Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns


1
Departamento de Ciencias de la Vida, Universidad de Alcalá de Henares, E-28805 Madrid, Spain
2
Departamento de Ecología, Facultad de Ciencias, Universidad de Granada, E-18071 Granada, Spain
3
Department of Zoology, University of Granada, E-18071 Granada, Spain
*
Author to whom correspondence should be addressed.
Forests 2017, 8(8), 276; https://doi.org/10.3390/f8080276
Submission received: 10 July 2017
/
Revised: 19 July 2017
/
Accepted: 27 July 2017
/
Published: 31 July 2017
Abstract
:The assisted regeneration of oaks can be achieved through seeding or planting. Whereas direct seeding of acorns has several advantages over planting nursery-grown seedlings, the problem of seed predation by mammals precludes its widespread application. We investigated the potential of diesel as a mammal repellent to prevent the consumption of Holm oak acorns. We tested the effect of submerging acorns in diesel for 0, 6, 12, and 24 h on acorn predation in a field experiment in the Sierra Nevada Natural Park (Granada province, Spain), and the potential effect of the same treatments on seedling emergence and growth in a nursery experiment. The field experiment showed no reduction in acorn predation through diesel. On the other hand, submersion in diesel hindered the germination of one in every four sown acorns regardless of submersion time. Our results are in line with many failed attempts to protect acorns with chemical repellents and suggest that an effective, universally-abundant, inexpensive, and easy-to-apply acorn protector to enhance the success of seeding operations is yet to be found.
1. Introduction
The assisted regeneration of oaks (Quercus spp.) can be achieved through direct seeding of acorns or the planting of nursery-grown seedlings [1,2]. While planted seedlings avoid the initial demographic filters of germination and emergence, transplanted seedlings often suffer great mortality rates, especially under Mediterranean climate conditions [3,4,5]. During initial growth, oaks develop a deep tap root that grants access to ground water. Growth in containers entangles this root or requires its pruning prior to transportation to the field, which yields suboptimal root architectures [6,7]. In this way, planted seedlings face their first summer without proper access to water and soil nutrients and have a low probability of survival [7]. The negative effects of root deformation in those trees that do survive can prevail until adulthood [6]. Direct seeding of acorns, besides allowing the normal development of the root system, provides greater flexibility in terms of the timing and the location of sowing [8], as well as important economic advantages [9], as compared to planting. However, many species of mammal predators, including rodents, lagomorphs and ungulates, prey on acorns [10,11,12,13,14]. As a consequence, direct sowing of acorns is usually avoided due to the large losses to predators and the uncertainty that it generates despite its many advantages [2,8,15,16].
Methods to protect acorns from predators have been investigated for centuries [17]. Broadly speaking, the available methods can be classified into the management of predator populations and habitats, physical protectors, and chemical repellents. While some recently-invented physical protectors [18,19] offer a solution to acorn predation under some circumstances, their use increases the cost of sowing, thus reducing the important economic advantage of this method. On the other hand, some chemical repellents could be effective [15,20], yet many can produce toxic effects on the environment [20] or on the seeds that they are meant to protect [11,20]. Finding a method to protect acorns from mammals that fulfills the conditions of being effective against predators, not harmful, inexpensive, easy to use, and universally available, thus continues to represent a major unresolved challenge in forest science.
Here, we aim to test the effectiveness of a common and inexpensive substance as a chemical repellent for acorns. In 1990, Bäumler and colleagues [20] published that diesel could represent the most effective protection for acorns among several substances that they tested, yet they did not measure potential diesel effects on plant performance beyond the stage of germination and we are not aware of any recent studies that have addressed the possible mammal repellency of diesel for sowing. To fill this gap and confirm the effectiveness of diesel as a repellent, we performed a field experiment in the Natural Protected Area of Sierra Nevada to assess the rate of acorn consumption in two replicated mammal-exclosure treatments—one with rodents only and one with rodents plus ungulates—and under four levels of diesel application: no submersion and submersion for 6, 12, and 24 h. We also performed a nursery experiment to search for possible effects of the diesel treatment on seedling emergence and growth. Overall, we aim to assess whether diesel—a cheap, easy-to-use, and universally available substance—also provides a positive balance in the trade-off between the effectiveness against mammal predators and potential negative effects on plant emergence and performance. A successful outcome could represent a major advance in forestation practice, favor the paradigm shift from planting seedlings to seeding acorns, and consequently reduce the cost—or increase the scale—of assisted regeneration of oaks.
2. Materials and Methods
We performed a field experiment to test the effectiveness of acorn submersion in diesel as a mammal repellent, and a nursery experiment to search for potential effects of this treatment on seedling emergence and growth.
For both studies, we picked Holm oak (Quercus ilex subsp. ballota (Desf.) Samp.) acorns from trees located near the field study site. The acorns were harvested three to four weeks before the experiments and stored under cold conditions (4 °C) until the time of sowing. Acorns were weighed, and their identity was noted with a white marker on the shell. The day before sowing, acorns from each parent tree were retrieved from cold storage, randomly divided in four groups, and mixed with those of other parent trees. Each group was then assigned to one of the four diesel treatments, consisting of either complete submersion in diesel (bought at a local petrol station) for 6 h, 12 h, or 24 h or no submersion (control). Submersion occurred for all treatments simultaneously, and treated acorns were placed in a wire mesh basket during transportation to the field. The control acorns were always handled by a different person than diesel-treated acorns to avoid their contamination.
2.1. Acorn Predation Experiment
The acorn predation experiment was conducted under field conditions at the Loma de los Panaderos (La Cortijuela Botanical Garden, Sierra Nevada National Park, SE Spain; 37°5′ N, 3°28′ W). Previous studies have documented high rates of acorn predation in this area, both by rodents and wild boars [21,22,23]. We used two fenced plots of ca. 3000 m2 (some 200 m apart from each other) built in 1997 where ungulates were excluded [24], as well as three unfenced plots of similar dimensions located just beside the fenced plots (snow precluded the use of a third exclosure). The exclosures generated two levels of acorn predators: rodents only (inside exclosures), and rodents plus ungulates (outside) [23]. The main rodent species in the area are wood mice (Apodemus sylvaticus) and Mediterranean pine voles (Microtus duodecimcostatus), whereas ungulates include Iberian ibex (Capra pyrenaica) and wild boars (Sus scrofa). The site, located at an elevation of 1800 m, has Mediterranean-type climate with a mean rainfall of 830 L m−2 year−1, a mean temperature of the coldest month (January) of 3.5 °C, and a mean temperature of the hottest month (August) of 21.6 °C. Vegetation in the area is dominated by successional shrubs intermingled with interspaces of bare ground and some scattered trees [25].
On 13 January 2017, we randomly placed 30 feeding stations in each of the five plots and placed four acorns (one per level of diesel) at each station in the corners of a 75 cm × 75 cm square, making a total of 600 monitored acorns (5 plots × 30 stations × 4 acorns). Feeding stations were at least 10 m apart from each other. The acorns were placed on the ground within cavities dug 1 cm deep in the soil to prevent them from rolling away with the wind, and their identity and position was noted. Acorn removal was monitored after 2 and 9 days, noting the state (removed or not) of each acorn. A further revision was made after 55 days, by which point 100% of acorns had disappeared. Snow precluded revising at an intermediate date and we are not certain whether acorns were removed by predators or by the harsh weather conditions before the last revision, so we interpret only the data from the first two revisions (no precipitation happened between sowing and the second revision). We considered removed acorns in these two revisions as depredated because previous studies found that >98% of acorns found by rodents are finally depredated and that there is no secondary caching by the main acorn dispersers in the area [22].
2.2. Nursery Experiment
To assess the potential effects of diesel on seedling emergence and initial development, we conducted a nursery experiment under outdoor conditions in a nursery located 1 km from the University of Granada, at an elevation of 735 m. We sowed 78 acorns of each diesel treatment in 300 mL plastic containers filled with 50% local soil (44.8% sand, 41.8% silt, 13.3% clay, and 0.8% organic matter [23]) and 50% peat (Kekkila Garden Brown 025W; Kekkila Group, Vantaa, Finland). The acorns were placed 1 cm below the soil surface and protected from domestic rodents with a 0.5 cm wire mesh screen. The containers were kept under outdoor conditions and irrigated regularly. Sowing was conducted on 20 December 2016, and the experiment was monitored after 89, 96, 103, 110, 117, 124, and 181 days for seedling emergence. On 20 June 2017 (at the end of the growing season), seedling height was measured and seedlings were harvested, oven-dried at 60 °C until constant weight, and weighed.
2.3. Statistical Analyses
The analyses described below were run in R version 3.3.1 [26].
To assess the effect of treatments on acorn consumption in the field experiment, we fitted logistic regression in mixed models, using the glmer function of the lme4 package [27]. We treated Exclosure (a two-level categorical variable) and Diesel (a four-level categorical variable) as fixed factors, also including the Exclosure × Diesel interaction in the model. In the random effects, we specified plot and feeding station nested within plot.
The data from the nursery experiment were analyzed with mixed effects models too. We tested the effect of Diesel and Acorn Weight—the fixed effects—on the probability of emergence with logistic regression in lme4, as described above; on stem height and dry biomass with linear mixed-effect models using the lme function of the nlme package [28]; and on the time to emergence with mixed-effects Cox models using the coxme function of the coxme package [29]. In the random effects part of each model, we allowed for a random intercept due to parent tree.
We tested the significance of the factors on deletion, using likelihood ratio tests [30]. The significance of the difference between individual factor levels in the diesel treatment was tested by merging two levels into one and performing likelihood ratio tests between the original model and the one with the new factor [31].
3. Results and Discussion
3.1. Acorn Predation
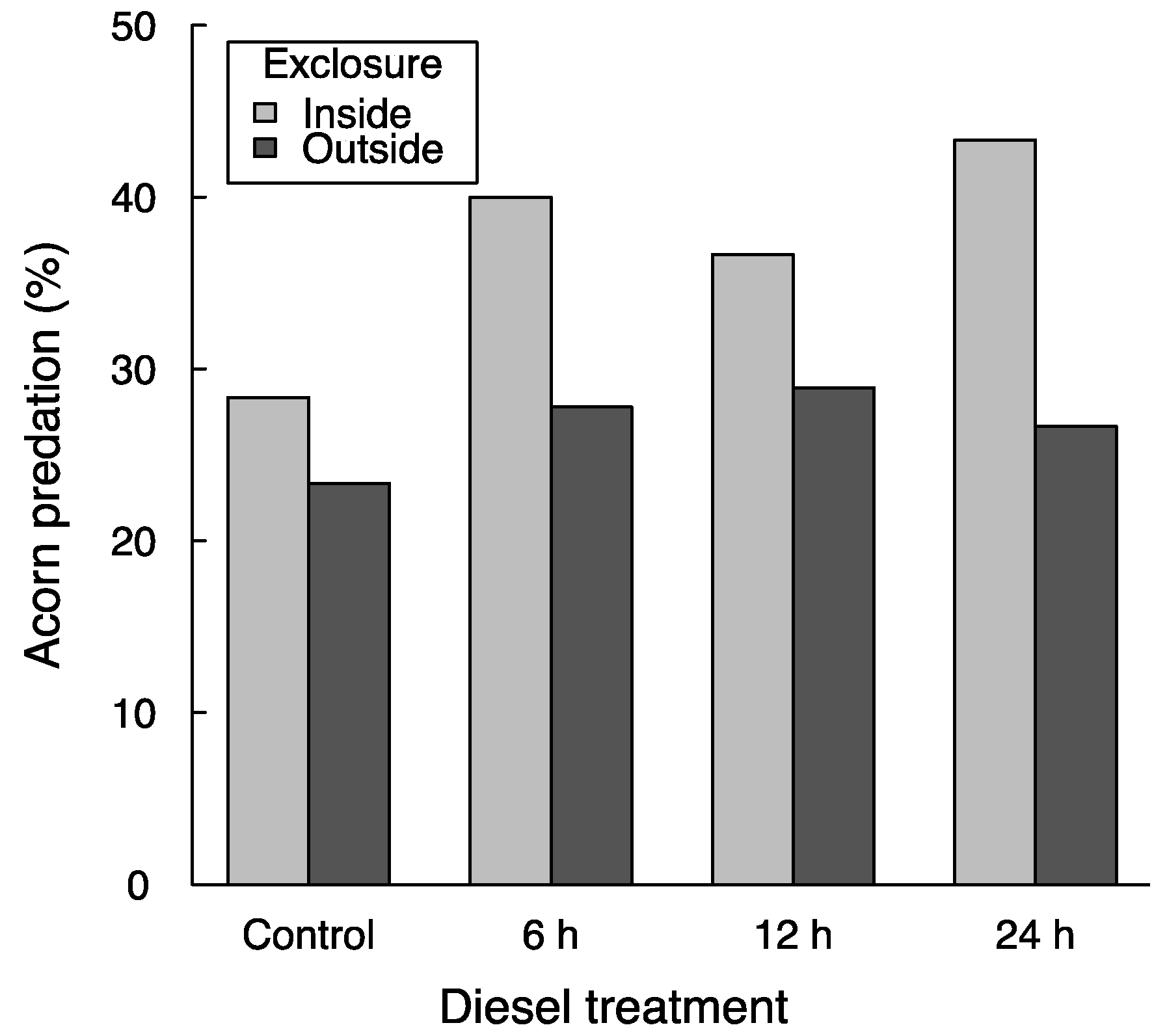
Two days after sowing, 8.8% of the acorns had been consumed, and 30.8% had been consumed after 9 days. In this second revision, 37.1% of the acorns within ungulate exclosures and 26.7% outside exclosures were depredated, although this difference was not significant (χ2 = 0.16, p = 0.69). It has often been described that rodents are more active in areas devoid of ungulates due to the removal of their trampling, competition, and even predation [12,32]. Although this seems to be the case in our study (predation by rodents within the exclosures was greater than that caused by rodents and ungulates combined outside exclosures), spatial variability between exclosure blocks and low replication of the blocks likely prevented the differences from being significant (when fitting the data obviating the spatial structure of the experiment in glm instead of with random effects in glmer, the exclosure effect becomes significant at p < 0.001). In any case, the use of the exclosures in this experiment aimed to assess the potential difference in the effect of diesel under the presence of different acorn predators (i.e., possible exclosure × diesel interactions; see below).
Acorn removal averaged 25.3% without diesel and between 32.0% and 33.3% in the three diesel treatments—yet these differences were not significant (χ2 = 2.71, p = 0.10). Similar to other experiments that have unsuccessfully tested the use of chemical repellents to protect acorns from mammal predators (e.g., [11]), our study shows no reduction in acorn predation by diesel. On the contrary, predation was about 7% greater with diesel than without it, and further monitoring could have led this difference to statistical significance. Thus, rather than deterring mammal predators, it is possible that diesel attracted them, which is in opposition with the study upon which we based our hypotheses [20]. This effect—or lack of an effect—of diesel, however, could vary with the identity of acorn predators, as do several other acorn-protector methods [2,8,19,23]. Instead, our results show that the lack of a reduction in acorn predation by diesel application was consistent across the two levels of acorn predators (Figure 1; diesel × exclosure interaction: χ2 = 0.91, p = 0.34). This suggests that diesel is ineffective in preventing predation under the presence of rodents and of ungulates—the two most commonly-cited guilds of acorn predators that decimate the outcomes of direct seeding operations with Quercus [2,8,11,16]. It should be noted though that differences in predator response to diesel could exist even within each of the two guilds considered in this study—yet our results show that diesel coating was not a general solution to predation, especially given that acorn predators usually include several species acting simultaneously [2,8,10,18,23].
It should be noted that the overall percentage of acorns that were consumed in this study was low compared to other acorn removal experiments performed at the same site in other years [21,22,23], especially given that here acorns were left on the ground instead of buried [10,33]. This can partially be explained because of the cold weather conditions during the study, which likely reduced the activity of rodents [34]. Besides, snow precluded monitoring the experiment for longer, which would most likely have yielded greater acorn removal rates. In any case, given the lack of reduction in predation with diesel and the indication that it might actually have attracted—rather than repelled—consumers under both predator scenarios, we consider further experimentation with this substance futile.
3.2. Seedling Emergence and Growth
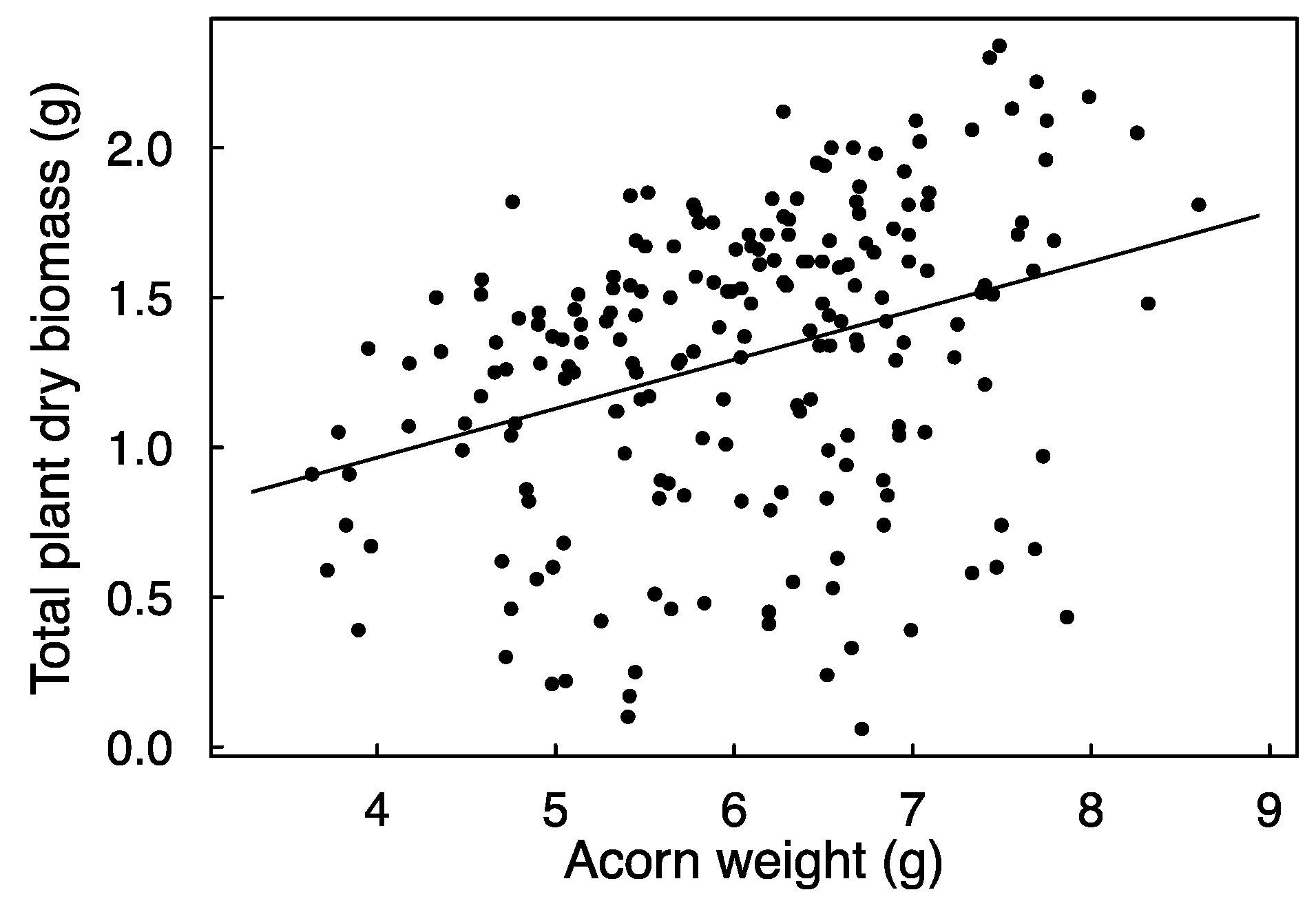
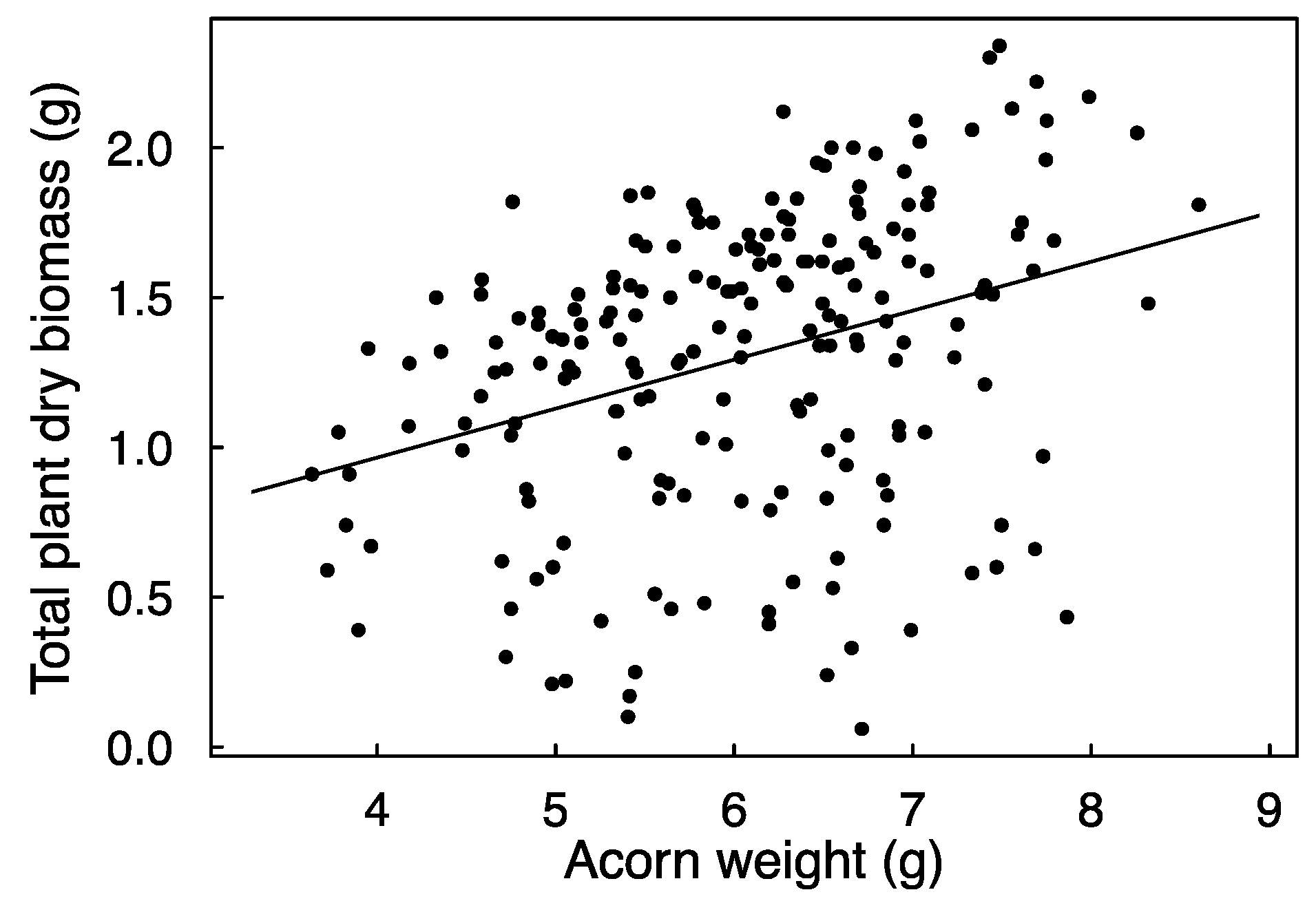
The application of diesel significantly reduced the probability of emergence from 83.3% of untreated acorns to 59.4% of treated acorns. This effect appeared regardless of submersion time, and there was no effect of acorn weight (Table 1). The time between sowing and emergence averaged 113.4 ± 1.89 days (mean ± SE), and it was neither significantly affected by diesel nor by acorn weight or the diesel × weight interaction (Table 1). Similarly, none of the tested factors affected the height of the stems of emerged plants at the end of the growing season (Table 1). Seedling biomass was positively related to acorn weight (Figure 2) but not significantly affected by diesel (Table 1).
The reduction in emergence due to submersion in diesel is in line with results that have previously been published [20] and with our expectations. However, such reduction in emergence does not necessarily eliminate the possibility of using a substance as a repellent: its effectiveness results from the trade-off between a positive effect of reducing predation and the potential negative effect on the plants. For instance, the 24% reduction in emergence found in this study could still have yielded an overall positive balance in case there had been a 50% reduction in acorn consumption, especially as the effects of acorn submersion in diesel do not seem to extend beyond the ontogenetic stages of germination and emergence. Such trade-off would, in turn, be balanced with the economic cost of acquiring and applying the repellent. In our case, however, rather than a trade-off between a positive and a negative outcome of diesel, our results suggest that the net outcome is the addition of two negative effects.
4. Conclusions
In this study, submersion in diesel failed to provide a solution to acorn predation. Regardless of the time that acorns were submerged (6, 12 or 24 h), diesel reduced the emergence of acorns and did not appear to have any positive effect in deterring mammal predators—on the contrary, it seemed to attract them. Finding effective protection for sown acorns and other large-seeded tree species continues to be a critical task that could lead to vast economic savings and important benefits for plant survival and performance related to the turning from planting nursery-grown seedlings to direct-seeding of acorns. The efforts in finding such protection should be directed to methods other than the application of diesel.
Acknowledgments
We thank Javier Ruiz Guzmán for valuable assistance in fieldwork. The Direction of the Natural Protected Area of Sierra Nevada provided fieldwork permissions and support. AL acknowledges postdoctoral fellowship from Spanish MINECO (FJCI-2015-23687). This work was supported by project IFAPA PR.AVA.AVA01.19.
Author Contributions
A.B.L. and J.C. conceived and designed the study; all authors performed the experiments; A.B.L. analyzed the data and wrote the initial draft; all authors revised the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Twedt, D.J.; Wilson, R.R. Development of oak plantations established for wildlife. For. Ecol. Manag. 2002, 162, 287–298. [Google Scholar] [CrossRef]
- Dey, D.C.; Jacobs, D.; Mcnabb, K.; Miller, G.; Baldwin, V.; Foster, G. Artificial Regeneration of Major Oak (Quercus) Species in the Eastern United States—A Review of the Literature. For. Sci. 2008, 54, 77–106. [Google Scholar]
- Benayas, J.M.R.; Navarro, J.; Espigares, T.; Nicolau, J.M.; Zavala, M.A. Effects of artificial shading and weed mowing in reforestation of Mediterranean abandoned cropland with contrasting Quercus species. For. Ecol. Manag. 2005, 212, 302–314. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Castro, J.; Delgado-Capel, M.J.; Molinas-González, C.; Pulgar, M.; Marañón-Jiménez, S.; Delgado-Huertas, A.; Querejeta, J.I. Restoring for the present or restoring for the future: Enhanced performance of two sympatric oaks (Quercus ilex and Quercus pyrenaica) above the current forest limit. Restor. Ecol. 2015, 23, 936–946. [Google Scholar] [CrossRef]
- Valdecantos, A.; Cortina, J.; Vallejo, V.R. Nutrient status and field performance of tree seedlings planted in Mediterranean degraded areas. Ann. For. Sci. 2006, 63, 249–256. [Google Scholar] [CrossRef]
- Zadworny, M.; Jagodziński, A.M.; Łakomy, P.; Ufnalski, K.; Oleksyn, J. The silent shareholder in deterioration of oak growth: Common planting practices affect the long-term response of oaks to periodic drought. For. Ecol. Manag. 2014, 318, 133–141. [Google Scholar] [CrossRef]
- Tsakaldimi, M.; Tsitsoni, T.; Ganatsas, P.; Zagas, T. A comparison of root architecture and shoot morphology between naturally regenerated and container-grown seedlings of Quercus ilex. Plant Soil 2009, 324, 103–113. [Google Scholar] [CrossRef]
- Allen, J.A.; Keeland, B.D.; Stanturf, J.A.; Clewell, A.F.; Kennedy, H.E., Jr. A Guide to Bottomland Hardwood Restoration; General Technical Report SRS-40; U.S. Department of Agriculture, U.S. Department of the Interior and Forest Service: Washington, DC, USA; U.S. Geological Survey: Reston, VA, USA, 2001; p. vii-132.
- Bullard, S.H.; Hodges, J.D.; Johnson, R.L.; Straka, T.J. Economics of direct seeding and planting for establishing oak stands on old-field sites in the South. South. J. Appl. For. 1992, 16, 35–40. [Google Scholar]
- Crawley, M.J.; Long, C.R. Alternate bearing, predator satiation and seedling recruitment in Quercus robur L. J. Ecol. 1995, 83, 683–696. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Castro, J.; Puerta-Piñero, C.; Rey Benayas, J.M. Suitability of the management of habitat complexity, acorn burial depth, and a chemical repellent for post-fire reforestation of oaks. Ecol. Eng. 2013, 53, 15–22. [Google Scholar] [CrossRef]
- Pérez-Ramos, I.M.; Marañón, T. Factors affecting post-dispersal seed predation in two coexisting oak species: Microhabitat, burial and exclusion of large herbivores. For. Ecol. Manag. 2008, 255, 3506–3514. [Google Scholar] [CrossRef]
- Puerta-Piñero, C.; Sánchez-Miranda, A.; Leverkus, A.; Castro, J. Management of burnt wood after fire affects post-dispersal acorn predation. For. Ecol. Manag. 2010, 260, 345–352. [Google Scholar] [CrossRef]
- Birkedal, M.; Löf, M.; Olsson, G.E.; Bergsten, U. Effects of granivorous rodents on direct seeding of oak and beech in relation to site preparation and sowing date. For. Ecol. Manag. 2010, 259, 2382–2389. [Google Scholar] [CrossRef]
- Farlee, L.D. Direct seeding of fine hardwood tree species. In Managing Fine Hardwoods after a Half Century of Research, Proceedings of the Seventh Walnut Council Research Symposium, Madison, WI, USA, 1–3 August 2011; General Technical Report NRS-P-115; Van Sambeek, J.W., Jackson, E.A., Coggeshall, M.V., Thomas, A.L., Michler, C.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA; Northern Research Station: Newtown Square, PA, USA, 2013; pp. 31–47. [Google Scholar]
- Madsen, P.; Löf, M. Reforestation in southern Scandinavia using direct seeding of oak (Quercus robur L.). Forestry 2005, 78, 55–64. [Google Scholar] [CrossRef]
- Smith, H.G.; Hopmans, P.; Sheridan, G.J.; Lane, P.N.J.; Noske, P.J.; Bren, L.J. Impacts of wildfire and salvage harvesting on water quality and nutrient exports from radiata pine and eucalypt forest catchments in South-Eastern Australia. For. Ecol. Manag. 2012, 263, 160–169. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Rojo, M.; Castro, J. Habitat complexity and individual acorn protectors enhance the post-fire restoration of oak forests via seed sowing. Ecol. Eng. 2015, 83, 276–280. [Google Scholar] [CrossRef]
- Reque, J.A.; Martin, E. Designing acorn protection for direct seeding of quercus species in high predation areas. For. Syst. 2015, 24, e018. [Google Scholar] [CrossRef]
- Bäumler, W.; Mielke, H.; Zahner, V. Behandlung von Eicheln mit Repellentien gegen Nagetierschäden. Curr. J. Pest Sci. 1990, 63, 15–19. [Google Scholar] [CrossRef]
- Puerta-Piñero, C. Intermediate spatial variations on acorn predation shapes Holm oak establishment within a Mediterranean landscape context. Plant Ecol. 2010, 210, 213–224. [Google Scholar] [CrossRef]
- Gómez, J.M.; Puerta-Piñero, C.; Schupp, E.W. Effectiveness of rodents as local seed dispersers of Holm oaks. Oecologia 2008, 155, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Leverkus, A.B.; Fuster, F. A new device to foster oak forest restoration via seed sowing. New For. 2015, 46, 919–929. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M.; Gómez-Aparicio, L. Benefits of using shrubs as nurse plants for reforestation in Mediterranean mountains: A 4-year study. Restor. Ecol. 2004, 12, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Use of shrubs as nurse plants: A new technique for reforestation in Mediterranean mountains. Restor. Ecol. 2002, 10, 297–305. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 10 July 2017).
- Bates, D.; Mächler, M.; Bolker, B. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1–128; 2016; Available online: http://CRAN.R-project.org/package=nlme (accessed on 10 July 2017).
- Therneau, T.M. coxme: Mixed Effects Cox Models; R Package Version 2.2–5; 2015; Available online: http://CRAN.R-project.org/package=coxme (accessed on 10 July 2017).
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000. [Google Scholar]
- Crawley, M.J. The R Book, 2nd ed.; Wiley & Sons: New Delhi, India, 2013; ISBN 9780262232586. [Google Scholar]
- Gómez, J.M.; Hódar, J.A. Wild boars (Sus scrofa) affect the recruitment rate and spatial distribution of holm oak (Quercus ilex). For. Ecol. Manag. 2008, 256, 1384–1389. [Google Scholar] [CrossRef]
- Gómez, J.M. Importance of microhabitat and acorn burial on Quercus ilex early recruitment: Non-additive effects on multiple demographic processes. Plant Ecol. 2004, 172, 287–297. [Google Scholar] [CrossRef]
- Wróbel, A.; Bogdziewicz, M. It is raining mice and voles: Which weather conditions influence the activity of Apodemus flavicollis and Myodes glareolus? Eur. J. Wildl. Res. 2015, 61, 475–478. [Google Scholar] [CrossRef]
Figure 1.
Average acorn removal in the field experiment after 9 days. Exclosures prevented the access of ungulate acorn predators. The diesel treatment included four levels of submersion time of acorns in diesel. The differences in average removal rates between exclosure and diesel treatments were not significant (see Results).
Figure 1.
Average acorn removal in the field experiment after 9 days. Exclosures prevented the access of ungulate acorn predators. The diesel treatment included four levels of submersion time of acorns in diesel. The differences in average removal rates between exclosure and diesel treatments were not significant (see Results).
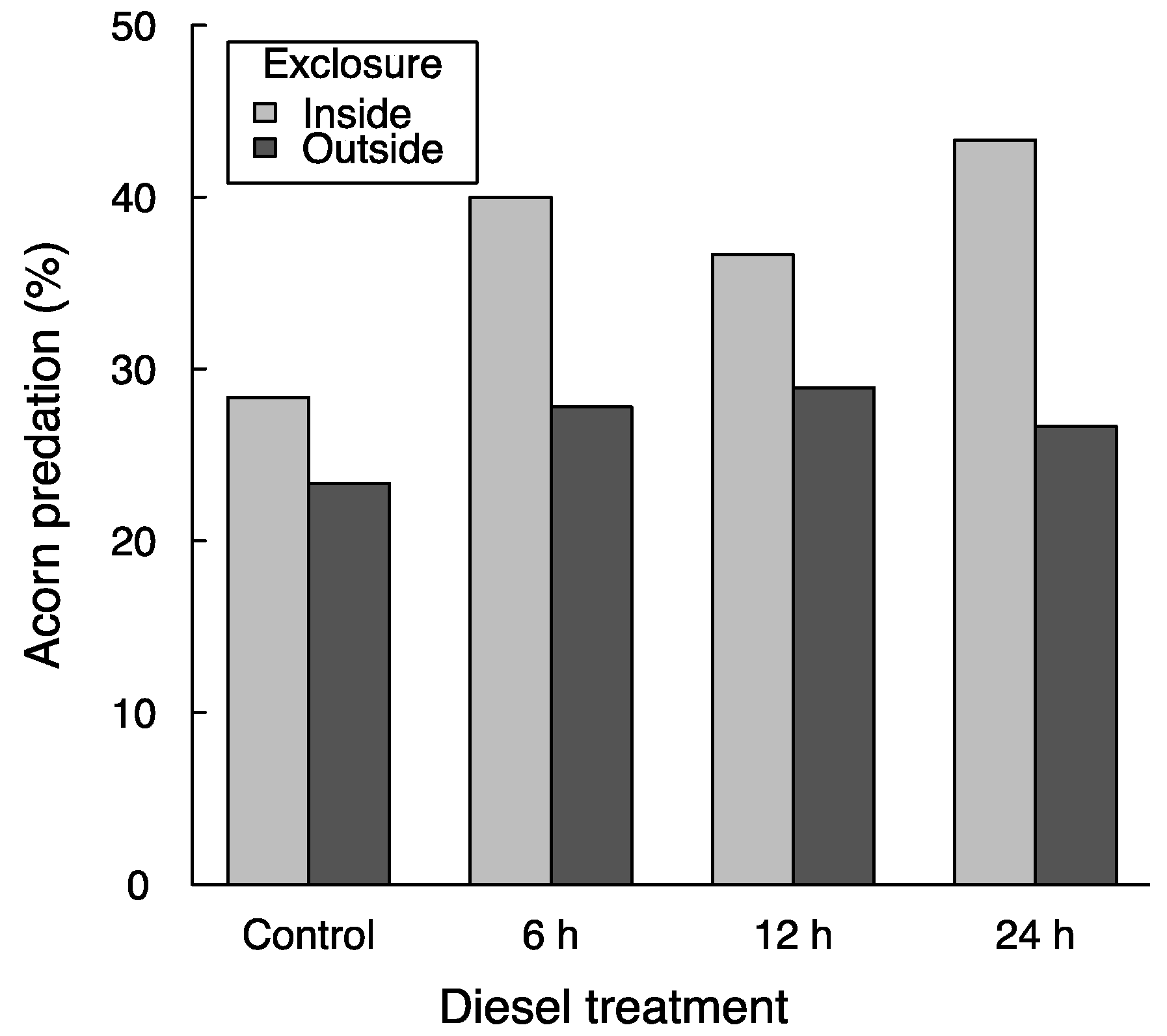
Figure 2.
Relationship between acorn weight and plant dry biomass. The line represents a simple linear regression line (the significance of this effect was assessed with mixed effects models; see Methods). R2 conditional of the model (i.e., variance explained by fixed and random effects combined) is 0.21.
Figure 2.
Relationship between acorn weight and plant dry biomass. The line represents a simple linear regression line (the significance of this effect was assessed with mixed effects models; see Methods). R2 conditional of the model (i.e., variance explained by fixed and random effects combined) is 0.21.

{kind=link}
{kind=link}
Table 1.
Results of mixed effects models for the nursery experiment.
| Term | df | Emergence Probability | Emergence Time | Stem Height | Seedling Dry Biomass | ||||
|---|---|---|---|---|---|---|---|---|---|
| χ2 | p | χ2 | p | χ2 | p | χ2 | p | ||
| Diesel (D) | 3 | 17.71 | <0.001 | 6.26 | 0.10 | 3.21 | 0.36 | 5.83 | 0.12 |
| Acorn weight (W) | 1 | 2.03 | 0.15 | 0.43 | 0.51 | 2.60 | 0.11 | 29.39 | <0.001 |
| D × W | 3 | --- 1 | --- 1 | 3.66 | 0.30 | 1.15 | 0.77 | 1.51 | 0.68 |
1 Not tested due to lack of model convergence; Numbers in bold indicate significant results.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Leverkus, A.B.; Carrión, M.; Molina-Morales, M.; Castro, J. Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns. Forests 2017, 8, 276. https://doi.org/10.3390/f8080276
AMA Style
Leverkus AB, Carrión M, Molina-Morales M, Castro J. Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns. Forests. 2017; 8(8):276. https://doi.org/10.3390/f8080276
Chicago/Turabian StyleLeverkus, Alexandro B., Manuel Carrión, Mercedes Molina-Morales, and Jorge Castro. 2017. "Effectiveness of Diesel as a Mammal Repellent for Direct Seeding of Acorns" Forests 8, no. 8: 276. https://doi.org/10.3390/f8080276
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.