Differential Effects of Coarse Woody Debris on Microbial and Soil Properties in Pinus densiflora Sieb. et Zucc. Forests
by
 , and
, and
Seongjun Kim
1, 
Guanlin Li
1,
Seung Hyun Han
1,
Hanna Chang
1,
Hyun-Jun Kim
1,2 and
Yowhan Son
1,* 1
Department of Environmental Science and Ecological Engineering, Korea University, Seoul 02841, Korea
2
BK21 Plus Eco-leader Education center, Korea University, Seoul 02841, Korea
*
Author to whom correspondence should be addressed.
Forests 2017, 8(8), 292; https://doi.org/10.3390/f8080292
Submission received: 25 July 2017
/
Revised: 8 August 2017
/
Accepted: 10 August 2017
/
Published: 11 August 2017
(This article belongs to the Special Issue Coarse Woody Debris of Forests in a Changing World)
Abstract
:Although coarse woody debris (CWD) is important for soil functioning, the mechanism which affects soil properties beneath CWD are unclear. Here, initial changes in microbial and soil properties were studied using homogenous CWD samples in eight Korean red pine (Pinus densiflora Sieb. et Zucc.) forests. For each forest, CWD samples (diameter: 11.1 ± 0.1 cm; length: 10.2 ± 0.0 cm) from similarly aged Korean red pine trees were laid on the mineral soil surface from May to June, 2016, and soils were sampled at points beneath CWD and at a distance of 1 m from the CWD after 1 year. Soils beneath the CWD had higher moisture but lower inorganic nitrogen (N) and a higher microbial biomass C (carbon)/N ratio than those sampled 1 m from the CWD. No differences in total C and N, labile C, pH, and C substrate utilization between the soils were significant. The difference in inorganic N between the soils decreased with increasing CWD decomposition, whereas that for microbial biomass fraction in total C and N increased correspondingly. Our results showed that soil microbial affinity for retaining N might become higher than that for retaining C under the presence of CWD, which possibly alters N availability and generates a spatial heterogeneity in forest soils.
1. Introduction
Coarse woody debris (CWD) in forest ecosystems plays diverse roles in nutrient recycling and influences soil properties [1]. It contains large amounts of C and nutrients [2,3], and thus decomposition of CWDs serves a process providing bioavailable carbon (C) and nutrients into the mineral soil [4]. The presence of CWD alters soil physical structure by stabilizing the soil surface and providing organic matter that aggregate soil particles [5]. These changes occur because CWD closely interact with soil microbial biomass accumulation and resource utilization [5,6]. Consequently, soils beneath CWD can show different soil developments and nutrient dynamics compared to bulk soils beneath organic layers, known as “pedogenic hot-spots” [7,8].
Given these significances of CWD, consequences of the CWD removal during management interventions are an important area of forest ecosystem research [9]. Recent studies have found negative effects of CWD removal on soil nitrogen (N) availability and C pool as well as soil microbial communities and metabolic potential [10,11,12]. However, the interaction between CWD and soils remains one of the most unknown subjects in forest ecology. High heterogeneities in size and physicochemical properties of CWD led to variations in the effects of CWD on soil properties, which has limited the designing of a controlled experiment to monitor the relationship between CWD and soils [8]. Furthermore, only few studies have clarified changes in soil development and nutrient dynamics alongside CWD decomposition process, and particularly, the initial changes in soils beneath CWD [5]. Initial CWD decomposition by microbes and arthropods governs the magnitude of the fixation of N into CWD and the release of N from CWD in the long term [13]. Hence, studying the initial interaction between CWD and soils could provide insights into the long-term N availability in forest soils.
The goal of the current study was to understand the change in soil functioning under the presence of CWD using homogenous CWD samples from Korean red pine (Pinus densiflora Sieb. et Zucc.) forests. This species accounts for approximately half of the coniferous forest cover in South Korea and is used for various purposes [14,15]. Studying the roles of pine CWD is important, given that forest management activities and outbreak of pine wilt disease in South Korea can generate large volumes of pine CWD [16,17]. Nevertheless, not many studies have focused on the interactions between pine CWD and soils of South Korea. Here, initial changes in microbial biomass, microbial substrate utilization, and soil properties were investigated because initial microbial responses can trigger further changes in N availability and C pool of soils beneath CWD [5]. We hypothesized that (1) soils beneath CWD would differ in microbial and soil properties compared to soils away from CWD and (2) the magnitude of this difference would be related to CWD decomposition rate.
2. Materials and Methods
2.1. Study Sites
A total of eight Korean red pine forests in South Korea were selected for the current study (Table 1). All forests were subjected to a temperate climate with hot, humid summer and cold, dry winter. Five of eight forests were located in the central part of the Korean peninsula, while the others were located in the southern part. The forests differed in tree density, altitude, and microclimate that might account for potential confounding effects of these factors on CWD decomposition and soils (Table 1).
Each forest consisted of three 4 m × 10 m subplots, which included 18 cylindrical pine CWD samples (total 54 CWD samples per forest). The CWD samples were laid on the mineral soil surface after removing the upper litter and humus layers between May and June, 2016. The CWD samples were placed vertically to the slope and fastened with cable ties and strings. All CWD samples used for the current study were of similarly aged (approximately 20-year-old) Korean red pine trees, which were logged in March 2016; thus, the CWD samples had consistent sizes and properties (Table A1).
2.2. Sampling and Field Measurement
Mineral soils associated with the CWD samples were collected in May to June 2017. For each forest, 10 CWD samples were randomly selected, and mineral soils at 0–10 cm depth were collected from one point beneath each of the selected CWD (SoilCWD) and the other point 1 m from the CWD (Soilcontrol). The collected soils were mixed into a composite SoilCWD and a composite Soilcontrol sample for each study forest. Each composite soil sample was divided into two subsamples: one was sealed in a plastic bag and stored at 4 °C until analyses of microbial biomass, substrate utilization, soil moisture, and soil inorganic N within one week, whereas the other sample was air-dried and sieved through a 2-mm mesh for use in soil pH, total C, total N, and labile C analyses.
Fresh mass of all CWD samples was measured prior to laying (May–June 2016) and one year after laying (May–June 2017). To quantify the dry-to-fresh mass ratio of the CWD, one CWD sample was randomly collected from each subplot, and oven-dried at 85 °C. Dry mass of the un-sampled CWD was estimated using fresh mass of each sample and the dry-to-fresh mass ratio of the CWD sample. The decomposition of the CWD was calculated based on the % change in dry mass relative to initial dry mass for one year (between May–June 2016 and May–June 2017). Air temperature and relative humidity in each forest during the study period were monitored every 30 min with a temperature and humidity sensor (Hobo Pro V2 U23-001, Onset, Bourne, MA, USA) (Table 1).
2.3. Soil Analysis
Soil moisture was measured by oven-drying fresh soil samples at 105 °C. Soil inorganic N (NH4+ and NO3−) was extracted with 2 M KCl solution. NH4+ and NO3− were analyzed according to the methods of Mulvaney [18] and Miranda et al. [19], respectively. Soil inorganic N was calculated as the sum of NH4+ and NO3− concentrations. Soil pH was measured using a 1:5 soil-to-water ratio with a refillable pH electrode (ROSS Ultra pH/ATC Triode, Thermo Scientific Orion, Waltham, MA, USA). Total C and N were determined by dry combustion at 1000 °C with an elemental analyzer (vario Macro, Elementar Analysensysteme GmbH, Langenselbold, Germany). Labile C was quantified by utilizing the field-kit protocol of Weil et al. [20], who used 0.02 M KMnO4 solution.
Microbial biomass C and N were measured by the chloroform fumigation–extraction method [21]. For 24 h, 8 g of fresh soils was fumigated using chloroform at room temperature and atmospheric pressure [22]. Non-fumigated soils were prepared at the same time. The extracted C and N were measured with a total organic C analyzer (TOC-V CPH, Shimadzu, Kyoto, Japan) after 2 h of aeration in a fume hood and 0.5 M K2SO4 extraction. Microbial biomass C and N were calculated using the difference in C or N concentration between fumigated and non-fumigated soils (extraction coefficient: 0.45 for C and 0.68 for N) [21,23]. Microbial coefficients for C and N were calculated by dividing microbial biomass C and N with total C and N, respectively [24].
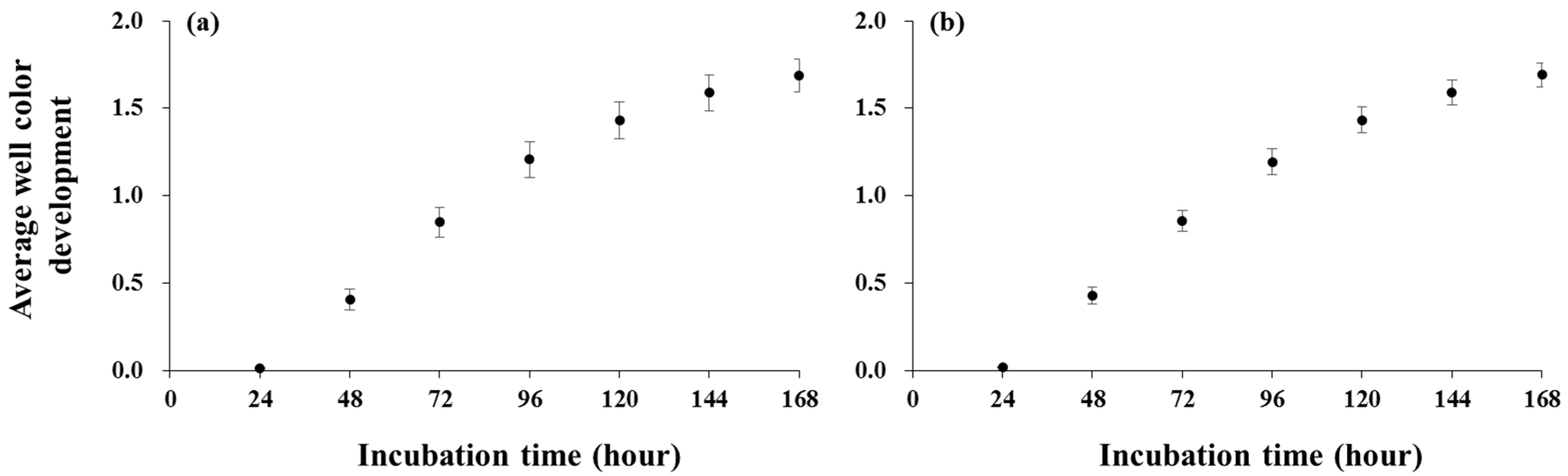
Microbial substrate utilization was measured with BIOLOGTM EcoPlate (BIOLOG Inc., Hayward, CA, USA). The EcoPlate is a 96-well transparent microplate containing 31 kinds of C substrates, which are commonly found in root exudates and soils (substrate groups are: carbohydrates, polymers, amino acids, amines, carboxylic acids, and phenols). The procedure was based on the method of Li et al. [25]. Ten grams of fresh soils were diluted using a 1:1000 soil-to-autoclaved water ratio by shaking at 200 rpm for 30 min. Of this, 0.14 mL of the soil suspension was dispensed into each well of the EcoPlate, which was incubated at 25 °C for seven days. The color development was measured every 24 h at a wavelength of 590 nm in a microplate reader (PowerWaveXS, BioTek, Winooski, VT, USA). To quantify the substrate utilization, the average well color development (AWCD) was calculated for all substrates as well as each substrate group by averaging the net absorbance measured after 96 h of incubation [26]. This time point was selected for calculating the AWCD because changes in the AWCD slowed down since this incubation time (Figure A1).
2.4. Statistical Analysis
Differences in microbial and soil properties were standardized as follows [8]:
where XCWD and Xcontrol indicate values for SoilCWD and Soilcontrol, respectively.
Δ% to Soilcontrol (%) = (XCWD − Xcontrol) Xcontrol × 100
The generalized linear mixed model approach was used to assess the significance of differences in microbial and soil properties between SoilCWD and Soilcontrol (n = 8; p < 0.05). The model included a fixed effect for the CWD presence and a G-side random effect for the study forest. If the model was statistically significant, Tukey’s HSD test was used to identify the significantly different means (p < 0.05). In addition, correlation and linear regression analyses were applied to describe pairwise relationships between microbial properties, soil properties, and CWD decomposition (n = 8; p < 0.05). Statistical analyses were done with proc GLIMMIX and CORR procedures in SAS 9.4 software (SAS Institute Inc., Cary, NC, USA).
3. Results
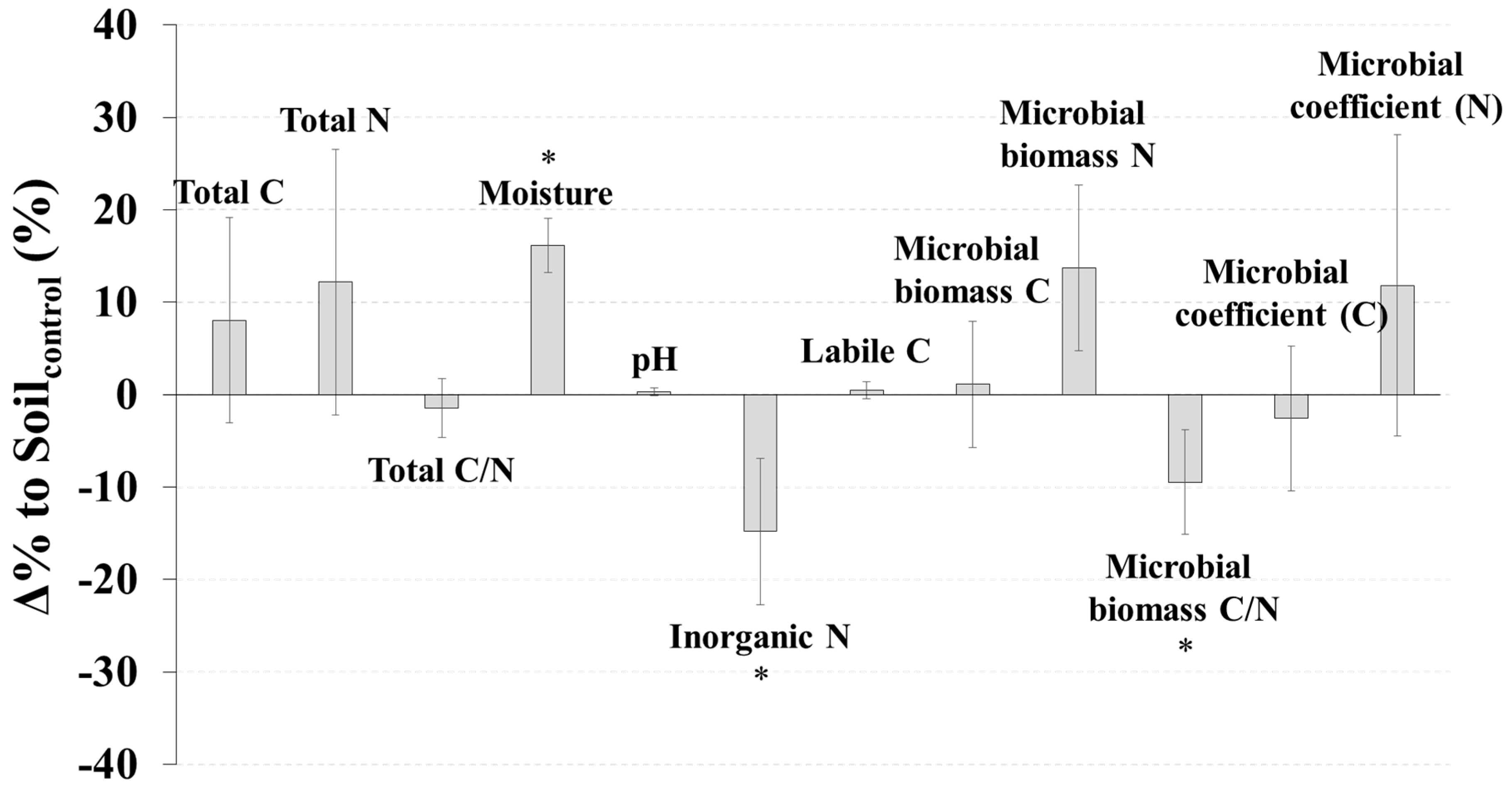
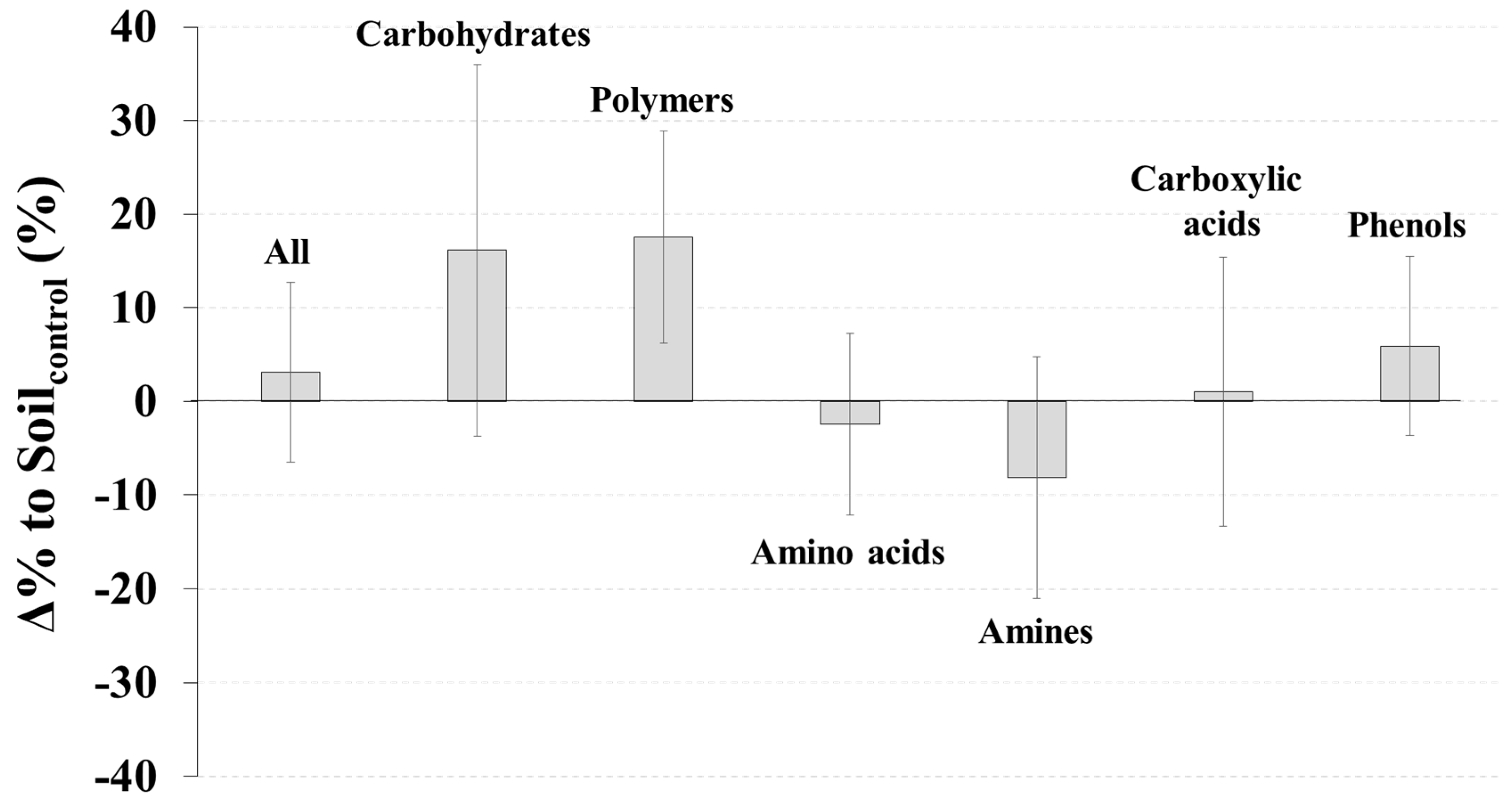
SoilCWD had significantly higher soil moisture (p < 0.01), but lower inorganic N (p < 0.05) and a lower microbial biomass C/N ratio (p < 0.05) than Soilcontrol (Figure 1). The Δ% to Soilcontrol for moisture, inorganic N, and the microbial C/N ratio were 16.1%, −14.8%, and −9.5%, respectively (Figure 1). No significant differences between SoilCWD and Soilcontrol were detected for the other microbial and soil properties (p > 0.05) (Figure 1); however, N-related properties (i.e., total N, inorganic N, microbial biomass N, and microbial coefficient for N) generally had higher |Δ% to Soilcontrol| than C-related properties (i.e., total C, labile C, microbial biomass C, and microbial coefficient for C). Differences in the AWCD between SoilCWD and Soilcontrol were not significant for all substrates and each substrate group (p > 0.05) (Figure 2).
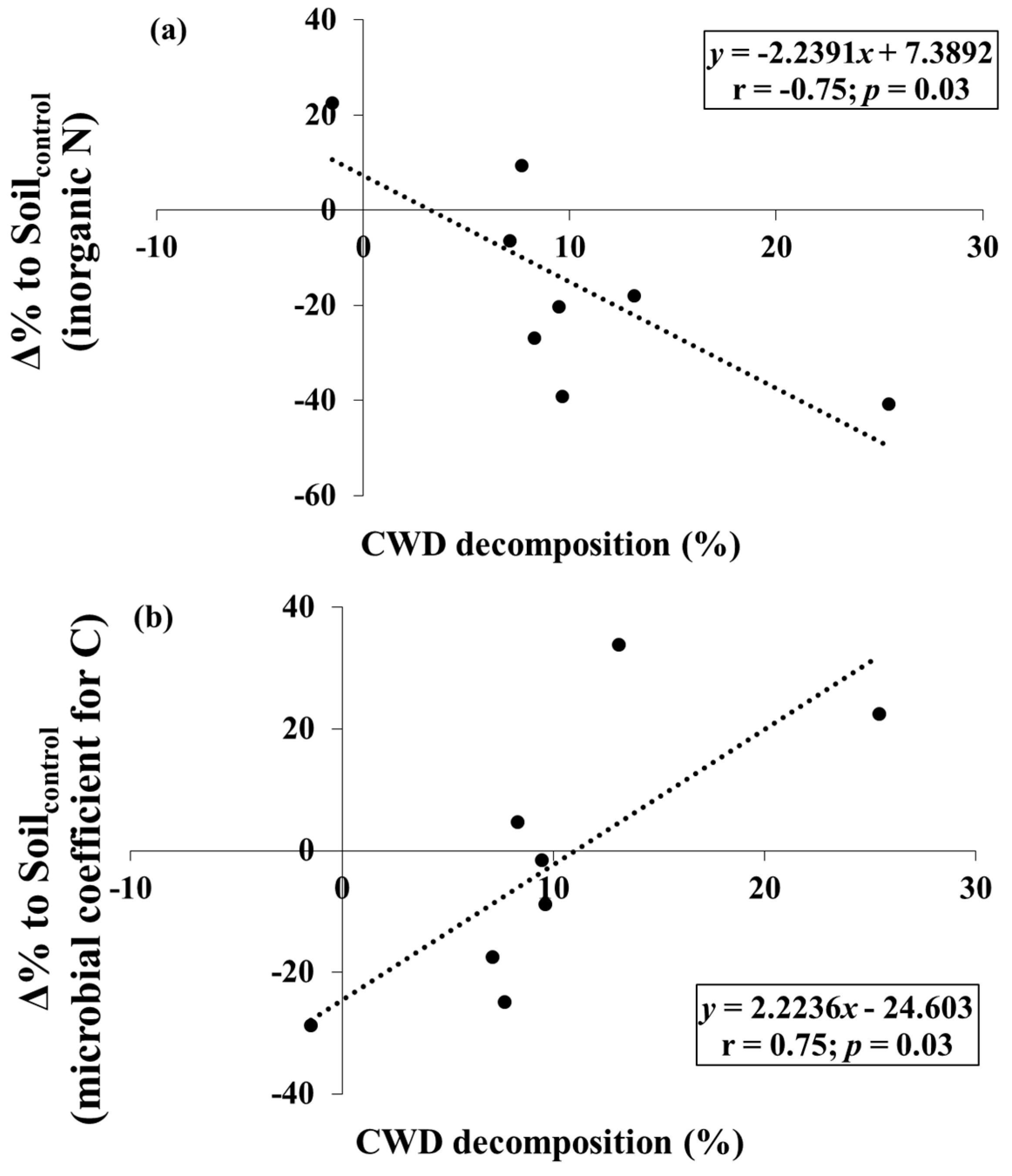
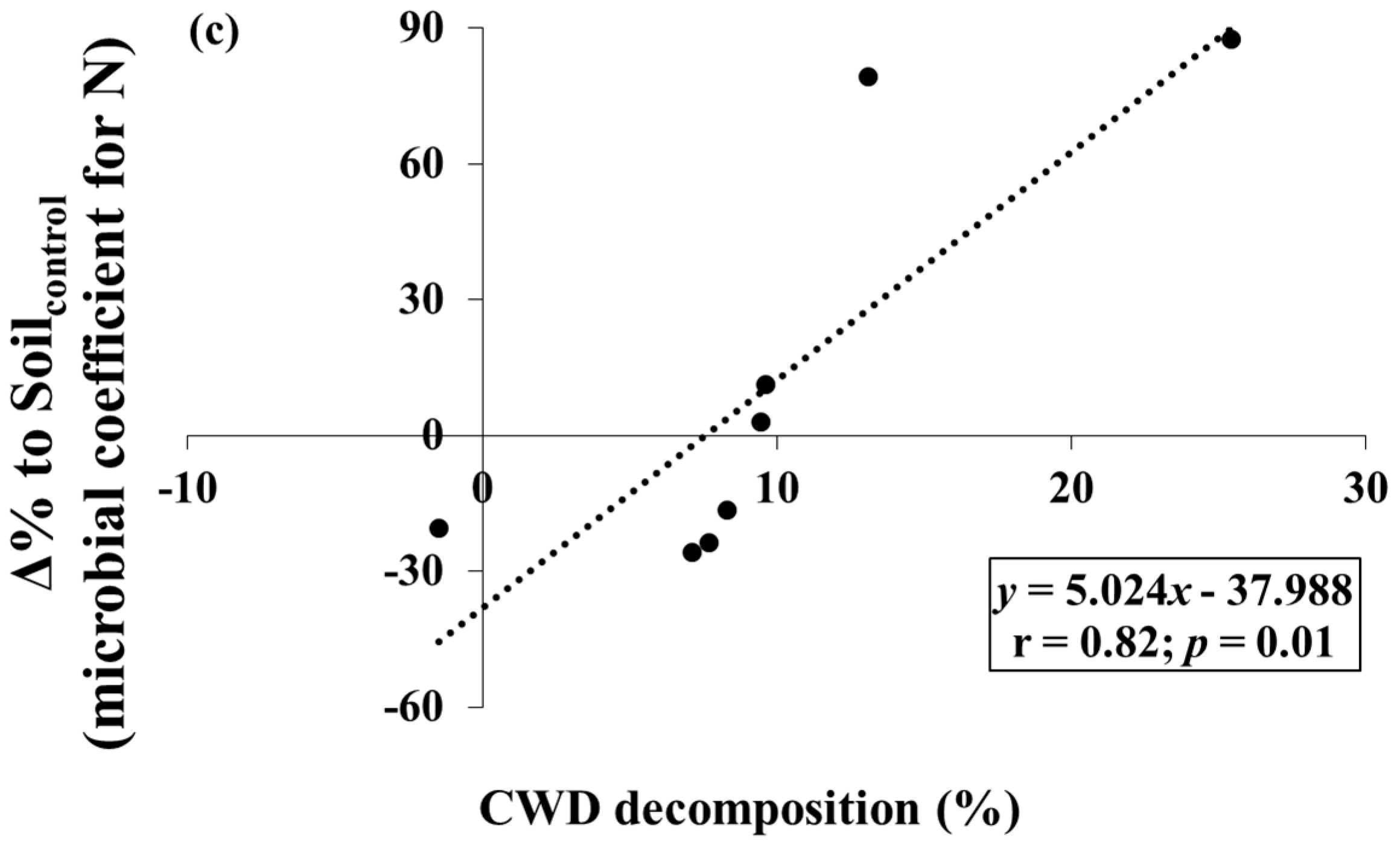
The CWD samples lost −1.5 to 25.4% of the initial mass during the study period (Table 1). Several Δ% to Soilcontrol for C- and N-related properties tended to vary with levels of CWD decomposition. The Δ% to Soilcontrol for inorganic N was negatively correlated to CWD decomposition (p < 0.05) (Figure 3a). The Δ% to Soilcontrol for microbial coefficients for C and N were positively correlated to CWD decomposition (p < 0.05), but the slope and correlation coefficient of microbial coefficient for N were higher than those of microbial coefficient for C (Figure 3b,c).
4. Discussion
Significant changes in inorganic N and the microbial biomass C/N ratio indicate that the presence of CWD could have differential effects on microbial C and N acquisitions even in the short term. Decreases in the microbial C/N ratio are generally caused by faster microbial immobilization of N than that of C in response to relative N deficiency [27]. As such, our results mean that microbial affinity for retaining N might be greater than that for retaining C under the presence of CWD. Overall, these results are in line with the general expectation that adding substrates with a high C/N ratio would enhance the N sequestration to microbial biomass, and reduce available N for plants accordingly [28].
The differential effect of CWD might result from the input of leachate containing a high C/N ratio from CWD, which possibly caused N limitation relative to C for soil microbes [6]. This relative N limitation might enhance soil microbial affinity for N uptake as well as N immobilization into pine CWD [2]. However, soil microbes beneath CWD might be unable to fully utilize additional C sources in the short term because soil N availability could act as a limiting factor for microbial growth under the N limited condition by CWD [28]. This effect of CWD could contribute to the decrease in inorganic N without changing labile C and AWCD, both of which reflect bioavailability and microbial utilization potential for soil C [26,29]. Considering that our results are inconsistent with a previous result showing that adding CWD increased AWCD after seven years [30], whether the detected patterns will persist over the long term should be confirmed through further monitoring.
It was also found that shifts in inorganic N and microbial biomass were correlated to the variation in CWD decomposition over the Korean red pine forests. The decreasing pattern in Δ% to Soilcontrol for inorganic N with increasing CWD decomposition shows that N limitation in soils beneath CWD might be more distinct under fast CWD decomposition, although CWD decomposition is known to add bioavailable C and nutrient sources into the soil [4]. Increases in microbial coefficients (fractions of microbial biomass in total soil) are closely related to the level of soil C and N limitation [27]; therefore, the steeper slope of regression of Δ% to Soilcontrol for microbial coefficients for N than that for microbial coefficients for C also exhibits the increase in N limitation relative to C under fast CWD decomposition.
Variations in Δ% to Soilcontrol because of differing CWD decomposition allow us to speculate that factors influencing CWD decomposition could also control the magnitude of differences between SoilCWD and Soilcontrol. It was found that CWD decomposition was generally faster in the southern forests than in the central forests, which might be due to higher temperature and increased arthropod activity in southern Korea [31]. Fast CWD decomposition due to these factors might accelerate substrate emission from CWD and thus promote microbial N immobilization to CWD and soils [2,3].
Given that SoilCWD and Soilcontrol were only 1 m apart, differences between SoilCWD and Soilcontrol demonstrate that the presence and decomposition of pine CWD could result in a small-scale spatial heterogeneity in forest soils. Moist and C substrate-rich conditions beneath CWD might be favorable for retaining N by soil microbes, and contribute to the small-scale spatial heterogeneity in N availability between SoilCWD and Soilcontrol accordingly [6,7]. These results further support a recent finding of Stutz and Lang [8], who emphasized CWD as “pedogenic hot-spots” in terms of changes in physicochemical properties of soils. Combining our results with their finding, we suggested that CWD could be considered to be spots where both pedogenic and microbial processes are concentrated. It could confirm the importance of appropriate CWD management in the context of sustaining soil functioning and fertility [9,10,11,12].
The detected initial change in microbial affinity also has implications for the long-term shift in soil N availability. Microbial biomass N generally acts as a potentially mineralizable N fraction, and its turnover could provide biologically available N sources for plants in the long term [32]. Microbial N retention could also contribute to maintaining soil N availability by stabilizing inorganic or dissolved forms of N, which easily leach from the soil [22]. In this context, our results indicate that the increase in microbial affinity for retaining N beneath CWD could provide additional N sources persisting over the long term, although microbial N immobilization competes with plant N uptake in the short term [29]. These findings are consistent with previous observations, showing that the CWD removal could reduce soil nutrient availability, and consequently decline the forest productivity in the long term [9].
5. Conclusions
In summary, the pine CWD altered the moisture, inorganic N, and microbial biomass C/N ratio of soils beneath them. It is notable that the effects of CWD on microbial and soil properties were related to variations in CWD decomposition across the study forests. These results demonstrate that the presence and decomposition of CWD might have differential effects on C and N acquisitions by soil microbes, and thereby lead to a spatial heterogeneity in forest soils. Changes in microbial and soil properties resulting from CWD might help conserve the long-term N availability by aiding microbes to stabilize N in bulk soils. Because the current study dealt with the initial stage of CWD decomposition only, long-term studies should be followed to thoroughly elucidate the mechanism that affects microbial and soil properties beneath CWD. However, findings from the current study will contribute to the understanding of the interaction between CWD and soils, for which some answers are still unknown [8].
Acknowledgments
This study was supported by the National Research Foundation of Korea (2015R1D1A1A01057124). We thank Heejae Jo, Hyungsub Kim, and Jusub Kim of the Korea University for their assistance during field measurements. We also thank Prof. Choonsig Kim of the Gyeongnam National University of Science and Technology for his cooperation during establishment of the southern forest sites.
Author Contributions
S.K. conceived the main idea of the study, analyzed data, and wrote the manuscript. G.L.L. participated in microbial analyses. S.H.H., H.C. and H.-J.K. participated in field measurements. Y.S. supervised the entire process of the study. All co-authors contributed to the writing of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funder played no role in the process of the current study.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Initial size and properties of coarse woody debris used for the current study.
| Diameter (cm) | Length (cm) | Volume (cm−3) | Dry Mass (g) | Density (g cm−3) | C Concentration (%) | N Concentration (%) |
|---|---|---|---|---|---|---|
| 11.1 (0.1) | 10.2 (0.0) | 995.7 (16.0) | 776.4 (6.7) | 0.79 (0.0) | 48.9 (0.3) | 0.04 (0.0) |
Values in parenthesis indicate standard errors.
Figure A1.
Average well color development for soils 1 m from coarse woody debris (a) and soils beneath coarse woody debris (b) throughout the incubation time. Error bars indicate standard errors.
Figure A1.
Average well color development for soils 1 m from coarse woody debris (a) and soils beneath coarse woody debris (b) throughout the incubation time. Error bars indicate standard errors.
References
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Noh, N.J.; Yoon, T.K.; Kim, R.-H.; Bolton, N.W.; Kim, C.; Son, Y. Carbon and nitrogen accumulation and decomposition from coarse woody debris in a naturally regenerated Korean red pine (Pinus densiflora S.et Z.) forest. Forests 2017, 8, 214. [Google Scholar] [CrossRef]
- Yoon, T.K.; Noh, N.J.; Kim, S.; Han, S.; Son, Y. Coarse woody debris respiration of Japanese red pine forests in Korea: Controlling factors and contribution to the ecosystem carbon cycle. Ecol. Res. 2015, 30, 723–734. [Google Scholar] [CrossRef]
- Laiho, R.; Prescott, C.E. The contribution of coarse woody debris to carbon, nitrogen, and phosphorus cycles in three Rocky Mountain coniferous forests. Can. J. For. Res. 1999, 29, 1592–1603. [Google Scholar] [CrossRef]
- Marañón-Jiménez, S.; Castro, J. Effect of decomposing post-fire coarse woody debris on soil fertility and nutrient availability in a Mediterranean ecosystem. Biogeochemistry 2013, 112, 519–535. [Google Scholar] [CrossRef]
- Hafner, S.D.; Groffman, P.M. Soil nitrogen cycling under litter and coarse woody debris in a mixed forest in NewYork State. Soil Biol. Biochem. 2005, 37, 2159–2162. [Google Scholar] [CrossRef]
- Brais, S.; Drouin, P. Interactions between deadwood and soil characteristics in a natural boreal trembling aspen–Jack pine stand. Can. J. For. Res. 2012, 42, 1456–1466. [Google Scholar] [CrossRef]
- Stutz, K.; Lang, F. Potentials and unknowns in managing coarse woody debris for soil functioning. Forests 2017, 8, 37. [Google Scholar] [CrossRef]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth—A meta analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar] [CrossRef]
- Adamczyk, S.; Kitunen, V.; Lindroos, A.-J.; Adamczyk, B.; Smolander, A. Soil carbon and nitrogen cycling processes and composition of terpenes five years after clear cutting a Norway spruce stand: Effects of logging residues. For. Ecol. Manag. 2016, 381, 318–326. [Google Scholar] [CrossRef]
- Cardenas, E.; Kranabetter, J.M.; Hope, G.; Maas, K.R.; Hallam, S.; Mohn, W.W. Forest harvesting reduces the soil metagenomics potential for biomass decomposition. ISME J. 2015, 9, 2465–2476. [Google Scholar] [CrossRef] [PubMed]
- Persson, T.; Lenoir, F.; Vegerfors, B. Long-term effects of stump harvesting and site preparation on pools and fluxes of soil carbon and nitrogen in central Sweden. Scand. J. For. Res. 2017, 32, 222–229. [Google Scholar] [CrossRef]
- Ulyshen, M.D. Insect-mediated nitrogen dynamics in decomposing wood. Ecol. Entomol. 2015, 40, 97–112. [Google Scholar] [CrossRef]
- Kim, D.-H.; Kim, J.-H.; Park, J.-H.; Ewane, E.B.; Lee, D.-H. Correlation between above-ground and below-ground biomass of 13-year-old Pinus densiflora S.et Z. planted in a post-fire area in Samcheok. For. Sci. Technol. 2016, 12, 115–124. [Google Scholar]
- Pyo, J. Developing the site index equation using a generalized algebraic difference approach for Pinus densiflora in central region, Korea. For. Sci. Technol. 2017, 13, 87–91. [Google Scholar] [CrossRef]
- Ko, S.; Yoon, T.K.; Kim, S.; Kim, C.; Lee, S.-T.; Seo, K.W.; Son, Y. Thinning intensity effects on carbon storage of soil, forest floor and coarse woody debris in Pinus densiflora stands. J. Korean For. Soc. 2014, 103, 30–36. (In Korean) [Google Scholar] [CrossRef]
- Shin, S.-C. Pine wilt disease in Korea. In Pine wilt disease; Zhao, K., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 26–32. [Google Scholar]
- Mulvaney, R.L. Nitrogen-inorganic forms. In Methods of Soil Analysis. Part 3—Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 1146–1155. [Google Scholar]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Weil, R.R.; Islam, K.R.; Stine, M.; Gruver, J.B.; Samson-Liebig, S.E. Estimating active carbon for soil quality assessment: A simplified method for laboratory and field use. Am. J. Altern. Agric. 2003, 18, 3–17. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Perakis, S.S.; Hedin, L.O. Fluxes and fates of nitrogen in soil of an unpolluted old-growth temperate forest, southern Chile. Ecology 2001, 82, 2245–2260. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the KEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Sparling, G.P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Aust. J. Soil Res. 1992, 30, 195–207. [Google Scholar] [CrossRef]
- Li, G.; Kim, S.; Park, M.J.; Son, Y. Short-term effects of experimental warming and precipitation manipulation on soil microbial biomass C and N, community substrate utilization patterns and community composition. Pedoshpere, 2017, 27, 714–724. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [PubMed]
- Xu, X.; Thornton, P.E.; Post, W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- Hodge, A.; Robinson, D.; Fitter, A. Are microorganisms more effective than plants at competing for nitrogen? Trends Plant Sci. 2000, 5, 304–308. [Google Scholar] [CrossRef]
- Culman, S.W.; Snapp, S.S.; Freeman, M.A.; Schipanski, M.E.; Beniston, J.; Lal, R.; Drinkwater, L.E.; Franzluebbers, A.J.; Glover, J.D.; Lee, J.; et al. Permanganate oxidizable carbon reflects a processed soil fraction that is sensitive to management. Soil Sci. Soc. Am. J. 2012, 76, 494–504. [Google Scholar] [CrossRef]
- Kwak, J.-H.; Chang, S.X.; Naeth, M.A.; Schaaf, W. Coarse woody debris increases microbial community functional diversity but not enzyme activities in reclaimed oil sands soils. PLoS ONE 2015, 10, e0143857. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Jeong, S.-Y. Ecological characteristics of termite (Reticulitermes speratus kyushuensis) for preservation of wooden cultural heritage. Conserv. Stud. 2004, 37, 327–348. (In Korean) [Google Scholar]
- Bonde, T.A.; Schnurer, J.; Rosswall, T. Microbial biomass as a fraction of potentially mineralizable nitrogen in soils from long-term field experiments. Soil Biol. Biochem. 1988, 20, 447–452. [Google Scholar] [CrossRef]
Figure 1.
Δ% to Soilcontrol (Equation (1)) for microbial and soil properties. Error bars indicate standard errors, and asterisks indicate significant differences (p < 0.05).
Figure 1.
Δ% to Soilcontrol (Equation (1)) for microbial and soil properties. Error bars indicate standard errors, and asterisks indicate significant differences (p < 0.05).
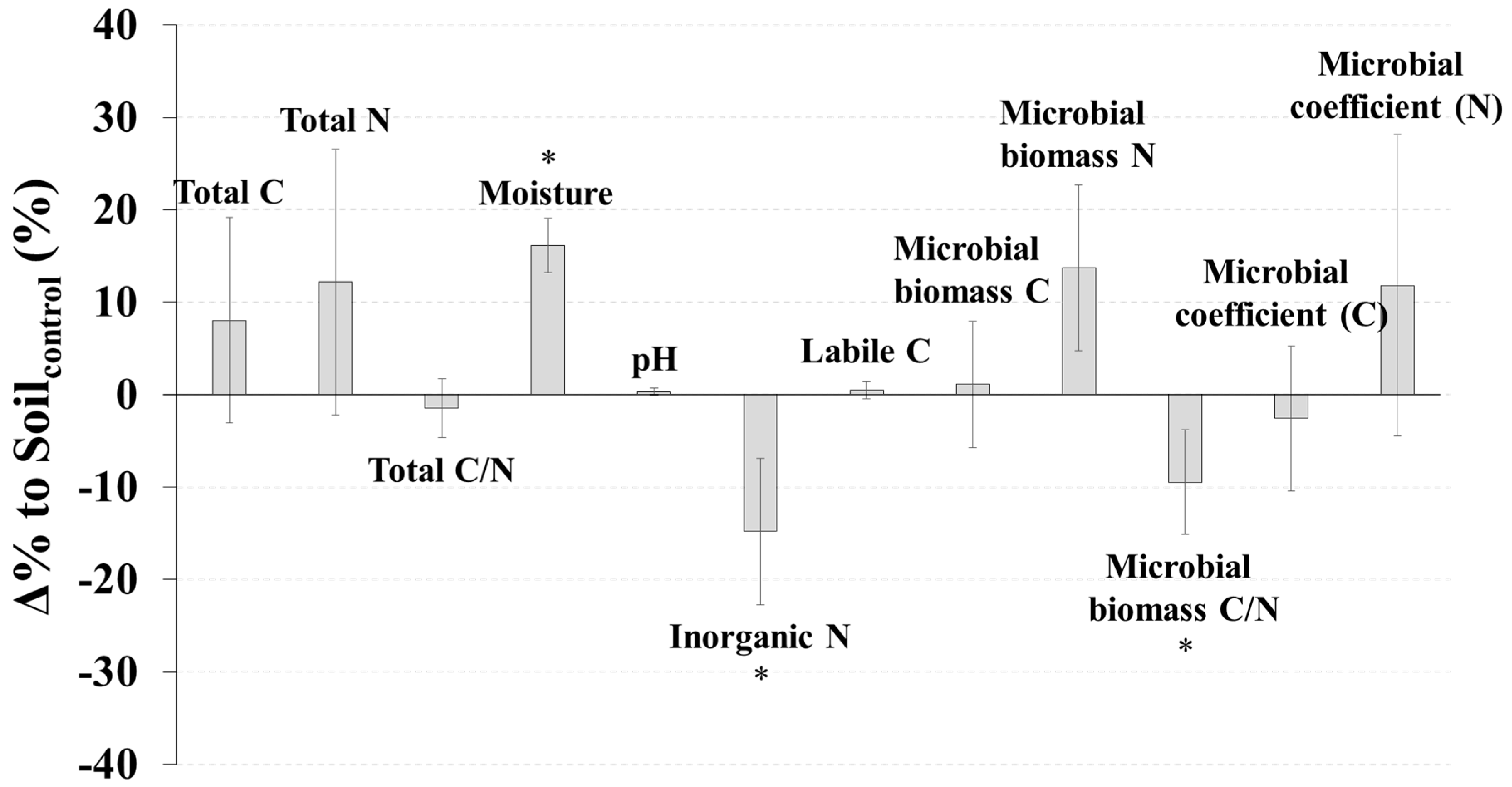
Figure 2.
Δ% to Soilcontrol (Equation (1)) for results of BIOLOG™ EcoPlate assay (average well color development). Error bars indicate standard errors.
Figure 2.
Δ% to Soilcontrol (Equation (1)) for results of BIOLOG™ EcoPlate assay (average well color development). Error bars indicate standard errors.
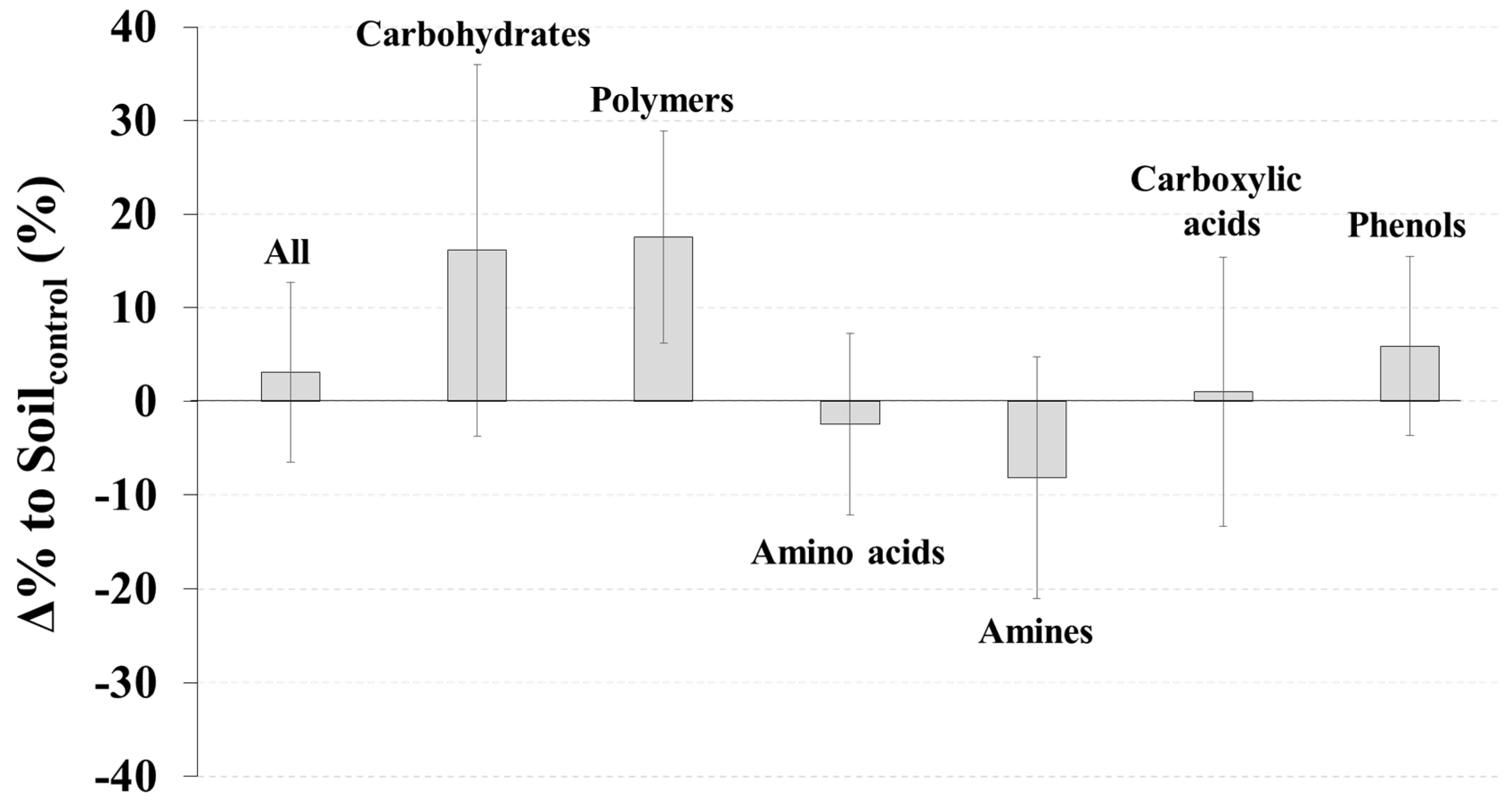
Figure 3.
Relationships between (a) Δ% to Soilcontrol (Equation (1)) for inorganic N and coarse woody debris (CWD) decomposition; (b) Δ% to Soilcontrol for microbial coefficients for C and CWD decomposition; and (c) Δ% to Soilcontrol for microbial coefficients for N and CWD decomposition.
Figure 3.
Relationships between (a) Δ% to Soilcontrol (Equation (1)) for inorganic N and coarse woody debris (CWD) decomposition; (b) Δ% to Soilcontrol for microbial coefficients for C and CWD decomposition; and (c) Δ% to Soilcontrol for microbial coefficients for N and CWD decomposition.

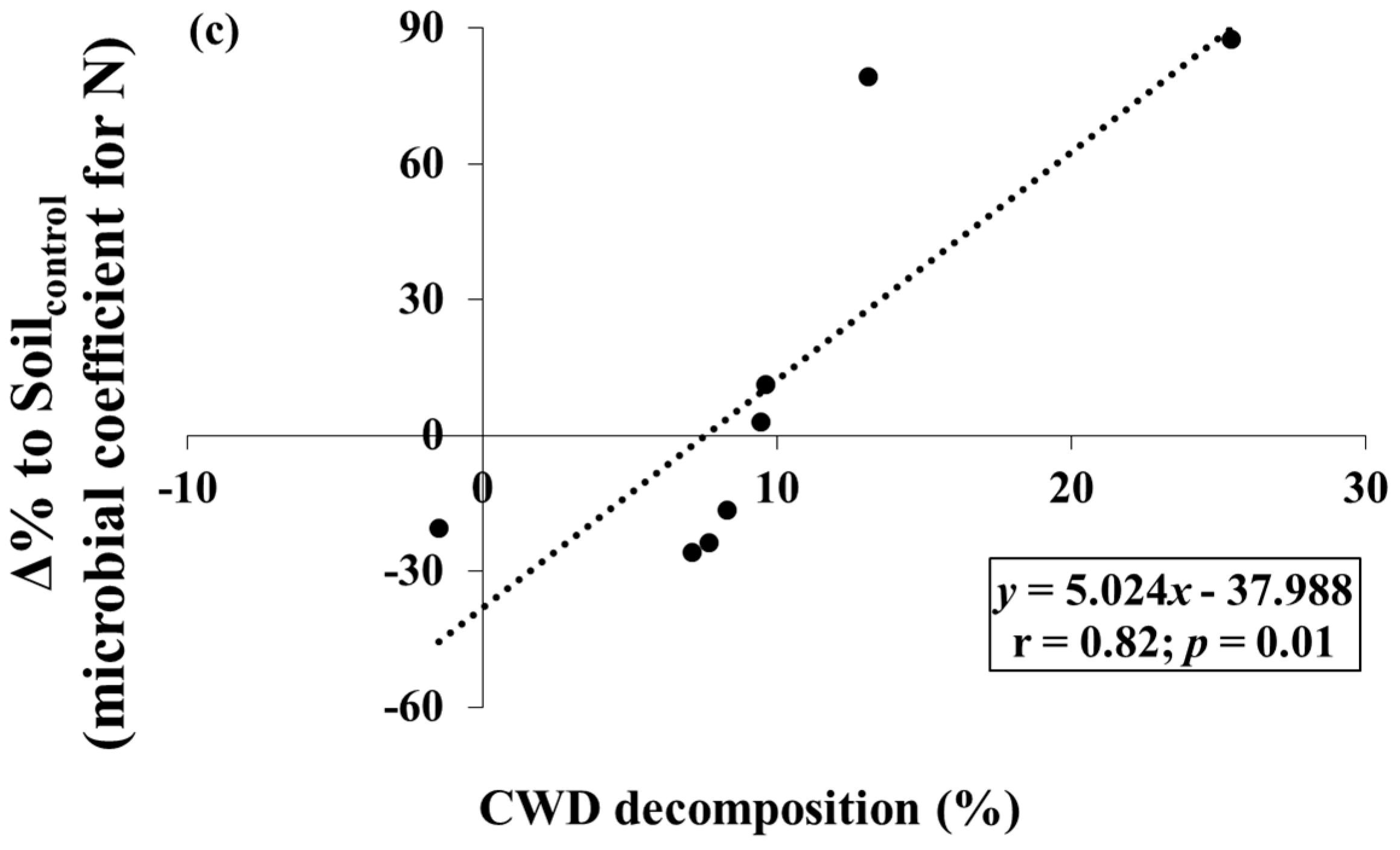
Table 1.
Conditions and coarse woody debris (CWD) decomposition in the study forests.
| Location | Tree Density (stem ha−1) | Altitude (m) | Air Temperature (°C) | Relative Humidity (%) | CWD Decomposition (%) | |
|---|---|---|---|---|---|---|
| Central 1 | 37°44′N, 127°10′E | 2500 | 500 | 11.7 | 75.5 | −1.5 |
| Central 2 | 37°46′N, 127°10′E | 1140 | 470 | 10.8 | 73.1 | 7.1 |
| Central 3 | 37°47′N, 127°10′E | 650 | 420 | 11.1 | 73.1 | 9.5 |
| Central 4 | 37°30′N, 128°56′E | 610 | 690 | 9.4 | 77.9 | 8.3 |
| Central 5 | 38°02′N, 128°22′E | 1050 | 610 | 8.8 | 80.8 | 7.7 |
| Southern 1 | 35°12′N, 128°10′E | 1560 | 180 | 14.5 | 71.8 | 9.6 |
| Southern 2 | 35°12′N, 128°10′E | 600 | 170 | 14.4 | 73.1 | 13.1 |
| Southern 3 | 35°21′N, 128°10′E | 1200 | 430 | 13.1 | 74.4 | 25.4 |
Air temperature and relative humidity represent mean annual values during the study period. CWD decomposition indicates the % change in CWD mass relative to initial mass over the study period.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, S.; Li, G.; Han, S.H.; Chang, H.; Kim, H.-J.; Son, Y. Differential Effects of Coarse Woody Debris on Microbial and Soil Properties in Pinus densiflora Sieb. et Zucc. Forests. Forests 2017, 8, 292. https://doi.org/10.3390/f8080292
AMA Style
Kim S, Li G, Han SH, Chang H, Kim H-J, Son Y. Differential Effects of Coarse Woody Debris on Microbial and Soil Properties in Pinus densiflora Sieb. et Zucc. Forests. Forests. 2017; 8(8):292. https://doi.org/10.3390/f8080292
Chicago/Turabian StyleKim, Seongjun, Guanlin Li, Seung Hyun Han, Hanna Chang, Hyun-Jun Kim, and Yowhan Son. 2017. "Differential Effects of Coarse Woody Debris on Microbial and Soil Properties in Pinus densiflora Sieb. et Zucc. Forests" Forests 8, no. 8: 292. https://doi.org/10.3390/f8080292
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.