48]. However, it is unclear how roots respond to increased N availability; this response can include increases in productivity, even though decreases in biomass occur [55]. This occurs when increases in root turnover with N availability result in root biomass decreases over long time scales, even though C allocated to the roots likely increases. The root response is strongly associated with root sensitivity [56]; the heterogeneity of the soil, substrate, and climate can result in different root responses that will increase root vulnerability to damage or mortality.
Finally, increases in N deposition can increase N in forest biomass [57]. This increase in biomass will lead to changes in N cycling as biomass eventually flows from the canopy into the litter pool. The enhanced amount of N in the litter pool can cause faster decomposition [57] and increase N mineralization on the forest floor.
Nitrogen deposition can also increase susceptibility to disturbance factors. An increase in biomass N can lead to lowered resistance to pathogen infection or herbivory [58]. This increased vulnerability to pests can result in increased wildfires [59]. Ultimately, these changing conditions could lead to changes in species composition and biodiversity as a result of shifts in nutrient availability or toxicity, environment favorability, and competition [7].
3. Observations of Drought Impacts
Under drought conditions, plants typically undergo C starvation or hydraulic failure [60] (Figure 2 and Table S1). Either condition can be fatal. Carbon starvation occurs when stomata close to constrain water loss, resulting in significantly lower C assimilation that will not only halt growth, but also risk insufficient reserves to sustain plant maintenance requirements. Hydraulic failure occurs when xylems become damaged or collapse, limiting a plant’s ability to extract water. The timing of drought and phenology will play a key role that drives plant response [61]. For example, a drought event that coincides with the peak growth period will result in higher plant mortality than drought during a less active growth period. Another important consideration is drought intensity versus drought frequency [62]. A severe drought with a long duration will have a different impact on plant response and survival than short, frequent droughts. The length of time a plant has been exposed to drought cycles is an important driver of trait changes that increase drought tolerance. Over short time scales, stomata regulate water loss, but, over longer time scales, changes in allometry will occur to optimize hydraulic conductance [22]. Plant traits may change similarly across short and long drought intervals; for example, leaf area may decrease with decreasing precipitation, while other processes may be delayed (i.e., partitioning changes between the leaf and roots) or homeostatic (e.g., xylem vulnerability) [22]. Figure 2 shows some of the effects forests may experience under drought.
There are many ecological traits that can make a system more or less tolerant to drought, thereby leading to improved water use efficiency, including control over stomatal conductance, allometric plasticity, hydraulic redistribution [63], or even long-term acclimation. To survive drought, plants may also reduce C demand, which can be achieved by leaf senescence and the down-regulation of respiration [64].
Many field studies have observed increased stomatal closure [20,28,65] and increased water use efficiency during droughts [20]. However, observations also found species that have less stomatal control and may continue to transpire well into a drought event [65]. This may provide an advantage during short-term droughts but not necessarily during longer, more intense drought events. The ability to moderate water uptake and growth may be absent in the tropics; Rowland et al. [66] found no change in the growth rate of trees in the Amazon under a long-term drought.
Stomata closure will reduce photosynthesis and, over long periods, lead to lower biomass accumulation, litter production, and ultimately N mineralization in the soil [67]. Additional nutrient limitations can occur during drought because of low soil moisture levels, which act to reduce nutrient flow and diffusion in soils. These N limitations can exacerbate the effects of drought by limiting photosynthesis further.
Since plants continue to demand C for metabolic respiration, when stomata close during drought, plants rely on reserves to meet C requirements. The ability to mobilize and transport stored C may be impaired by drought but is critical for species survival [64]. Carbon reserves can help trees avoid C starvation. As these reserves are depleted, a tree may suffer from C starvation. Although this theory lacks testing, McDowell et al. [60] suspects that C starvation may result in the mortality of isohydric species under drought conditions.
Another adaptation to drought is through increases in the root-to-shoot ratio to maintain respiration during the early stages of drought, as suggested by the optimal partitioning theory. Hertel et al. [21] found a doubling of root production and an increase in root-to-leaf biomass and production under drought in a mature beech forest. Even under significant decreases in a plant’s relative growth rate, its root growth may remain constant [68]. The ability to re-translocate biomass from leaves and stems or utilize stored nonstructural carbohydrates can increase survival chances [62]. Furthermore, altering the morphological characteristics of roots (e.g., length and surface area) can also aid in fulfilling water demands [69].
The ability to extend root systems to deep soil layers [70] or move water through the soil column from depth via hydraulic redistribution can also increase a plant’s chance of surviving a drought. Not only can this effect transfer water upward into dry soils, allowing plants to maintain transpiration and photosynthesis during dry seasons or at night, it can move water downward to be protected from evaporation or competition. The obvious benefit of hydraulic redistribution is maintaining water potential below hydraulic failure limits, but neighboring plants can also take advantage of the new water that is available [71]. The quantity of water hydraulically transferred ranges from 0.04 mm H2O d−1 up to 1.3 mm H2O d−1 and may be as much as 80% of the water that the plant transpires [72].
Several other mechanisms can also play a role in drought mortality such as increased vulnerability to pests or fungal infection and fire [30,73]. These disturbances can have a devastating impact on forest mortality and the C cycle. Similar to N deposition impacts on vegetation, changes in biomass, composition, mortality, and nutrient cycling can lead to shifts in species competition and distribution [60] and the impairment of the mobilization and transport of stored C [64,73].
4. Interactions between N and Drought
The interactions between N and drought are difficult to determine because (1) the effects can depend on the timing of N deposition relative to drought; (2) most experiments are done with young trees or herbaceous plants and not with mature vegetation; (3) many studies impose only weak drought conditions that might not result in drought–N feedbacks [74]; and (4) the impacts vary with ecosystem and plant traits. However, in general, the effects from drought and N deposition are interdependent [24] and not always additive [75,76]. A list of studies that have examined N deposition-drought interactions is included in Table S1.
Drought seems to negate the increase in productivity observed from increased N deposition [77,78], although the effects of N addition may alleviate some of the impacts of drought on growth [78]. This can be partially attributed to the countering effects of N and drought on photosynthesis; in particular, N deposition tends to increase photosynthetic capacity, while drought limits photosynthesis. Other interactions between these two types of stress also occur. For example, N deposition tends to increase water use efficiency (WUE), but, during a severe drought, plants may lose the ability to utilize N, leading to a loss of biomass [77]. Some ecosystems experience a decline in above- and belowground biomass [24,79] and a loss of root functionality [79]. In some cases, changes in allocation between above- and belowground biomass result in higher root-to-shoot ratios [76].
The timing of a drought event is also an important factor considering N level effects. Plants are more susceptible to drought when N availability increases before drought because it leads to higher productivity, thereby increasing evaporative demands [24]. This can even result in a loss of N allocation control and adaptive strategies to mitigate drought effects [24]. Finally, increased N availability can lower concentrations of compounds that are important for stress tolerance, which could also increase mortality under more severe drought conditions [80].
Plant age also plays a role in determining the response of N deposition combined with drought. Palátová [81] found that the reduction in root biomass due to combined N deposition and drought was more severe for young seedlings than older trees. Trees tend to allocate more biomass to roots as they age, so younger trees are more susceptible to drought, and drought following N deposition further increases sensitivity to drought [76].
However, N deposition during drought may not always be detrimental. For example, drought can lock nutrients in an immobilized state by reducing the soil water available for decomposition [28], but N deposition can increase available nutrients. Increased N deposition can also aid the recovery after drought of some species [25]. For trees that preferentially shift allocations to stems under N deposition and drought [82], water stress may be alleviated by the extra water storage in the tree. In desert systems, N deposition relieved some of the negative effects of water stress through increases in root weight, leaf number, leaf area, biomass, and decreased root-to-shoot ratios [80,83].
5. Earth System Models
ESMs are designed to predict the climate state by integrating feedback between the atmosphere, land, and ocean. One goal of these models is to capture biosphere-atmosphere interactions in order to understand what will be the vegetation response to changing environmental conditions. The land component has undergone considerable growth over the last decade and now has complexity that rivals atmospheric models. This growth has greatly improved the climate forecasting ability of ESMs, but these models still fall short of providing a good estimate of the land C sink. The following section and Table 1 details some important processes already included in many LSMs that are necessary to capture vegetation responses to increased N and drought. The limitations of these model advances are also highlighted.
5.1. Nitrogen
An obvious and crucial component for capturing N deposition impacts on plants is the treatment of N, but early versions ESMs only considered the C cycle and neglected the N cycle completely. Carbon-only models miss a significant N deficit and therefore overestimate C sequestration by ecosystems under climate change [102,103]. Many of the latest versions of ESMs now include N (and a rare few include phosphorus). However, because N is often a limiting nutrient in ecosystems, the focus is on plant response under limited N conditions and on the effects of the progressive N limitation [104] that is expected under elevated CO2. Although some models offer prescribed N, more sophisticated ones employ an N pool (bulk or speciated) that is available to both plants and decomposers. Nitrogen additions come from sources including lightning, deposition, mineralization, and biological fixation. Losses are from plant uptake, immobilization, leaching, and nitrification/denitrification processes. Models can represent N limitation in different ways, including using N to scale photosynthesis [105,106], downscaling potential gross primary productivity (GPP) to reflect N availability [38,84,85,107], defining a C cost of N uptake [108], optimizing N allocation for leaf processes [109], or adapting a flexible C:N ratio for N allocation [106]. Nitrogen uptake is scaled depending on demand, based on stoichiometry (see Section 5.2) and availability, where photosynthesis and decomposition may be downscaled. More recent developments have led to the development of the Fixation and Uptake of Nitrogen (FUN) model [108], which expands N acquisition to include processes of passive uptake, active uptake, re-translocation, and symbiotic N fixation through a C cost. Other advances in the Community Land Model (CLM) expand N uptake to include methods using Michaelis-Menten equations [106,110] or equilibrium chemistry approximation [111,112]. Additional processes such as abiotic (i.e., mineral surface) competition for soil nutrients (Nutrient COMpetition model, [111]) are anticipated for the next generation of the CLM and the Accelerated Climate Model for Energy (ACME).
5.2. Allometry
ESMs rarely include dynamic responses to changes in resource availability. The most common allocation approach assigns C to each plant component (usually leaf, stem, and root) via fixed ratios that vary with plant functional type (PFT), but not spatially or temporally [38,85,88,95,107,113,114,115]. For models that include N (and less often P), N uptake plays a strong role in governing C assimilation and drives competition between plants and decomposers. In these cases, C allocation requires an additional constraint of fulfilling C:N:P ratios. Nitrogen uptake is controlled by plant demand and decomposition requirements determined from fixed C partitioning and C:N stoichiometry. When N and P demands to maintain stoichiometric C:N:P are unmet, photosynthesis is downscaled [116,117]. This simple approach works well for regions that have relatively stable environments, but it does not permit plant plasticity responses to changing nutrients. This method of C allocation has been highlighted as a weakness of these models [118,119] and has resulted in some models swapping to more dynamic allocation schemes that allow responses to stress, as in Friedlingstein et al. [120]. Carbon allocation in two ESMs (CTEM [86,87] and ORCHIDEE [94]) is governed by the most limiting resource; light limitation results in more biomass being allocated to the stem, and water and N limitations result in more biomass being allocated to the roots. However, because N is not explicit in these models, the N limitation is parameterized.
The simple representation of plant allometry in ESMs can have a significant influence on biogeochemistry since the allocation of biomass in the form of C to woody versus non-woody (or photosynthetic versus non-photosynthetic) plant components drives biogeochemical cycling, the quality and quantity of litter, and the duration of C storage. Our ability to model C uptake and subsequent storage therefore depends on including C allocation relationships and their responses under changes in resource availability.
5.3. Roots
Another component of ESMs that is oversimplified is root forms and function. For example, most root algorithms in ESMs consist of a fixed rooting depth and distribution [93]. This constrains water uptake to the root zone (weighted by effective root fraction in each layer) and does not allow differential water uptake from soil layers due to changes in root kinetics and morphology, nor does it allow changes in root distribution or depth to increase the water available to the plant. Water uptake is generally a function of plant demand (for evapotranspiration needs), root distribution, and soil water content [121]. The most common model approach to determine water limitation is to calculate a water stress scalar, which can be a function of soil water matric potential, matric potential when stomata are open or closed, or soil water content [121]. That water stress scalar is then multiplied by physiological variables such as transpiration, stomatal conductance, photosynthetic capacity, and maintenance respiration to capture water stress on plants. One approach to improve the evapotranspiration in the CLM is to add root hydraulic redistribution [122], which can transport water either up or down the soil column. Hydraulic redistribution does not influence root growth or distribution, but it does enhance water uptake by shifting the water distribution in the root zone.
Nitrogen uptake (when included) is often less complex in ESMs; most N uptake is from a bulk N mineralization pool that depends on supply and demand rather than root biomass or profile, although some recent work has focused on uptake as a function of root biomass [106]. Essentially, plants are given every opportunity to extract all available N necessary for growth. Although this allows an implicit metric for allowing roots an opportunity to adjust their uptake, it is not realistic and lacks feedbacks on the biogeochemistry from changes in the root profile.
Recently, studies focused on modeling root growth to maximize various plant traits have been conducted. For example, Sivandran and Bras [123] used optimization techniques for root growth in the TIN-based Real-Time Integrated Basin Simulator coupled to the Vegetation Generator for Interactive Evolution (tRIBS + VEGGIE) model to maximize plant transpiration. The improved root parameterizations allowed more root C to be allocated to soil layers with high soil moisture, increased water uptake, and decreased plant water stress. Another study by McMurtrie et al. [124] altered the vertical distribution of root mass to maximize N uptake. However, the underlying limitation in all studies is that they focus on maximizing only one limiting resource, namely, water or N. This limits the studies’ relevance to environments that share those resource limitations so they may not reflect the future state of those regions or be suitable for global application.
6. Models Development Priorities
While ESMs have advanced considerably, there are several processes that still require attention if we are to capture the effects from N deposition and drought as shown in the pathways in Figure 1 and Figure 2. Future model development should focus on the following goals (see Table 2 for summary): (1) allow a more flexible coupling of C and N in models; (2) integrate a dynamic C partitioning algorithm; (3) improve the structure and function of roots; (4) include succession and age classes; (5) include intra-plant competition; and (6) develop methods of trait-based modeling rather than the traditional PFT approach. Some of these developments are being addressed, at least in part, for the next generation of models. For others, support needs to come from observation and the empirical community to develop robust methodology for inclusion in models.
6.1. Flexibility of C:N Coupling in Models
Given the increase in foliar N under increased N, allowing adaptations in the stoichiometry of C and N would improve model responses [118]. The main impact of this will be to decrease C:N in leaves, driving increases in productivity and changes to soil and litter N content that would be present under increasing N deposition. Changes in C:N ratios occur not just over the lifecycle of the plant, but also in response to changes in nutrient availability, which are not captured in models. Changes in resource availability will result in changes to plant C allocation and partitioning. Furthermore, plant responses will be limited under fixed C:N ratios, which in turn drive changes in belowground biogeochemistry and ultimately C uptake and storage. However, our understanding of how C:N ratios change with plant age and resource status is limited. Therefore, more observations of how C:N is partitioned within the plant over optimal conditions and under climate change are needed.
6.2. Adaptive Dynamics Approach to C Allocation
Possibly the most challenging improvement that would likely provide the most benefit for models predicting C uptake would be to address how C is partitioned within the plant and how that varies over time and with changes to the environment. According to Franklin et al. [114], the difficulty is that C allocation is the result of several processes, which makes a mechanistic approach to represent C allocation difficult. The most robust approach for modeling allocation is adaptive dynamics that include evolution strategies that can emerge through population dynamics and can result in an evolutionary stable strategy [114]. This type of implementation will be difficult in an ESM, given the complexity of integrating dynamic C allocation and the computational demand required to run at high resolution over global scales. However, optimization techniques such as those proposed by Lynch [131] may be an alternative; vegetation may be able to take on the form of optimal response through a cost-benefit approach or game theoretical optimization [114]. This approach allows a flexible plant response to local and regional environmental conditions and nutrient availability. In addition, this functionality would allow PFTs in models to optimize nutrient capture by focusing resources to improve the uptake of the most limiting resources. In a scenario with N deposition and drought, a PFT would be able to alter its root:shoot ratio to increase allocation to roots to increase water uptake or to the stem to increase light competition.
6.3. Improving Form and Function of Roots
In order to allow ecosystems to respond to changes in the environment such as climate change, roots must be allowed to adapt to the heterogeneity of water and nutrients in the soil. Improving the form and function of roots will allow PFTs the opportunity to respond to the heterogeneity of resources, thus increasing the potential for N and/or water uptake. Increasing root depth alone is not sufficient to improve modeled water uptake in water-stressed systems; models also need to consider root distribution, plasticity, and hydraulics [132]. At a minimum, root profiles should include time-varying structures as leaves and stems. Although models tend to have homogeneous soil horizontally (in a grid cell), the vertical structure of resources is dynamic. Allowing roots to proliferate in soil layers where resources are concentrated gives PFTs the chance to adapt to changes in environment and can further change the vertical distribution of C and N. Baker et al. [63] improved the modeled Net Ecosystem Exchange cycle in the Simple Biosphere Model compared with observations in the Amazon by adding hydraulic redistribution and soil depth to 10 m. Other elements of root systems that should be included in models are root order and classification (which will differ in respiration, uptake, turnover, and storage capacity), root phenology and turnover, and resource uptake response to heterogeneity of resources [133]. Warren et al. [121] provided additional suggestions for improving root representation in models, including scaling root function across temporal and spatial scales and including root traits that inform function and hydraulic redistribution. Although most work in ESM development has focused on improving aboveground productivity, some effort has targeted belowground activities. For example, recent work used optimization techniques to modify root growth to maximize plant transpiration [123] and N uptake [124]. Arora and Boer [93] developed a method to represent root distribution as a function of root biomass, which is a proxy for plant age, to allow root depth to increase when plants are young but grow horizontally when plants are mature. This was implemented in the Integrated Science Assessment Model [125] to capture seasonal leaf area index and GPP in northern high-latitude ecosystems. These methods should be expanded to optimize the most limiting resource, rather than focusing on only one limiting resource.
6.4. Succession
Most models do not consider succession or stand age, despite the evidence that old-growth forests do not respond as strongly as young trees to short-term changes in soil moisture or N availability [134]. This results in a homogeneous distribution of vegetation rather than the heterogeneous plant cover that occurs due to the variation of soils and climate across landscapes. This also limits a model’s ability to capture disturbance and recovery events, which are major drivers of C and nutrient cycling. Including succession could help models capture different responses to N and drought stress as a result of tree age by, for example, simulating the higher mortality of young trees. A method to capture succession was implemented by Fisher et al. [135] in the Community Land Model by separating vegetation into cohorts of age, PFT, and height; this method was tested against deciduous–evergreen forest boundaries [126]. These techniques can capture the variability of individual PFTs within a grid cell, adding large-scale heterogeneity in plant distribution, and can be the first step toward a more trait-based modeling approach (see Section 6.6).
6.5. Competition
In many LSMs, plants compete for resources such as light, water, and nutrients within a grid cell based on their weight on the grid cell. In these cases, the competition is between different PFTs and not within a PFT. Dynamic vegetation models can force one PFT to replace another when climate conditions are favorable [136]. A few LSMs have adopted Lotka-Volterra predator/prey equations [137] to represent colonization rates. However, no model currently implements a game-theoretic approach to the competition for resources that can produce an overabundance of biomass for resource uptake to outcompete neighbors. This method has the potential to improve estimates of plant production, respond to changing resources, and lead to an evolutional stable strategy [138]. PFTs would have the opportunity to alter biomass allocation (i.e., shifts in the root:shoot ratio) to outcompete neighbors for limiting resources, which would lead to shifts in productivity and vegetation species distribution. Although it is hindered by the current model’s resolution, as continuously expanding computational capabilities move toward high-resolution models [139], game theory is an achievable target.
6.6. Trait-Based Modeling
The current PFT approach to modeling vegetation is limited to feedback that results from changes in species distribution since most models assume that attributes within a PFT do not change with climate [140]. To capture the adaptation and evolution of vegetation, the concept of trait-based modeling was introduced by Lavorel and Garnier [141]. Trait-based modeling links plant traits that act as plant responses to the environment with plant traits that represent the effect of plants on the ecosystem in order to capture the plant assemblage of a region. A trait can be defined as a feature that describes the morphology, physiology, or phenology characteristics of an individual, which can vary across an environmental gradient [142]. Trait based modeling allows dynamic vegetation to move beyond simple climate based temperature controls on vegetation existence and establishment. Therefore, species distribution would be a response to N and drought in the ecosystem. This approach has been used to improve two dynamic vegetation models (aDGVM, [128]; JeDi-DGVM, [129]). The Jedi-DGVM outperformed other leading dynamic vegetation models for Leaf Area Index (LAI), NPP, CO2 seasonality, C fluxes, and, in some regions, C stocks [129]. Incorporating plant traits in the CSIRO Atmospheric Biosphere Land Exchange (CABLE) model improved the biogeographical distribution of major forests that have multiple dominant PFTs [130]. Using plant traits to capture whole-plant hydraulics has even been suggested to improve C and water use responses to drought [143]. The benefit of trait-based modeling is that the environment acts as a filter for trait composition, analogous to evolutional selection processes [140], and is not limited by climate and geography. This is particularly important considering that McNeil et al. [144] found that species’ foliar N responses to N deposition were dependent on two main plant traits, leaf mass area and shade tolerance. Matheny et al. [136] also suggested several plant traits that could influence the availability of water to a plant that span across leaves, stems, and roots. The challenge is finding data to support the choice of traits and how different traits co-vary.
7. Conclusions
We have provided a review of the impacts of N and drought on ecosystems and a list of future model recommendations that serves to address the missing processes needed in LSMs to capture those interactions. While this list is extensive, it is by no means exhaustive. Our suggestions target two co-occurring stressors, but additional stresses will also be present that should be considered such as warming, elevated CO2, and herbivory. For example, herbivory could reduce the N benefits of elevated productivity from consumption [145] or be a source of N deposition. Effects from elevated CO2 can vary; CO2 fertilization can help mitigate extreme heat and drought [146], or fertilization effects on productivity may be eliminated during drought and nutrient limitation [147]. These additional impacts on ecosystems are important, considering that CO2 fertilization currently plays a larger role than N deposition in increasing productivity [148].
Another example is the interactions that exist between N availability, drought, and air temperature. It is thought that forest productivity should increase with temperature at the global scale, but droughts often associated with increases in temperature have lowered the NPP potential in some regions [149]. For instance, tropical forests can increase Net Ecosystem Exchange (NEE) with modest increases in air temperature, and drought effects could be ameliorated with increases in atmospheric CO2 [150]. The interactions of temperature with drought and N can also affect the surface properties of forests beyond evapotranspiration. Drought sensitive forests (deciduous forests) tend to have a higher surface albedo than drought resistant ones (e.g., conifer forests), affecting the regional energy balance [151]. Further, increases in temperature can increase decomposition and N mineralization rates in the absence of drought, but N mineralization will not respond to temperature if moisture is the limiting factor [152]. These biophysical properties are important in understanding the global net effect of drought and N deposition on biosphere climate interactions [153] and are not the focus of this work. It is important to note that, in most elevated temperature ecosystem experiments, drought is a secondary effect caused by elevated temperature treatments [154,155], making it difficult to distinguish primary from integrated responses of forests to drought and temperature on biogeochemical and biophysical processes.
This study only focused on two stressors to ecosystems, N and drought. Although some of the identified processes targeted by this manuscript may help models to capture impacts from other climate-related effects, additional ecosystem processes will need to be addressed in the future. As the community integrates additional processes in models to capture vegetation responses to N inputs, we can revisit the resulting C uptake to evaluate the sink capacity of the terrestrial surface. However, special care should be taken when implementing additional parameters and processes into models, particularly so that models are not over-parameterized and so that the resulting C cycle response is not constrained by incomplete observational data. In the near term, model sensitivity studies can be used to determine the most sensitive parameters and processes that drive changes in C and nutrient cycles. More research on the structural uncertainty of models can provide insight on those processes that have strong feedbacks or introduce instability. Finally, when rigorous model testing through benchmarking is complete, we can focus on additional questions related to the fate of C in the land.
Improving estimates of the global terrestrial C sink is a priority for ESM development. In order to improve the predictions and reduce uncertainty, model development should focus on the processes that will be affected by multiple co-occurring stressors such as N deposition and drought. We have suggested avenues of model improvement that are possible in the near future with hopes that future generations of models can benefit and capture the response to increased N availability and drought.
Supplementary Materials
The following are available online at www.mdpi.com/1999-4907/8/8/267/s1, Table S1, Summary of previous studies that have investigated N deposition, drought, and the interactions of ecosystems.
Acknowledgments
Argonne National Laboratory’s work was supported under U.S. Department of Energy contract DE-AC02-06CH11357. B.D. is supported by the Accelerated Climate Modeling for Energy (ACME) program, sponsored by the US Department of Energy, Office of Science, Office of Biological and Environmental Research. We also thank US Forest Service grant 15-JV-11242302-038. We thank Hormoz BassiriRad, Henry Howe, and Eduardo Dias de Oliveira for their discussion and comments on early versions of the draft. We also thank the anonymous reviewers for helpful suggestions on organization and flow, which greatly improved the manuscript.
Author Contributions
B.D. conceived the review concept, collected and analyzed the relevant information. B.D. and M.G. wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Friedlingstein, P.; Meinshausen, M.; Arora, V.K.; Jones, C.J.; Anav, A.; Liddicoat, S.K.; Knutti, R. Uncertainties in CMIP5 climate projections due to carbon cycle feedbacks. J. Clim. 2014, 27, 511–526. [Google Scholar] [CrossRef]
- Seneviratne, S.I.; Nicholls, N.; Easterling, D.; Goodess, C.M.; Kanae, S.; Kossin, J.; Luo, Y.; Marengo, J.; McInnes, K.; Rahimi, M.; et al. Changes in climate extremes and their impacts on the natural physical environment. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; Field, C.B., Barros, V., Stocker, T.F., Qin, D., Dokken, D.J., Ebi, K.L., Mastrandrea, M.D., Mach, K.J., Plattner, G.-K., Allen, S.K., et al., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 109–230. [Google Scholar]
- Vitousek, P.M.; Aber, J.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, G.D. Human alteration of the global nitrogen cycle: Causes and consequences. Issues Ecol. 1997, 1, 1–17. [Google Scholar]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Dentener, F.; Drevet, J.; Lamarque, J.F.; Bey, I.; Eickhout, B.; Fiore, A.M.; Hauglustaine, D.; Horowitz, L.W.; Krol, M.; Kulshrestha, U.C.; et al. Nitrogen and sulfur deposition on regional and global scales: A multimodel evaluation. Glob. Biogeochem. Cycles 2006, 20. [Google Scholar] [CrossRef] [Green Version]
- Aber, J.D.; Goodale, C.L.; Ollinger, S.V.; Smith, M.-L.; Magill, A.H.; Martin, M.E.; Hallett, R.A.; Stoddard, J.L. Is Nitrogen deposition altering the nitrogen status of northeastern forests? Bioscience 2003, 53, 375–389. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardo, L.H.; Fenn, M.E.; Goodale, C.L.; Geiser, L.H.; Driscoll, C.T.; Allen, E.B.; Baron, J.S.; Bobbink, R.; Bowman, W.D.; Clark, C.M.; et al. Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States. Ecol. Appl. 2011, 21, 3049–3082. [Google Scholar] [CrossRef]
- Bobbink, R.; Roelofs, J.G.M. Nitrogen critical loads for natural and semi-natural ecosystems: The empirical approach. Water Air. Soil Pollut. 1995, 85, 2413–2418. [Google Scholar] [CrossRef]
- Melillo, J.; Steudler, P.; Aber, J.; Newkirk, K.; Lux, H.; Bowles, F.; Catricala, C.; Magill, A.; Ahrens, T.; Morrisseau, S. Soil warming and carbon-cycle feedbacks to the climate system. Science 2002, 298, 2173–2176. [Google Scholar] [CrossRef] [PubMed]
- Pastor, J.; Post, W. Response of northern forests to CO2-induced climate change. Nature 1988, 334, 55–58. [Google Scholar] [CrossRef]
- Peterjohn, W.T.; Melillo, J.M.; Steudler, P.A.; Newkirk, K.M.; Bowles, F.P.; Aber, J.D. Responses of trace gas fluxes and N availability to experimentailly elevated soil temperatures. Ecol. Appl. 1994, 4, 617–625. [Google Scholar] [CrossRef]
- Clark, C.M.; Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 2008, 451, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Simkin, S.M.; Allen, E.B.; Bowman, W.D.; Clark, C.M.; Belnap, J.; Brooks, M.L.; Cade, B.S.; Collins, S.L.; Geiser, L.H.; Gilliam, F.S.; et al. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 4086–4091. [Google Scholar] [CrossRef] [PubMed]
- BassiriRad, H.; Lussenhop, J.F.; Sehtiya, H.L.; Borden, K.K. Nitrogen deposition potentially contributes to oak regeneration failure in the Midwestern temperate forests of the USA. Oecologia 2015, 177, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Minocha, R.; Turlapati, S.A.; Long, S.; McDowell, W.H.; Minocha, S.C. Long-term trends of changes in pine and oak foliar nitrogen metabolism in response to chronic nitrogen amendments at Harvard Forest, MA. Tree Physiol. 2015, 35, 894–909. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.; Lohse, K.A.; Hall, S.J. The globalization of nitrogen deposition: Consequences for terrestrial ecosystems. Ambio 2002, 31, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.T.; Yoh, M.; Mo, J.M.; Gundersen, P.; Zhou, G.Y. Response of nitrogen leaching to nitrogen deposition in disturbed and mature forests of southern China. Pedosphere 2009, 19, 111–120. [Google Scholar] [CrossRef]
- Dise, N.B.; Wright, R.F. Nitrogen leaching from European forests in relation to nitrogen deposition. For. Ecol. Manag. 1995, 71, 153–161. [Google Scholar] [CrossRef]
- Broeckx, L.S.; Verlinden, M.S.; Berhongaray, G.; Zona, D.; Fichot, R.; Ceulemans, R. The effect of a dry spring on seasonal carbon allocation and vegetation dynamics in a poplar bioenergy plantation. GCB Bioenergy 2014, 6, 473–487. [Google Scholar] [CrossRef]
- Hertel, D.; Strecker, T.; Müller-Haubold, H.; Leuschner, C. Fine root biomass and dynamics in beech forests across a precipitation gradient—Is optimal resource partitioning theory applicable to water-limited mature trees? J. Ecol. 2013, 101, 1183–1200. [Google Scholar] [CrossRef]
- Martin-Stpaul, N.K.; Limousin, J.M.; Vogt-Schilb, H.; Rodríguez-Calcerrada, J.; Rambal, S.; Longepierre, D.; Misson, L. The temporal response to drought in a Mediterranean evergreen tree: Comparing a regional precipitation gradient and a throughfall exclusion experiment. Glob. Chang. Biol. 2013, 19, 2413–2426. [Google Scholar] [CrossRef] [PubMed]
- Padilla, F.M.; Miranda, J.D.; Jorquera, M.J.; Pugnaire, F.I. Variability in amount and frequency of water supply affects roots but not growth of arid shrubs. Plant Ecol. 2009, 204, 261–270. [Google Scholar] [CrossRef]
- Friedrich, U.; von Oheimb, G.; Kriebitzsch, W.U.; Schleßelmann, K.; Weber, M.S.; Härdtle, W. Nitrogen deposition increases susceptibility to drought—Experimental evidence with the perennial grass Molinia caerulea (L.) Moench. Plant Soil 2012, 353, 59–71. [Google Scholar] [CrossRef]
- Kinugasa, T.; Tsunekawa, A.; Shinoda, M. Increasing nitrogen deposition enhances post-drought recovery of grassland productivity in the Mongolian steppe. Oecologia 2012, 170, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Boisier, J.P.; Ciais, P.; Ducharne, A.; Guimberteau, M. Projected strengthening of Amazonian dry season by constrained climate model simulations. Nat. Clim. Chang. 2015, 5, 656–660. [Google Scholar] [CrossRef]
- Cook, B.I.; Ault, T.R.; Smerdon, J.E. Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci. Adv. 2015, 1, e1400082. [Google Scholar] [CrossRef] [PubMed]
- Hanson, P.J.; Weltzin, J.F. Drought disturbance from climate change: Repsonse of United States forests. Sci. Total Enivron. 2000, 262, 205–220. [Google Scholar] [CrossRef]
- Phillips, O.L.; Aragão, L.E.O.C.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; López-González, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A.; et al. Drought sensitivity of the Amazon rainforest. Science 2009, 323, 1344–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Cui, X.; Wang, H.; Chen, A. Drought: The most important physical stress of terrestrial ecosystems. Acta Ecol. Sin. 2014, 34, 179–183. [Google Scholar] [CrossRef]
- Thornton, P.E.; Doney, S.C.; Lindsay, K.; Moore, J.K.; Mahowald, N.; Randerson, J.T.; Fung, I.; Lamarque, J.F.; Feddema, J.J.; Lee, Y.H. Carbon-nitrogen interactions regulate climate-carbon cycle feedbacks: Results from an atmosphere-ocean general circulation model. Biogeosciences 2009, 6, 2099–2120. [Google Scholar] [CrossRef]
- Koirala, S.; Yeh, P.J.F.; Hirabayashi, Y.; Kanae, S.; Oki, T. Global-scale land surface hydrologic modeling with the representation of water table dynamics. J. Geophys. Res. Atmos. 2014, 119, 75–89. [Google Scholar] [CrossRef]
- Yan, B.; Dickinson, R.E. Modeling hydraulic redistribution and ecosystem response to droughts over the Amazon Basin using Community Land Model 4.0 (CLM4). J. Geophys. Res. Biogeosci. 2014, 119, 2130–2143. [Google Scholar] [CrossRef]
- Leuzinger, S.; Quinn Thomas, R. How do we improve Earth system models? Integrating Earth system models, ecosystem models, experiments and long-term data. New Phytol. 2011, 191, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Ukkola, A.M.; De Kauwe, M.G.; Pitman, A.J.; Best, M.J.; Abramowitz, G.; Haverd, V.; Decker, M.; Haughton, N. Land surface models systematically overestimate the intensity, duration and magnitude of seasonal-scale evaporative droughts. Environ. Res. Lett. 2016, 11. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Zhou, S.X.; Medlyn, B.E.; Pitman, A.J.; Wang, Y.P.; Duursma, R.A.; Prentice, I.C. Do land surface models need to include differential plant species responses to drought? Examining model predictions across a mesic-xeric gradient in Europe. Biogeosciences 2015, 12, 7503–7518. [Google Scholar] [CrossRef]
- Fisher, J.B.; Badgley, G.; Blyth, E. Global nutrient limitation in terrestrial vegetation. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Gerber, S.; Hedin, L.O.; Oppenheimer, M.; Pacala, S.W.; Shevliakova, E. Nitrogen cycling and feedbacks in a global dynamic land model. Glob. Biogeochem. Cycles 2010, 24. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationship in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.A.; Howarth, R.W. Nutrients in synergy. Nature 2007, 449, 1000–1001. [Google Scholar] [CrossRef] [PubMed]
- Bala, G.; Devaraju, N.; Chaturvedi, R.K.; Caldeira, K.; Nemani, R. Nitrogen deposition: How important is it for global terrestrial carbon uptake? Biogeosciences 2013, 10, 7147–7160. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Burton, A.J.; Zak, D.R.; Talhelm, A.F. Simulated chronic nitrogen deposition increases carbon storage in northern temperate forests. Glob. Chang. Biol. 2008, 14, 142–153. [Google Scholar]
- Elvir, J.A.; Rustad, L.; Wiersrna, G.B.; Fernandez, I.; White, A.S.; White, G.J. Eleven-year response of foliar chemistry to chronic nitrogen and sulfur additions at the Bear Brook Watershed in Maine. Can. J. For. Res. 2005, 35, 1402–1410. [Google Scholar]
- Boggs, J.L.; Mcnulty, S.G.; Gavazzi, M.J.; Myers, J.M. Tree growth, foliar chemistry, and nitrogen cycling across a nitrogen deposition gradient in southern Applachian deciduous forests. Can. J. For. Res. 2005, 35, 1901–1913. [Google Scholar]
- Pitcairn, C.E.R.; Leith, I.D.; Fowler, D.; Hargreaves, K.J.; Moghaddam, M.; Kennedy, V.H.; Granat, L. Foliar nitrogen as an indicator of nitrogen deposition and critical loads exceedance on a European scale. Water Air. Soil Pollut. 2001, 130, 1037–1042. [Google Scholar]
- Fleischer, K.; Rebel, K.T.; Van Der Molen, M.K.; Erisman, J.W.; Wassen, M.J.; Van Loon, E.E.; Montagnani, L.; Gough, C.M.; Herbst, M.; Janssens, I.A.; et al. The contribution of nitrogen deposition to the photosynthetic capacity of forests. Glob. Biogeochem. Cycles 2013, 27, 187–199. [Google Scholar]
- Talhelm, A.F.; Pregitzer, K.S.; Burton, A.J. No evidence that chronic nitrogen additions increase photosynthesis in mature sugar maple forests. Ecol. Appl. 2011, 21, 2413–2424. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Maughan, M.W.; Sun, J.; Feng, X.; Miguez, F.; Lee, D.; Dietze, M.C. Impact of nitrogen allocation on growth and photosynthesis of Miscanthus (Miscanthus × giganteus). GCB Bioenergy 2012, 4, 688–697. [Google Scholar]
- Quinn Thomas, R.; Canham, C.D.; Weathers, K.C.; Goodale, C.L. Increased tree carbon storage in response to nitrogen deposition in the US. Nat. Geosci. 2010, 3, 13–17. [Google Scholar]
- De Vries, W.; Du, E.; Butterbach-Bahl, K. Short and long-term impacts of nitrogen deposition on carbon sequestration by forest ecosystems. Curr. Opin. Environ. Sustain. 2014, 9–10, 90–104. [Google Scholar]
- Xia, J.; Wan, S. Global response patterns of terrestrial plant species to nitrogen addition. New Phytol. 2008, 179, 428–439. [Google Scholar] [PubMed]
- Ibáñez, I.; Zak, D.R.; Burton, A.J.; Pregitzer, K.S. Chronic nitrogen deposition alters tree allometric relationships: Implications for biomass production and carbon storage. Ecol. Appl. 2016, 26, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Fang, J. Weak growth response to nitrogen deposition in an old-growth boreal forest. Ecosphere 2014, 5. [Google Scholar] [CrossRef]
- Verma, P.; Sagar, R.; Verma, H.; Verma, P.; Singh, D.K. Changes in species composition, diversity and biomass of herbaceous plant traits due to N amendment in a dry tropical environment of India. J. Plant Ecol. 2014, 8, 321–332. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Bergeron, Y. Influence of environmental variability on root dynamics in northern forests. CRC Crit. Rev. Plant Sci. 2009, 28, 179–197. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Eissenstat, D.M.; Lovett, G.M.; Bowden, R.D.; Rustad, L.E.; Driscoll, C.T. Root stress and nitrogen deposition: Consequences and research priorities. New Phytol. 2013, 197, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhang, W.; Chen, H.; Mo, J. Impacts of nitrogen deposition on soil nitrogen cycle in forest ecosystems: A review. Acta Ecol. Sin. 2015, 35, 35–43. [Google Scholar] [CrossRef]
- Bobbink, R.; Lamers, L.P.M. Effects of increased nitrogen deposition. In Air Pollution and Plant Life; Bell, J.N.B., Treshow, M., Eds.; John Wiley and Sons: Chichester, UK, 2002; pp. 201–235. [Google Scholar]
- Grulke, N.E.; Minnich, R.A.; Paine, T.; Riggan, P. Air pollution increases forest susceptibility to wildfires: A case study for the San Bernardino Mountains in southern California. In Wildland Fires and Air Pollution; Bytnerowicz, A., Arbaugh, M., Riebau, A., Anderson, C., Eds.; Elsiver: New York, NY, USA, 2009; pp. 365–403. [Google Scholar]
- Mcdowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Weißhuhn, K.; Auge, H.; Prati, D. Geographic variation in the response to drought in nine grassland species. Basic Appl. Ecol. 2011, 12, 21–28. [Google Scholar] [CrossRef]
- Klein, T.; Cohen, S.; Yakir, D. Hydraulic adjustments underlying drought resistance of Pinus halepensis. Tree Physiol. 2011, 31, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Baker, I.T.; Prihodko, L.; Denning, A.S.; Goulden, M.; Miller, S.; Da Rocha, H.R. Seasonal drought stress in the amazon: Reconciling models and observations. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef]
- Sala, A.; Piper, F.; Hoch, G. Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytol. 2010, 186, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.A.; Pineda, M.; de la Cruz Jiménez, J.; Vergara, M.F.; Rao, I.M. Contrasting strategies to cope with drought conditions by two tropical forage C4 grasses. AoB Plants 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Rowland, L.; da Costa, A.C.L.; Galbraith, D.R.; Oliveira, R.S.; Binks, O.J.; Oliveira, A.A.R.; Pullen, A.M.; Doughty, C.E.; Metcalfe, D.B.; Vasconcelos, S.S.; et al. Death from drought in tropical forests is triggered by hydraulics not carbon starvation. Nature 2015, 528, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Lotter, D.; Valentine, A.J.; Archer Van Garderen, E.; Tadross, M. Physiological responses of a fynbos legume, Aspalathus linearis to drought stress. S. Afr. J. Bot. 2014, 94, 218–223. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Belowground drought response of European beech: Fine root biomass and carbon partitioning in 14 mature stands across a precipitation gradient. Glob. Chang. Biol. 2008, 14, 2081–2095. [Google Scholar] [CrossRef]
- Hanson, P.J.; Tschaplinski, T.J.; Wullschleger, S.D.; Todd, D.E.; Augé, R.M. The Resilience of Upland-Oak Forest Canopy Trees to Chronic and Acute Precipitation Maniuplations; Buckley, D.S., Clatterbuck, W.K., Eds.; U.S. Department of Agriculture: Morgantown, WV, USA; Southern Research Station: Asheville, NC, USA, 2007. [Google Scholar]
- Prieto, I.; Armas, C.; Pugnaire, F.I. Water release through plant roots: New insights into its consequences at the plant and ecosystem level. New Phytol. 2012, 193, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Neumann, R.B.; Cardon, Z.G. The magnitude of hydraulic redistribution by plant roots: A review and synthesis of empirical and modeling studies. New Phytol. 2012, 194, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Kleczewski, N.M.; Herms, D.A.; Bonello, P. Nutrient and water availability alter belowground patterns of biomass allocation, carbon partitioning, and ectomycorrhizal abundance in Betula nigra. Trees 2012, 26, 525–533. [Google Scholar] [CrossRef]
- Meyer-Grünefeldt, M.; Friedrich, U.; Klotz, M.; Von Oheimb, G.; Härdtle, W. Nitrogen deposition and drought events have non-additive effects on plant growth—Evidence from greenhouse experiments. Plant Biosyst. 2013, 149, 424–432. [Google Scholar] [CrossRef]
- Meyer-Grünefeldt, M.; Calvo, L.; Marcos, E.; Von Oheimb, G.; Härdtle, W. Impacts of drought and nitrogen addition on Calluna heathlands differ with plant life-history stage. J. Ecol. 2015, 103, 1141–1152. [Google Scholar] [CrossRef]
- Liu, X.; Fan, Y.; Long, J.; Wei, R.; Kjelgren, R.; Gong, C.; Zhao, J. Effects of soil water and nitrogen availability on photosynthesis and water use efficiency of Robinia pseudoacacia seedlings. J. Environ. Sci. 2013, 25, 585–595. [Google Scholar] [CrossRef]
- Wang, M.; Shi, S.; Lin, F.; Hao, Z.; Jiang, P.; Dai, G. Effects of soil water and nitrogen on growth and photosynthetic response of Manchurian Ash (Fraxinus mandshurica) seedlings in northeastern China. PLoS ONE 2012, 7, e30754. [Google Scholar] [CrossRef] [PubMed]
- Palátová, E. Effect of increased nitrogen depositions and drought stress on the development of young Norway spruce Picea abies (L.) Karst. stands. Dendrobiology 2004, 51, 41–45. [Google Scholar]
- Zhou, X.; Zhang, Y.; Ji, X.; Downing, A.; Serpe, M. Combined effects of nitrogen deposition and water stress on growth and physiological responses of two annual desert plants in northwestern China. Environ. Exp. Bot. 2011, 74, 1–8. [Google Scholar] [CrossRef]
- Palátová, E. Effect of increased nitrogen depositions and drought stress on the development of scots pine (Pinus sylvestris L.)—II. Root system response. J. For. Sci. 2002, 48, 237–247. [Google Scholar]
- Albuquerque, W.G.; Severino, L.S.; Beltrao, N.E.M.; Azevedo, C.A.V.; Da Silva Filho, J.L. Growth and biomass allocation of Jutropha curcas plants as influenced by nitrogen under different soil moisture regimes. Res. Crop. 2013, 14, 928–934. [Google Scholar]
- Verburg, P.S.J.; Kapitzke, S.E.; Stevenson, B.A.; Bisiaux, M. Carbon allocation in Larrea tridentata plant-soil systems as affected by elevated soil moisture and N availability. Plant Soil 2014, 378, 227–238. [Google Scholar] [CrossRef]
- Oleson, K.W.; Lawrence, D.M.; Gordon, B.; Drewniak, B.; Huang, M.; Koven, C.D.; Levis, S.; Li, F.; Riley, W.J.; Subin, Z.M.; et al. Technical Description of Version 4.5 of the Community Land Model (CLM); National Center for Atmospheric Research: Boulder, CO, USA, 2013. [Google Scholar]
- Wang, Y.P.; Law, R.M.; Pak, B. A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere. Biogeosciences 2010, 7, 2261–2282. [Google Scholar] [CrossRef] [Green Version]
- Arora, V.K.; Boer, G.J. A parameterization of leaf phenology for the terrestrial ecosystem component of climate models. Glob. Chang. Biol. 2005, 11, 39–59. [Google Scholar] [CrossRef]
- Arora, V.K.; Boer, G.J.; Christian, J.R.; Curry, C.L.; Denman, K.L.; Zahariev, K.; Flato, G.M.; Scinocca, J.F.; Merryfield, W.J.; Lee, W.G. The effect of terrestrial photosynthesis down regulation on the twentieth-century carbon budget simulated with the CCCma Earth System Model. J. Clim. 2009, 22, 6066–6088. [Google Scholar] [CrossRef]
- Shevliakova, E.; Pacala, S.W.; Malyshev, S.; Hurtt, G.C.; Milly, P.C.D.; Caspersen, J.P.; Sentman, L.T.; Fisk, J.P.; Wirth, C.; Crevoisier, C. Carbon cycling under 300 years of land use change: Importance of the secondary vegetation sink. Glob. Biogeochem. Cycles 2009, 23. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Collatz, G.J.; Ball, J.T.; Grivet, C.; Berry, J.A. Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: A model that includes a laminar boundary layer. Agric. For. Meteorol. 1991, 54, 107–136. [Google Scholar] [CrossRef]
- Zeng, X. Global vegetation root distribution for land modeling. J. Hydrometeorol. 2001, 2, 525–530. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Arora, V.K.; Boer, G.J. A representation of variable root distribution in dynamic vegetation models. Earth Interact. 2003, 7, 1–19. [Google Scholar] [CrossRef]
- Krinner, G.; Viovy, N.; de Noblet-Ducoudré, N.; Ogée, J.; Polcher, J.; Friedlingstein, P.; Ciais, P.; Sitch, S.; Prentice, I.C. A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Zaehle, S.; Friend, A.D. Carbon and nitrogen cycle dynamics in the O-CN land surface model: Model description, site-scale evaluation, and sensitivity to parameter estimates. Glob. Biogeochem. Cycles 2010, 24. [Google Scholar] [CrossRef]
- Best, M.J.; Pryor, M.; Clark, D.B.; Rooney, G.G.; Essery, R.L.H.; Ménard, C.B.; Edwards, J.M.; Hendry, M.A.; Porson, A.; Gedney, N.; et al. The Joint UK Land Environment Simulator (JULES), model description—Part 1: Energy and water fluxes. Geosci. Model Dev. 2011, 4, 677–699. [Google Scholar] [CrossRef]
- Clark, D.B.; Mercado, L.M.; Sitch, S.; Jones, C.D.; Gedney, N.; Best, M.J.; Pryor, M.; Rooney, G.G.; Essery, R.L.H.; Blyth, E.; et al. The Joint UK Land Environment Simulator (JULES), model description—Part 2: Carbon fluxes and vegetation dynamics. Geosci. Model Dev. 2011, 4, 701–722. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.; Prentice, I.C.; Sykes, M.T. Representation of vegetation dynamics in the modelling of terrestrial ecosystems: Comparing two contrasting approaches within European climate space. Glob. Ecol. Biogeogr. 2001, 10, 621–637. [Google Scholar] [CrossRef]
- Smith, B.; Wärlind, D.; Arneth, A.; Hickler, T.; Leadley, P.; Siltberg, J.; Zaehle, S. Implications of incorporating N cycling and N limitations on primary production in an individual-based dynamic vegetation model. Biogeosciences 2014, 11, 2027–2054. [Google Scholar] [CrossRef] [Green Version]
- Friend, A.D.; Kiang, N.Y. Land surface model development for the GISS GCM: Effects of improved canopy physiology on simulated climate. J. Clim. 2005, 18, 2883–2902. [Google Scholar] [CrossRef]
- Collatz, G.J.; Ribas-Carbo, M.; Berry, J.A. Coupled photosynthesis-stomatal conductance model for leaves of C4 plants. Aust. J. Plant Physiol. 1992, 19, 519–538. [Google Scholar] [CrossRef]
- Zaehle, S.; Jones, C.D.; Houlton, B.; Lamarque, J.F.; Robertson, E. Nitrogen availability reduces CMIP5 projections of twenty-first-century land carbon uptake. J. Clim. 2015, 28, 2494–2511. [Google Scholar] [CrossRef]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Su, S.; Currie, W.S.; Dukes, J.S.; Finzi, A.; Hartwig, U.; Hungate, B.; McMurtrie, R.E.; Oren, R.; Parton, W.J.; et al. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience 2004, 54, 731–739. [Google Scholar] [CrossRef]
- Zaehle, S.; Medlyn, B.E.; De Kauwe, M.G.; Walker, A.P.; Dietze, M.C.; Hickler, T.; Luo, Y.; Wang, Y.-P.; El-masri, B.; Thornton, P.; et al. Evaluation of 11 terrestrial carbon-nitrogen cycle models against observations from two temperate free-air CO2 enrichment studies. New Phytol. 2014, 202, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, B.; Riley, W.J.; Koven, C.D.; Mu, M.; Randerson, J.T. Representing leaf and root physiological traits in CLM improves global carbon and nitrogen cycling predictions. J. Adv. Model. Earth Syst. 2016, 8, 598–613. [Google Scholar] [CrossRef]
- Parida, B.R. The Influence of Plant Nitrogen Availability on the Global Carbon Cycle and NO Emissions. Ph.D. Thesis, University of Hamburg, Hamburg, Germany, 2011. [Google Scholar]
- Fisher, J.B.; Sitch, S.; Malhi, Y.; Fisher, R.A.; Huntingford, C.; Tan, S.Y. Carbon cost of plant nitrogen acquisition: A mechanistic, globally applicable model of plant nitrogen uptake, retranslocation, and fixation. Glob. Biogeochem. Cycles 2010, 24. [Google Scholar] [CrossRef]
- Ali, A.A.; Xu, C.; Rogers, A.; McDowell, N.G.; Medlyn, B.E.; Fisher, R.A.; Wullschleger, S.D.; Reich, P.B.; Vrugt, J.A.; Bauerle, W.L.; et al. Global-scale environmental control of plant photosynthetic capacity. Ecol. Appl. 2015, 25, 2349–2365. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.Q.; Zaehle, S.; Templer, P.H.; Goodale, C.L. Global patterns of nitrogen limitation: Confronting two global biogeochemical models with observations. Glob. Chang. Biol. 2013, 19, 2986–2998. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Riley, W.J.; Tang, J. A new theory of plant-microbe nutrient competition resolves inconsistencies between observations and model predictions. Ecol. Appl. 2017, 27, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.Y.; Riley, W.J. A total quasi-steady-state formulation of substrate uptake kinetics in complex networks and an example application to microbial litter decomposition. Biogeosciences 2013, 10, 8329–8351. [Google Scholar] [CrossRef]
- Tjiputra, J.F.; Roelandt, C.; Bentsen, M.; Lawrence, D.M.; Lorentzen, T.; Schwinger, J.; Seland, Ø.; Heinze, C. Evaluation of the carbon cycle components in the Norwegian Earth System Model (NorESM). Geosci. Model Dev. 2013, 6, 301–325. [Google Scholar] [CrossRef]
- Franklin, O.; Johansson, J.; Dewar, R.C.; Dieckmann, U.; McMurtrie, R.E.; Brännström, A.K.; Dybzinski, R. Modeling carbon allocation in trees: A search for principles. Tree Physiol. 2012, 32, 648–666. [Google Scholar] [CrossRef] [PubMed]
- Goll, D.S.; Brovkin, V.; Parida, B.R.; Reick, C.H.; Kattge, J.; Reich, P.B.; Van Bodegom, P.M.; Niinemets, U. Nutrient limitation reduces land carbon uptake in simulations with a model of combined carbon, nitrogen and phosphorus cycling. Biogeosciences 2012, 9, 3547–3569. [Google Scholar] [CrossRef] [Green Version]
- Zaehle, S.; Dalmonech, D. Carbon-nitrogen interactions on land at global scales: Current understanding in modelling climate biosphere feedbacks. Curr. Opin. Environ. Sustain. 2011, 3, 311–320. [Google Scholar] [CrossRef]
- Thornton, P.E.; Lamarque, J.F.; Rosenbloom, N.A.; Mahowald, N.M. Influence of carbon-nitrogen cycle coupling on land model response to CO2 fertilization and climate variability. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Medlyn, B.E.; Zaehle, S.; De Kauwe, M.G.; Walker, A.P.; Dietze, M.C.; Hanson, P.J.; Hickler, T.; Jain, A.K.; Luo, Y.; Parton, W.; et al. Using ecosystem experiments to improve vegetation models. Nat. Clim. Chang. 2015, 5, 528–534. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Medlyn, B.E.; Zaehle, S.; Walker, A.P.; Dietze, M.C.; Wang, Y.P.; Luo, Y.; Jain, A.K.; El-Masri, B.; Hickler, T.; et al. Where does the carbon go? A model-data intercomparison of vegetation carbon allocation and turnover processes at two temperate forest free-air CO2 enrichment sites. New Phytol. 2014, 203, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Friedlingstein, P.; Joel, G.; Field, C.B.; Fung, I.Y. Toward an allocation scheme for global terrestrial carbon models. Glob. Chang. Biol. 1999, 5, 755–770. [Google Scholar] [CrossRef]
- Warren, J.M.; Hanson, P.J.; Iversen, C.M.; Kumar, J.; Walker, A.P.; Wullschleger, S.D. Root structural and functional dynamics in terrestrial biosphere models—Evaluation and recommendations. New Phytol. 2015, 205, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Riley, W.J.; Niu, J. Incorporating root hydraulic redistribution in CLM4.5: Effects on predicted site and global evapotranspiration, soil moisture, and water storage. J. Adv. Model. Earth Syst. 2015, 7, 1828–1848. [Google Scholar] [CrossRef]
- Sivandran, G.; Bras, R.L. Dynamic root distributions in ecohydrological modeling: A case study at Walnut Gulch Experimental Watershed. Water Resour. Res. 2013, 49, 3292–3305. [Google Scholar] [CrossRef]
- McMurtrie, R.E.; Iversen, C.M.; Dewar, R.C.; Medlyn, B.E.; Näsholm, T.; Pepper, D.A.; Norby, R.J. Plant root distributions and nitrogen uptake predicted by a hypothesis of optimal root foraging. Ecol. Evol. 2012, 2, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- El Masri, B.; Shu, S.; Jain, A.K. Implementation of a dynamic rooting depth and phenology into a land surface model: Evaluation of carbon, water, and energy fluxes in the high latitude ecosystems. Agric. For. Meteorol. 2015, 211–212, 85–99. [Google Scholar] [CrossRef]
- Fisher, R.A.; Muszala, S.; Verteinstein, M.; Lawrence, P.; Xu, C.; McDowell, N.G.; Knox, R.G.; Koven, C.; Holm, J.; Rogers, B.M.; et al. Taking off the training wheels: The properties of a dynamic vegetation model without climate envelopes, CLM4.5(ED). Geosci. Model Dev. 2015, 8, 3593–3619. [Google Scholar] [CrossRef]
- Mcnickle, G.G.; Gonzalez-meler, M.A.; Lynch, D.J.; Baltzer, J.L.; Brown, J.S. The world’s biomes and primary production as a triple tragedy of the commons foraging game played among plants. Proc. R. Soc. B 2016, 283. [Google Scholar] [CrossRef] [PubMed]
- Scheiter, S.; Langan, L.; Higgins, S.I. Next-generation dynamic global vegetation models: Learning from community ecology. New Phytol. 2013, 198, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Pavlick, R.; Drewry, D.T.; Bohn, K.; Reu, B.; Kleidon, A. The Jena Diversity-Dynamic Global Vegetation Model (JeDi-DGVM): A diverse approach to representing terrestrial biogeography and biogeochemistry based on plant functional trade-offs. Biogeosciences 2013, 10, 4137–4177. [Google Scholar] [CrossRef]
- Lu, X.; Wang, Y.-P.; Wright, I.J.; Reich, P.B.; Shi, Z.; Dai, Y. Incorporation of plant traits in a land surface model helps explain the global biogeographical distribution of major forest functional types. Glob. Ecol. Biogeogr. 2016, 26, 304–317. [Google Scholar] [CrossRef]
- Lynch, D.J. Impacts of Fine-Roots on Terrestrial Net Primary Productivity and Soil Nutrient Cycling. Ph.D. Thesis, University of Illinois at Chicago, Chicago, IL, USA, 2015. [Google Scholar]
- Nippert, J.B.; Holdo, R.M. Challenging the maximum rooting depth paradigm in grasslands and savannas. Funct. Ecol. 2015, 29, 739–745. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Lucash, M.S.; McCormack, M.L.; Sivandran, G. Improving the representation of roots in terrestrial models. Ecol. Model. 2014, 291, 193–204. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.; Mcdowell, N.; Purves, D.; Moorcroft, P.; Sitch, S.; Cox, P.; Huntingford, C.; Meir, P.; Woodward, F.I. Assessing uncertainties in a second-generation dynamic vegetation model caused by ecological scale limitations. New Phytol. 2010, 187, 666–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quillet, A.; Peng, C.H.; Garneau, M. Toward dynamic global vegetation models for simulating vegetation-climate interactions and feedbacks: Recent developments, limitations, and future challenges. Environ. Rev. 2010, 18, 333–353. [Google Scholar] [CrossRef]
- Arora, V.K.; Boer, G.J. Simulating competition and coexistence between plant functional types in a dynamic vegetation model. Earth Interact. 2006, 10, 1–30. [Google Scholar] [CrossRef]
- McNickle, G.G.; Brown, J.S. Evolutionarily stable strategies for nutrient foraging and below-ground competition in plants. Evol. Ecol. Res. 2012, 14, 667–687. [Google Scholar]
- Department of Energy. Available online: https://climatemodeling.science.energy.gov/sites/default/files/ACME_Overview_Brochure.pdf (accessed on 2 May 2017).
- Van Bodegom, P.M.; Douma, J.C.; Witte, J.P.M.; Ordoñez, J.C.; Bartholomeus, R.P.; Aerts, R. Going beyond limitations of plant functional types when predicting global ecosystem-atmosphere fluxes: Exploring the merits of traits-based approaches. Glob. Ecol. Biogeogr. 2012, 21, 625–636. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Garnier, E.; Navas, M.L. A trait-based approach to comparative functional plant ecology: Concepts, methods and applications for agroecology. A review. Agron. Sustain. Dev. 2012, 32, 365–399. [Google Scholar] [CrossRef]
- Matheny, A.M.; Mirfenderesgi, G.; Bohrer, G. Trait-based representation of hydrological functional properties of plants in weather and ecosystem models. Plant Divers. 2016, 39, 1–12. [Google Scholar] [CrossRef]
- McNeil, B.E.; Read, J.M.; Driscoll, C.T. Foliar nitrogen responses to elevated atmospheric nitrogen deposition in nine temperate forest canopy species. Environ. Sci. Technol. 2007, 41, 5191–5197. [Google Scholar] [CrossRef] [PubMed]
- Throop, H.L. Nitrogen deposition and herbivory affect biomass production and allocation in an annual plant. Oikos 2005, 111, 91–100. [Google Scholar] [CrossRef]
- Roy, J.; Picon-Cochard, C.; Augusti, A.; Benot, M.-L.; Thiery, L.; Darsonville, O.; Landais, D.; Piel, C.; Defossez, M.; Devidal, S.; et al. Elevated CO2 maintains grassland net carbon uptake under a future heat and drought extreme. Proc. Natl. Acad. Sci. USA 2016, 113, 6224–6229. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Hobbie, S.E.; Lee, T.D. Plant growth enhancement by elevated CO2 eliminated by joint water and nitrogen limitation. Nat. Geosci. 2014, 7, 920–924. [Google Scholar] [CrossRef]
- Los, S.O. Analysis of trends in fused AVHRR and MODIS NDVI data for 1982-2006: Indication for a CO2 fertilization effect in global vegetation. Glob. Biogeochem. Cycles 2013, 27, 318–330. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Meler, M.A.; Rucks, J.S.; Aubanell, G. Mechanistic insights on the responses of plant and ecosystem gas exchange to global environmental change: Lessons from Biosphere 2. Plant Sci. 2014, 226, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.B.; Randerson, J.T.; Canadell, J.G.; Anderson, R.G.; Avissar, R.; Baldocchi, D.D.; Bonan, G.B.; Caldeira, K.; Diffenbaugh, N.S.; Field, C.B. Protecting climate with forests. Environ. Res. Lett. 2008, 3. [Google Scholar] [CrossRef]
- Hyvönen, R.; Ågren, G.I.; Linder, S.; Persson, T.; Cotrufo, M.F.; Ekblad, A.; Freeman, M.; Grelle, A.; Janssens, I.A.; Jarvis, P.G.; et al. The likely impact of elevated CO2, nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: A literature review. New Phytol. 2007, 173, 463–480. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.G.; Canadell, J.G.; Randerson, J.T.; Jackson, R.B.; Hungate, B.A.; Baldocchi, D.D.; Ban-Weiss, G.A.; Bonan, G.B.; Caldeira, K.; Cao, L.; et al. Biophysical considerations in forestry for climate protection. Front. Ecol. Environ. 2011, 9, 174–182. [Google Scholar] [CrossRef]
- Wu, Z.; Dijkstra, P.; Koch, G.W.; Peñuelas, J.; Hungate, B.A. Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef]
- Gonzalez-Meler, M.A.; Silva, L.B.C.; Dias-De-Oliveira, E.; Flower, C.E.; Martinez, C.A. Experimental air warming of a Stylosanthes capitata, vogel dominated tropical pasture affects soil respiration and nitrogen dynamics. Front. Plant Sci. 2017, 8, 46. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of N deposition on ecosystems, based on the concepts of Bobbink et al. [7]. Up arrows represent an increase, and down arrows represent a decrease. Three main pathways exist: changes in foliar or leaf N, changes to biomass partitioning, and increases in biomass N. Changes in leaf or plant N generally lead to a positive feedback by increasing N in the litter pool. Changes in the partitioning of biomass can lead to changes in competition for resources that will ultimately affect species distribution. In general, the impacts of N deposition occur over short timescales (<5 years), while changes to species composition and disturbance occur over longer timescales [7].
Figure 1.
Effects of N deposition on ecosystems, based on the concepts of Bobbink et al. [7]. Up arrows represent an increase, and down arrows represent a decrease. Three main pathways exist: changes in foliar or leaf N, changes to biomass partitioning, and increases in biomass N. Changes in leaf or plant N generally lead to a positive feedback by increasing N in the litter pool. Changes in the partitioning of biomass can lead to changes in competition for resources that will ultimately affect species distribution. In general, the impacts of N deposition occur over short timescales (<5 years), while changes to species composition and disturbance occur over longer timescales [7].
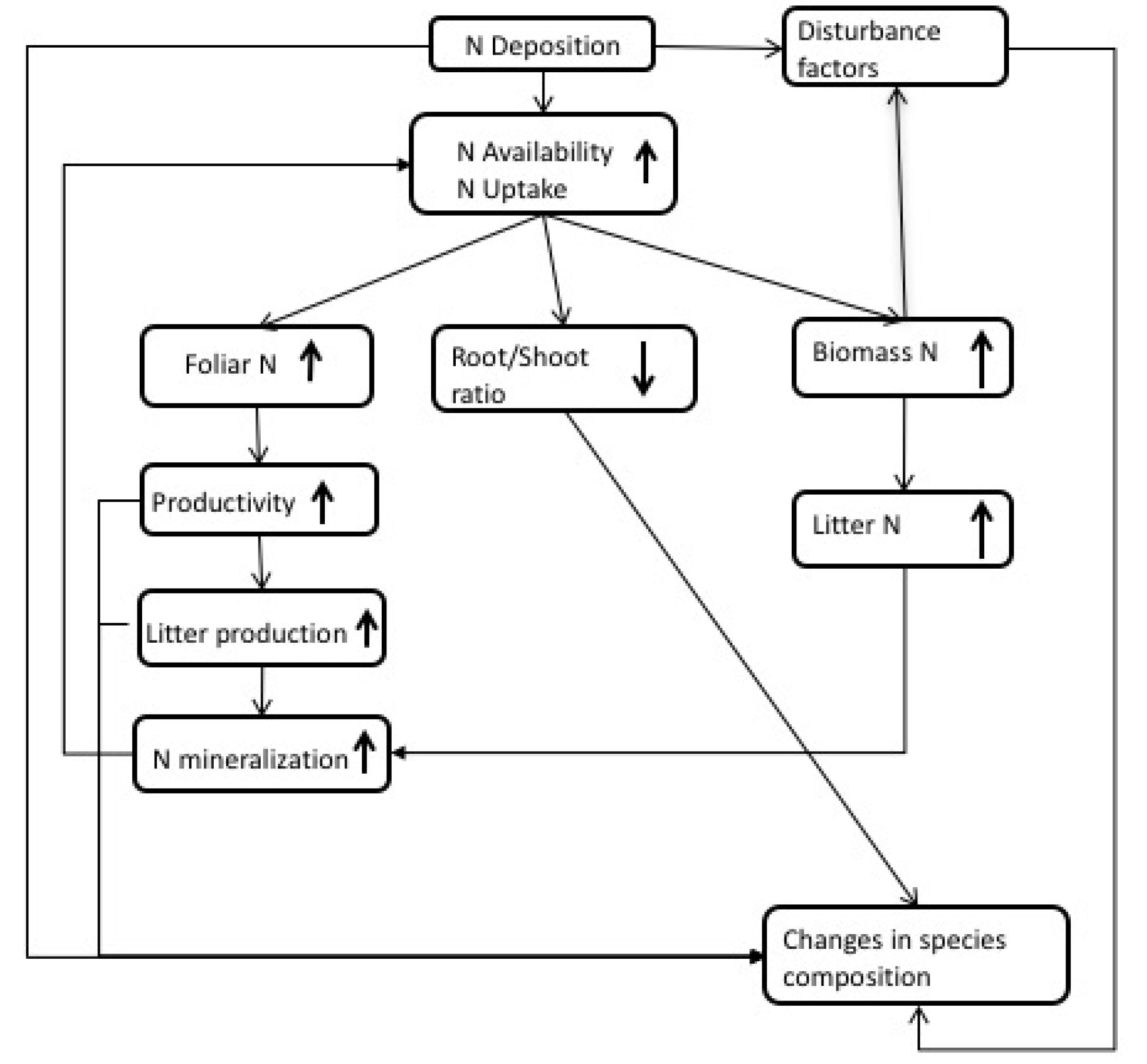
Figure 2.
Effects of drought on ecosystems. Up arrows represent an increase, and down arrows represent a decrease. Four main pathways exist: changes in C availability, increases in N limitation, changes to C mobilization and transport, and changes to biomass partitioning (i.e., root:shoot). Changes in C availability and N limitation both lead to decreases in photosynthesis, which has a positive feedback on N limitation when less biomass is contributing N to litter pools. Changes in C availability and a lack of C mobilization can lead to death from carbon starvation. Several pathways result in altered species composition from shifts in nutrient availability and competition. In general, the impacts from drought occur over short timescales (<5 years), while changes to species composition and disturbance occur over longer timescales.
Figure 2.
Effects of drought on ecosystems. Up arrows represent an increase, and down arrows represent a decrease. Four main pathways exist: changes in C availability, increases in N limitation, changes to C mobilization and transport, and changes to biomass partitioning (i.e., root:shoot). Changes in C availability and N limitation both lead to decreases in photosynthesis, which has a positive feedback on N limitation when less biomass is contributing N to litter pools. Changes in C availability and a lack of C mobilization can lead to death from carbon starvation. Several pathways result in altered species composition from shifts in nutrient availability and competition. In general, the impacts from drought occur over short timescales (<5 years), while changes to species composition and disturbance occur over longer timescales.
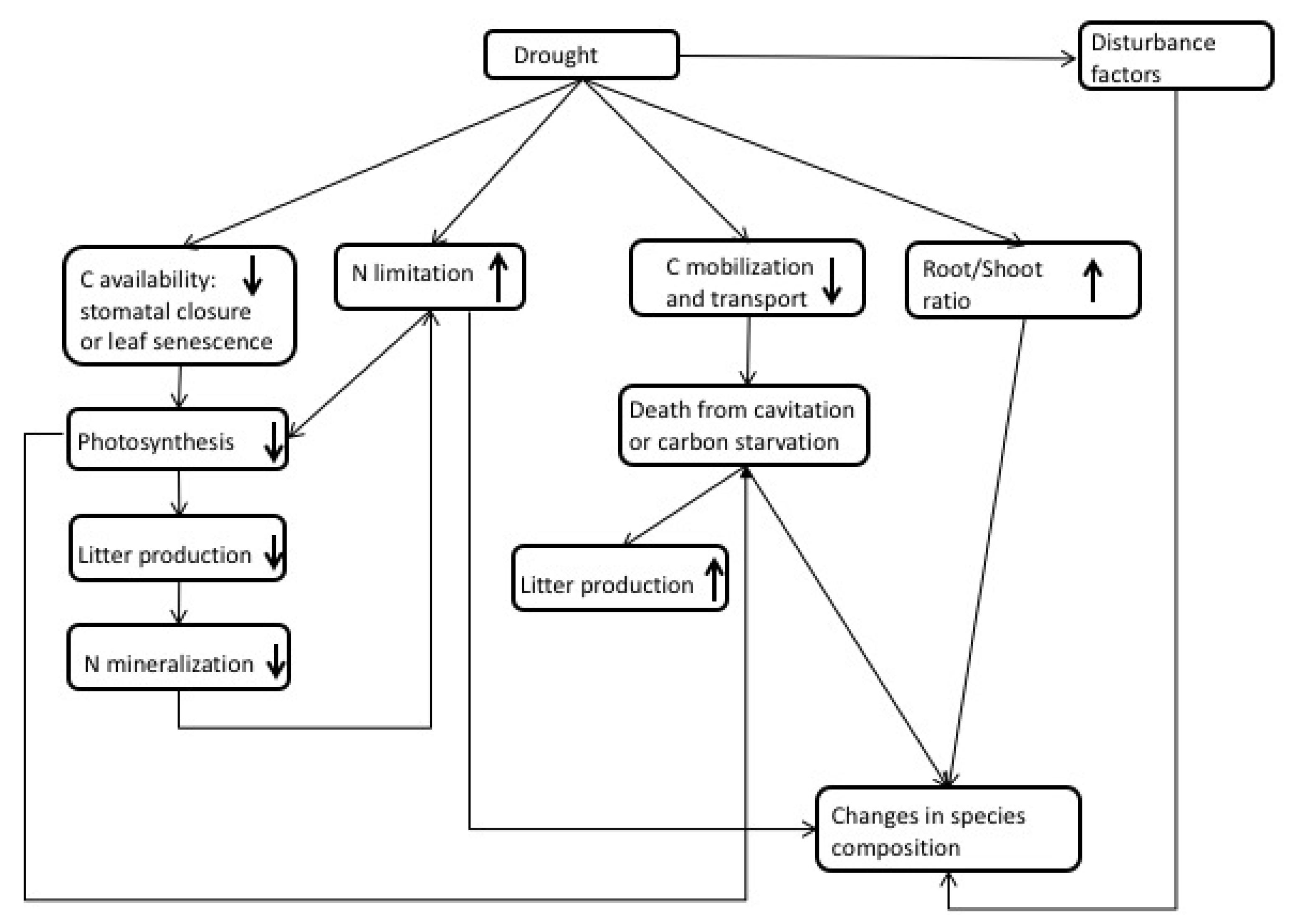

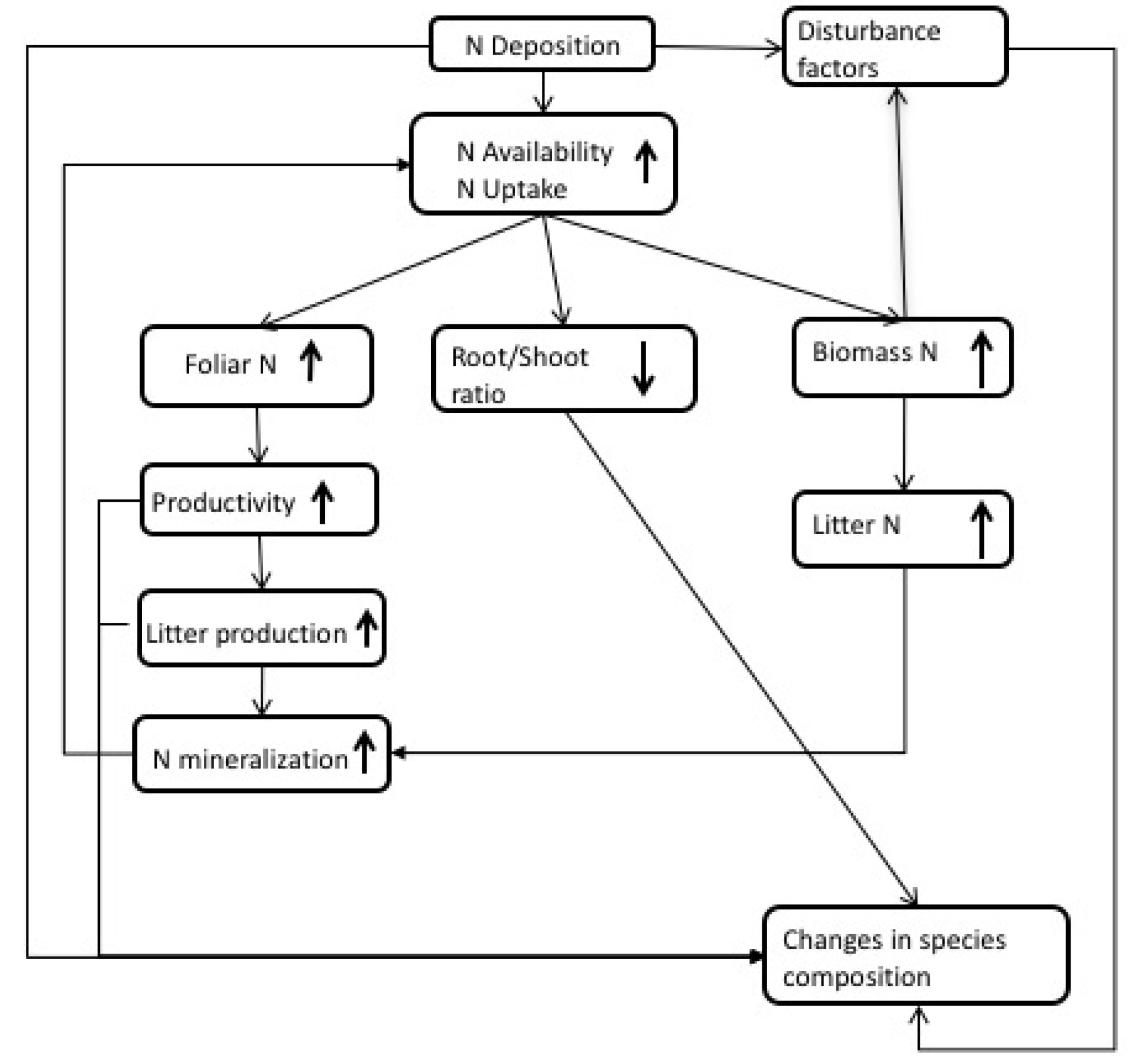{kind=link}
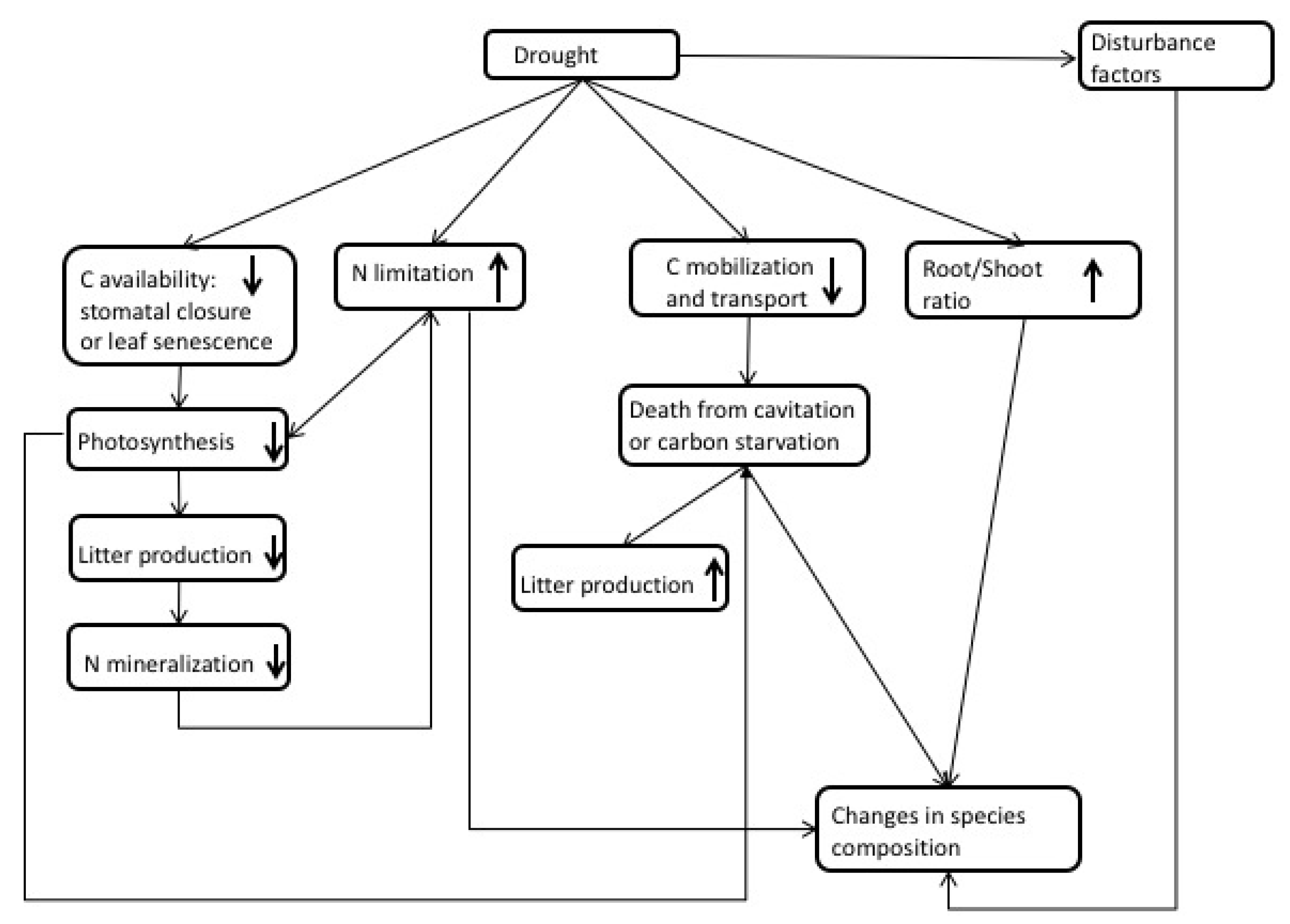{kind=link}
Table 1.
Representation of key model features from a subset of terrestrial ecosystem models.
| Process | Model | |||
| CLM4.5 | CABLE | CTEM | LM3 | |
| Reference | [84] | [85] | [86,87] | [38,88] |
| Time step | 30 min to one hour | 30 min | 30 min to one day | 30 min |
| Plant Functional Type (PFTs) | 14 natural and two generic crop types | 15 natural and one crop type | seven natural and two crop types (C3 and C4) | five natural |
| Dynamic vegetation | Dependent on climate or prescribed | NA | Dependent on climate or prescribed | Dependent on climate and light |
| Plant C | ||||
| Photosynthesis | [89,90] | [89] | [89,90] | [89,90] |
| Phenology | Evergreen, stress deciduous, seasonal deciduous, and crop | Biome dependent, four states, input from remote sensing | Four leaf states: maximum growth, normal growth, leaf fall, and dormancy | Drought and cold deciduous seasonal |
| Allocation | Fixed fraction | Fixed fraction | Dependent on light, water, phenological status | Functional balance to maintain root-to-shoot ratio |
| Plant N | ||||
| Uptake | Dependent on N pool size, plant demand | Dependent on N pool size, plant demand | NA | Michaelis-Menten kinetics, dependent on N pool size and root biomass; priority given to immobilization |
| Fixation | Function of Net Primary Productivity (NPP) | External input | NA | Dependent on plant N demand, NPP, and light availability; C cost paid for biological nitrogen fixation (BNF) |
| Stoichiometry (C:N) | Flexible (within 0.8 N:C) | Fixed (PFT dependent) | NA | Fixed (PFT dependent) |
| Plant water | ||||
| Uptake | Dependent on plant demand, root profile, and soil matric potential | Dependent on plant demand, root fraction, and soil water content | Dependent on soil moisture content | NA |
| Root architecture | Double exponential for water uptake [91]; single exponential for soil C/N cycling [92] | Exponential [86] | Prescribed maximum rooting depth, root distribution dependent on time and PFT [93] | NA |
| Process | Model | |||
| ORCHIDEE | O-CN | JULES | LPJ-GUESS | |
| Reference | [94] | [94,95] | [96,97] | [98,99] |
| Time step | 30 min to one day | 30 min to one day | 30 min to one day | 1 day |
| PFTs | 10 natural and two agricultural grasses | 10 natural and two agricultural grasses | 5 natural | 11 natural |
| Dynamic vegetation | Dependent on climate, stand structure, and light | Dependent on climate, stand structure, and light | Dependent on NPP and tree-shrub-grass hierarchy from the Lotka-Volterra competition approach | Dependent on climate, stand structure, light and soil resources, disturbance, and succession |
| Plant C | ||||
| Photosynthesis | [89,90] | [100] | [90,101] | [90,101] |
| Phenology | Drought and cold deciduous seasonal | Drought and cold deciduous seasonal | Cold deciduous | Evergreen, drought, and cold deciduous |
| Allocation | Rule-based response to external limits; dependent on light, water, and N | Pipe model to maintain root-to-shoot ratio | Fixed fraction | Functional balance to maintain root-to-shoot ratio |
| Plant N | ||||
| Uptake | Implicit, dependent on soil humidity and soil temperature | Michaelis-Menten kinetics, dependent on fine root biomass, plant N status, N pool size, and soil temperature | NA | Dependent on N pool size, plant demand, root mass, and soil temperature |
| Fixation | NA | Calculate potential N fixation from evapotranspiration | NA | Calculate potential N fixation from evapotranspiration |
| Stoichiometry (C:N) | Prescribed | Flexible (provided range) | Fixed fraction | Flexible (provided range) |
| Plant water | ||||
| Uptake | Dependent on plant demand, root fraction, and soil water content | Dependent on plant demand, root fraction, and soil water content | Dependent on plant demand, root fraction, and available soil moisture | Dependent on plant demand and soil water in root zone |
| Root architecture | Exponential root profile | Exponential root profile | Double exponential | Two soil layers; more roots in lower layer (except grass) |
Table 2.
Summary of recommendations for model development.
| Recommendation | Description | Impact | Example(s) |
|---|---|---|---|
| Flexibility of CN coupling | • Allows C:N ratios in the leaf to vary with N availability • Dynamic partitioning of N in the plant | • Effects N in the leaf with influences on photosynthesis | Fixation and Uptake of Nitrogen (FUN) model [108] Leaf Utilization of Nitrogen for Assimilation (LUNA) [109] Community Land Model (CLM)/Accelerated Climate Model for Energy (ACME) [106] |
| Adaptive dynamics approach to C partitioning | • Flexibility in C allocation to account for plant plasticity across environmental conditions | • Optimize nutrient uptake • Increase tissue allocation to respond to limiting resource | CLM/ACME [106] |
| Improve form and function of roots | • Time varying root structure (depth and distribution) • Variable root depth, traits, plasticity, and hydraulics that scale across space and time | • Adapt to heterogeneity of water and nutrients in soil • Optimizes below ground resource uptake | Dynamic root depth [125] Maximize N [124] Maximize evapotranspiration (ET) [123] |
| Succession | • Representing age class • Variable growth dynamics and response to stress with age | • Capture disturbance and recovery • Heterogeneity in plant distribution, improved canopy light dynamics | Ecosystem Demography (ED) model [126] |
| Competition | • Inter- and intra-species competition for resources (e.g., light, water, N, etc.) • Allows competition both within and between PFTs | • Alters allocation of resources to outcompete neighbors • Possibly altering productivity or shift vegetation distribution | Triple Tragedy of Commons [127] Competition with consumers [111] |
| Trait-based modeling | • Varying morphology, physiology, or phenology characteristics of individuals across an environmental gradient • Environment acts as filter for trait composition | • Adaptation and evolution of species to environmental conditions • Dynamic vegetation moves beyond simple rules of existence and/or establishment | Adaptive Dynamic Global Vegetation Model (aDGVM) [128] Jena Diversity-Dynamic Global Vegetation Model (JeDi-DGVM) [129] CSIRO Atmospheric Biosphere Land Exchange (CABLE) [130] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Drewniak, B.; Gonzalez-Meler, M.A. Earth System Model Needs for Including the Interactive Representation of Nitrogen Deposition and Drought Effects on Forested Ecosystems. Forests 2017, 8, 267. https://doi.org/10.3390/f8080267
AMA Style
Drewniak B, Gonzalez-Meler MA. Earth System Model Needs for Including the Interactive Representation of Nitrogen Deposition and Drought Effects on Forested Ecosystems. Forests. 2017; 8(8):267. https://doi.org/10.3390/f8080267
Chicago/Turabian StyleDrewniak, Beth, and Miquel A. Gonzalez-Meler. 2017. "Earth System Model Needs for Including the Interactive Representation of Nitrogen Deposition and Drought Effects on Forested Ecosystems" Forests 8, no. 8: 267. https://doi.org/10.3390/f8080267
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.